Toxin Degradation by Rumen Microorganisms: A Review

 , , and
, , and 
Abstract
: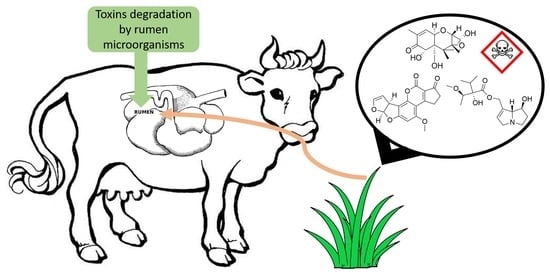
1. Introduction
2. Rumen Microbial Detoxification of Plant Toxins
2.1. Non-Protein Amino Acids
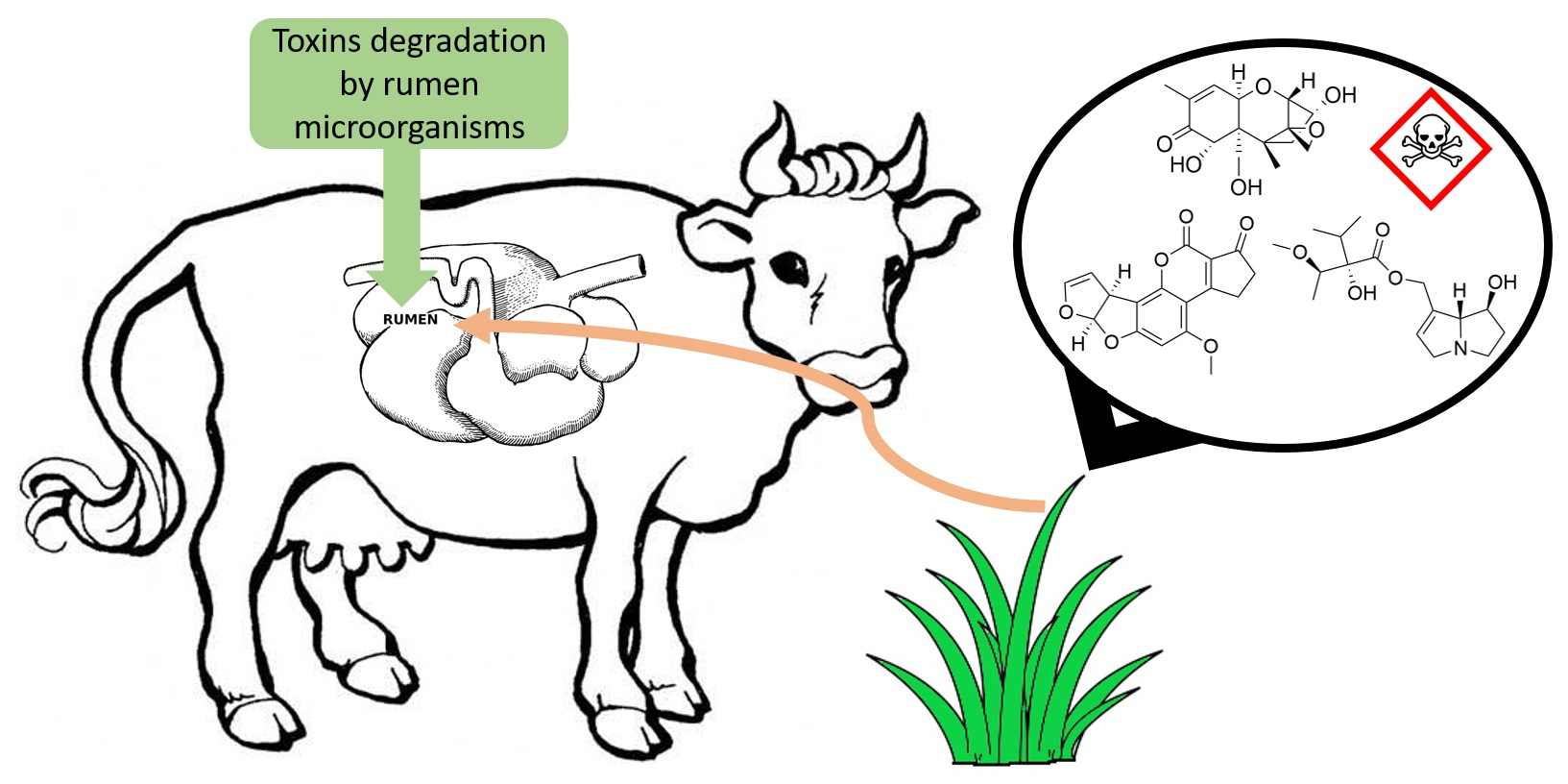
2.1.1. Mimosine
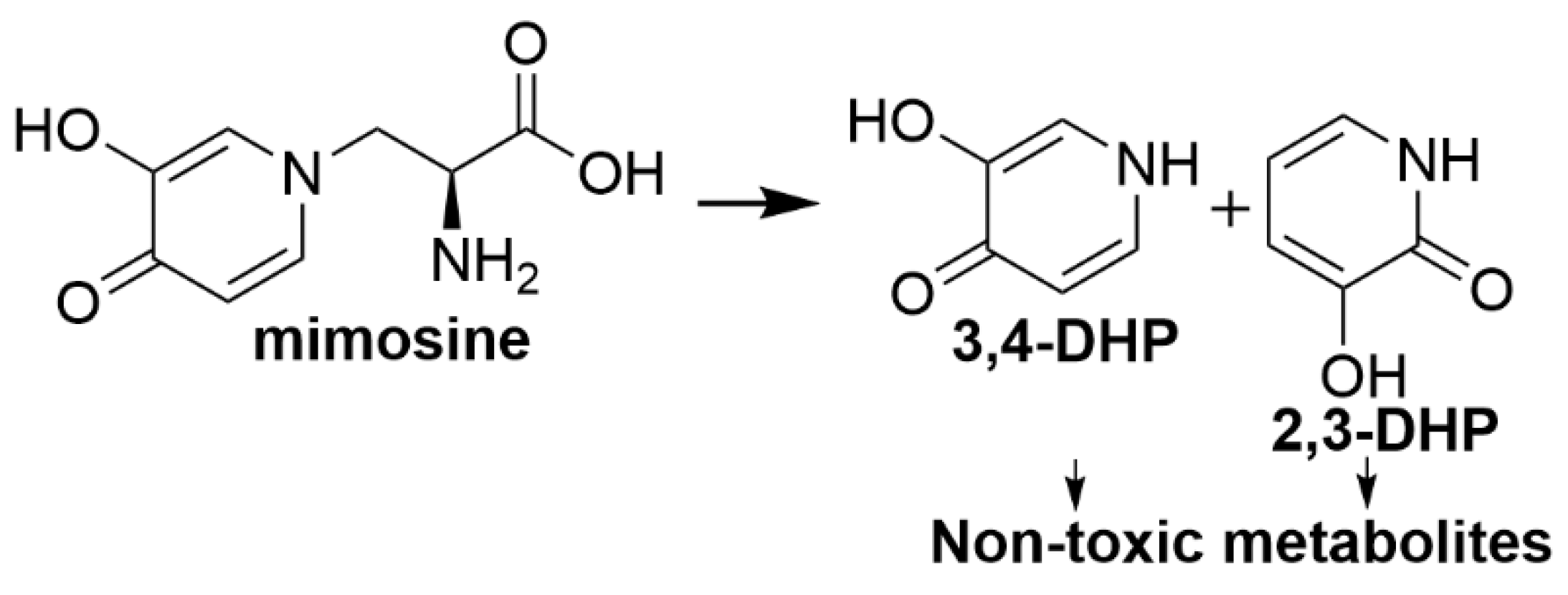
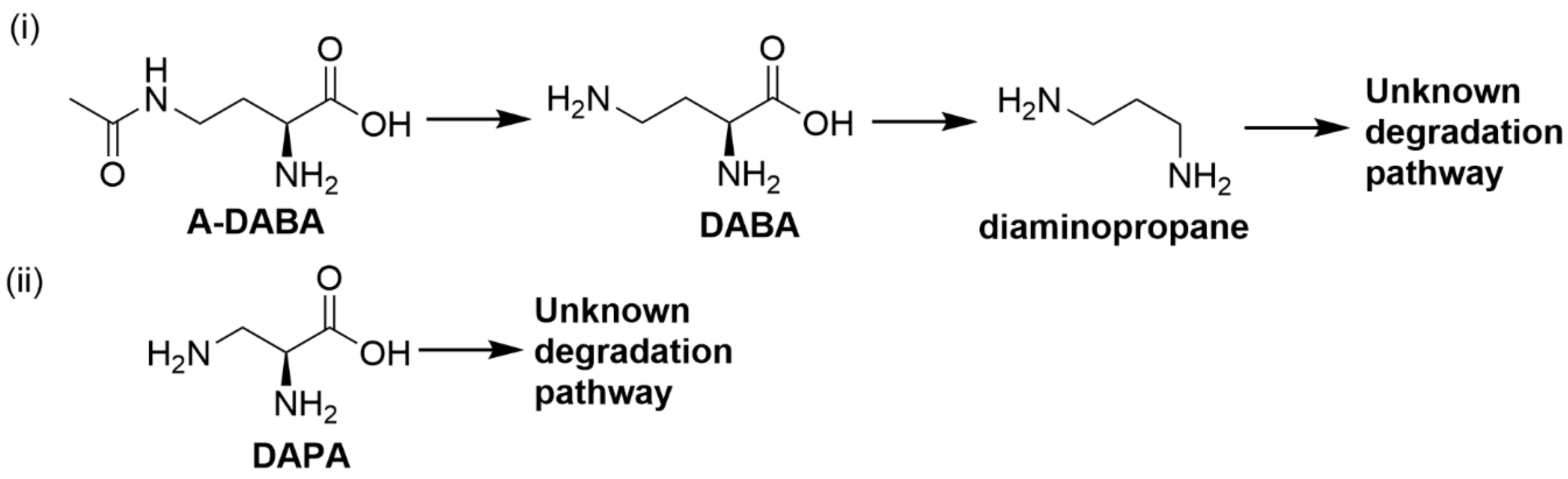
2.1.2. 4-N-Acetyl-2,4-diaminobuytric Acid (A-DABA)
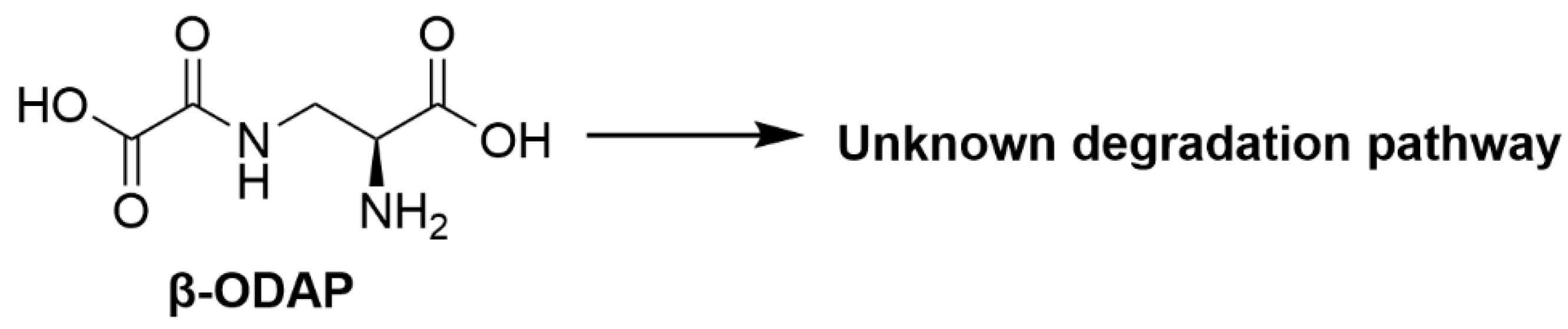
2.1.3. β-N-Oxalyl-l-α,β-diaminopropionic Acid (β-ODAP)
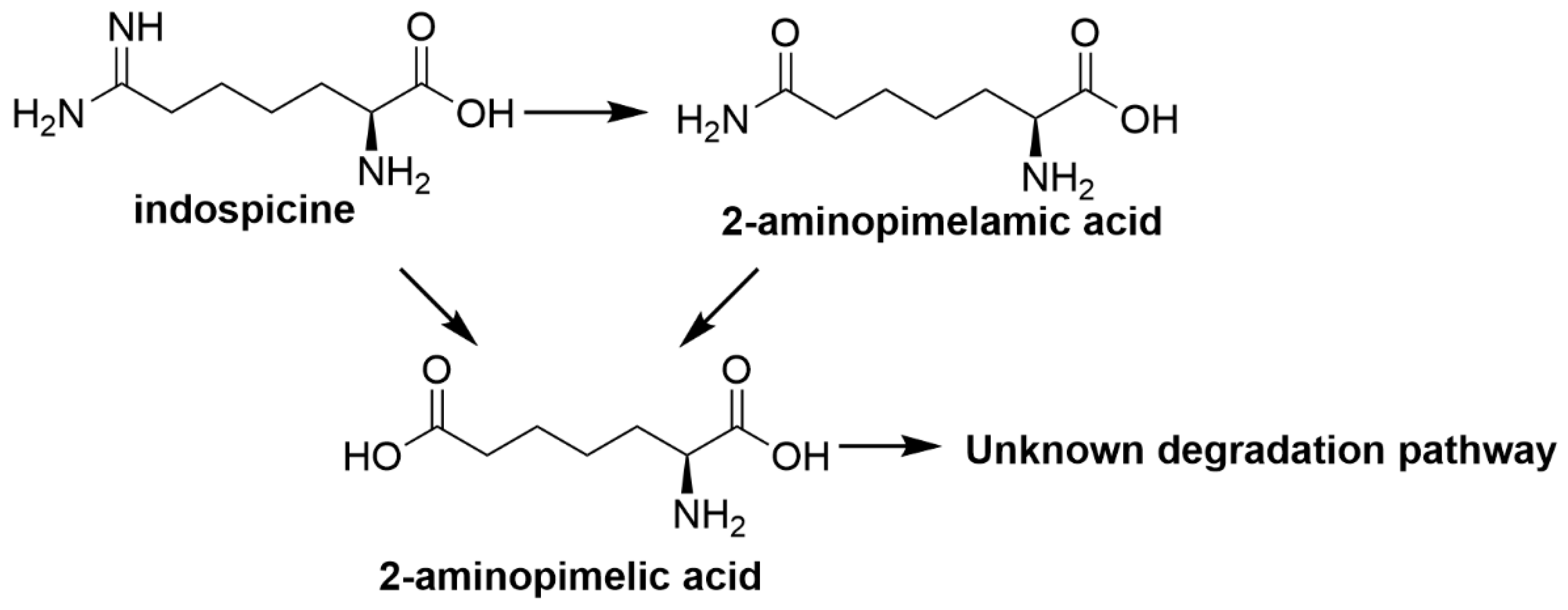
2.1.4. Indospicine
2.2. Fluoroacetate
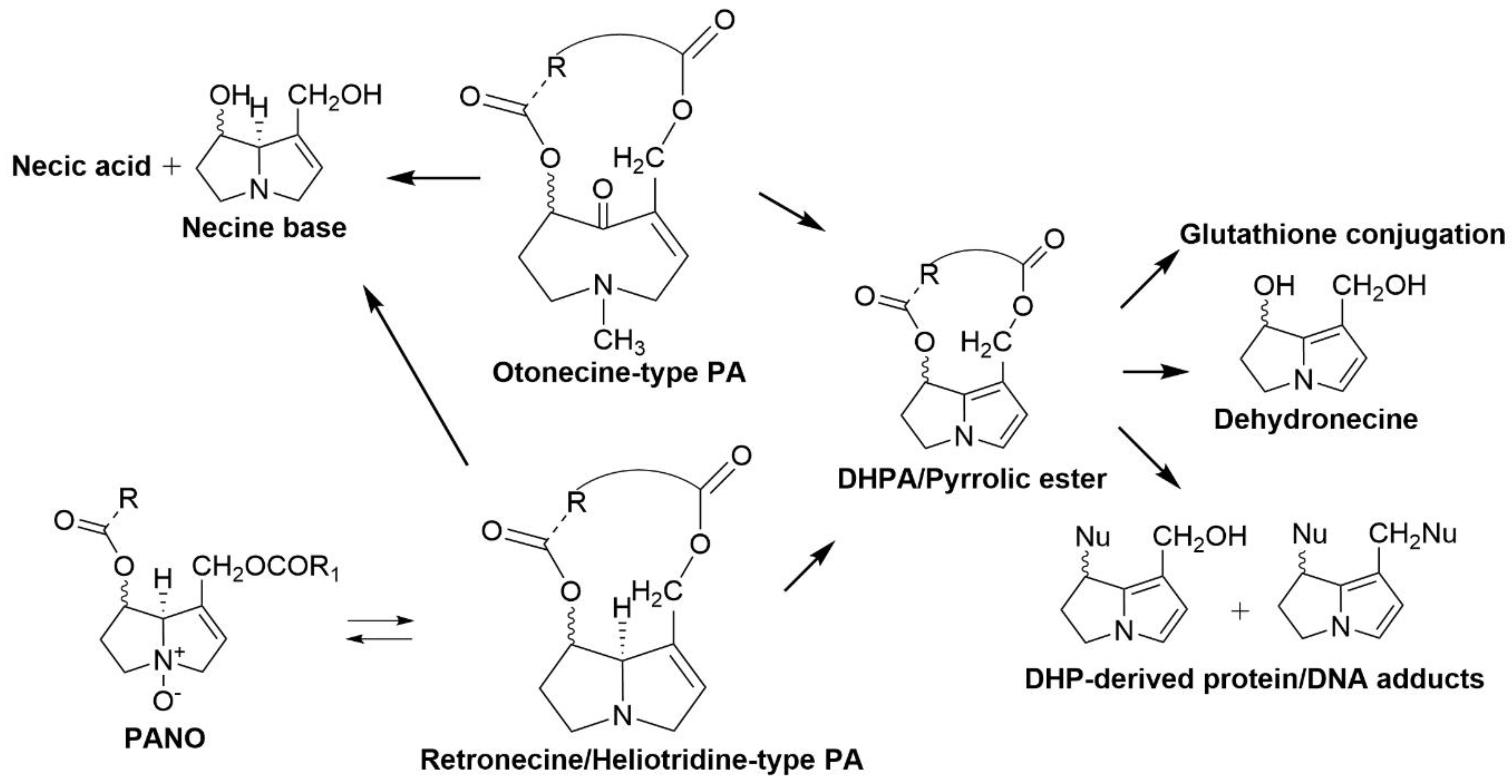
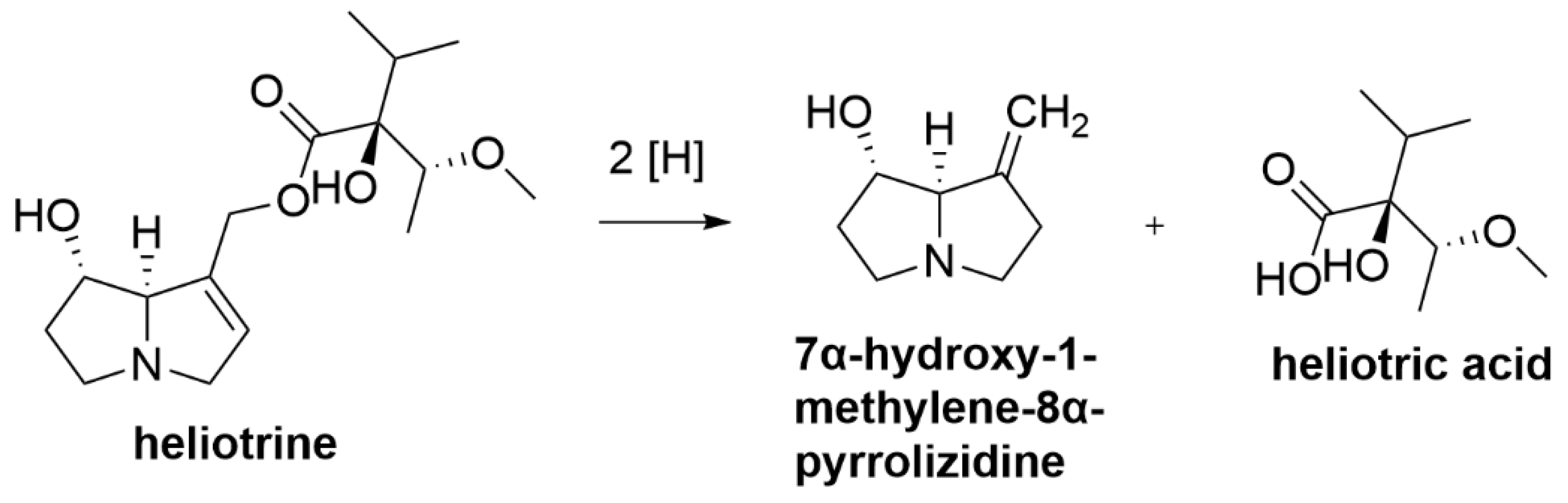
2.3. Pyrrolizidine Alkaloids
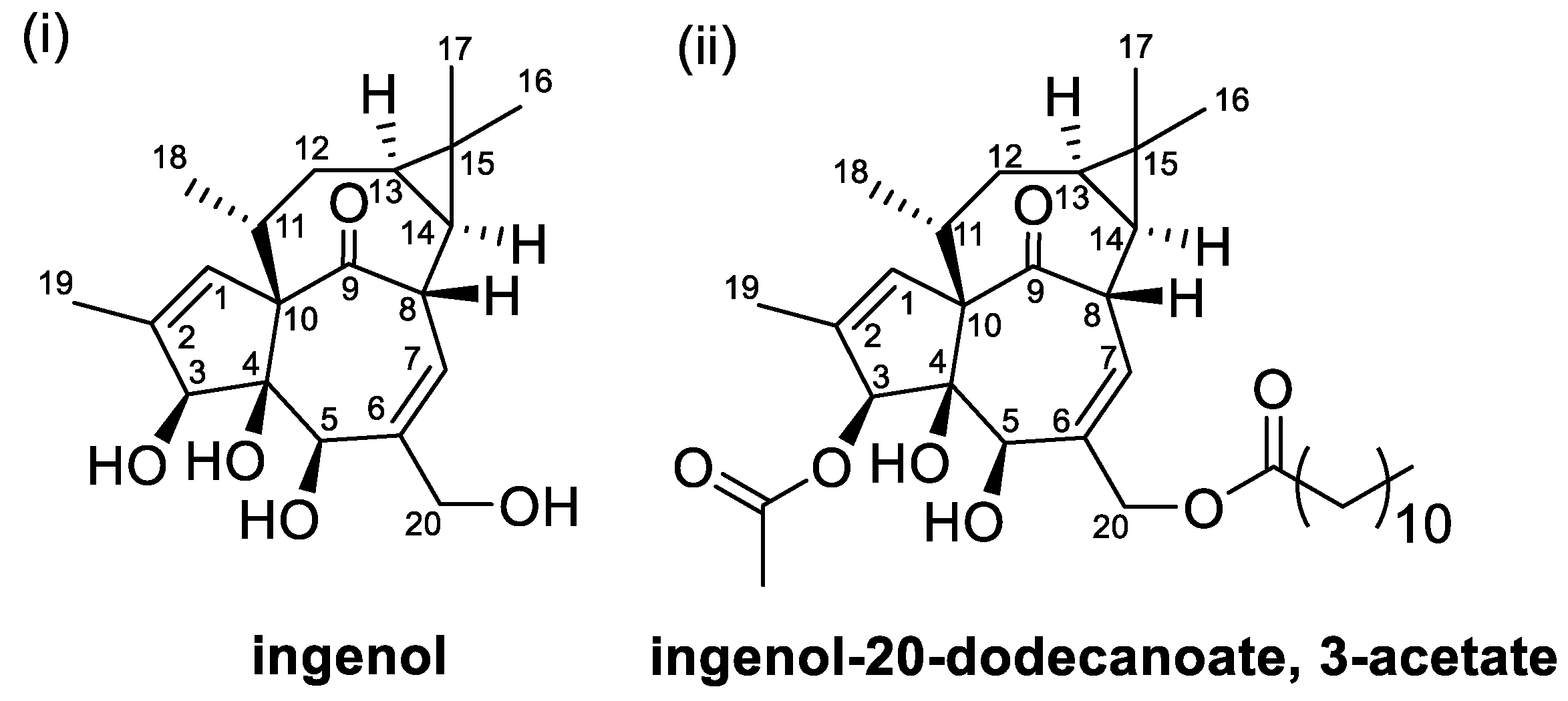
2.4. Diterpenoids of Leafy Spurge
3. Rumen Microbial Detoxification of Cyanotoxins
4. Rumen Microbial Detoxification of Mycotoxins
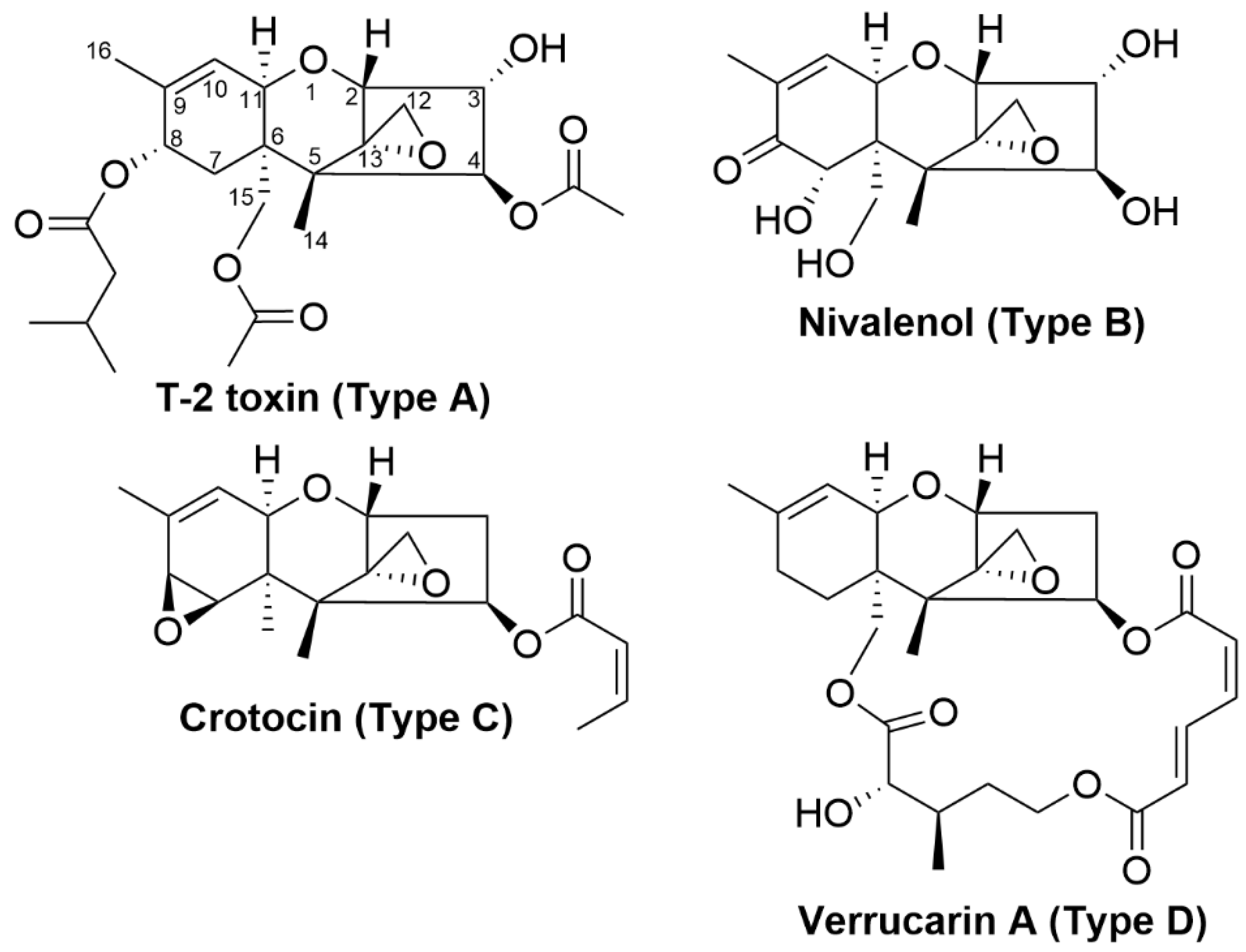
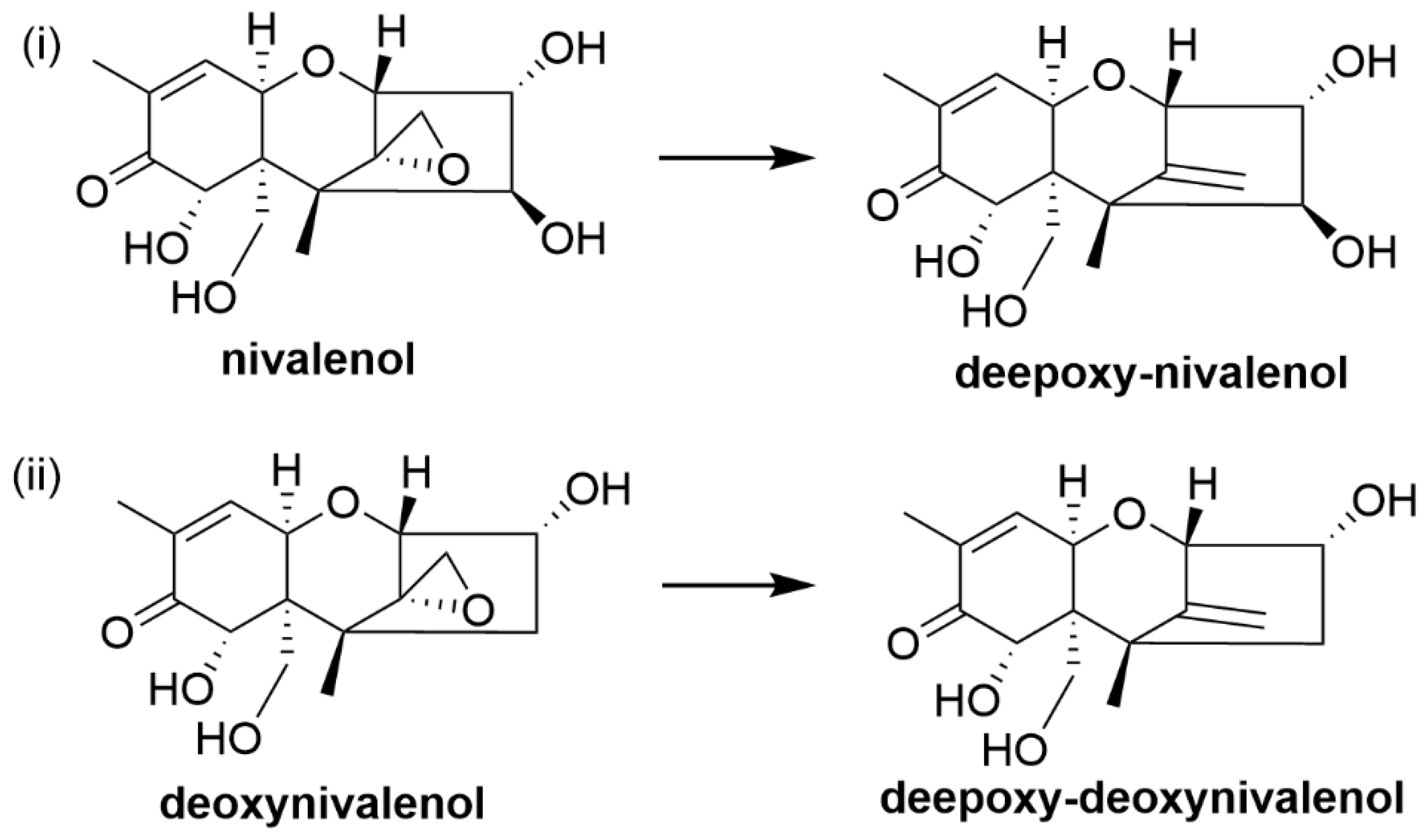
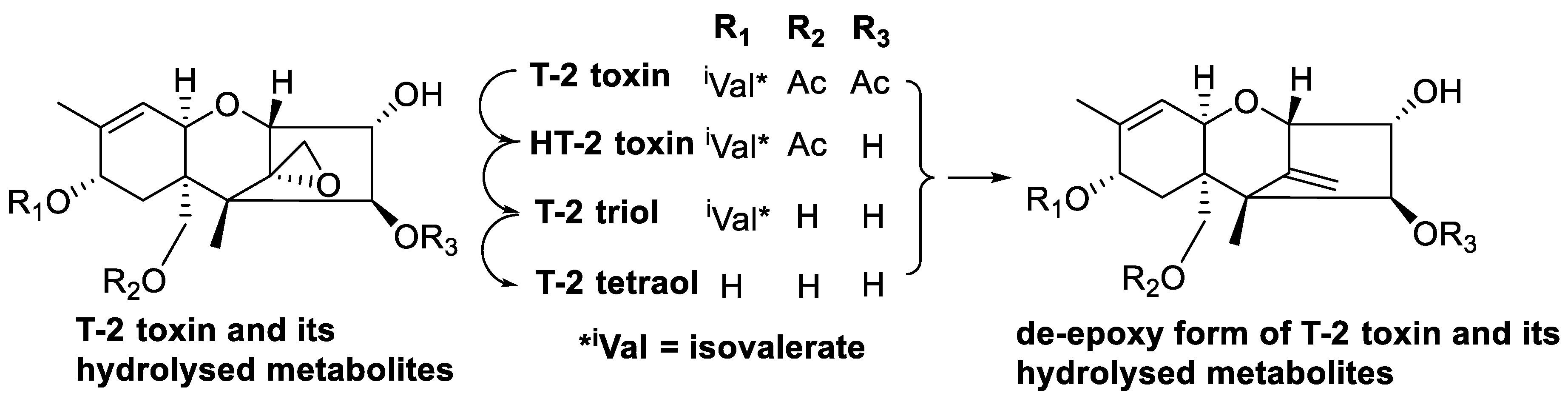
4.1. Trichothecenes
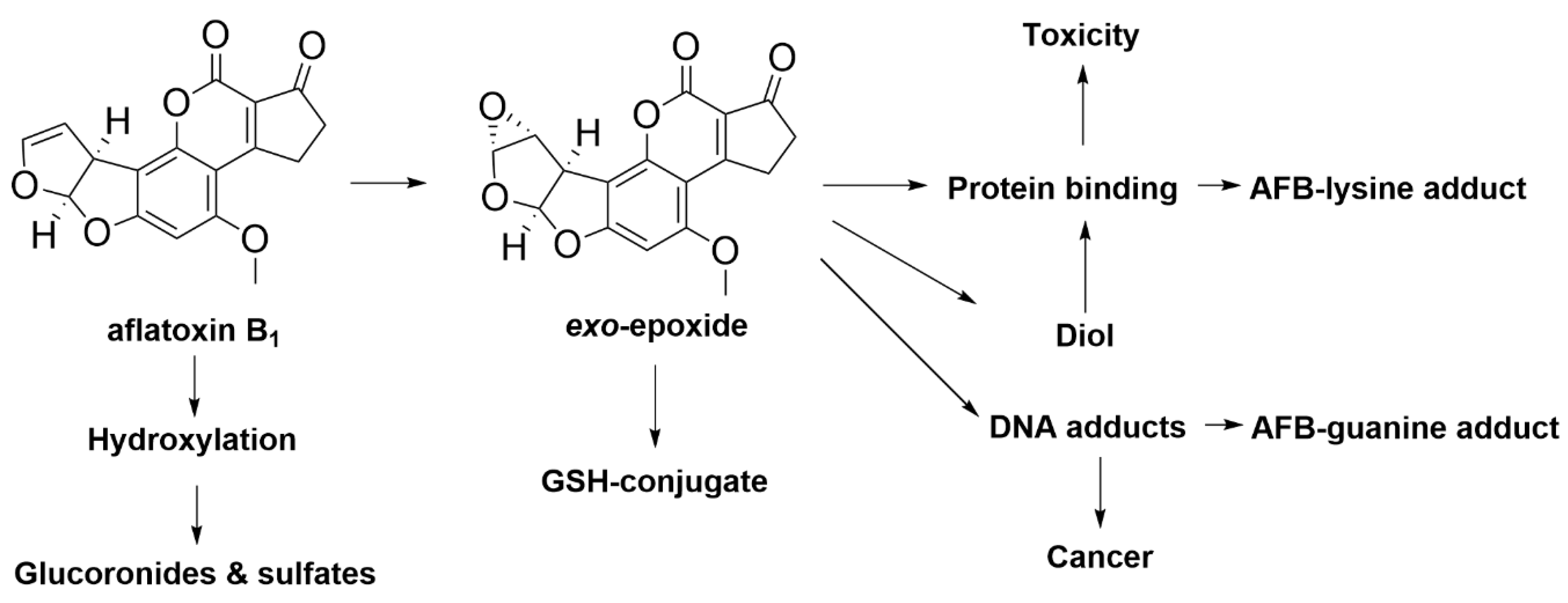
4.2. Aflatoxin B1
4.3. Ochratoxin A
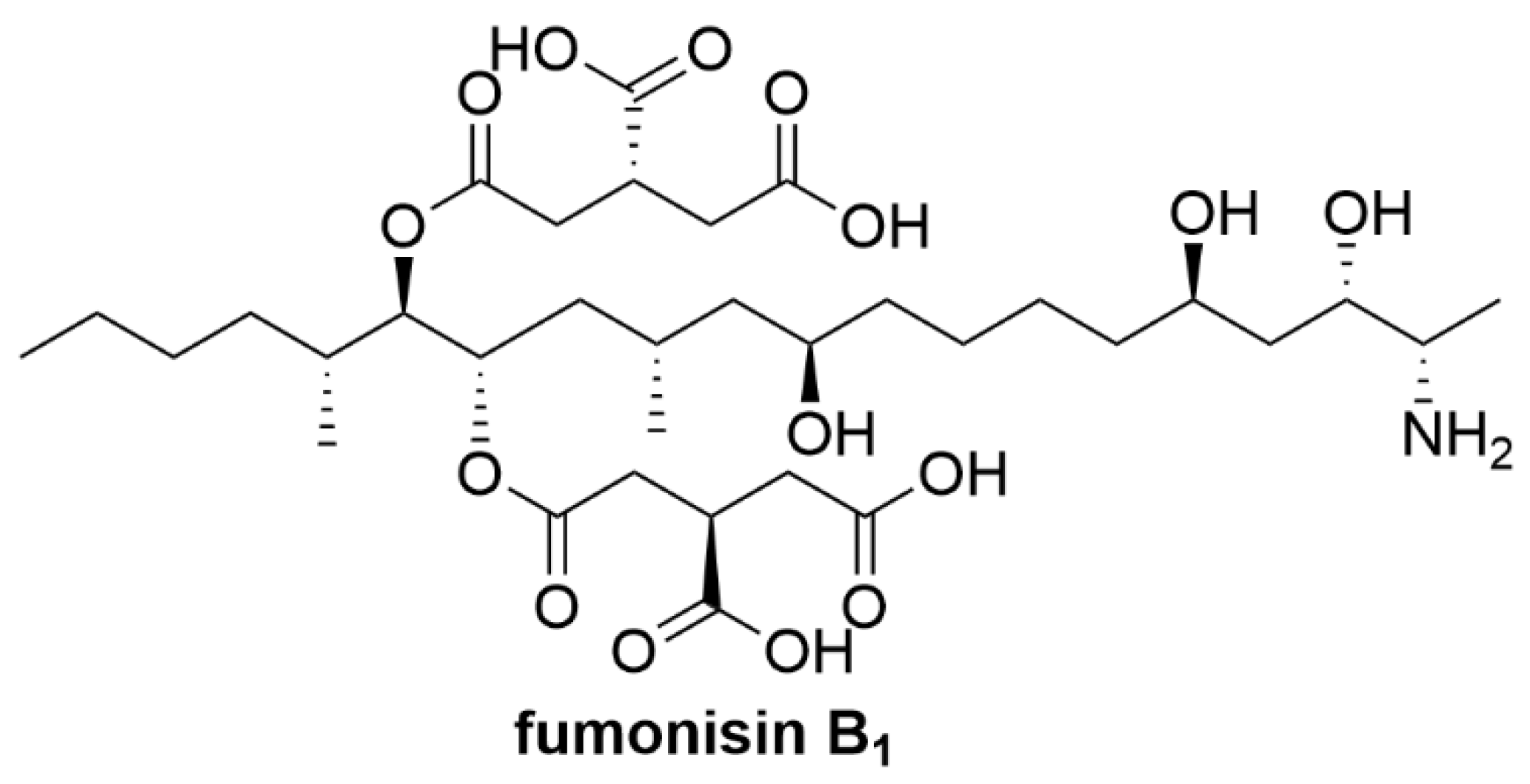
4.4. Fumonisins
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kabera, J.N.; Semana, E.; Mussa, A.R.; He, X. Plant secondary metabolites: Biosynthesis, classification, function and pharmacological properties. J. Pharm. Pharmacol. 2014, 2, 377–392. [Google Scholar]
- Yang, L.; Wen, K.; Ruan, X.; Zhao, Y.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, T. From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef]
- McSweeney, C.; Mackie, R. Micro-organisms and Ruminant Digestion: State of Knowledge, Trends and Future Prospects; Background Study Paper No. 61; Food and Agriculture Organization of the United Nation: Rome, Italy, 2012. [Google Scholar]
- Krause, D.; Smith, W.; Brooker, J.; McSweeney, C. Tolerance mechanisms of streptococci to hydrolysable and condensed tannins. Anim. Feed Sci. Technol. 2005, 121, 59–75. [Google Scholar] [CrossRef]
- Smith, G.S. Toxification and detoxification of plant compounds by ruminants: An overview. J. Range Manag. 1992, 45, 25–30. [Google Scholar] [CrossRef]
- Wadhwa, M.; Bakshi, M.; Makkar, H.P. Modifying gut microbiomes in large ruminants: Opportunities in non-intensive husbandry systems. Anim. Front. 2016, 6, 27–36. [Google Scholar] [CrossRef]
- Allison, M.J.; Mayberry, W.R.; McSweeney, C.S.; Stahl, D.A. Synergistes jonesii, gen. sp. nov.: A rumen bacterium that degrades toxic pyridinediols. Syst. Appl. Microbiol. 1992, 15, 522–529. [Google Scholar] [CrossRef]
- Hagler, W.; Danko, G.; Horvath, L.; Palyusik, M.; Mirocha, C. Transmission of zearalenone and its metabolite into ruminant milk. Acta Vet. Acad. Sci. Hung. 1980, 28, 209–216. [Google Scholar]
- Kuiper-Goodman, T.; Scott, P.; Watanabe, H. Risk assessment of the mycotoxin zearalenone. Regul. Toxicol. Pharmacol. 1987, 7, 253–306. [Google Scholar] [CrossRef]
- Joint Food and Agriculture Organization/World Health Organization Working Group. Guidelines for the Evaluation of Probiotics in Food; Report of a Joint FAO/WHO: London, ON, Canada, 2002. [Google Scholar]
- Retta, K.S. Role of probiotics in rumen fermentation and animal performance: A review. Int. J. Livest. Prod. 2016, 7, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Soccol, C.R.; Vandenberghe, L.P.D.S.; Spier, M.R.; Medeiros, A.B.P.; Yamaguishi, C.T.; Lindner, J.D.D.; Pandey, A.; Thomaz-Soccol, V. The potential of probiotics: A review. Food Technol. Biotechnol. 2010, 48, 413–434. [Google Scholar]
- Hegarty, M.P.; Court, R.D.; Christie, G.S.; Lee, C.P. Mimosine in Luecaena leucocephala is metabolised to a goitrogen in ruminants. Aust. Vet. J. 1976, 52, 490. [Google Scholar] [CrossRef]
- Jones, R.; Megarrity, R. Comparative toxicity responses of goats fed on Leucaena leucocephala in Australia and Hawaii. Aust. J. Agric. Res. 1983, 34, 781–790. [Google Scholar] [CrossRef]
- Odenyo, A.A.; Osuji, P.O.; Karanfil, O.; Adinew, K. Microbiological evaluation of Acacia angustissima as a protein supplement for sheep. Anim. Feed Sci. Technol. 1997, 65, 99–112. [Google Scholar] [CrossRef]
- Rao, S.; Adiga, P.; Sarma, P. The isolation and characterization of β-N-oxalyl-l-α, β-diaminopropionic acid: A neurotoxin from the seeds of Lathyrus sativus. Biochemistry 1964, 3, 432–436. [Google Scholar] [CrossRef]
- Van Moorhem, M.; Lambein, F.; Leybaert, L. Unraveling the mechanism of β-N-oxalyl-α, β-diaminopropionic acid (β-ODAP) induced excitotoxicity and oxidative stress, relevance for neurolathyrism prevention. Food Chem. Toxicol. 2011, 49, 550–555. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Al Jassim, R.A.; Cawdell-Smith, A.J. The occurrence and toxicity of indospicine to grazing animals. Agriculture 2015, 5, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Nordfeldt, S.; Younge, O.R. Toxicity of Creeping Indigo to Livestock; Hawaii Agricultural Experiment Station, University of Hawaii: Honolulu, HI, USA, 1949; p. 1. [Google Scholar]
- Jeganathan, P. Toxic effects of feeding Indigofera endecaphylla (Jacq.) to calves. Ceylon Vet. J. 1953, 1, 83–85. [Google Scholar]
- Maskasame, C. Toxicity and Nutritional Value of Some Promising Pasture Legumes to Rats and Sheep. Master’s Thesis, The University of Queensland, Brisbane, Australia, 1985. [Google Scholar]
- Dalziel, J.M. The Useful Plants of West Tropical Africa; Crown Agents for the Colonies: London, UK, 1937; p. 612. [Google Scholar]
- Yelf, J. The toxicity of creeping indigo in Fiji. Fiji Agric. J. 1959, 29, 9–10. [Google Scholar]
- Bell, A.; Newton, L.; Everist, S.; Legg, J. Acacia georginae poisoning of cattle and sheep. Aust. Vet. J. 1955, 31, 249–257. [Google Scholar] [CrossRef]
- Medeiros, R.; Neto, S.; Barbosa, R.C.; Lima, E.F.; Riet-Correa, F. Sudden bovine death from Mascagnia rigida in Northeastern Brazil. Vet. Hum. Toxicol. 2002, 44, 286–288. [Google Scholar] [PubMed]
- Sherley, M. The traditional categories of fluoroacetate poisoning signs and symptoms belie substantial underlying similarities. Toxicol. Lett. 2004, 151, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Bull, L.B.; Culvenor, C.; Dick, A. The Pyrrolizidine Alkaloids: Their Chemistry, Pathogenicity and Other Biological Properties; North-Holland Publishing Co.: Amsterdam, The Netherlands, 1968; p. 293. [Google Scholar]
- Wachenheim, D.; Blythe, L.; Craig, A. Effects of antibacterial agents on in vitro ovine ruminal biotransformation of the hepatotoxic pyrrolizidine alkaloid jacobine. Appl. Environ. Microbiol. 1992, 58, 2559–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, D.; Pfister, J.; Molyneux, R.; Kechele, P. Cynoglossum officinale toxicity in calves. J. Comp. Pathol. 1991, 104, 403–410. [Google Scholar] [CrossRef]
- Goeger, D.; Cheeke, P.; Schmitz, J.A.; Buhler, D. Toxicity of tansy ragwort (Senecio jacobaea) to goats. Am. J. Vet. Res. 1982, 43, 252–254. [Google Scholar]
- Molyneux, R.; Johnson, A.; Stuart, L. Delayed manifestation of Senecio-induced pyrrolizidine alkaloidosis in cattle. Vet. Hum. Toxicol. 1988, 30, 201–205. [Google Scholar]
- Lym, R.G.; Kirby, D.R. Cattle foraging behavior in leafy spurge (Euphorbia esula)-infested rangeland. Weed Technol. 1987, 314–318. [Google Scholar] [CrossRef]
- Seip, E.; Hecker, E. Skin irritant ingenol esters from Euphorbia esula. Planta Med. 1982, 46, 215–218. [Google Scholar] [CrossRef]
