Transcriptomic Insights into the Antifungal Effects of Magnolol on the Growth and Mycotoxin Production of Alternaria alternata



Abstract
:1. Introduction
2. Results
2.1. Antifungal Effects of Magnolol on A. alternata
2.2. Transcriptomic Profiles of A. alternata in Response to Magnolol
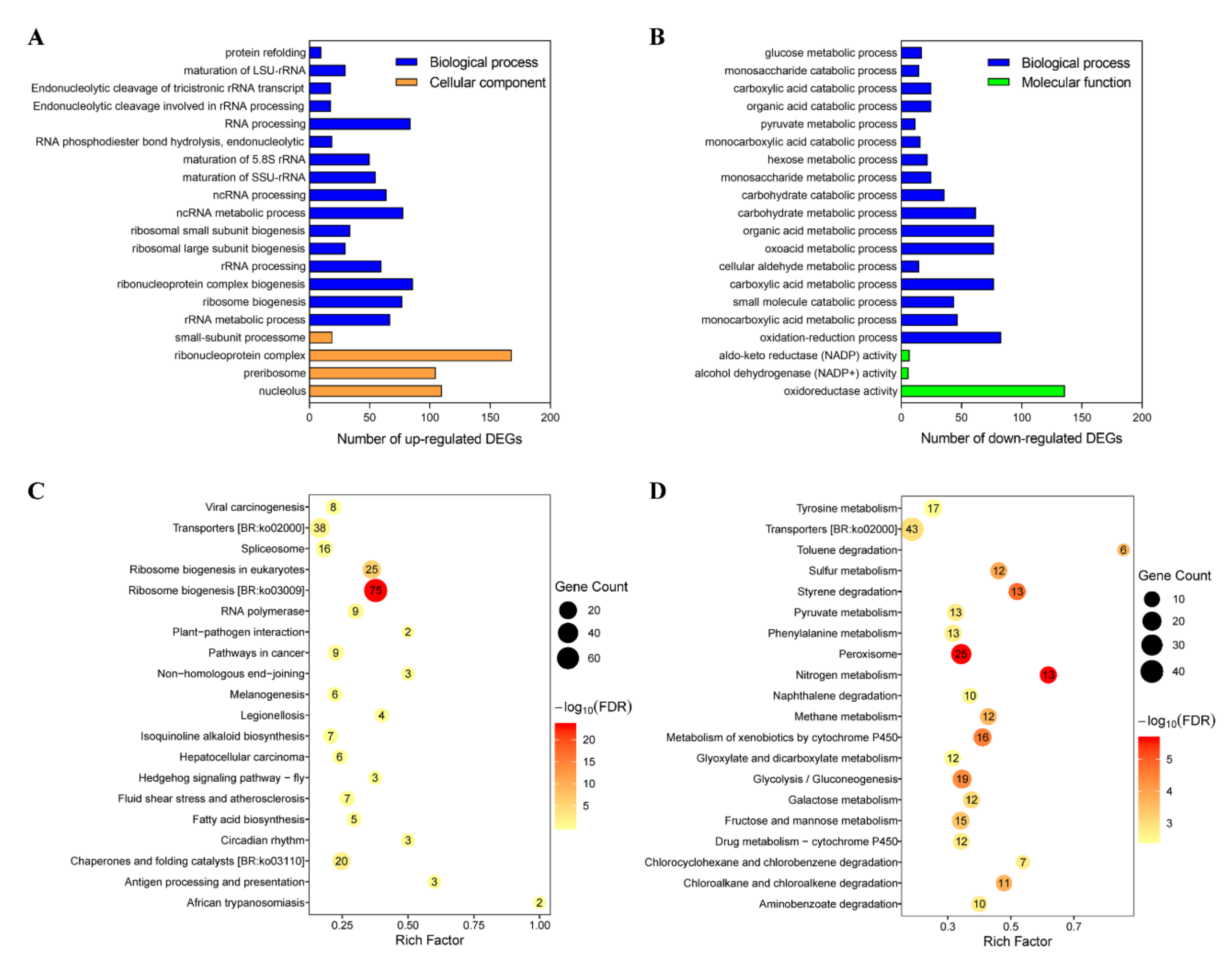
2.3. Functional Analysis of DEGs
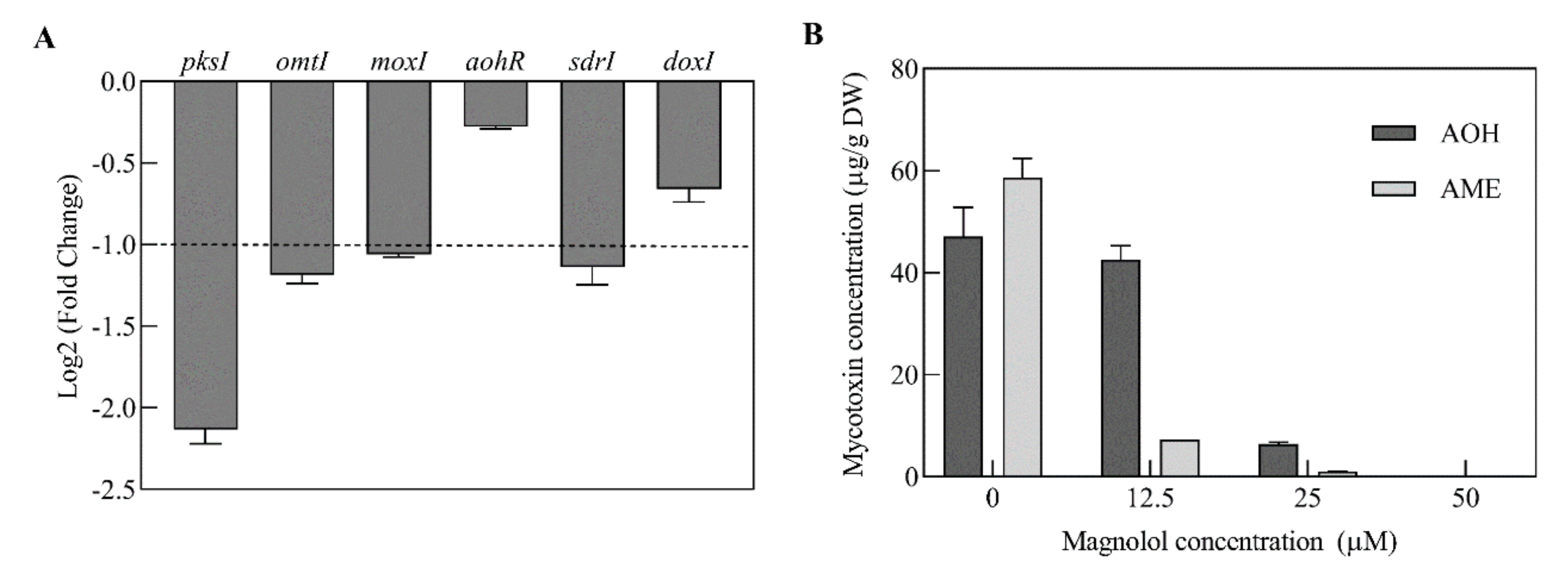
2.4. Genes Involved in Mycotoxin Biosynthesis
2.5. Genes Involved in the Primary Metabolism
2.6. Genes Related to Stress Response
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strain and Culture Condition
5.2. Antifungal Effects of Magnolol on A. alternata
5.3. Scanning Electron Microscopy (SEM) Analysis
5.4. AOH and AME Analysis
5.5. RNA Extraction, cDNA Library Construction, and RNA-Seq Analysis
5.6. Antioxidant Enzymatic Activities and Glutathione Measurement
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Logrieco, A.; Moretti, A.; Solfrizzo, M. Alternaria toxins and plant diseases: An overview of origin, occurrence and risks. World Mycotoxin J. 2009, 2, 129–140. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Reverberi, M.; Geisen, R. Mycotoxins in harvested fruits and vegetables: Insights in producing fungi, biological role, conducive conditions, and tools to manage postharvest contamination. Postharvest Biol. Technol. 2016, 122, 95–105. [Google Scholar] [CrossRef]
- Wei, D.; Wang, Y.; Jiang, D.; Feng, X.; Li, J.; Wang, M. Survey of Alternaria toxins and other mycotoxins in dried fruits in China. Toxins (Basel) 2017, 9, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solfrizzo, M. Recent advances on Alternaria mycotoxins. Curr. Opin. Food Sci. 2017, 17, 57–61. [Google Scholar] [CrossRef]
- Solhaug, A.; Eriksen, G.S.; Holme, J.A. Mechanisms of action and toxicity of the mycotoxin alternariol: A review. Basic Clin. Pharmacol. Toxicol. 2016, 119, 533–539. [Google Scholar] [CrossRef]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S.T.; Ippolito, A.; Sanzani, S.M.; Fischer, R. Alternariol as virulence and colonization factor of Alternaria alternata during plant infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef]
- Oufensou, S.; Scherm, B.; Pani, G.; Balmas, V.; Fabbri, D.; Dettori, M.A.; Carta, P.; Malbrán, I.; Migheli, Q.; Delogu, G. Honokiol, magnolol and its monoacetyl derivative show strong anti-fungal effect on Fusarium isolates of clinical relevance. PLoS ONE 2019, 14, e0221249. [Google Scholar] [CrossRef]
- Behbehani, J.; Shreaz, S.; Irshad, M.; Karched, M. The natural compound magnolol affects growth, biofilm formation, and ultrastructure of oral Candida isolates. Microb. Pathog. 2017, 113, 209–217. [Google Scholar] [CrossRef]
