Dimethylformamide Inhibits Fungal Growth and Aflatoxin B1 Biosynthesis in Aspergillus flavus by Down-Regulating Glucose Metabolism and Amino Acid Biosynthesis



Abstract
:1. Introduction
2. Results
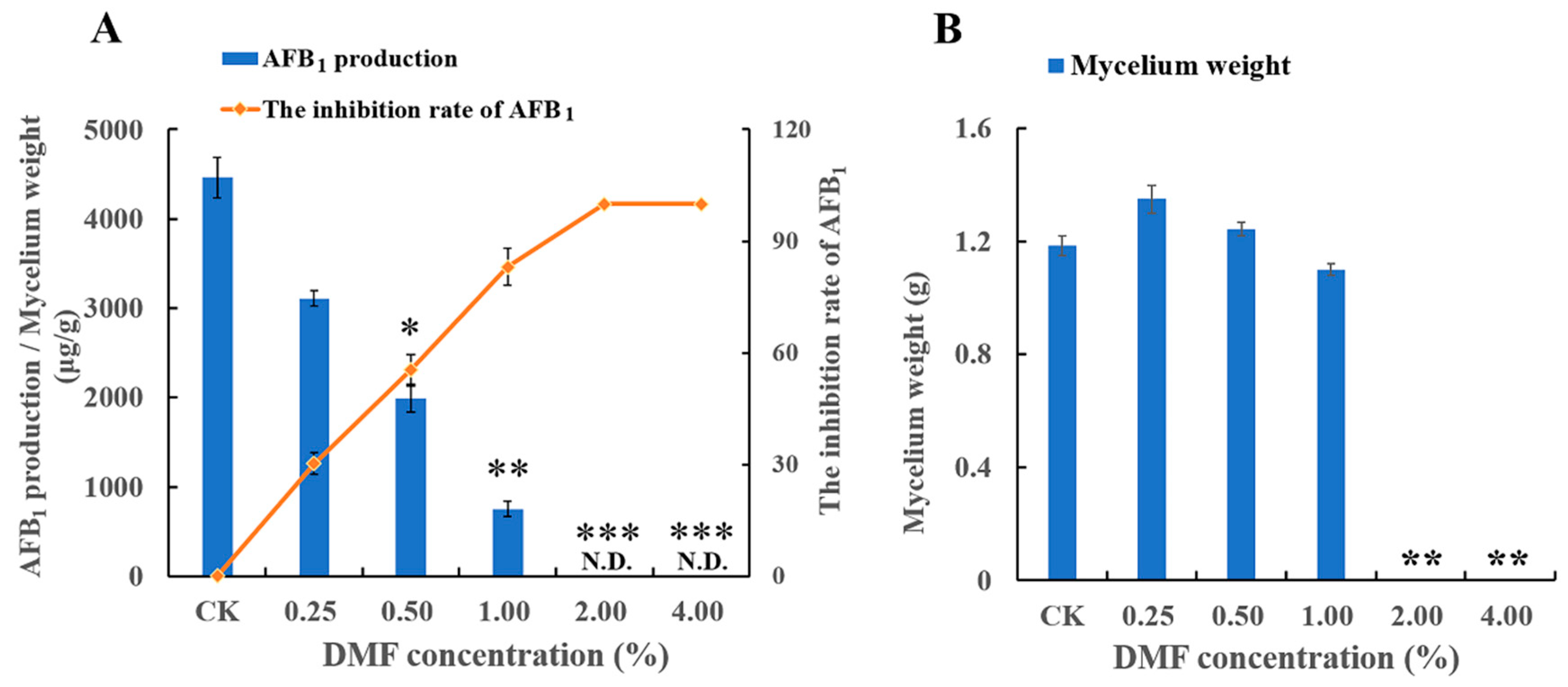
2.1. Inhibitory Potential of DMF Acts on Growth and Toxicity by A. flavus
2.2. Changes on Gene Expression Profile of A. flavus Treated with DMF
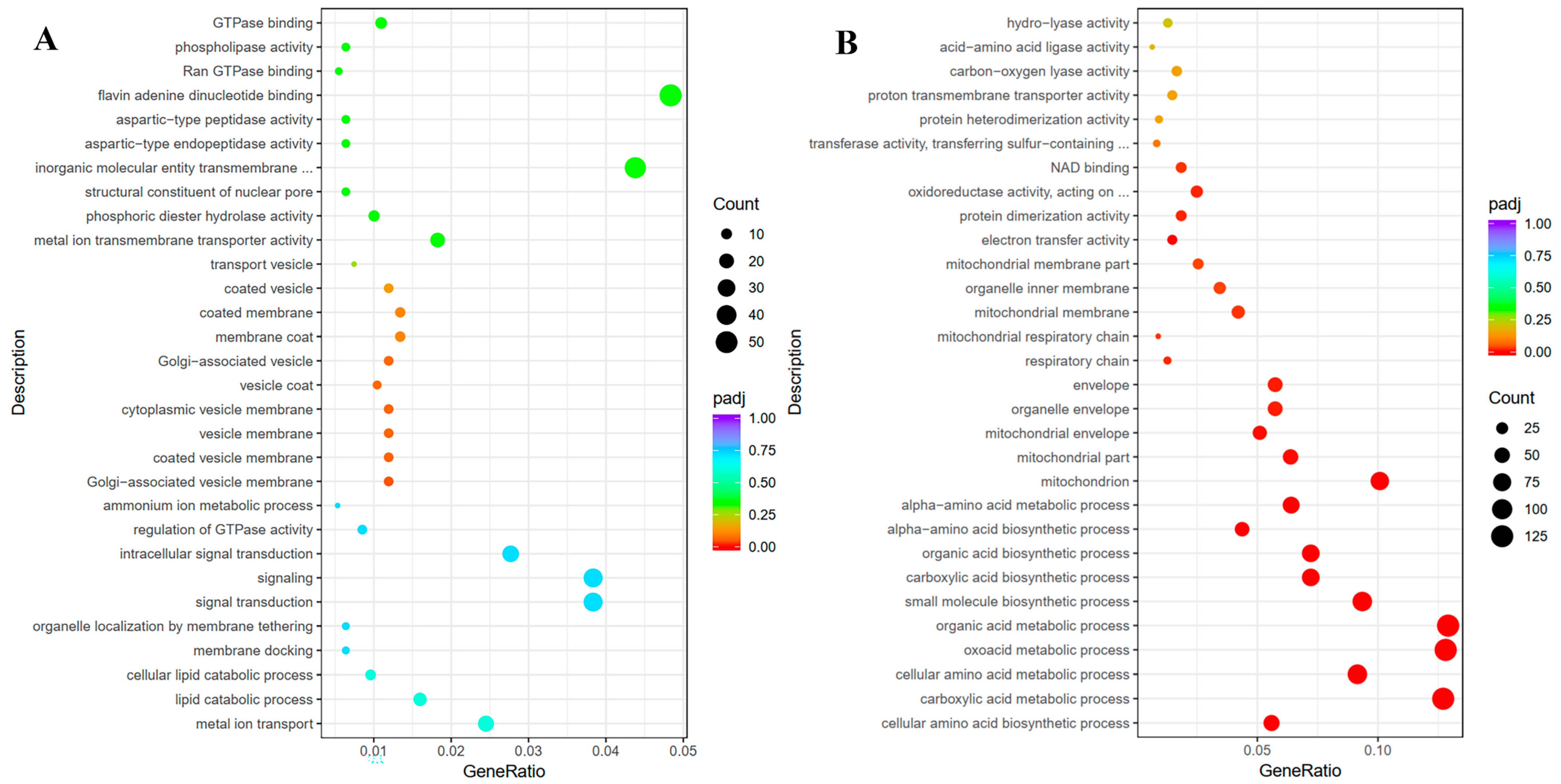
2.3. Functions and Involved Pathways of Significant DEGs
2.4. Genes Related to Pigment Biosynthesis and Fungal Development
2.5. Genes Related to the Biosynthesis of Aflatrem, Aflatoxins, and Cyclopiazonic Acid
2.6. Genes Involved in Cell Wall
2.7. Genes Involved in Glucose Metabolism Pathway
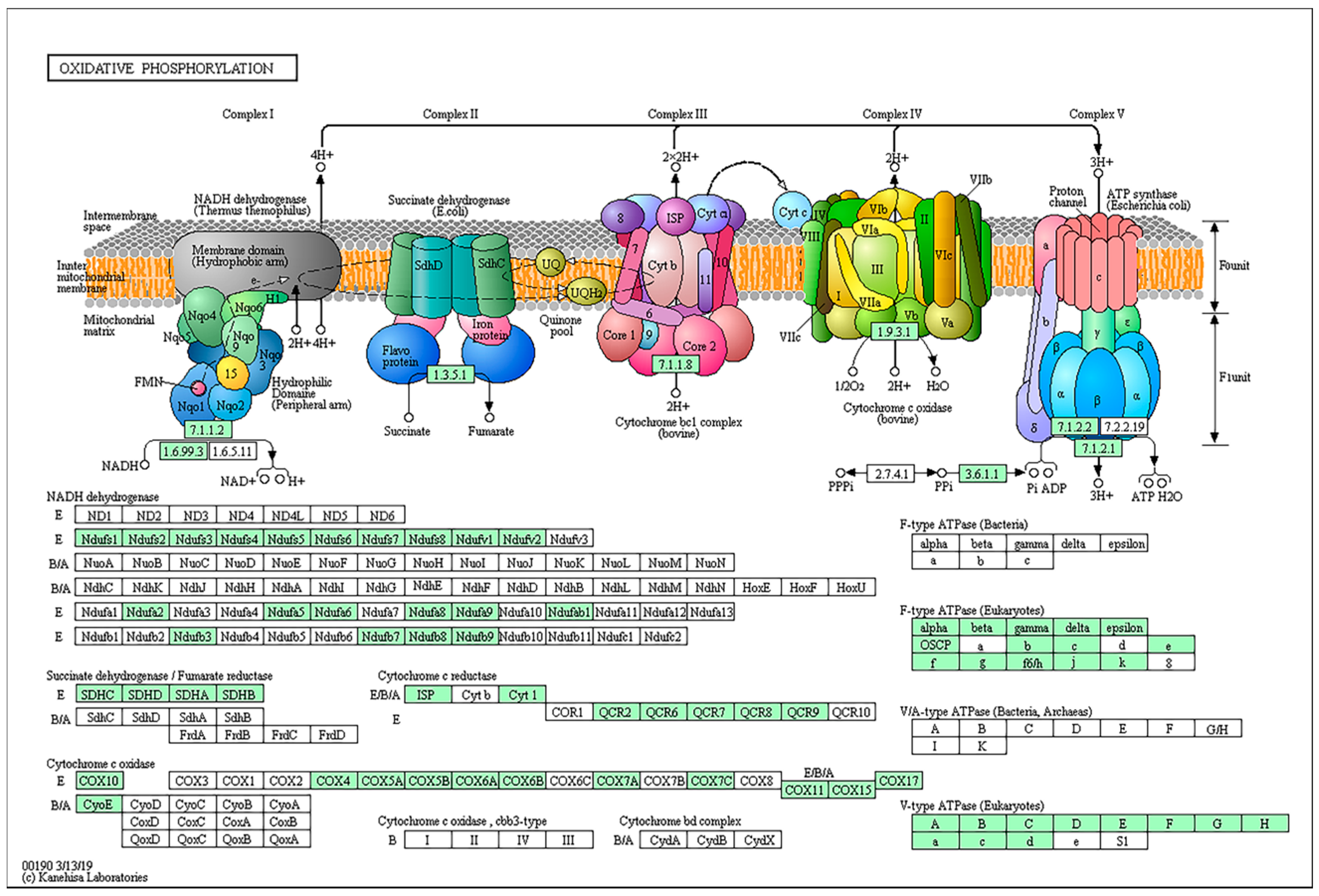
2.8. Genes Involved in Oxidative Phosphorylation and Amino Acid Biosynthesis/Metabolism
2.9. Genes Involved in MAPK Pathway, Oxylipins, GPCRs and Oxidative Stress Response
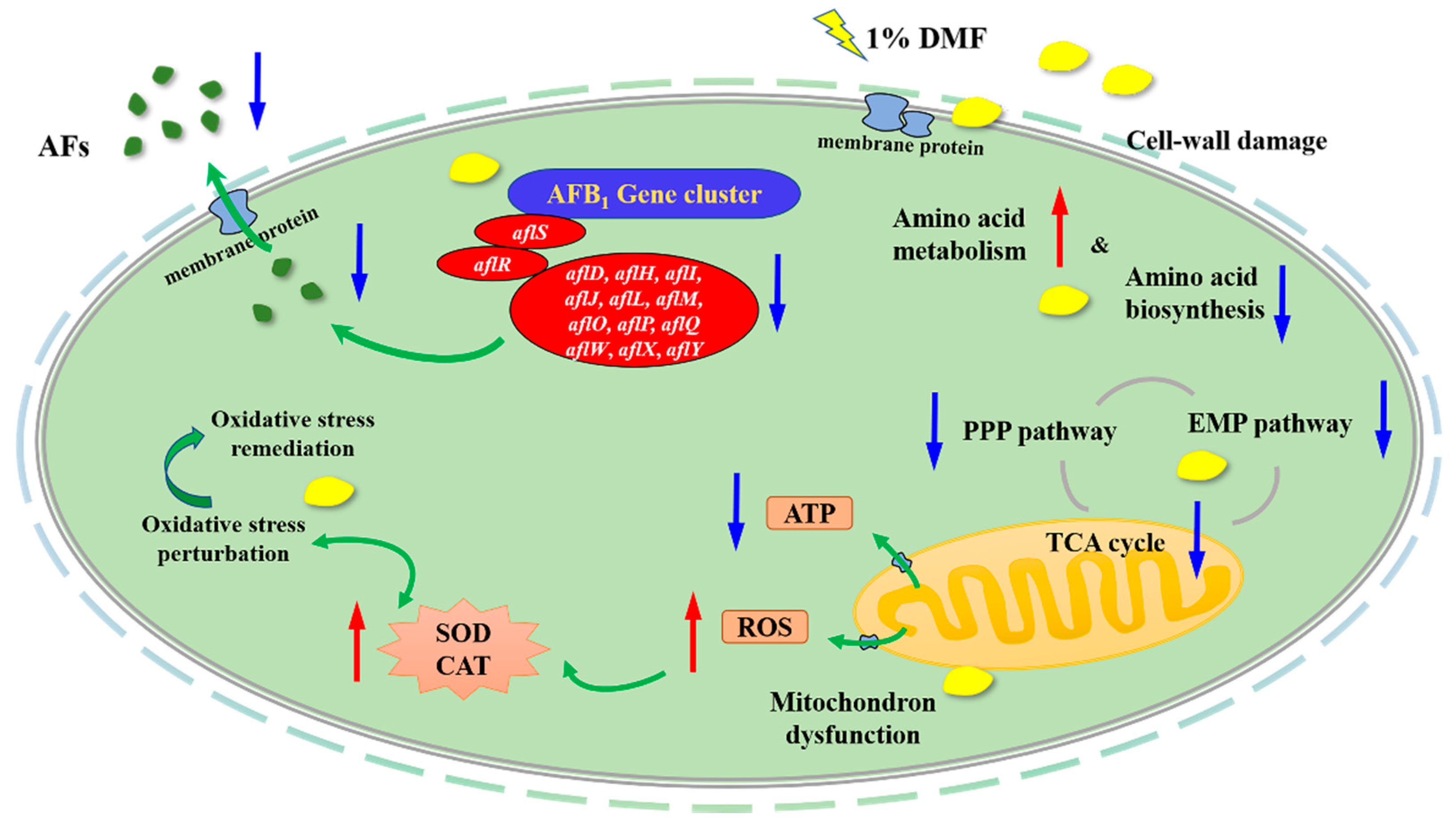
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strain, Chemicals and Treatment
5.2. Determination of Fungal Growth and AFB1 Production
5.3. Construction of cDNA Libraries and RNA Sequencing
5.4. Analysis of Sequence Data
5.5. RT-qPCR Analysis of AFs Biosynthesis Genes
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rajasekaran, K.; Sayler, R.J.; Sickler, C.M.; Majumdar, R.; Jaynes, J.; Cary, J.W. Control of Aspergillus flavus growth and aflatoxin production in transgenic maize kernels expressing a tachyplesin-derived synthetic peptide, AGM182. Plant Sci. 2018, 270, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Klich, M.A. Environmental and developmental factors influencing aflatoxin production by Aspergillus flavus and Aspergillus parasiticus. Mycoscience 2007, 48, 71–80. [Google Scholar] [CrossRef]
- Groopman, J.D.; Kensler, T.W.; Wild, C.P. Protective interventions to prevent aflatoxin-induced carcinogenesis in developing countries. Annu. Rev. Public Health 2008, 29, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, K.; Salleh, B.; Saad, B.; Abbas, H.; Abel, C.; Shier, W. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer [IARC]. Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 56, 245–395. [Google Scholar]
- International Agency for Research on Cancer [IARC]. Some traditional herbal medicines, some mycotoxins, naphthalene and styrene. IARC Monogr. Eval. Carcinog. Risks Hum. 2002, 82, 1–556. [Google Scholar]
- Luttfullah, G.; Hussain, A. Studies on contamination level of aflatoxins in some dried fruits and nuts of pakistan. Food Control. 2011, 22, 426–429. [Google Scholar] [CrossRef]
- Atayde, D.D.; Reis, T.A.; Ignácio, J.G.; Zorzete, P.; Reis, G.M.; Benedit, C. Mycobiota and aflatoxins in a peanut variety grown in different regions in the state of so paulo, brazil. Crop Prot. 2012, 33, 7–12. [Google Scholar] [CrossRef]