- Wood, R. Acute animal and human poisonings from cyanotoxin exposure—A review of the literature. Environ. Int. 2016, 91, 276–282. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Zhao, Q.; Pu, C.; Qiu, Z.; Zhang, R.; Shu, W. A cross-sectional investigation of chronic exposure to microcystin in relationship to childhood liver damage in the Three Gorges Reservoir Region, China. Environ. Health Perspect. 2011, 119, 1483–1488. [Google Scholar] [CrossRef] [Green Version]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, P.L.; Bhattacharya, R. The cyanobacterial toxin microcystin-LR induced DNA damage in mouse liver in vivo. Toxicology 1996, 114, 29–36. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P.; Li, L.; Xu, J. First identification of the hepatotoxic microcystins in the serum of a chronically exposed human population together with indication of hepatocellular damage. Toxicol. Sci. 2009, 108, 81–89. [Google Scholar] [CrossRef]
- Rao, P.L.; Bhattacharya, R.; Parida, M.; Jana, A.; Bhaskar, A. Freshwater cyanobacterium Microcystis aeruginosa (UTEX 2385) induced DNA damage in vivo and in vitro. Environ. Toxicol. Pharmacol. 1998, 5, 1–6. [Google Scholar] [CrossRef]
- Zhan, L.; Sakamoto, H.; Sakuraba, M.; Wu, D.-S.; Zhang, L.-S.; Suzuki, T.; Hayashi, M.; Honma, M. Genotoxicity of microcystin-LR in human lymphoblastoid TK6 cells. Mutat. Res. 2004, 557, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rocha, O.; Ansari, K.; Doohan, F. Effects of trichothecene mycotoxins on eukaryotic cells: A review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, C.; Vaughn, M.; Campbell, J.; Wei, C.; Stafford, M.; Hansen, B. Mycotoxins in Human and Animal Health; Pathotox Publishers Inc.: Park Forest, IL, USA, 1977; p. 807. [Google Scholar]
- Del Regno, M.; Adesso, S.; Popolo, A.; Quaroni, A.; Autore, G.; Severino, L.; Marzocco, S. Nivalenol induces oxidative stress and increases deoxynivalenol pro-oxidant effect in intestinal epithelial cells. Toxicol. Appl. Pharm. 2015, 285, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, L.W.; Hagler, W. Mycotoxins in dairy cattle: Occurrence, toxicity, prevention and treatment. Proc. Southwest Nutr. Conf. 2005, 124–138. [Google Scholar]
- Keyl, A.; Booth, A. Aflatoxin effects in livestock. J. Am. Oil Chem. Soc. 1971, 48, 599–604. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, H.; De Saeger, S.; De Boevre, M.; Sun, F.; Zhang, S.; Cao, X.; Wang, Z. In vitro and in vivo metabolism of ochratoxin A: A comparative study using ultra-performance liquid chromatography-quadrupole/time-of-flight hybrid mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 3579–3589. [Google Scholar] [CrossRef]
- Mobashar, M.; Hummel, J.; Blank, R.; Südekum, K.-H. Ochratoxin A in ruminants—A review on its degradation by gut microbes and effects on animals. Toxins 2010, 2, 809. [Google Scholar] [CrossRef] [Green Version]
- Mally, A.; Keim-Heusler, H.; Amberg, A.; Kurz, M.; Zepnik, H.; Mantle, P.; Völkel, W.; Hard, G.C.; Dekant, W. Biotransformation and nephrotoxicity of ochratoxin B in rats. Toxicol. Appl. Pharm. 2005, 206, 43–53. [Google Scholar] [CrossRef]
- Rutigliano, L.; Valentini, L.; Martino, N.A.; Pizzi, F.; Zanghì, A.; Dell’Aquila, M.E.; Minervini, F. Ochratoxin A at low concentrations inhibits in vitro growth of canine umbilical cord matrix mesenchymal stem cells through oxidative chromatin and DNA damage. Reprod. Toxicol. 2015, 57, 121–129. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Aljawish, A.; El-Nekeety, A.A.; Abdel-Aziem, S.H.; Hassan, N.S. Chitosan nanoparticles plus quercetin suppress the oxidative stress, modulate DNA fragmentation and gene expression in the kidney of rats fed ochratoxin A-contaminated diet. Food Chem. Toxicol. 2017, 99, 209–221. [Google Scholar] [CrossRef]
- Costa, J.G.; Saraiva, N.; Guerreiro, P.S.; Louro, H.; Silva, M.J.; Miranda, J.P.; Castro, M.; Batinic-Haberle, I.; Fernandes, A.S.; Oliveira, N.G. Ochratoxin A-induced cytotoxicity, genotoxicity and reactive oxygen species in kidney cells: An integrative approach of complementary endpoints. Food Chem. Toxicol. 2016, 87, 65–76. [Google Scholar] [CrossRef]
- Cui, J.; Liu, J.; Wu, S.; Wang, Y.; Shen, H.; Xing, L.; Wang, J.; Yan, X.; Zhang, X. Oxidative DNA damage is involved in ochratoxin A-induced G2 arrest through ataxia telangiectasia-mutated (ATM) pathways in human gastric epithelium GES-1 cells in vitro. Arch. Toxicol. 2013, 87, 1829–1840. [Google Scholar] [CrossRef]
- Rahimtula, A.; Béréziat, J.-C.; Bussacchini-Griot, V.; Bartsch, H. Lipid peroxidation as a possible cause of ochratoxin A toxicity. Biochem. Pharmacol. 1988, 37, 4469–4477. [Google Scholar] [CrossRef]
- Harwig, J.; Munro, I. Mycotoxins of possible importance in diseases of Canadian farm animals. Can. Vet. J. 1975, 16, 125–141. [Google Scholar]
- Ribelin, W.; Fukushima, K.; Still, P. The toxicity of ochratoxin to ruminants. Can. J. Comp. Med. 1978, 42, 172–176. [Google Scholar]
- Osweiler, G.; Kehrli, M.; Stabel, J.; Thurston, J.; Ross, P.; Wilson, T. Effects of fumonisin-contaminated corn screenings on growth and health of feeder calves. J. Anim. Sci. 1993, 71, 459–466. [Google Scholar] [CrossRef]
- Mathur, S.; Constable, P.D.; Eppley, R.M.; Tumbleson, M.E.; Smith, G.W.; Tranquilli, W.J.; Morin, D.E.; Haschek, W.M. Fumonisin B1 increases serum sphinganine concentration but does not alter serum sphingosine concentration or induce cardiovascular changes in milk-fed calves. Toxicol. Sci. 2001, 60, 379–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edrington, T.; Kamps-Holtzapple, C.; Harvey, R.; Kubena, L.; Elissalde, M.; Rottinghaus, G. Acute hepatic and renal toxicity in lambs dosed with fumonisin-containing culture material. J. Anim. Sci. 1995, 73, 508–515. [Google Scholar] [CrossRef] [Green Version]
- Gurung, N.; Rankins, D., Jr.; Shelby, R.; Goel, S. Effects of fumonisin B1-contaminated feeds on weanling Angora goats. J. Anim. Sci. 1998, 76, 2863–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.J.; Megarrity, R.G. Successful transfer of DHP-degrading bacteria from Hawaiian goats to Australian ruminants to overcome the toxicity of Leucaena. Aust. Vet. J. 1986, 63, 259–262. [Google Scholar] [CrossRef]
- Allison, M.J.; Hammond, A.C.; Jones, R.J. Detection of ruminal bacteria that degrade toxic dihydroxypyridine compounds produced from mimosine. Appl. Environ. Microbiol. 1990, 56, 590–594. [Google Scholar] [CrossRef] [Green Version]
- Klieve, A.; Ouwerkerk, D.; Turner, A.; Roberton, R. The production and storage of a fermentor-grown bacterial culture containing Synergistes jonesii, for protecting cattle against mimosine and 3-hydroxy-4 (1H)-pyridone toxicity from feeding on Leucaena leucocephala. Aust. J. Agric. Res. 2002, 53, 1–5. [Google Scholar] [CrossRef]
- McSweeney, C.; Blackall, L.; Collins, E.; Conlan, L.; Webb, R.; Denman, S.; Krause, D. Enrichment, isolation and characterisation of ruminal bacteria that degrade non-protein amino acids from the tropical legume Acacia angustissima. Anim. Feed Sci. Technol. 2005, 121, 191–204. [Google Scholar] [CrossRef]
- Peng, H.H.; Brooker, J. Isolation of ODAP-degrading bacteria from the sheep rumen. In Lathyrus Lathyrism Newsletter; The University of Western Australia: Perth, Australia, 2000; Volume 1, p. 33. [Google Scholar]
- Marichamy, S.; Yigzaw, Y.; Gorton, L.; Mattiasson, B. Isolation of obligate anaerobic rumen bacteria capable of degrading the neurotoxin β-ODAP (β-N-oxalyl-l-α, β-diaminopropionic acid) as evaluated by a liquid chromatography/biosensor analysis system. J. Sci. Food Agric. 2005, 85, 2027–2032. [Google Scholar] [CrossRef]
- Tan, E.T.T.; Al Jassim, R.; D’Arcy, B.R.; Fletcher, M.T. In vitro biodegradation of hepatotoxic indospicine in Indigofera spicata and its degradation derivatives by camel foregut and cattle rumen fluids. J. Agric. Food Chem. 2017, 65, 7528–7534. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.K.; Webb, R.I.; Sly, L.I.; Denman, S.E.; McSweeney, C.S. Isolation and survey of novel fluoroacetate-degrading bacteria belonging to the phylum Synergistetes. FEMS Microbiol. Ecol. 2012, 80, 671–684. [Google Scholar] [CrossRef] [Green Version]