- Choi, N.H.; Choi, G.J.; Min, B.S.; Jang, K.S.; Choi, Y.H.; Kang, M.S.; Park, M.S.; Choi, J.E.; Bae, B.K.; Kim, J.C. Effects of neolignans from the stem bark of Magnolia obovata on plant pathogenic fungi. J. Appl. Microbiol. 2009, 106, 2057–2063. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Lu, M.-H.; Guo, D.-S.; Zhai, Y.-Y.; Miao, D.; Yue, J.; Yuan, C.; Zhao, M.; An, D. Antifungal Effect of magnolol and honokiol from Magnolia officinalis on Alternaria alternata causing tobacco brown spot. Molecules 2019, 24, 2140. [Google Scholar] [CrossRef] [Green Version]
- Sarrica, A.; Kirika, N.; Romeo, M.; Salmona, M.; Diomede, L. Safety and toxicology of magnolol and honokiol. Planta Med. 2018, 84, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzonkalik, K.; Herrling, T.; Syldatk, C.; Neumann, A. The influence of different nitrogen and carbon sources on mycotoxin production in Alternaria alternata. Int. J. Food Microbiol. 2011, 147, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Estiarte, N.; Crespo-Sempere, A.; Marín, S.; Sanchis, V.; Ramos, A.J. Exploring polyamine metabolism of Alternaria alternata to target new substances to control the fungal infection. Food Microbiol. 2017, 65, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [Green Version]
- Pfannmüller, A.; Boysen, J.M.; Tudzynski, B. Nitrate assimilation in Fusarium fujikuroi is controlled by multiple levels of regulation. Front. Microbiol. 2017, 8, 381. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon catabolite repression in filamentous fungi. Int. J. Mol. Sci. 2018, 19, 48. [Google Scholar] [CrossRef] [Green Version]
- Beattie, S.R.; Mark, K.M.K.; Thammahong, A.; Ries, L.N.A.; Dhingra, S.; Caffrey-Carr, A.K.; Cheng, C.; Black, C.C.; Bowyer, P.; Bromley, M.J.; et al. Filamentous fungal carbon catabolite repression supports metabolic plasticity and stress responses essential for disease progression. PLoS Pathog. 2017, 13, e1006340. [Google Scholar] [CrossRef]
- Yang, M.; Lu, L.; Li, S.; Zhang, J.; Li, Z.; Wu, S.; Guo, Q.; Liu, H.; Wang, C. Transcriptomic insights into benzenamine effects on the development, aflatoxin biosynthesis, and virulence of Aspergillus flavus. Toxins (Basel) 2019, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhi, Q.Q.; Li, J.Y.; Keller, N.P.; He, Z.M. The antioxidant gallic acid inhibits aflatoxin formation in Aspergillus flavus by modulating transcription factors farB and creA. Toxins (Basel) 2018, 10, 270. [Google Scholar] [CrossRef] [Green Version]
- Fasoyin, O.E.; Wang, B.; Qiu, M.; Han, X.; Chung, K.R.; Wang, S. Carbon catabolite repression gene creA regulates morphology, aflatoxin biosynthesis and virulence in Aspergillus flavus. Fungal Genet. Biol. 2018, 115, 41–51. [Google Scholar] [CrossRef]