- The U.S. Food and Drug Administration [FDA]. CPG Sec. 570.375 Aflatoxin in Peanuts and Peanut Products; 2005. Available online: https://www.fda.gov/media/72073/download (accessed on 29 October 2020).
- The U.S. Food and Drug Administration [FDA]. CPG Sec 555.400 Foods-Adulteration with Aflatoxin; 2005. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/cpg-sec-555400-foods-adulteration-aflatoxin (accessed on 29 October 2020).
- Commission Regulation (EU) No 165/2010 of 26 February 2010 Amending Regulation (EC) No 1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs as Regards Aflatoxins (Text with EEA Relevance) OJ L 50, 27.2. 2010, pp. 8–12 (BG, ES, CS, DA, DE, ET, EL, EN, FR, IT, LV, LT, HU, MT, NL, PL, PT, RO, SK, SL, FI, SV) Special Edition in Croatian: Chapter 13 Volume 038 pp. 244–248. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32010R0165&qid=1603901359517 (accessed on 29 October 2020).
- Bhatnagar-Mathur, P.; Sunkara, S.; Bhatnagar-Panwar, M.; Waliyar, F.; Sharma, K.K. Biotechnological advances for combating Aspergillus flavus and aflatoxin contamination in crops. Plant Sci. 2015, 234, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Ceker, S.; Agar, G.; Alpsoy, L.; Nardemir, G.; Kizil, H.E. Antagonistic effects of Satureja hortensis essential oil against AFB1 on human lymphocytes in vitro. Cytol. Genet. 2014, 48, 327–332. [Google Scholar] [CrossRef]
- Abass, M.H. In vitro antifungal activity of different plant hormones on the growth and toxicity of nigrospora spp. on date palm (Phoenix Dactylifera L.). Open Plant Sci. J. 2017, 10, 10–20. [Google Scholar] [CrossRef]
- Xu, L.; Zhao, Q.W.; Luo, J.; Ma, W.L.; Jin, Y.; Li, C.H.; Hou, Y.F.; Feng, M.Y.; Wang, Y.; Chen, J.; et al. Integration of proteomics, lipidomics, and metabolomics reveals novel metabolic mechanisms underlying N, N-dimethylformamide induced hepatotoxicity. Ecotoxicol. Environ. Saf. 2020, 205, 111166. [Google Scholar] [CrossRef] [PubMed]
- Watt, S.; Schubert, F.; Wood, V.; Goodhead, I.; Penkett, C.J.; Rogers, J.; Bhler, J.; Wilhelm, B.T.; Marguerat, S. Dynamic repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution. Nature 2008, 453, 1239–1243. [Google Scholar]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Borries, A.; Vogel, J.; Sharma, C.M. Differential RNA Sequencing(dRNA-Seq): Deep-Sequencing-Based Analysis of Primary Transcriptomes. Tag-Based Next Generation Sequencing; Wiley-VCH Verlag Gmbh&Co. KGaA: Weinheim, Germany, 2012. [Google Scholar]
- Lv, C.; Wang, P.; Ma, L.; Zheng, M.; Liu, Y.; Xing, F. Large-scale comparative analysis of eugenol-induced/repressed genes expression in Aspergillus flavus using RNA-seq. Front. Microbiol. 2018, 9, 1116. [Google Scholar] [CrossRef]