- Looft, T.; Levine, U.; Stanton, T. Cloacibacillus porcorum sp. nov., a mucin-degrading bacterium from the swine intestinal tract and emended description of the genus Cloacibacillus. Int. J. Syst. Evol. Microbiol. 2013, 63, 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Camboim, E.K.; Almeida, A.P.; Tadra-Sfeir, M.Z.; Junior, F.G.; Andrade, P.P.; McSweeney, C.S.; Melo, M.A.; Riet-Correa, F. Isolation and identification of sodium fluoroacetate degrading bacteria from caprine rumen in Brazil. Sci. World J. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.; Khan, S.; Webb, R.; Denman, S.; McSweeney, C. Characterisation and survey in cattle of a rumen Pyrimadobacter sp. which degrades the plant toxin fluoroacetate. FEMS Microbiol. Ecol. 2020, 1–11. [Google Scholar] [CrossRef]
- Gregg, K.; Cooper, C.L.; Schafer, D.J.; Sharpe, H.; Beard, C.E.; Allen, G.; Xu, J. Detoxification of the plant toxin fluoroacetate by a genetically modified rumen bacterium. Bio/Technology 1994, 12, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Gregg, K.; Hamdorf, B.; Henderson, K.; Kopecny, J.; Wong, C. Genetically modified ruminal bacteria protect sheep from fluoroacetate poisoning. Appl. Environ. Microbiol. 1998, 64, 3496–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanigan, G.W. Peptococcus heliotrinreducans, sp.nov., a cytochrome-producing anaerobe which metabolizes pyrrolizidine alkaloids. Microbiology 1976, 94, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, G.; Smith, R. Reduction of heliotrine by a rumen microorganism. Aust. J. Biol. Sci. 1968, 21, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Hovermale, J.T.; Craig, A.M. Metabolism of pyrrolizidine alkaloids by Peptostreptococcus heliotrinreducens and a mixed culture derived from ovine ruminal fluid. Biophys. Chem. 2002, 101–102, 387–399. [Google Scholar] [CrossRef]
- Rattray, R.M.; Craig, A.M. Molecular characterization of sheep ruminal enrichments that detoxify pyrrolizidine alkaloids by denaturing gradient gel electrophoresis and cloning. Microb. Ecol. 2007, 54, 264–275. [Google Scholar] [CrossRef]
- Kronberg, S.L.; Walker, J.W. Ruminal metabolism of leafy spurge in sheep and goats: A potential explanation for differential foraging on spurge by sheep, goats, and cattle. J. Chem. Ecol. 1993, 19, 2007–2017. [Google Scholar] [CrossRef]
- Waterman, R.C.; Richardson, K.D.; Lodge-Ivey, S.L. Effects of Euphorbia esula L. (leafy spurge) on cattle and sheep in vitro fermentation and gas production. J. Sci. Food Agric. 2011, 91, 2053–2060. [Google Scholar] [CrossRef] [PubMed]
- Manubolu, M.; Madawala, S.R.; Dutta, P.C.; Malmlöf, K. In vitro biodegradation of cyanotoxins in the rumen fluid of cattle. BMC Vet. Res. 2014, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- King, R.R.; McQueen, R.E.; Levesque, D.; Greenhalgh, R. Transformation of deoxynivalenol (vomitoxin) by rumen microorganisms. J. Agric. Food Chem. 1984, 32, 1181–1183. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Cote, L.-M.; Swanson, S.; Buck, W. Confirmation of DOM-1, a de-epoxidation metabolite of deoxynivalenol, in biological fluids of lactating cows. Agric. Biol. Chem. 1986, 50, 227–229. [Google Scholar] [CrossRef]
- Hedman, R.; Pettersson, H. Transformation of nivalenol by gastrointestinal microbes. Arch. Anim. Nutr. 1997, 50, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Prelusky, D.B.; Veira, D.M.; Trenholm, H.L.; Hartin, K.E. Excretion profiles of the mycotoxin deoxynivalenol, following oral and intravenous administration to sheep. Toxicol. Sci. 1986, 6, 356–363. [Google Scholar] [CrossRef]
- He, P.; Young, L.; Forsberg, C. Microbial transformation of deoxynivalenol (vomitoxin). Appl. Environ. Microbiol. 1992, 58, 3857–3863. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, E.; Binder, E.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef]
- Verheecke, C.; Liboz, T.; Mathieu, F. Microbial degradation of aflatoxin B1: Current status and future advances. Int. J. Food Microbiol. 2016, 237, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Schatzmayr, G.; Zehner, F.; Täubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res. 2006, 50, 543–551. [Google Scholar] [CrossRef]
- Galtier, P.; Alvinerie, M. In vitro transformation of ochratoxin A by animal microbial floras. Ann. Res. Vet. 1976, 7, 91–98. [Google Scholar]
- Kiessling, K.-H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hult, K.; Teiling, A.; Gatenbeck, S. Degradation of ochratoxin A by a ruminant. Appl. Environ. Microbiol. 1976, 32, 443–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhaya, S.D.; Yang, L.; Seo, J.K.; Kim, M.H.; Lee, C.K.; Lee, C.H.; Ha, J.K. Effect of feed types on ochratoxin A disappearance in goat rumen fluid. Asian-Australas. J. Anim. Sci. 2010, 24, 198–205. [Google Scholar] [CrossRef]
- Caloni, F.; Spotti, M.; Auerbach, H.; den Camp, H.O.; Gremmels, J.F.; Pompa, G. In vitro metabolism of fumonisin B1 by ruminal microflora. Vet. Res. Commun. 2000, 24, 379–387. [Google Scholar] [CrossRef]
- Gurung, N.; Rankins, D., Jr.; Shelby, R. In vitro ruminal disappearance of fumonisin B1 and its effects on in vitro dry matter disappearance. Vet. Hum. Toxicol. 1999, 41, 196–199. [Google Scholar]
- Nunn, P.B.; Bell, E.A.; Watson, A.A.; Nash, R.J. Toxicity of non-protein amino acids to humans and domestic animals. Nat. Prod. Commun. 2010, 5, 485–504. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Corrêa, K.C.S.; Fett-Neto, A.G. Abiotic stresses and non-protein amino acids in plants. Crit. Rev. Plant. Sci. 2019, 38, 411–430. [Google Scholar] [CrossRef]
- Staszek, P.; Weston, L.A.; Ciacka, K.; Krasuska, U.; Gniazdowska, A. l-Canavanine: How does a simple non-protein amino acid inhibit cellular function in a diverse living system? Phytochem. Rev. 2017, 16, 1269–1282. [Google Scholar] [CrossRef]
- Rodgers, K.J.; Samardzic, K.; Main, B.J. Toxic Nonprotein Amino Acids; Springer: Dordrecht, The Netherlands, 2015; Volume 1, p. 20. [Google Scholar]
- Derakhshani, H.; Corley, S.W.; Al Jassim, R. Isolation and characterization of mimosine, 3, 4 DHP and 2, 3 DHP degrading bacteria from a commercial rumen inoculum. J. Basic Microbiol. 2016, 56, 580–585. [Google Scholar] [CrossRef]
- Vietmeyer, N.; Cottom, B.; Ruskin, F. Leucaena, Promising Forage and Tree Crop for the Tropics; National Academy of Sciences: Washington, DC, USA, 1977; p. 115. [Google Scholar]
- Gray, S. A review of research on Leucaena leucocephala. Trop. Grassl. 1968, 2, 19–30. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Gonzalez, V.; Plucknett, D.L. Varietal Variation and Yield Trials of Leucaena leucocephala (Koa Haole) in Hawaii; University of Hawaii: Honolulu, HI, USA, 1972; pp. 1–29. [Google Scholar]
- Hegarty, M.; Schinckel, P. Reaction of sheep to the consumption of Leucaena glauca Benth. and to its toxic principle mimosine. Aust. J. Agric. Res. 1964, 15, 153–167. [Google Scholar] [CrossRef]
- Lowry, J.B.; Tangendjaja, B. Autolysis of mimosine to 3-hydroxy-4-1 (H) pyridone in green tissues of Leucaena leucocephala. J. Sci. Food Agric. 1983, 34, 529–533. [Google Scholar] [CrossRef]
- Jones, R.J. Does ruminal metabolism of mimosine explain the absence of Leucaena toxicity in Hawaii? Aust. Vet. J. 1981, 57, 55–56. [Google Scholar] [CrossRef] [PubMed]
- Halliday, M.J.; Pakereng, C.; Edison, R.G.; Ara, P.; Dida, P.R.; Nulik, J.; Kana Hau, D.; McMillan, H.E.; Shelton, H. Effectiveness of inoculation with rumen fluid containing Synergistes jonesii to control DHP toxicity in ruminants in eastern Indonesia. Trop. Grassl. 2019, 7, 252–257. [Google Scholar] [CrossRef]
- Tan, P.; Wang, X.; Wang, J. Rumen bacteria degrading toxic mimosine and dihydroxypyridine compounds in China. Acta Microbiol. Sin. 1994, 34, 379–384. [Google Scholar]