- Da Cruz Cabral, L.; Rodríguez, A.; Delgado, J.; Patriarca, A. Understanding the effect of postharvest tomato temperatures on two toxigenic Alternaria spp. strains: Growth, mycotoxins and cell-wall integrity-related gene expression. J. Sci. Food Agric. 2019, 99, 6689–6695. [Google Scholar] [CrossRef] [PubMed]
- Maggio-Hall, L.A.; Wilson, R.A.; Keller, N.P. Fundamental contribution of β-oxidation to polyketide mycotoxin production in planta. Mol. Plant-Microbe Interact. 2005, 18, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, Y.; Antonenkov, V.D.; Glumoff, T.; Hiltunen, J.K. Peroxisomal β-oxidation-A metabolic pathway with multiple functions. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 1413–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montibus, M.; Pinson-Gadais, L.; Richard-Forget, F.; Barreau, C.; Ponts, N. Coupling of transcriptional response to oxidative stress and secondary metabolism regulation in filamentous fungi. Crit. Rev. Microbiol. 2015, 41, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Sieńko, M.; Natorff, R.; Skoneczny, M.; Kruszewska, J.; Paszewski, A.; Brzywczy, J. Regulatory mutations affecting sulfur metabolism induce environmental stress response in Aspergillus nidulans. Fungal Genet. Biol. 2014, 65, 37–47. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, N.; Wang, D.; Wang, M. Effects of essential oil citral on the growth, mycotoxin biosynthesis and transcriptomic profile of Alternaria alternata. Toxins (Basel) 2019, 11, 553. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Sun, X.; Yu, D.; Xu, J.; Chung, K.; Li, H. Genomic and transcriptomic analyses of the tangerine pathotype of Alternaria alternata in response to oxidative stress. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.H.; Lin, C.H.; Chung, K.R. A nonribosomal peptide synthetase mediates siderophore production and virulence in the citrus fungal pathogen Alternaria alternata. Mol. Plant Pathol. 2013, 14, 497–505. [Google Scholar] [CrossRef]
- Pani, G.; Scherm, B.; Azara, E.; Balmas, V.; Jahanshiri, Z.; Carta, P.; Fabbri, D.; Dettori, M.A.; Fadda, A.; Dessì, A.; et al. Natural and natural-like phenolic inhibitors of type B trichothecene in vitro production by the wheat (Triticum sp.) pathogen Fusarium culmorum. J. Agric. Food Chem. 2014, 62, 4969–4978. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yuan, S.; Sun, J.; Li, Q.; Jiang, W.; Cao, J. Ethyl p-coumarate exerts antifungal activity in vitro and in vivo against fruit Alternaria alternata via membrane-targeted mechanism. Int. J. Food Microbiol. 2018, 278, 26–35. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, N.; Wang, Y.; Jiang, D.; Feng, X. Characterization of phenolic compounds from early and late ripening sweet cherries and their antioxidant and antifungal activities. J. Agric. Food Chem. 2017, 65, 5413–5420. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Hu, S.C.S.; Yen, F.L.; Hsu, L.F.; Lee, I.T.; Lin, Z.C.; Tsai, M.H.; Huang, C.L.; Liang, C.J.; Chiang, Y.C. Magnolol nanoparticles exhibit improved water solubility and suppress TNF-α-induced VCAM-1 expression in endothelial cells. J. Biomed. Nanotechnol. 2017, 13, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Tao, N.; Yang, W.; Jing, G. Cinnamaldehyde damaged the cell membrane of Alternaria alternata and induced the degradation of mycotoxins in vivo. Ind. Crops Prod. 2018, 112, 427–433. [Google Scholar] [CrossRef]
- Palumbo, J.D.; O’Keeffe, T.L.; Mahoney, N.E. Inhibition of ochratoxin A production and growth of Aspergillus species by phenolic antioxidant compounds. Mycopathologia 2007, 164, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Atanasova-Penichon, V.; Barreau, C.; Richard-Forget, F. Antioxidant secondary metabolites in cereals: Potential involvement in resistance to Fusarium and mycotoxin accumulation. Front. Microbiol. 2016, 7, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, D.; Fetzner, R.; Burkhardt, B.; Podlech, J.; Metzler, M.; Dang, H.; Lawrence, C.; Fischer, R. Identification of a polyketide synthase required for alternariol (AOH) and alternariol-9-methyl ether (AME) formation in Alternaria alternata. PLoS ONE 2012, 7, e40564. [Google Scholar] [CrossRef]
- Wang, P.; Ma, L.; Jin, J.; Zheng, M.; Pan, L.; Zhao, Y.; Sun, X.; Liu, Y.; Xing, F. The anti-aflatoxigenic mechanism of cinnamaldehyde in Aspergillus flavus. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Jiang, T.; Zhang, X.F.; Wang, X.F.; Zhang, D.P. Arabidopsis 3-Ketoacyl-CoA Thiolase-2 (KAT2), an enzyme of fatty acid β-oxidation, is involved in ABA signal transduction. Plant Cell Physiol. 2011, 52, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef]
- Han, X.; Qiu, M.; Wang, B.; Yin, W.B.; Nie, X.; Qin, Q.; Ren, S.; Yang, K.; Zhang, F.; Zhung, Z.; et al. Functional analysis of the nitrogen metabolite repression regulator gene nmrA in Aspergillus flavus. Front. Microbiol. 2016, 7, 1794. [Google Scholar] [CrossRef] [Green Version]