- Zhao, X.; Zhi, Q.Q.; Li, J.Y.; Keller, N.P.; He, Z.M. The antioxidant gallic acid inhibits aflatoxin formation in Aspergillus flavus by modulating transcription factors FarB and CreA. Toxins 2018, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Ma, L.; Jin, J.; Zheng, M.; Pan, L.; Zhao, Y. The antiaflatoxigenic mechanism of cinnamaldehyde in Aspergillus flavus. Sci. Rep. 2019, 9, 10499. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya-Bayram, Ö.; Palmer, J.M.; Keller, N.; Braus, G.H.; Bayram, Ö. One Juliet and four Romeos: VeA and its methyltransferases. Front. Microbiol. 2015, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, R.L.; Jones, S.B.; Gerasimowicz, W.V.; Zaica, L.L.; Stahl, H.G.; Ocker, L.A. Regulation of aflatoxin biosynthesis: Assessment of the role of cellular energy status as a regulator of the induction of aflatoxin production. Appl. Environ. Microbiol. 1987, 53, 1224–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, K.C. Predicted roles of the uncharacterized clustered genes in aflatoxin biosynthesis. Toxins 2009, 1, 37–58. [Google Scholar] [CrossRef] [Green Version]
- Chang, P. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Price, M.S.; Yu, J.; Nierman, W.C.; Kim, H.; Pritchard, B.; Jacobus, C.A.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A. The aflatoxin pathway regulator AFLR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiol. Lett. 2006, 255, 275–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef]
- Yin, H.B.; Chen, C.H.; Kollanoor-Johny, A.; Darre, M.J.; Venkitanarayanan, K. Controlling Aspergillus flavus and Aspergillus parasiticus growth and aflatoxin production in poultry feed using carvacrol and trans-cinnamaldehyde. Poult. Sci. 2015, 94, 2183–2190. [Google Scholar] [CrossRef]
- Ren, Y.; Jin, J.; Zheng, M.; Yang, Q.; Xing, F. Ethanol inhibits aflatoxin B1 biosynthesis in Aspergillus flavus by up-regulating oxidative stress-related genes. Front. Microbiol. 2020, 10, 2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.Q.; Zhao, X.X.; Zhi, Q.Q.; Zhao, M.; He, Z.M. Transcriptomic profiling of Aspergillus flavus in response to 5-azacytidine. Fungal Genet. Biol. 2013, 56, 78–86. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Payne, G.A. Genetic regulation of aflatoxin biosynthesis: From gene to genome. Fungal Genet. Biol. 2009, 46, 113–125. [Google Scholar] [CrossRef]
- Davis, N.D.; Diener, U.L. Growth and aflatoxin production by Aspergillus parasiticus from various carbon sources. Appl. Microbiol. 1968, 16, 158–159. [Google Scholar] [CrossRef] [Green Version]