- Aung, A.; Ter, U.M.; Gessler, F.; Böhnel, H. Isolation of mimosine degrading bacteria from rumen juice and mass production by Göttingen bioreactor technology. J. Agric. Sci. Technol. Iran 2011, 1, 764–772. [Google Scholar]
- Rincón, M.; Allison, M.; Michelangeli, F.; De Sanctis, Y.; Domínguez-Bello, M. Anaerobic degradation of mimosine-derived hydroxypyridines by cell free extracts of the rumen bacterium Synergistes jonesii. FEMS Microbiol. Ecol. 1998, 27, 127–132. [Google Scholar] [CrossRef]
- Jouany, J.; Michalet-Doreau, B.; Doreau, M. Manipulation of the rumen ecosystem to support high-performance beef cattle-Review. Asian-Australas. J. Anim. Sci. 2000, 13, 96–114. [Google Scholar] [CrossRef]
- Nguyen, B.C.Q.; Tawata, S. The chemistry and biological activities of mimosine: A review. Phytother. Res. 2016, 30, 1230–1242. [Google Scholar] [CrossRef]
- Smith, A.H.; Odenyo, A.A.; Osuji, P.O.; Wallig, M.A.; Kandil, F.E.; Seigler, D.S.; Mackie, R.I. Evaluation of toxicity of Acacia angustissima in a rat bioassay. Anim. Feed Sci. Technol. 2001, 91, 41–57. [Google Scholar] [CrossRef]
- Evans, C.S.; Shah, A.J.; Adlard, M.W.; Arce, M.L.R. Non-protein amino acids in seeds of neotropical species of Acacia. Phytochemistry 1992, 32, 123–126. [Google Scholar] [CrossRef]
- Rasmussen, M.A.; Foster, J.G.; Allison, M.J. Lathyrus sylvestris (flatpea) toxicity in sheep and ruminal metabolism of flatpea neurolathyrogens. In Poisonous Plants: Proceedings of the Third International Symposium; Iowa State University Press: Ames, IA, USA, 1992; pp. 377–381. [Google Scholar]
- Yan, Z.Y.; Spencer, P.S.; Li, Z.X.; Liang, Y.M.; Wang, Y.F.; Wang, C.Y.; Li, F.M. Lathyrus sativus (grass pea) and its neurotoxin ODAP. Phytochemistry 2006, 67, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Liu, F.; Chen, P.; Jez, J.M.; Krishnan, H.B. β-N-Oxalyl-l-α, β-diaminopropionic acid (β-ODAP) content in Lathyrus sativus: The integration of nitrogen and sulfur metabolism through β-cyanoalanine synthase. Int. J. Mol. Sci. 2017, 18, 526. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, G.; Li, Y.; Wang, J.; Liang, Z. Toxicological study on grass pea vine (Lathyrus sativus L.) and its toxic-component BOAA. Sci. Agric. Sin. 1989, 22, 86–93. [Google Scholar]
- Chen, Y.; Li, Z.; Lv, F.; Bao, X.; Liu, S.; Liu, X.; Zhang, G.; Li, Y. Studies on the screening of low toxic species of Lathyrus, analysis of toxins and toxicology. J. Lanzhou Univ. (Nat. Sci.) 1992, 28, 93–98. [Google Scholar]
- Kuo, Y.H.; Bau, H.M.; Rozan, P.; Chowdhury, B.; Lambein, F. Reduction efficiency of the neurotoxin β-ODAP in low-toxin varieties of Lathyrus sativus seeds by solid state fermentation with Aspergillus oryzae and Rhizopus microsporus var chinensis. J. Sci. Food Agric. 2000, 80, 2209–2215. [Google Scholar] [CrossRef]
- Nair, A.J.; Khatri, G.; Santha, I.; Mehta, S. Cloning of ODAP degrading gene and its expression as fusion protein in Escherichia coli. J. Plant Biochem. Biotechnol. 1994, 3, 103–106. [Google Scholar] [CrossRef]
- Mabberley, D. The Plant Book. A Portable Dictionary of the Vascular Plants; Cambridge University Press: Cambridge, UK, 1997; p. 858. [Google Scholar]
- Fletcher, M.T.; Reichmann, K.G.; Ossedryver, S.M.; McKenzie, R.A.; Carter, P.D.; Blaney, B.J. Accumulation and depletion of indospicine in calves (Bos taurus) fed creeping indigo (Indigofera spicata). Anim. Prod. Sci. 2016, 58, 568–576. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Kelly, W.R.; McEwan, D.; Williams, O.J.; Cameron, R. Hepatotoxicity of dogs of horse meat contaminated with indospicine. Aust. Vet. J. 1988, 65, 337–340. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Pound, A.W. Indospicine, a hepatotoxic amino acid from Indigofera spicata: Isolation, structure, and biological studies. Aust. J. Biol. Sci. 1970, 23, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Hutton, E.M.; Windrum, G.M.; Kratzing, C.C. Studies on the toxicity of Indigofera endecaphylla. I. Toxicity for rabbits. J. Nutr. 1958, 64, 321–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FitzGerald, L.; Fletcher, M.; Paul, A.; Mansfield, C.; O’Hara, A. Hepatotoxicosis in dogs consuming a diet of camel meat contaminated with indospicine. Aust. Vet. J. 2011, 89, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, M.P. Toxic amino acids in foods of animals and man. Proc. Nutr. Soc. Aust. 1986, 11, 73–81. [Google Scholar]
- Madsen, N.P.; Hegarty, M.P. Inhibition of rat liver homogenate arginase activity in vitro by the hepatotoxic amino acid indospicine. Biochem. Pharmacol. 1970, 19, 2391–2393. [Google Scholar] [CrossRef]
- Pollitt, S. Residue Implications of Indospicine, a Toxic, Non-Protein Amino Acid. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2001. [Google Scholar]
- Young, M.P. Investigation of the Toxicity of Horsemeat due to Contamination by Indospicine. Ph.D. Thesis, University of Queensland, Brisbane, Australia, 1992. [Google Scholar]
- Pearn, J.H.; Hegarty, M. Indospicine—The teratogenic factor from Indigofera spicata extract causing cleft palate. Br. J. Exp. Pathol. 1970, 51, 34–36. [Google Scholar]
- Dowling, R.M.; McKenzie, R.A. Poisonous Plants: A field Guide; Queensland Department of Primary Industries: Brisbane, Australia, 1993; p. 164. [Google Scholar]
- Bogdan, A. Observations on palatability of some leguminous plants of Kenya. East Afr. Agric. J. 1949, 15, 38–40. [Google Scholar] [CrossRef]
- Nordfeldt, S.; Henke, L.A.; Morita, K.; Matsumoto, H.; Takahashi, M.; Younge, O.R.; Willers, E.; Cross, R. Feeding tests with Indigofera endecaphylla Jacq. (Creeping indigo) and some observations on its poisonous effects on domestic animals. In Technical Bulletin; University of Hawaii: Honolulu, HI, USA, 1952; p. 23. [Google Scholar]
- Nath, K.; Malik, N.; Singh, O. Chemical composition and nutritive value of Indigofera enneaphylla and I. cordifolia as sheep feeds. Aust. J. Exp. Agric. 1971, 11, 178–185. [Google Scholar] [CrossRef]
- De Wolfe, R.H. Kinetics and Mechanisms of Reactions of Amidines; John Wiley & Sons: Bristol, UK, 1975; Volume 1. [Google Scholar]
- Tan, E.T.; Yong, K.W.; Wong, S.-H.; D’Arcy, B.R.; Al Jassim, R.; De Voss, J.J.; Fletcher, M.T. Thermo-alkaline treatment as a practical degradation strategy to reduce indospicine contamination in camel meat. J. Agric. Food Chem. 2016, 64, 8447–8453. [Google Scholar] [CrossRef]
- Ellard, K.; Seidel, P. Development of a Sustainable Camel Industry: A Compilation of Two Reports for the Rural Industries Research and Development Corporation; Rural Industries Research & Development Corporation: Canberra, Australia, 2000; p. 70. [Google Scholar]
- Harper, D.B.; O’Hagan, D.; Murphy, C.D. Fluorinated natural products: Occurrence and biosynthesis. In Natural Production of Organohalogen Compounds; Gribble, G., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 141–169. [Google Scholar]
- Leong, L.E.X.; Khan, S.; Davis, C.K.; Denman, S.E.; McSweeney, C.S. Fluoroacetate in plants—A review of its distribution, toxicity to livestock and microbial detoxification. J. Anim. Sci. Biotechnol. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Lauble, H.; Kennedy, M.; Emptage, M.; Beinert, H.; Stout, C. The reaction of fluorocitrate with aconitase and the crystal structure of the enzyme-inhibitor complex. Proc. Natl. Acad. Sci. USA 1996, 93, 13699–13703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.T.; Cook, D.; Pfister, J.A.; Allen, J.G.; Colegate, S.M.; Riet-Correa, F.; Taylor, C.M. Monofluoroacetate-containing plants that are potentially toxic to livestock. J. Agric. Food Chem. 2014, 62, 7345–7354. [Google Scholar] [CrossRef]