- Ries, L.N.A.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse regulation of the CreA carbon catabolite repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotaka, M.; Johnson, C.; Lamb, H.K.; Hawkins, A.R.; Ren, J.; Stammers, D.K. Structural Analysis of the Recognition of the negative regulator NmrA and DNA by the zinc finger from the GATA-type transcription factor AreA. J. Mol. Biol. 2008, 381, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Giese, H.; Sondergaard, T.E.; Sørensen, J.L. The AreA transcription factor in Fusarium graminearum regulates the use of some nonpreferred nitrogen sources and secondary metabolite production. Fungal Biol. 2013, 117, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Macios, M.; Caddick, M.X.; Weglenski, P.; Scazzocchio, C.; Dzikowska, A. The GATA factors AREA and AREB together with the co-repressor NMRA, negatively regulate arginine catabolism in Aspergillus nidulans in response to nitrogen and carbon source. Fungal Genet. Biol. 2012, 49, 189–198. [Google Scholar] [CrossRef]
- Reverberi, M.; Gazzetti, K.; Punelli, F.; Scarpari, M.; Zjalic, S.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Aoyap1 regulates OTA synthesis by controlling cell redox balance in Aspergillus ochraceus. Appl. Microbiol. Biotechnol. 2012, 95, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Ponts, N.; Pinson-Gadais, L.; Barreau, C.; Richard-Forget, F.; Ouellet, T. Exogenous H2O2 and catalase treatments interfere with Tri genes expression in liquid cultures of Fusarium graminearum. FEBS Lett. 2007, 581, 443–447. [Google Scholar] [CrossRef] [Green Version]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Punelli, F.; Camera, E.; Fabbri, C.; Picardo, M.; Fanelli, C.; Fabbri, A.A. Modulation of antioxidant defense in Aspergillus parasiticus is involved in aflatoxin biosynthesis: A role for the ApyapA gene. Eukaryot. Cell 2008, 7, 988–1000. [Google Scholar] [CrossRef] [Green Version]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, K.; Yang, H.; Zhang, Z.; Yuan, Y.; Yue, T. Effect of cinnamaldehyde and citral combination on transcriptional profile, growth, oxidative damage and patulin biosynthesis of Penicillium expansum. Front. Microbiol. 2018, 9, 597. [Google Scholar] [CrossRef]
- Sun, Q.; Shang, B.; Wang, L.; Lu, Z.; Liu, Y. Cinnamaldehyde inhibits fungal growth and aflatoxin B1 biosynthesis by modulating the oxidative stress response of Aspergillus flavus. Appl. Microbiol. Biotechnol. 2016, 100, 1355–1364. [Google Scholar] [CrossRef]
- Rodríguez-Peña, J.M.; García, R.; Nombela, C.; Arroyo, J. The high-osmolarity glycerol (HOG) and cell wall integrity (CWI) signalling pathways interplay: A yeast dialogue between MAPK routes. Yeast 2010, 27, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Graf, E.; Schmidt-Heydt, M.; Geisen, R. HOG MAP kinase regulation of alternariol biosynthesis in Alternaria alternata is important for substrate colonization. Int. J. Food Microbiol. 