- Maggio-Hall, L.A.; Wilson, R.A.; Keller, N.P. Fundamental contribution of beta-oxidation to polyketiden mycotoxin production in planta. Mol. Plant Microbe Interact. 2005, 18, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Maggio-Hall, L.A.; Keller, N.P. Mitochondrial β-oxidation in Aspergillus nidulans. Mol. Microbiol. 2004, 54, 1173–1185. [Google Scholar] [CrossRef]
- Reverberi, M.; Punelli, M.; Smith, C.A.; Zjalic, S.; Scarpari, M.; Scala, V.; Cardinali, G.; Aspite, N.; Pinzari, F.; Payne, G.A.; et al. How peroxisomes affect aflatoxin biosynthesis in Aspergillus flavus. PLoS ONE 2012, 7, e48097. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Liang, Y.; Zhang, J.; Liu, C. Aspergillus flavus grown in peptone as the carbon source exhibits spore density-and peptone concentration-dependent aflatoxin biosynthesis. BMC Microbiol. 2012, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Ruijter, G.J.G.; Visser, J. Carbon repression in Aspergilli. FEMS Microbiol. Lett. 1997, 151, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Shroff, R.A.; Lockington, R.A.; Kelly, J.M. Analysis of mutations in the creA gene involved in carbon catabolite repression in Aspergillus nidulans. Can. J. Microbiol. 1996, 42, 950–959. [Google Scholar] [CrossRef]
- Wilkinson, J.R.; Yu, J.; Bland, J.M.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E. Amino acid supplementation reveals differential regulation of aflatoxin biosynthesis in Aspergillus flavus NRRL 3357 and Aspergillus parasiticus SRRC 143. Appl. Microbiol. Biotechnol. 2007, 74, 1308–1319. [Google Scholar] [CrossRef]
- Adye, J.C.; Mateles, R.I. Incorporation of labeled compound into aflatoxin. Biochim. Et Biophys. Acta 1964, 86, 418–420. [Google Scholar] [CrossRef]
- Naik, M.; Modi, V.V.; Patel, N.C. Studies on aflatoxin synthesis in Aspergillus flavus. Indian J. Exp. Biol. 1970, 8, 345–346. [Google Scholar] [PubMed]
- Payne, G.A.; Hagler, W.M. Effect of specific amino acids on growth and aflatoxin production by Aspergillus parasiticus and Aspergillus flavus in defined media. Appl. Environ. Microbiol. 1983, 46, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Roze, L.V.; Chanda, A.; Laivenieks, M.; Beaudry, R.M.; Artymovich, K.A.; Koptina, A.V.; Awad, D.W.; Valeeva, D.; Jones, A.D.; Linz, J.E. Volatile profiling reveals intracellular metabolic changes in Aspergillus parasiticus: veA regulates branched chain amino acid and ethanol metabolism. BMC Biochem. 2010, 11, 33. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Hu, S.S.; Sarreal, S.B.; Li, R.W. Suppression of aflatoxin biosynthesis in Aspergillus flavus by 2-phenylethanol is associated with stimulated growth and decreased degradation of branched-chain amino acids. Toxins 2015, 7, 3887–3902. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Lu, J.; Zhang, W.; Suzukizuki, I.; Li, G. Enhanced electron-transfer reactivity of cytochrome b5 by Dimethylsulfoxide and N, N′-Dimethylformamide. Anal. Sci. 2002, 18, 1031–1033. [Google Scholar]