- Calver, M.; King, D. Controlling vertebrate pests with fluoroacetate: Lessons in wildlife management, bio-ethics, and co-evolution. J. Biol. Educ. 1986, 20, 257–262. [Google Scholar] [CrossRef] [Green Version]
- McIlroy, J. The sensitivity of Australian animals to 1080 poison. I. Intraspecific variation and factors affecting acute toxicity. Wildl. Res. 1981, 8, 369–383. [Google Scholar] [CrossRef]
- Robison, W.H. Acute toxicity of sodium monofluoroacetate to cattle. J. Wildl. Manag. 1970, 34, 647–648. [Google Scholar] [CrossRef]
- Twigg, L.; Mead, R.; King, D. Metabolism of fluoroacetate in the skink (Tiliqua rugosa) and the rat (Rattus norvegicus). Aust. J. Biol. Sci. 1986, 39, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Twigg, L.; King, D.; Davis, H.; Saunders, D.; Mead, R. Tolerance to, and metabolism of, fluoroacetate in the emu. Wildl. Res. 1988, 15, 239–247. [Google Scholar] [CrossRef]
- Oliver, A.; King, D.; Mead, R. Fluoroacetate tolerance, a genetic marker in some Australian mammals. Aust. J. Zool. 1979, 27, 363–372. [Google Scholar] [CrossRef]
- King, D.; Oliver, A.; Mead, R. The adaptation of some Western Australian mammals to food plants containing fluoroacetate. Aust. J. Zool. 1978, 26, 699–712. [Google Scholar] [CrossRef]
- Becker, M.; Carneiro, F.M.; Oliveira, L.P.D.; Silva, M.I.V.D.; Riet-Correa, F.; Lee, S.T.; Pescador, C.A.; Nakazato, L.; Colodel, E.M. Induction and transfer of resistance to poisoning by Amorimia pubiflora in sheep whith non-toxic dosis of the plant and ruminal content. Cienc. Rural 2016, 46, 674–680. [Google Scholar] [CrossRef]
- Duarte, A.L.L.; Medeiros, R.M.T.; Carvalho, F.K.L.; Lee, S.T.; Cook, D.; Pfister, J.A.; Costa, V.M.M.; Riet-Correa, F. Induction and transfer of resistance to poisoning by Amorimia (Mascagnia) septentrionalis in goats. J. Appl. Toxicol. 2014, 34, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Fetzner, S.; Lingens, F. Bacterial dehalogenases: Biochemistry, genetics, and biotechnological applications. Microbiol. Mol. Biol. Rev. 1994, 58, 641–685. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, M.F.A.; Paula, D.A.J.; Riet-Correa, F.; Dutra, V.; Nakazato, L. Detection and characterization of bovine rumen microorganisms resistant to sodium fluoroacetate. Acta Sci. Vet. 2019, 47. [Google Scholar] [CrossRef]
- Santos, A.C.; Riet-Correa, F.; Heckler, R.F.; Lima, S.C.; Silva, M.L.; Rezende, R.; Carvalho, N.M.; Lemos, R.A. Repeated administration of non-toxic doses of sodium monofluoroacetate does not protect against poisoning by this compound in sheep. Pesqui. Vet. Bras. 2014, 34, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Uzor, P.F. Alkaloids from plants with antimalarial activity: A review of recent studies. Evid. Based Complement. Altern. Med. 2020, 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Arora, S. Alkaloids-important therapeutic secondary metabolites of plant origin. J. Crit. Rev. 2015, 2, 1–8. [Google Scholar]
- Smith, L.; Culvenor, C. Plant sources of hepatotoxic pyrrolizidine alkaloids. J. Nat. Prod. 1981, 44, 129–152. [Google Scholar] [CrossRef]
- Stegelmeier, B.; Edgar, J.; Colegate, S.; Gardner, D.; Schoch, T.; Coulombe, R.; Molyneux, R. Pyrrolizidine alkaloid plants, metabolism and toxicity. J. Nat. Toxins 1999, 8, 95–116. [Google Scholar]
- Moreira, R.; Pereira, D.M.; Valentão, P.; Andrade, P.B. Pyrrolizidine alkaloids: Chemistry, pharmacology, toxicology and food safety. Int. J. Mol. Sci. 2018, 19, 1668. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.P.; Xia, Q.; Lin, G.; Chou, M.W. Pyrrolizidine alkaloids—Genotoxicity, metabolism enzymes, metabolic activation, and mechanisms. Drug Metab. Rev. 2004, 36, 1–55. [Google Scholar] [CrossRef]
- Robertson, J.; Stevens, K. Pyrrolizidine alkaloids. Nat. Prod. Rep. 2014, 31, 1721–1788. [Google Scholar] [CrossRef]
- Zhao, Y.; Xia, Q.; Yin, J.J.; Lin, G.; Fu, P.P. Photoirradiation of dehydropyrrolizidine alkaloids—Formation of reactive oxygen species and induction of lipid peroxidation. Toxicol. Lett. 2011, 205, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Prakash, A.S.; Pereira, T.N.; Reilly, P.E.; Seawright, A.A. Pyrrolizidine alkaloids in human diet. Mutat. Res. 1999, 443, 53–67. [Google Scholar] [CrossRef]
- Huxtable, R.; Yan, C.; Wild, S.; Maxwell, S.; Cooper, R. Physicochemical and metabolic basis for the differing neurotoxicity of the pyrrolizidine alkaloids, trichodesmine and monocrotaline. Neurochem. Res. 1996, 21, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Mei, N.; Fu, P.P. Genotoxicity of pyrrolizidine alkaloids. J. Appl. Toxicol. 2010, 30, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Wiedenfeld, H.; Edgar, J. Toxicity of pyrrolizidine alkaloids to humans and ruminants. Phytochem. Rev. 2011, 10, 137–151. [Google Scholar] [CrossRef]
- Pass, D.; Hogg, G.; Russell, R.; Edgar, J.; Tence, I.; Rikard-Bell, L. Poisoning of chickens and ducks by pyrrolizidine alkaloids of Heliotropium europaeum. Aust. Vet. J. 1979, 55, 284–288. [Google Scholar] [CrossRef]
- Harper, P.; Walker, K.; Krahenbuhl, R.; Christie, B. Pyrrolizidine alkaloid poisoning in calves due to contamination of straw by Heliotropium europeum. Aust. Vet. J. 1985, 62, 382–383. [Google Scholar] [CrossRef]
- Seaman, J. Hepatogenous chronic copper poisoning in sheep associated with grazing Echium plantagineum. Aust. Vet. J. 1985, 62, 247–248. [Google Scholar] [CrossRef]
- Giesecke, P. Serum biochemistry in horses with Echium poisoning. Aust. Vet. J. 1986, 63, 90–91. [Google Scholar] [CrossRef]
- Jones, R.; Drummond, G.; Chatham, R. Heliotropium europaeum poisoning of pigs. Aust. Vet. J. 1981, 57, 395–396. [Google Scholar] [CrossRef]
- Craig, A.M.; Latham, C.J.; Blythe, L.L.; Schmotzer, W.B.; O’Connor, O.A. Metabolism of toxic pyrrolizidine alkaloids from tansy ragwort (Senecio jacobaea) in ovine ruminal fluid under anaerobic conditions. Appl. Environ. Microbiol. 1992, 58, 2730–2736. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, R.; Wink, M. Do naïve ruminants degrade alkaloids in the rumen? J. Chem. Ecol. 2005, 31, 761–787. [Google Scholar] [CrossRef]
- Damir, H.A.; Adam, S.; Tartour, G. The effects of Heliotropium ovalifolium on goats and sheep. Br. Vet. J. 1982, 138, 463–472. [Google Scholar] [CrossRef]
- Culvenor, C.; Jago, M.; Peterson, J.; Smith, L.; Payne, A.; Campbell, D.; Edgar, J.; Frahn, J. Toxicity of Echium plantagineum (Paterson’s Curse). 1. Marginal toxic effects in Merino wethers from long-term feeding. Aust. J. Agric. Res. 1984, 35, 293–304. [Google Scholar] [CrossRef]
- Lodge-Ivey, S.; Rappe, M.; Johnston, W.; Bohlken, R.; Craig, A. Molecular analysis of a consortium of ruminal microbes that detoxify pyrrolizidine alkaloids. Can. J. Microbiol. 2005, 51, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Bangsund, D.A.; Nudell, D.J.; Sell, R.S.; Leistritz, F.L. Economic analysis of using sheep to control leafy spurge. J. Range Manag. 2001, 54, 322–329. [Google Scholar] [CrossRef]
- Mora, M.J.; Hervás, G.; Mantecón, Á.R.; Busqué, J.; Frutos, P. Ruminal degradation of mountain pastures infested by Euphorbia polygalifolia in cattle and sheep: Effect of previous grazing in invaded areas. J. Sci. Food Agric. 2006, 86, 2153–2160. [Google Scholar] [CrossRef]
- Halaweish, F.T.; Kronberg, S.; Hubert, M.B.; Rice, J.A. Toxic and aversive diterpenes of Euphorbia esula. J. Chem. Ecol. 2002, 28, 1599–1611. [Google Scholar] [CrossRef]
- Richardson, K.; Kelly, W.; Reil, M.; Waterman, R.; Lodge-Ivey, S. Effects of leafy spurge (Euphorbia esula) on ruminant gas production and in vitro digestion. In Proceedings of the Western Section American Society of Animal Science, Logan, UT, USA, 21–23 June 2006; p. 66. [Google Scholar]