2012, 157, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Kohut, G.; Ádám, A.L.; Fazekas, B.; Hornok, L. N-starvation stress induced FUM gene expression and fumonisin production is mediated via the HOG-type MAPK pathway in Fusarium proliferatum. Int. J. Food Microbiol. 2009, 130, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Stoll, D.; Schmidt-Heydt, M.; Geisen, R. Differences in the regulation of ochratoxin A by the HOG pathway in Penicillium and Aspergillus in response to high osmolar environments. Toxins (Basel) 2013, 5, 1282–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Cruz Cabral, L.; Delgado, J.; Patriarca, A.; Rodríguez, A. Differential response to synthetic and natural antifungals by Alternaria tenuissima in wheat simulating media: Growth, mycotoxin production and expression of a gene related to cell wall integrity. Int. J. Food Microbiol. 2019, 292, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.M.E.; Anderson, M.A.; Traven, A.; Van Der Weerden, N.L.; Bleackley, M.R. Activation of stress signalling pathways enhances tolerance of fungi to chemical fungicides and antifungal proteins. Cell. Mol. Life Sci. 2014, 71, 2651–2666. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Li, Z.; Wang, L.; Li, H.; Zhu, X.; Feng, X.; Wang, M. Effects of ultraviolet-c treatment on growth and mycotoxin production by Alternaria strains isolated from tomato fruits. Int. J. Food Microbiol. 2019, 311. [Google Scholar] [CrossRef]
- Meena, M.; Swapnil, P.; Upadhyay, R.S. Isolation, characterization and toxicological potential of Alternaria-mycotoxins (TeA, AOH and AME) in different Alternaria species from various regions of India. Sci. Rep. 2017, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Log2 (AaM/AaC) 1 | p-Value | Function |
|---|---|---|---|
| Nitrogen metabolism | |||
| CC77DRAFT_1016967 | −1.536 | 0.001 | nmrA |
| CC77DRAFT_1034395 | −2.647 | 0.000 | 2-nitropropane dioxygenase precursor |
| CC77DRAFT_1056077 | −2.376 | 0.013 | high affinity nitrate transporter NrtB |
| CC77DRAFT_937904 | −2.108 | 0.004 | nitrite reductase NiR |
| CC77DRAFT_937944 | −2.159 | 0.001 | nitrate reductase NR |
| CC77DRAFT_1011175 | −2.026 | 0.000 | cyanate hydratase |
| CC77DRAFT_383282 | −1.938 | 0.000 | formamidase FmdS |
| CC77DRAFT_937213 | −1.831 | 0.000 | carbonic anhydrase |
| CC77DRAFT_854456 | −1.897 | 0.004 | putative oxidoreductase |
| CC77DRAFT_201495 | −1.541 | 0.000 | carbon-nitrogen hydrolase |
| CC77DRAFT_1059060 | −1.677 | 0.000 | carbonic anhydrase |
| CC77DRAFT_505636 | −1.650 | 0.000 | glutamine synthetase |
| CC77DRAFT_261749 | −1.627 | 0.000 | glutamate synthase |
| CC77DRAFT_926595 | −1.065 | 0.000 | glutamine synthetase |
| Carbon utilization | |||
| CC77DRAFT_1058240 | −1.289 | 0.000 | CreA |