- Pe~Na-Valdivia, C.B.; Rodriguez-Flores, L.; Gómez-Puyou, M.T.; Lotina-Hennsen, B. Inhibition of photophosphorylation and electron transport by N, N-dimethylformamide. Biophys. Chem. 1991, 41, 169–174. [Google Scholar] [CrossRef]
- Madrigal-Perez, L.A.; Ramos-Gomez, M. Resveratrol inhibition of cellular respiration: New paradigm for an old mechanism. Int. J. Mol. Sci. 2016, 17, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayashree, T.; Subramanyam, C. Oxidative stress as a prerequisite for aflatoxin production by Aspergillus parasiticus. Free. Radic. Biol. Med. 2000, 29, 981–985. [Google Scholar] [CrossRef]
- Lee, M.J.; Sheppard, D.C. Recent advances in the understanding of the Aspergillus fumigatus cell wall. J. Microbiol. 2016, 54, 232–242. [Google Scholar] [CrossRef]
- Dichtl, K.; Helmschrott, C.; Dirr, F.; Wagener, J. Deciphering cell wall integrity signalling in Aspergillus fumigatus: Identification and functional characterization of cell wall stress sensors and relevant Rho GTPases. Mol. Microbiol. 2012, 83, 506–519. [Google Scholar] [CrossRef]
- Henry, C.; Latgé, J.P.; Beauvais, A. α-1, 3-glucans are dispensable in Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 26–29. [Google Scholar] [CrossRef] [Green Version]
- Meetei, P.A.; Rathore, R.S.; Prabhu, N.P.; Vindal, V. In silico screening for identification of novel β-1, 3-glucansynthase inhibitors using pharmacophore and 3D-QSAR methodologies. Springerplus 2016, 5, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henar, M.V.; Durán, A.; Roncero, C. Chitin synthases in yeast and fungi. EXS 1999, 87, 55–69. [Google Scholar]
- Yang, M.; Lu, L.; Li, S.; Zhang, J.; Li, Z.; Wu, S.; Guo, Q.; Liu, H.; Wu, C. Transcriptomic insights into benzenamine effects on the development, aflatoxin biosynthesis, and virulence of Aspergillus flavus. Toxins 2019, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Feng, K.; Yang, H.; Zhang, Z.; Yuan, Y.; Yue, T. Effect of cinnamaldehyde and citral combination on transcriptional profile, growth, oxidative damage and patulin biosynthesis of Penicillium expansum. Front. Microbiol. 2018, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Yamazaki, D.; Takaya, N.; Takagi, M.; Ohta, A.; Horiuchi, H. A chitinase gene, chiB, involved in the autolytic process of Aspergillus nidulans. Curr. Genet. 2007, 51, 89. [Google Scholar] [CrossRef]
- Liang, D.; Xing, F.; Selvaraj, J.N.; Liu, X.; Wang, L.; Hua, H.; Zhou, L.; Zhao, Y.; Wang, Y.; Liu, Y. Inhibitory effect of cinnamaldehyde, citral and eugenol on aflatoxin biosynthetic gene expression and aflatoxin B1 biosynthesis in Aspergillus flavus. J. Food Sci. 2015, 80, M2917–M2924. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Priebe, S.; Linde, J.; Albrecht, D.; Guthke, R.; Brakhage, A.A. FungiFun: A web-based application for functional categorization of fungal genes and proteins. Fungal Genet. Biol. 2011, 48, 353–358. [Google Scholar] [CrossRef]