- Landgraf, B.K.; Fay, P.K.; Havstad, K.M. Utilization of leafy spurge (Euphorbia esula) by sheep. Weed Sci. 1984, 32, 348–352. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.W. Comparison of sheep and goat preferences for leafy spurge. J. Range Manag. 1994, 47, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.E.; Lacey, J.R.; Olson, B.E. Economic feasibility of grazing sheep on leafy spurge-infested rangeland in Montana. J. Range Manag. 1996, 49, 372–374. [Google Scholar] [CrossRef]
- Hein, D.G.; Miller, S.D. Influence of leafy spurge on forage utilization by cattle. J. Range Manag. 1992, 45, 405–407. [Google Scholar] [CrossRef]
- Winkler, J.D.; Hong, B.-C.; Bahador, A.; Kazanietz, M.G.; Blumberg, P.M. Synthesis of ingenol analogs with affinity for protein kinase C. Bioorg. Med. Chem. Lett. 1993, 3, 577–580. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S.; Francis, G.; Becker, K. Phorbol esters: Structure, biological activity, and toxicity in animals. Int. J. Toxicol. 2007, 26, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Kosten, T.; Contreras, R.J. Deficits in conditioned heart rate and taste aversion in area postrema-lesioned rats. Behav. Brain Res. 1989, 35, 9–21. [Google Scholar] [CrossRef]
- Kronberg, S.L.; Halaweish, F.T.; Hubert, M.B.; Weimer, P.J. Interactions between Euphorbia esula toxins and bovine ruminal microbes. J. Chem. Ecol. 2006, 32, 15–28. [Google Scholar] [CrossRef]
- Ettoumi, A.; El Khalloufi, F.; El Ghazali, I.; Oudra, B.; Amrani, A.; Nasri, H.; Bouaïcha, N. Bioaccumulation of Cyanobacterial Toxins in Aquatic Organisms and Its Consequences for Public Health; Nova Science Pubilshers: New York, NY, USA, 2011; pp. 1–34. [Google Scholar]
- Dawson, R. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver tumor promotion by the cyanobacterial cyclic peptide toxin microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef]
- Yoshida, T.; Makita, Y.; Nagata, S.; Tsutsumi, T.; Yoshida, F.; Sekijima, M.; Tamura, S.I.; Ueno, Y. Acute oral toxicity of microcystin-LR, a cyanobacterial hepatotoxin, in mice. Nat. Toxins 1997, 5, 91–95. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Jouany, J.-P. Mycotoxins in feeds and their fate in animals: A review. Anim. Res. 2002, 51, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Vasanthi, S.; Bhat, R.V. Mycotoxins in foods. Occurrence, health & economic significance & food control measures. Indian J. Med. Res. 1998, 108, 212–224. [Google Scholar] [PubMed]
- Upadhaya, S.D.; Park, M.; Ha, J.K. Mycotoxins and their biotransformation in the rumen: A review. Asian-Australas. J. Anim. 2010, 23, 1250–1260. [Google Scholar] [CrossRef]
- Taheur, F.B.; Kouidhi, B.; Al Qurashi, Y.M.A.; Salah-Abbès, J.B.; Chaieb, K. Biotechnology of mycotoxins detoxification using microorganisms and enzymes. Toxicon 2019, 160, 12–22. [Google Scholar] [CrossRef]
- Bertero, A.; Moretti, A.; Spicer, L.J.; Caloni, F. Fusarium molds and mycotoxins: Potential species-specific effects. Toxins 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, D.; Picken, C.; Murphy, L.; Buhr, M. Measurement of the relative binding affinity of zearalenone, alpha-zearalenol and beta-zearalenol for uterine and oviduct estrogen receptors in swine, rats and chickens: An indicator of estrogenic potencies. Comp. Biochem. Phys. C 1989, 94, 691–694. [Google Scholar] [CrossRef]
- Ueno, Y.; Tashiro, F. α-Zearalenol, a major hepatic metabolite in rats of zearalenone, an estrogenic mycotoxin of Fusarium species. J. Biochem. 1981, 89, 563–571. [Google Scholar] [CrossRef]
- Ueno, Y.; Hsieh, D.P. The toxicology of mycotoxins. Crit. Rev. Toxicol. 1985, 14, 99–132. [Google Scholar] [CrossRef]
- Pinton, P.; Oswald, I.P. Effect of deoxynivalenol and other Type B trichothecenes on the intestine: A review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, K.; Yamada, M.-A.; Saito, M. Inhibitory effect of nivalenol, a toxic metabolite of Fusarium nivale, on the growth cycle and biopolymer synthesis of HeLa cells. Jpn. J. Med. Sci. Biol. 1968, 21, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Commission Regulation (EC) No. 1126/2007 of 28 September 2007 Amending Regulation (EC) No. 1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs as Regards Fusarium Toxins in Maize and Maize Products. Off. J. Eur. Union 2007, 225, 14–17. [Google Scholar]
- Rotter, B.A. Invited review: Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. 1996, 48, 1–34. [Google Scholar] [CrossRef]
- Swanson, S.; Helaszek, C.; Buck, W.; Rood, H., Jr.; Haschek, W. The role of intestinal microflora in the metabolism of trichothecene mycotoxins. Food Chem. Toxicol. 1988, 26, 823–829. [Google Scholar] [CrossRef]
- Swanson, S.P.; Nicoletti, J.; Rood, H.D.; Buck, W.B.; Cote, L.M.; Yoshizawa, T. Metabolism of three trichothecene mycotoxins, T-2 toxin, diacetoxyscirpenol and deoxynivalenol, by bovine rumen microorganisms. J. Chromatogr. A 1987, 414, 335–342. [Google Scholar] [CrossRef]
- Eriksen, G.S.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Onji, Y.; Dohi, Y.; Aoki, Y.; Moriyama, T.; Nagami, H.; Uno, M.; Tanaka, T.; Yamazoe, Y. Deepoxynivalenol: A new metabolite of nivalenol found in the excreta of orally administered rats. J. Agric. Food Chem. 1989, 37, 478–481. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Takeda, H.; Ohi, T. Structure of a novel metabolite from deoxynivalenol, a trichothecene mycotoxin, in animals. Agric. Biol. Chem. 1983, 47, 2133–2135. [Google Scholar] [CrossRef]
- Kollarczik, B.; Gareis, M.; Hanelt, M. In vitro transformation of the Fusarium mycotoxins deoxynivalenol and zearalenone by the normal gut microflora of pigs. Nat. Toxins 1994, 2, 105–110. [Google Scholar] [CrossRef]
- Eriksen, G.S.; Pettersson, H.; Johnsen, K.; Lindberg, J. Transformation of trichothecenes in ileal digesta and faeces from pigs. Arch. Anim. Nutr. 2002, 56, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Binder, J.; Horvath, E.; Schatzmayr, G.; Ellend, N.; Danner, H.; Krska, R.; Braun, R. Screening for deoxynivalenol-detoxifying anaerobic rumen microorganisms. Cereal Res. Commun. 1997, 25, 343–346. [Google Scholar] [CrossRef]
- Binder, E.; Heidler, D.; Schatzmayr, G.; Thimm, N.; Fuchs, E.; Schuh, M.; Krska, R.; Binder, J. Microbial detoxification of mycotoxins in animal feed. In Mycotoxins and Phycotoxins in Perspective at the Turn of the Millennium, Proceedings of the 10th International IUPAC Symposium, Mycotoxins and Phycotoxins, Garujà, Brazil, 21–25 May 2000; RIVM: Utrecht, The Netherlands, 2000; pp. 271–277. [Google Scholar]
- Diaz, D.E. The Mycotoxin Blue Book, 1st ed.; Nottingham University Press: Nottingham, UK, 2005; pp. 25–56. [Google Scholar]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin B1 in affecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins 2011, 3, 566–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Commission Regulation (EU) No. 165/2010 of 26 February 2010 Amending Regulation (EC) No. 1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs as Regards Aflatoxins. Off. J. Eur. Union 2010, 50, 8–12. [Google Scholar]
- Wogan, G.N. Aflatoxins as risk factors for hepatocellular carcinoma in humans. Cancer Res. 1992, 52, 2114–2118. [Google Scholar]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Food Additives and Contaminants in Food, Proceedings of the 56th Meeting of the Joint FAO/WHO Expert Committee on Food Additives (JECFA); World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Bailey, G.S.; Williams, D.E.; Wilcox, J.S.; Loveland, P.M.; Coulombe, R.A.; Hendricks, J.D. Aflatoxin B1 carcinogenesis and its relation to DNA adduct formation and adduct persistence in sensitive and resistant salmonid fish. Carcinogenesis 1988, 9, 1919–1926. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.L.; Gallagher, E.P. Mechanisms of aflatoxin carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 1994, 34, 135–172. [Google Scholar] [CrossRef]