| CC77DRAFT_318215 | −2.128 | 0.000 | C2H2 transcription factor (AmdA) |
| CC77DRAFT_947345 | −1.766 | 0.000 | acetyl-CoA synthetase-like protein |
| CC77DRAFT_1063927 | −1.715 | 0.000 | C6 transcription factor FacB |
| CC77DRAFT_1020016 | −1.526 | 0.000 | pyruvate carboxylase |
| CC77DRAFT_30145 | −1.177 | 0.000 | mitochondrial uncoupling protein 2 |
| CC77DRAFT_997221 | −1.015 | 0.000 | mitochondrial carrier |
| CC77DRAFT_926519 | 1.412 | 0.000 | isocitrate lyase 2 |
| Cell wall biogenesis | |||
| CC77DRAFT_598626 | −1.007 | 0.000 | Rho GTPase1 |
| CC77DRAFT_1022845 | −1.230 | 0.000 | mannose-6-phosphate isomerase |
| CC77DRAFT_1066075 | −1.526 | 0.000 | mannose-6-phosphate isomerase |
| CC77DRAFT_944335 | −1.685 | 0.000 | quinone oxidoreductase putative |
| CC77DRAFT_61983 | −1.211 | 0.000 | chitinase |
| CC77DRAFT_985840 | −1.195 | 0.007 | chitin deacetylase 1 |
| CC77DRAFT_2778 | 1.515 | 0.000 | chitinase |
| CC77DRAFT_1061411 | 1.247 | 0.000 | nucleotide-diphospho-sugar transferase |
| CC77DRAFT_1039824 | 1.335 | 0.000 | hypothetical protein |
| CC77DRAFT_1064369 | 2.536 | 0.000 | glycoside hydrolase |
| CC77DRAFT_906789 | −1.612 | 0.009 | glycoside hydrolase |
| CC77DRAFT_931389 | −1.127 | 0.004 | glycoside hydrolase |
| Fatty acid beta-oxidation | |||
| CC77DRAFT_937137 | −3.031 | 0.000 | tropinone reductase 1 |
| CC77DRAFT_1024610 | −2.286 | 0.000 | 3-ketoacyl-CoA thiolase B |
| CC77DRAFT_38276 | −2.169 | 0.000 | acyl-CoA dehydrogenase |
| CC77DRAFT_1087261 | −1.963 | 0.021 | Delta3-Delta2-enoyl-CoA isomerase |
| CC77DRAFT_210867 | −1.642 | 0.000 | 3-oxoacyl-reductase |
| Gene | Log2 (AaM/AaC) 1 | p-Value | Function |
|---|---|---|---|
| Antioxidant enzyme | |||
| CC77DRAFT_905830 | 1.554 | 0.000 | Superoxide dismutase (SOD) |
| CC77DRAFT_911584 | 2.368 | 0.000 | SOD |
| CC77DRAFT_1050117 | −1.644 | 0.000 | SOD |
| CC77DRAFT_1021907 | −1.374 | 0.000 | SOD |
| CC77DRAFT_1036489 | −2.497 | 0.001 | Catalase (CAT) |
| CC77DRAFT_1013212 | −1.485 | 0.000 | CAT |
| CC77DRAFT_296007 | 3.497 | 0.000 | Peroxidase (POD) |
| CC77DRAFT_299044 | 3.421 | 0.000 | POD |
| CC77DRAFT_1039208 | 1.391 | 0.000 | POD |
| CC77DRAFT_960122 | 1.186 | 0.000 | POD |
| CC77DRAFT_1021328 | 4.768 | 0.000 | POD |
| CC77DRAFT_227615 | 1.812 | 0.000 | POD |
| CC77DRAFT_1096817 | −2.209 | 0.001 | POD |
| CC77DRAFT_1051184 | −1.353 | 0.015 | POD |
| CC77DRAFT_1008747 | −1.162 | 0.000 | POD |
| Glutathione metabolism | |||
| CC77DRAFT_1032529 | −3.152 | 0.000 | glutathione S-transferase II |
| CC77DRAFT_687848 | −1.445 | 0.000 | glutathione S-transferase II |
| CC77DRAFT_1050574 | −1.671 | 0.000 | glutamate-cysteine ligase |
| CC77DRAFT_1022906 | −1.210 | 0.000 | glutamate-cysteine ligase regulatory subunit |