- Li, X.; Jiang, Y.; Ma, L.; Ma, X.; Liu, Y.; Shan, J.; Ma, K.; Xing, F. Comprehensive transcriptome and proteome analyses reveal the modulation of aflatoxin production by Aspergillus flavus on different crop substrates. Front. Microbiol. 2020, 11, 1497. [Google Scholar] [CrossRef]
- Lu, L.X.; Zhou, F.; Zhou, Y.; Fan, X.L.; Ye, S.F.; Wang, L.; Chen, H.; Lin, Y.J. Expression profile analysis of the polygalacturonase-inhibiting protein genes in rice and their responses to phytohormones and fungal infection. Plant Cell Rep. 2012, 31, 1173–1187. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | CK * (FPKM) | D1 * (FPKM) | Log2 D1/CK | Annotated Gene Function |
|---|---|---|---|---|
| AFLA_016120 | 4.06 | 44.50 | 3.45 | O-methyltransferase family protein |
| AFLA_016130 | 4.70 | 45.64 | 3.28 | hypothetical protein |
| AFLA_016140 | 2.25 | 16.60 | 2.88 | Arp1 conidial pigment biosynthesis scytalone dehydratase |
| AFLA_006180 | 0.84 | 1.08 | 0.34 | Arb2/brown2 conidial pigment biosynthesis oxidase |
| AFLA_009340 | 2.11 | 4.24 | 1.01 | Mod-A developmental regulator, putative |
| AFLA_014260 | 0.62 | 1.99 | 1.67 | RodB/HypB conidial hydrophobin |
| AFLA_098380 | 0.11 | 0.13 | 0.28 | RodA/RolA conidial hydrophobin |
| AFLA_039530 | 4.14 | 19.88 | 2.26 | FluG |
| AFLA_044790 | 154.53 | 520.23 | 1.75 | conidiation-specific family protein |
| AFLA_044800 | 18.58 | 64.49 | 1.80 | conidiation protein Con-6, putative |
| AFLA_046990 | 166.39 | 188.53 | 0.18 | StuA APSES transcription factor |
| AFLA_018340 | 147.87 | 136.85 | −0.11 | GpaA/FadA G-protein complex alpha subunit |
| AFLA_081490 | 36.64 | 23.97 | −0.61 | Gda1/VelB nucleoside diphosphatase |
| AFLA_021090 | 2.06 | 4.23 | 1.04 | sporulation associated protein |
| AFLA_024890 | 25.68 | 32.99 | 0.36 | Fsr1/Pro1 cell differentiation and development protein |
| AFLA_029620 | 2.75 | 4.35 | 0.66 | AbaA transcription factor |
| AFLA_026900 | 17.70 | 32.20 | 0.86 | VosA developmental regulator |
| AFLA_066460 | 135.21 | 112.55 | −0.26 | VeA developmental regulator |
| AFLA_033290 | 40.16 | 27.03 | −0.57 | LaeA regulator of secondary metabolism |
| AFLA_134030 | 25.77 | 16.89 | −0.61 | developmental regulator FlbA |
| AFLA_136410 | 158.96 | 139.49 | −0.19 | transcriptional regulator Medusa |
| AFLA_137320 | 122.34 | 73.75 | −0.73 | C2H2 conidiation transcription factor FlbC |
| AFLA_052030 | 11.14 | 17.11 | 0.62 | WetA developmental regulatory protein |
| AFLA_071090 | 291.36 | 406.28 | 0.48 | EsdC GTP-binding protein |
| AFLA_079710 | 54.76 | 57.87 | 0.08 | HymA conidiophore development protein |