- Do, J.H.; Choi, D.-K. Aflatoxins: Detection, toxicity, and biosynthesis. Biotechnol. Bioprocess Eng. 2007, 12, 585–593. [Google Scholar] [CrossRef]
- Zhu, Y.; Hassan, Y.I.; Watts, C.; Zhou, T. Innovative technologies for the mitigation of mycotoxins in animal feed and ingredients—A review of recent patents. Anim. Feed. Sci. Technol. 2016, 216, 19–29. [Google Scholar] [CrossRef]
- Wang, R.; Fui, S.; Miao, C.; Feng, D. Effects of different mycotoxin adsorbents on performance, meat characteristics and blood profiles of avian broilers fed mold contaminated corn. Asian-Australas. J. Anim. 2005, 19, 72–79. [Google Scholar] [CrossRef]
- Allcroft, R.; Roberts, B.; Lloyd, M. Excretion of aflatoxin in a lactating cow. Food Chem. Toxicol. 1968, 6, 619–625. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Sung, H.G.; Lee, C.H.; Lee, S.Y.; Kim, S.W.; Cho, K.J.; Ha, J.K. Comparative study on the aflatoxin B1 degradation ability of rumen fluid from Holstein steers and Korean native goats. J. Vet. Sci. 2009, 10, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): Screening for aflatoxin B1 and ochratoxin A detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Ma, Q.; Zhao, L.; Lei, Y.; Shan, Y.; Ji, C. Isolation of Bacillus subtilis: Screening for aflatoxins B1, M1 and G1 detoxification. Eur. Food Res. Technol. 2011, 232, 957–962. [Google Scholar] [CrossRef]
- Zuo, R.; Chang, J.; Yin, Q.; Wang, P.; Yang, Y.; Wang, X.; Wang, G.; Zheng, Q. Effect of the combined probiotics with aflatoxin B1-degrading enzyme on aflatoxin detoxification, broiler production performance and hepatic enzyme gene expression. Food Chem. Toxicol. 2013, 59, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kong, Q.; Chi, C.; Shan, S.; Guan, B. Biotransformation of aflatoxin B1 and aflatoxin G1 in peanut meal by anaerobic solid fermentation of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus. Int. J. Food Microbiol. 2015, 211, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Megalla, S.E.; Mohran, M.A. Fate of aflatoxin B1 in fermented dairy products. Mycopathologia 1984, 88, 27–29. [Google Scholar] [CrossRef]
- Krogh, P. Ochratoxin A in Food; Academic Press: London, UK, 1987; pp. 97–112. [Google Scholar]
- European Commission. Commission Regulation (EU) No. 594/2012 of 5 July 2012 Amending Regulation (EC) No. 1881/2006 as Regards the Maximum Levels of the Contaminants Ochratoxin A, Non Dioxin-Like PCBs and Melamine in Foodstuffs. Off. J. Eur. Union 2012, 255, 14–17. [Google Scholar]
- Guillamont, E.M.; Lino, C.; Baeta, M.; Pena, A.; Silveira, M.; Vinuesa, J.M. A comparative study of extraction apparatus in HPLC analysis of ochratoxin A in muscle. Anal. Bioanal. Chem. 2005, 383, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Skaug, M.A. Analysis of Norwegian milk and infant formulas for ochratoxin A. Food Addit. Contam. 1999, 16, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Pozzo, L.; Cavallarin, L.; Nucera, D.; Antoniazzi, S.; Schiavone, A. A survey of ochratoxin A contamination in feeds and sera from organic and standard swine farms in northwest Italy. J. Sci. Food Agric. 2010, 90, 1467–1472. [Google Scholar] [CrossRef]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 years of research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omar, R.F.; Hasinoff, B.B.; Mejilla, F.; Rahimtula, A.D. Mechanism of ochratoxin A stimulated lipid peroxidation. Biochem. Pharmacol. 1990, 40, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Mally, A.; Pepe, G.; Ravoori, S.; Fiore, M.; Gupta, R.C.; Dekant, W.; Mosesso, P. Ochratoxin A causes DNA damage and cytogenetic effects but no DNA adducts in rats. Chem. Res. Toxicol. 2005, 18, 1253–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin-Kuan, M.; Cavin, C.; Delatour, T.; Schilter, B. Ochratoxin A carcinogenicity involves a complex network of epigenetic mechanisms. Toxicon 2008, 52, 195–202. [Google Scholar] [CrossRef]
- Kabak, B.; Brandon, E.F.; Var, I.; Blokland, M.; Sips, A.J. Effects of probiotic bacteria on the bioaccessibility of aflatoxin B1 and ochratoxin A using an in vitro digestion model under fed conditions. J. Environ. Sci. Health B 2009, 44, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Versantvoort, C.H.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.; Sips, A.J. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef]
- Hagelberg, S.; Hult, K.; Fuchs, R. Toxicokinetics of ochratoxin A in several species and its plasma-binding properties. J. Appl. Toxicol. 1989, 9, 91–96. [Google Scholar] [CrossRef]
- Oster, T.; Jayyosi, Z.; Creppy, E.E.; El Amri, H.S.; Batt, A.-M. Characterization of pig liver purified cytochrome P-450 isoenzymes for ochratoxin A metabolism studies. Toxicol. Lett. 1991, 57, 203–214. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef]
- Zepnik, H.; Pähler, A.; Schauer, U.; Dekant, W. Ochratoxin A-induced tumor formation: Is there a role of reactive ochratoxin A metabolites? Toxicol. Sci. 2001, 59, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Reljic, Z.; Zlatovic, M.; Savic-Radojevic, A.; Pekmezovic, T.; Djukanovic, L.; Matic, M.; Pljesa-Ercegovac, M.; Mimic-Oka, J.; Opsenica, D.; Simic, T. Is increased susceptibility to Balkan endemic nephropathy in carriers of common GSTA1 (*A/*B) polymorphism linked with the catalytic role of GSTA1 in ochratoxin A biotransformation? Serbian case control study and in silico analysis. Toxins 2014, 6, 2348–2362. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Madhyastha, S.; Marquardt, R.R.; Li, S.; Vodela, J.K.; Frohlich, A.; Kemppainen, B.W. Toxicity of ochratoxin A, its opened lactone form and several of its analogs: Structure–activity relationships. Toxicol. Appl. Pharm. 1996, 137, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Battacone, G.; Nudda, A.; Pulina, G. Effects of ochratoxin A on livestock production. Toxins 2010, 2, 1796–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elling, F.; Møller, T. Mycotoxic nephropathy in pigs. Bull. World Health Organ. 1973, 49, 411–418. [Google Scholar]
- Santin, E.; Paulillo, A.C.; Maiorka, P.C.; Alessi, A.C.; Krabbe, E.L.; Maiorka, A. The effects of ochratoxin/aluminosilicate interaction on the tissues and humoral immune response of broilers. Avian Pathol. 2002, 31, 73–79. [Google Scholar] [CrossRef]
- Duarte, S.C.; Lino, C.M.; Pena, A. Ochratoxin A in feed of food-producing animals: An undesirable mycotoxin with health and performance effects. Vet. Microbiol. 2011, 154, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Madhyastha, M.; Marquardt, R.; Frohlich, A. Hydrolysis of ochratoxin A by the microbial activity of digesta in the gastrointestinal tract of rats. Arch. Environ. Contam. Toxicol. 1992, 23, 468–472. [Google Scholar] [CrossRef]
- Sreemannarayana, O.; Frohlich, A.; Vitti, T.; Marquardt, R.; Abramson, D. Studies of the tolerance and disposition of ochratoxin A in young calves. J. Anim. Sci. 1988, 66, 1703–1711. [Google Scholar] [CrossRef]
- Xiao, H.; Marquardt, R.; Frohlich, A.; Phillips, G.; Vitti, T. Effect of a hay and a grain diet on the rate of hydrolysis of ochratoxin A in the rumen of sheep. J. Anim. Sci. 1991, 69, 3706–3714. [Google Scholar] [CrossRef]
- Schjøth, J.E.; Visconti, A.; Sundheim, L. Fumonisins in maize in relation to climate, planting time and hybrids in two agroecological zones in Zambia. Mycopathologia 2009, 167, 209–219. [Google Scholar] [CrossRef]
- Ahangarkani, F.; Rouhi, S.; Gholamour Azizi, I. A review on incidence and toxicity of fumonisins. Toxin Rev. 2014, 33, 95–100. [Google Scholar] [CrossRef]