| CC77DRAFT_77574 | −2.371 | 0.000 | thioredoxin-like protein |
| CC77DRAFT_1056344 | 1.282 | 0.001 | glutathione S-transferase |
| CC77DRAFT_356794 | 1.067 | 0.001 | glutathione S-transferase |
| Siderophore biosynthesis | |||
| CC77DRAFT_935643 | 1.464 | 0.000 | tyrosinase |
| CC77DRAFT_1061171 | 2.376 | 0.000 | Non-ribosomal peptide synthase, NPS6 |
| CC77DRAFT_1031046 | 2.732 | 0.000 | ABC transporter |
| CC77DRAFT_1020276 | 4.384 | 0.000 | L-ornithine 5-monooxygenase |
| CC77DRAFT_69570 | 2.126 | 0.000 | aerobactin siderophore biosynthesis protein iucB |
| CC77DRAFT_69691 | 4.699 | 0.000 | short-chain-fatty-acid-CoA ligase, sidI |
| CC77DRAFT_71629 | 4.417 | 0.000 | siderophore iron transporter, mirB |
| CC77DRAFT_69581 | −2.210 | 0.000 | MFS transporter |
| Sulfur metabolism | |||
| CC77DRAFT_1015694 | −3.069 | 0.000 | thiosulfate sulfurtransferas-like protein |
| CC77DRAFT_1020633 | −1.427 | 0.000 | sulfite reductase hemoprotein |
| CC77DRAFT_1022955 | −3.093 | 0.000 | o-acetylhomoserine ami |
| CC77DRAFT_1067622 | −2.515 | 0.000 | molybdopterin binding oxidoreductase |
| CC77DRAFT_1099663 | −1.502 | 0.000 | sulfide: quinone oxidoreductase |
| CC77DRAFT_675919 | −1.094 | 0.000 | carbohydrate phosphatase |
| CC77DRAFT_786633 | −1.703 | 0.000 | adenylyl-sulfate kinase |
| CC77DRAFT_925110 | −2.268 | 0.000 | phosphoadenosine phosphosulfate reductase |
| CC77DRAFT_951586 | −1.374 | 0.000 | ATP-sulfurylase |
| CC77DRAFT_971236 | −5.474 | 0.000 | methanesulfonate monooxygenase |
| CC77DRAFT_192219 | −1.215 | 0.000 | alpha/beta-hydrolase |
| CC77DRAFT_240786 | −1.437 | 0.000 | homoserine acetyltransferase family protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wang, D.; Yuan, S.; Feng, X.; Wang, M. Transcriptomic Insights into the Antifungal Effects of Magnolol on the Growth and Mycotoxin Production of Alternaria alternata. Toxins 2020, 12, 665. https://doi.org/10.3390/toxins12100665
Wang L, Wang D, Yuan S, Feng X, Wang M. Transcriptomic Insights into the Antifungal Effects of Magnolol on the Growth and Mycotoxin Production of Alternaria alternata. Toxins. 2020; 12(10):665. https://doi.org/10.3390/toxins12100665
Chicago/Turabian StyleWang, Liuqing, Duo Wang, Shuzhi Yuan, Xiaoyuan Feng, and Meng Wang. 2020. "Transcriptomic Insights into the Antifungal Effects of Magnolol on the Growth and Mycotoxin Production of Alternaria alternata" Toxins 12, no. 10: 665. https://doi.org/10.3390/toxins12100665
APA StyleWang, L., Wang, D., Yuan, S., Feng, X., & Wang, M. (2020). Transcriptomic Insights into the Antifungal Effects of Magnolol on the Growth and Mycotoxin Production of Alternaria alternata. Toxins, 12(10), 665. https://doi.org/10.3390/toxins12100665