| AFLA_080170 | 5.12 | 7.75 | 0.60 | FlbD MYB family conidiophore development protein, putative |
| AFLA_082850 | 1.68 | 2.20 | 0.39 | BrlA C2H2 type conidiation transcription factor |
| AFLA_083110 | 34.55 | 50.12 | 0.54 | conidiation-specific protein (Con-10), putative |
| AFLA_101920 | 8.41 | 14.18 | −0.75 | FluG extracellular developmental signal biosynthesis protein |
| AFLA_131490 | 62.27 | 62.35 | 0.00 | conserved hypothetical protein |
| Cluster ID | Gene ID | CK * (FPKM) | D1 * (FPKM) | Log2 D1/CK | Annotated Gene Function |
|---|---|---|---|---|---|
| 15 | AFLA_045460 | 2.99 | 0.90 | −1.73 | MFS multidrug transporter, putative |
| 15 | AFLA_045470 | 0 | 0 | NA | FAD dependent oxidoreductase, putative |
| 15 | AFLA_045480 | 0.06 | 0.07 | 0.20 | dimethylallyl tryptophan synthase, putative |
| 15 | AFLA_045490 | 0 | 0.03 | Up | hybrid PKS/NRPS enzyme, putative |
| 15 | AFLA_045500 | 0.04 | 0.07 | 0.85 | cytochrome P450, putative |
| 15 | AFLA_045510 | 0 | 0.02 | Up | integral membrane protein |
| 15 | AFLA_045520 | 0 | 0.07 | Up | integral membrane protein |
| 15 | AFLA_045530 | 0 | 0 | NA | hypothetical protein |
| 15 | AFLA_045540 | 0.04 | 0.06 | 0.76 | cytochrome P450, putative |
| 15 | AFLA_045550 | 3.64 | 6.43 | 0.82 | hypothetical protein |
| 15 | AFLA_045560 | 4.57 | 6.63 | 0.54 | carboxylic acid transport protein |
| 15 | AFLA_045570 | 2.23 | 0.62 | −1.85 | acetyl xylan esterase, putative |
| 54 | AFLA_139100 | 6.51 | 7.62 | 0.23 | aflYe/orf/Ser -Thr protein phosphatase family protein |
| 54 | AFLA_139110 | 5.18 | 7.60 | 0.55 | aflYd/sugR/sugar regulator |
| 54 | AFLA_139120 | 3.94 | 7.22 | 0.87 | aflYc/glcA/glucosidase |
| 54 | AFLA_139130 | 4.60 | 5.98 | 0.38 | aflYb/hxtA/putative hexose transporter |
| 54 | AFLA_139140 | 2.68 | 1.05 | −1.34 | aflYa/nadA/NADH oxidase |
| 54 | AFLA_139150 | 9.79 | 2.77 | −1.82 | aflY/hypA/hypP/hypothetical protein |
| 54 | AFLA_139160 | 10.32 | 4.95 | −1.06 | aflX/ordB/monooxygenase/oxidase |
| 54 | AFLA_139170 | 15.59 | 5.08 | −1.62 | aflW/moxY/monooxygenase |
| 54 | AFLA_139180 | 11.52 | 5.79 | −0.99 | aflV/cypX/cytochrome P450 monooxygenase |
| 54 | AFLA_139190 | 10.62 | 5.74 | −0.89 | aflK/vbs/VERB synthase |
| 54 | AFLA_139200 | 3.91 | 1.43 | −1.45 | aflQ/ordA/ord-1/oxidoreductase/cytochrome P450 monooxigenase |
| 54 | AFLA_139210 | 16.72 | 5.02 | −1.73 | aflP/omtA/omt-1/O-methyltransferase A |
| 54 | AFLA_139220 | 27.50 | 8.70 | −1.66 | aflO/omtB/dmtA/O-methyltransferase B |
| 54 | AFLA_139230 | 1.47 | 0.41 | −1.81 | aflI/avfA/cytochrome P450 monooxygenase |
| 54 | AFLA_139240 | 6.38 | 3.60 | −0.82 | aflLa/hypB/hypothetical protein |
| 54 | AFLA_139250 | 10.04 | 4.37 | −1.19 | aflL/verB/desaturase/P450 monooxygenase |
| 54 | AFLA_139260 | 6.54 | 3.59 | −0.86 | aflG/avnA/ord-1/cytochrome P450 monooxygenase |