- Voss, K.; Smith, G.; Haschek, W. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed. Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- Moreno, E.C.; Garcia, G.T.; Ono, M.A.; Vizoni, É.; Kawamura, O.; Hirooka, E.Y.; Ono, E.Y.S. Co-occurrence of mycotoxins in corn samples from the Northern region of Paraná State, Brazil. Food Chem. 2009, 116, 220–226. [Google Scholar] [CrossRef]
- Merrill, A.H.J.; Sullards, M.C.; Wang, E.; Voss, K.A.; Riley, R.T. Sphingolipid metabolism: Roles in signal transduction and disruption by fumonisins. Environ. Health Perspect. 2001, 109, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Howard, P.C.; Eppley, R.M.; Stack, M.E.; Warbritton, A.; Voss, K.A.; Lorentzen, R.J.; Kovach, R.M.; Bucci, T.J. Fumonisin B1 carcinogenicity in a two-year feeding study using F344 rats and B6C3F1 mice. Environ. Health Perspect. 2001, 109, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Giannitti, F.; Diab, S.S.; Pacin, A.M.; Barrandeguy, M.; Larrere, C.; Ortega, J.; Uzal, F.A. Equine leukoencephalomalacia (ELEM) due to fumonisins B1 and B2 in Argentina. Pesqui. Vet. Bras. 2011, 31, 407–412. [Google Scholar] [CrossRef]
- Ross, P.F.; Rice, L.G.; Osweiler, G.D.; Nelson, P.E.; Richard, J.L.; Wilson, T.M. A review and update of animal toxicoses associated with fumonisin-contaminated feeds and production of fumonisins by Fusarium isolates. Mycopathologia 1992, 117, 109–114. [Google Scholar] [CrossRef]
- Ledoux, D.R.; Brown, T.P.; Weibking, T.S.; Rottinghaus, G.E. Fumonisin toxicity in broiler chicks. J. Vet. Diagn. Investig. 1992, 4, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Srichana, D.; Rottinghaus, G.; Srichana, P.; Porter, J.; Kerley, M.; Ledoux, D.; Spain, J.; Ellersieck, M. Effect of fumonisin on growth of ruminal bacteria in batch culture. Thammasat Int. J. Sci. Technol. 2009, 14, 13–21. [Google Scholar]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin Name | Toxin Source | Effects on Ruminants |
|---|---|---|
| Mimosine | Leucaena leucocephala | Hair loss, goitre, poor cattle live-weight gain [14] and hypothyroidism [15] |
| 4-N-acetyl-2,4-diaminobutyric acid | Acacia angustissima | Head pressing, grinding of teeth, foaming at the mouth and jerking of the body [16] |
| β-N-oxalyl-l-α,β-diaminopropionic acid | Grass pea (Lathyrus sativus) | Lathyrism [17], induces oxidative stress and excitotoxicity resulting in motor neuron degeneration [18] |
| Indospicine | Indigofera spp. (particularly I. linnaei and I. spicata) | Hepatotoxic, teratogenic, loss in body weight, embryo-lethal effects [19,20,21,22] and reproductive losses [20,23,24] |
| Fluoroacetate | Naturally in over 40 plant species. Also used as pesticide (Compound 1080) | Death [25,26], acidosis, hypocalcaemia and heart failure [27] |
| Pyrrolizidine alkaloids | About 3% of all flowering plants, including Heliotropium, Senecio, Crotalaria, Echium, and Cynoglossum species | Loss of appetite, diarrhoea and depression [28] Death [29,30,31] and megalocytosis [32] |
| Ingenol and ingenol esters | Leafy spurge (Euphorbia esula L.) | Aversion to plant [33], irritant and tumour promoter [34] |
| Microcystins and nodularin | Cyanobacteria | Sudden death, reduced animal performance [35] and potential to be carcinogenic, hepatotoxic, immunotoxic, neurotoxic and genotoxic [36,37,38,39,40,41] |
| Trichothecenes (Nivalenol, deoxynivalenol and T-2 toxin) | Fungi including Fusarium, Trichoderma, Cephalosporium, Myrothecium, Spicellum, Stachybotrys and Trichothecium | Immunosuppression, reduced growth rate, reproductive disorders, feed refusal, vomiting [42], eukaryotic protein synthesis inhibition [43] and generation of free radicals [44] |
| Aflatoxin B1 | Aspergillus fungi | Reduced animal health, performance, reproduction [45], weight loss, liver damage, decreased milk yield and reduced feed utilisation efficiency [46] |
| Ochratoxin A | Aspergillus and Penicillium fungi | Nephrotoxic, hepatotoxic, teratogenic, carcinogenic [47,48,49,50], formation of free radicals [51,52,53,54] and fatal poisoning [55,56] |
| Fumonisins | Fusarium verticillioides and Fusarium proliferatum fungi | Liver, kidney damage [57,58,59,60] and lymphocyte blastogenesis [57] |
| Toxin | Identified Rumen Microorganisms | Role in Toxin Degradation |
|---|---|---|
| Mimosine | Synergistes jonesii | Degrades toxic mimosine metabolites, 3,4-dihydroxypyridine and 2,3-dihydroxypyridine into unidentified non-toxic metabolites [15,61,62,63] |
| 4-N-acetyl-2,4-diaminobutyric acid | Bacteria not identified | Hydrolyses 4-N-acetyl-2,4-diaminobutyric acid to diaminobutyric acid and diaminopropane followed by further degradation into non-toxic metabolites [64] |
| Diaminopropionic acid | Firmicutes strain LPLR3; Klebsiella strain LPSR1 | Degrade diaminopropionic acid to further non-toxic metabolites [64] |
| β-N-oxalyl-l-α,β-diaminopropionic acid | Megasphaera elsdenii; Clostridium bifermentans | Degradation pathway and metabolites not identified [65,66] |
| Indospicine | Bacteria not identified | Hydrolyses indospicine to 2-aminopimelamic acid and 2-aminopimelic acid followed by further metabolism to hypothesised non-toxic metabolites [67] |
| Fluoroacetate | Synergistes, Pigmentiphaga, Ancylobacter, Pyramidobacter spp.; Butyrivibro fibrisolvens genetically modified with dehalogenase gene from Moraxella sp. strain B | Degrade toxic fluoroacetate into non-toxic fluoride and acetate [68,69,70,71,72,73] |
| Pyrrolizidine alkaloids | Peptostreptococcus heliotrinreducens; L4M2 mixed rumen bacterial culture from sheep rumen, containing bacterial species Anaerovibrio, Desulfovibrio, Megasphaera, Prevotella and Synergistes | Reduce pyrrolizidine alkaloid, heliotrine into non-toxic 7α-hydroxy-1-methylene-8α-pyrrolizidine and heliotric acid [74,75,76,77] |
| Ingenol and ingenol esters | Bacteria not identified | Degradation pathway and metabolites not identified [78,79] |
| Microcystins and nodularin | Bacteria not identified | Toxin degradation observed but degradation pathway was not identified [80] |
| Nivalenol and deoxynivalenol | Eubacterium strain BBSH 797 | Nivalenol and deoxynivalenol degraded into their less-toxic de-epoxide metabolites [81,82,83,84,85] |
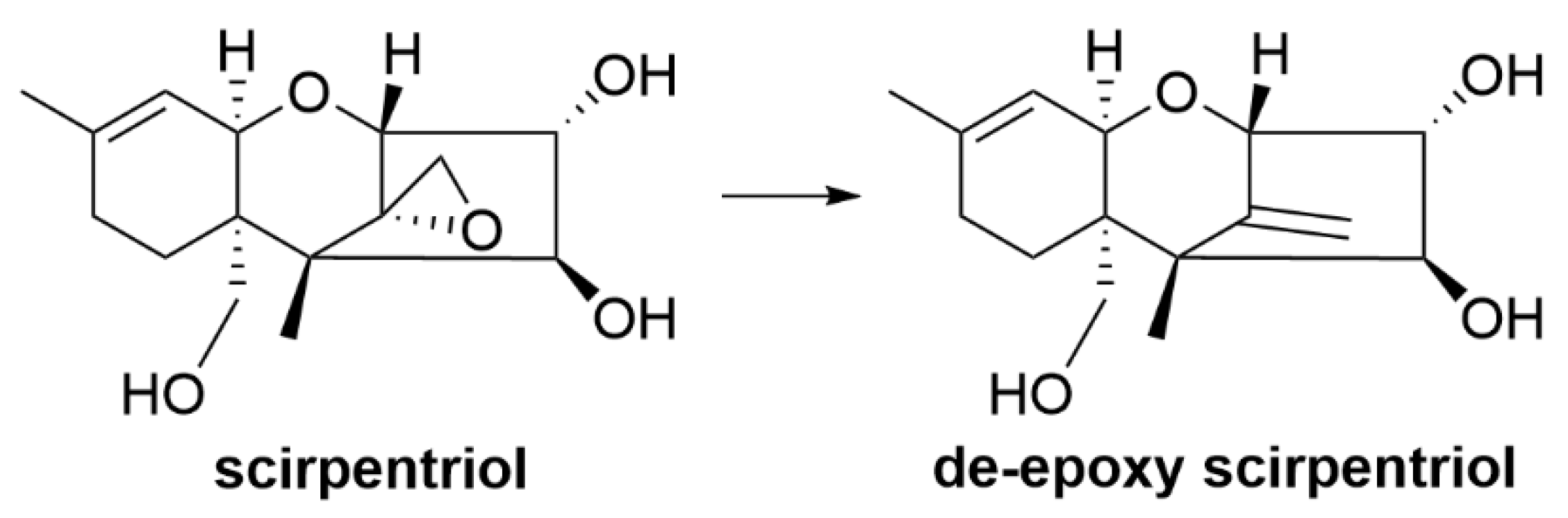
| T-2 toxin and scirpentriol | Eubacterium strain BBSH 797 | T-2 toxin and scirpentriol degraded into their less-toxic de-epoxide metabolites [86] |
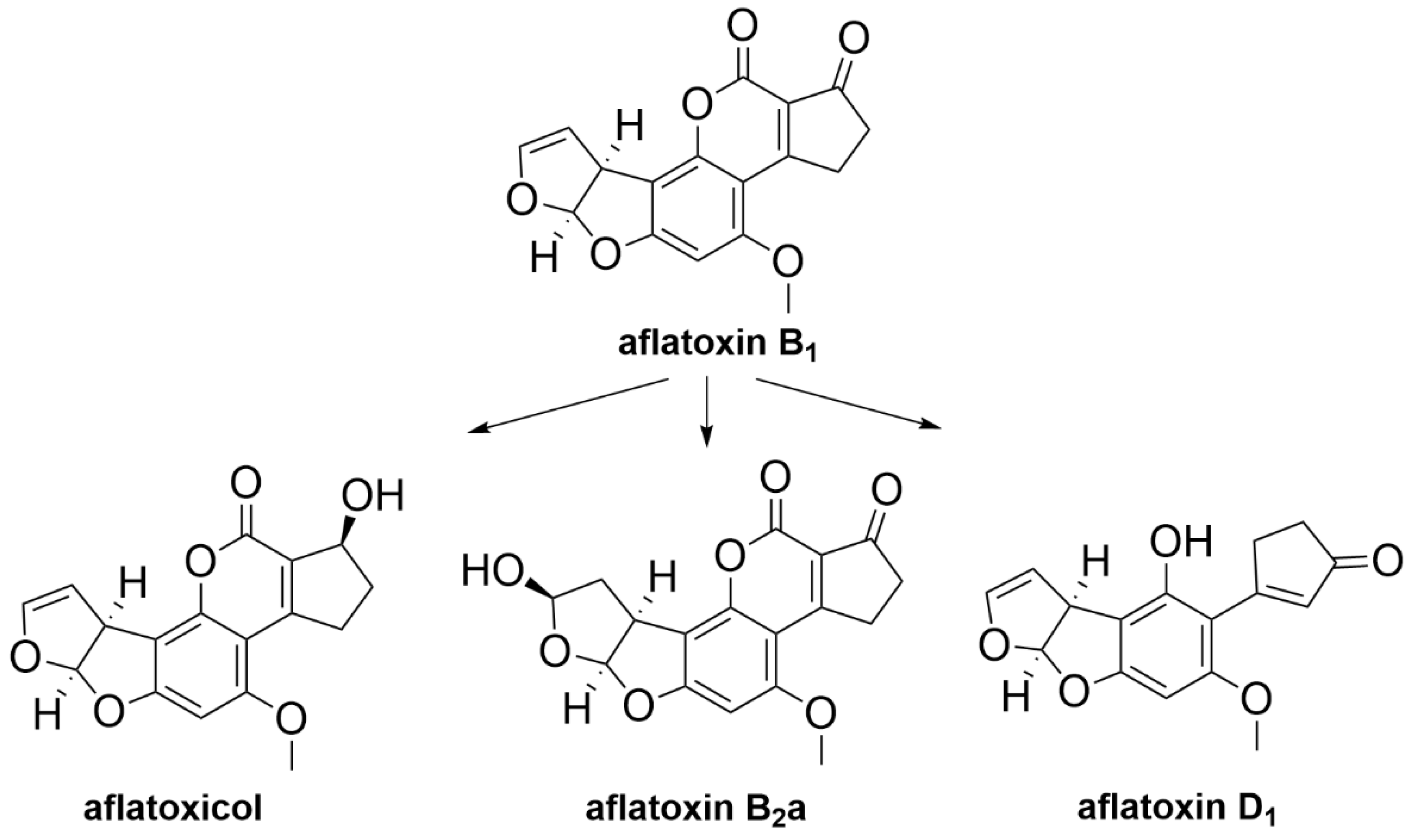
| Aflatoxin B1 | Streptococcus sp. and Lactobacillus sp. which may be present in the rumen | Degradation of aflatoxin B1 into less toxic aflatoxicol, less toxic aflatoxin B2a and non-toxic aflatoxin D1 [87] |
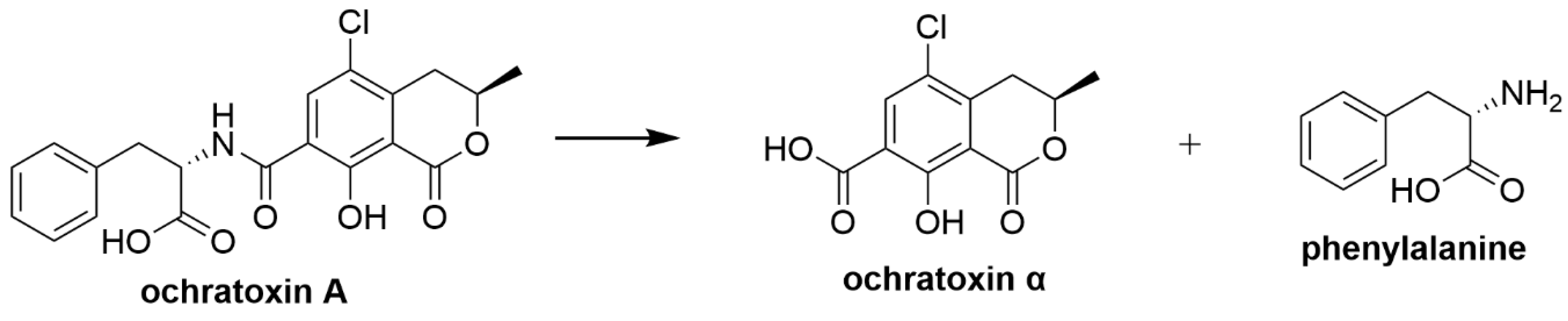
| Ochratoxin A | Rumen protozoa; Bacillus lichenformis; Lactobacillus vitulinus | Hydrolysis of ochratoxin A into non-toxic ochratoxin α and phenylalanine [48,88,89,90,91,92] |
| Fumonisins | Bacteria not identified | Degradation pathway and metabolites not identified [93,94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loh, Z.H.; Ouwerkerk, D.; Klieve, A.V.; Hungerford, N.L.; Fletcher, M.T. Toxin Degradation by Rumen Microorganisms: A Review. Toxins 2020, 12, 664. https://doi.org/10.3390/toxins12100664
Loh ZH, Ouwerkerk D, Klieve AV, Hungerford NL, Fletcher MT. Toxin Degradation by Rumen Microorganisms: A Review. Toxins. 2020; 12(10):664. https://doi.org/10.3390/toxins12100664
Chicago/Turabian StyleLoh, Zhi Hung, Diane Ouwerkerk, Athol V. Klieve, Natasha L. Hungerford, and Mary T. Fletcher. 2020. "Toxin Degradation by Rumen Microorganisms: A Review" Toxins 12, no. 10: 664. https://doi.org/10.3390/toxins12100664
APA StyleLoh, Z. H., Ouwerkerk, D., Klieve, A. V., Hungerford, N. L., & Fletcher, M. T. (2020). Toxin Degradation by Rumen Microorganisms: A Review. Toxins, 12(10), 664. https://doi.org/10.3390/toxins12100664