| 54 | AFLA_139270 | 176.54 | 181.24 | 0.03 | aflNa/hypD/hypothetical protein |
| 54 | AFLA_139280 | 4.94 | 4.62 | −0.09 | aflN/verA/monooxygenase |
| 54 | AFLA_139290 | 13.89 | 7.10 | −0.96 | aflMa/hypE/hypothetical protein |
| 54 | AFLA_139300 | 55.63 | 17.29 | −1.68 | aflM/ver-1/dehydrogenase/ketoreductase |
| 54 | AFLA_139310 | 15.86 | 8.02 | −0.98 | aflE/norA/aad/adh-2/NOR reductase/dehydrogenase |
| 54 | AFLA_139320 | 33.02 | 13.07 | −1.33 | aflJ/estA/esterase |
| 54 | AFLA_139330 | 28.77 | 13.33 | −1.10 | aflH/adhA/short chain alcohol dehydrogenase |
| 54 | AFLA_139340 | 108.53 | 90.40 | −0.26 | aflS/pathway regulator |
| 54 | AFLA_139360 | 76.49 | 57.18 | −0.41 | aflR/apa-2/afl-2/transcription activator |
| 54 | AFLA_139370 | 8.47 | 7.73 | −0.13 | aflB/fas-1/fatty acid synthase beta subunit |
| 54 | AFLA_139380 | 6.89 | 7.99 | 0.21 | aflA/fas-2/hexA/fatty acid synthase alpha subunit |
| 54 | AFLA_139390 | 39.85 | 19.86 | −1.00 | aflD/nor-1/reductase |
| 54 | AFLA_139400 | 13.55 | 8.20 | −0.72 | aflCa/hypC/hypothetical protein |
| 54 | AFLA_139410 | 10.41 | 6.66 | −0.64 | aflC/pksA/pksL1/polyketide synthase |
| 54 | AFLA_139420 | 130.55 | 123.18 | −0.08 | aflT/aflT/transmembrane protein |
| 54 | AFLA_139430 | 21.56 | 16.39 | −0.39 | aflU/cypA/P450 monooxygenase |
| 54 | AFLA_139440 | 22.49 | 16.67 | −0.43 | aflF/norB/dehydrogenase |
| 55 | AFLA_139460 | 485.20 | 216.59 | −1.16 | MFS multidrug transporter, putative |
| 55 | AFLA_139470 | 64.20 | 249.28 | 1.95 | FAD dependent oxidoreductase, putative |
| 55 | AFLA_139480 | 155.11 | 556.66 | 1.84 | dimethylallyl tryptophan synthase, putative |
| 55 | AFLA_139490 | 0.95 | 6.29 | 2.72 | hybrid PKS/NRPS enzyme, putative |
| Gene ID | CK * (FPKM) | D1 * (FPKM) | Log2 D1/CK | Annotated Gene Function |
|---|---|---|---|---|
| AFLA_038420 | 0.02 | 0.59 | 4.68 | endo-chitosanase B |
| AFLA_024770 | 0.89 | 4.22 | 2.25 | symbiotic chitinase, putative |
| AFLA_023460 | 5.01 | 17.45 | 1.80 | alpha-1,3-glucan synthase Ags1 |
| AFLA_134100 | 0.05 | 0.09 | 0.76 | alpha-1,3-glucan synthase Ags2 |
| AFLA_052800 | 293.07 | 314.70 | 0.10 | 1,3-beta-glucan synthase catalytic subunit FksP |
| AFLA_041060 | 0.05 | 0.01 | −2.88 | cell wall associated protein, putative |
| AFLA_104680 | 0.05 | 0.01 | −2.27 | class V chitinase ChiB1 |
| AFLA_013280 | 0.75 | 0.22 | −1.76 | class V chitinase, putative |
| AFLA_031380 | 88.88 | 34.17 | −1.38 | class V chitinase, putative |
| AFLA_054470 | 0.31 | 0.25 | −0.30 | class V chitinase Chi100 |
| AFLA_114760 | 24.13 | 10.06 | −1.26 | chitin synthase B |
| AFLA_086070 | 0.02 | 0.01 | −0.94 | chitin synthase, putative |
| AFLA_067530 | 43.44 | 29.37 | −0.56 | chitin biosynthesis protein (Chs7), putative |
| AFLA_137200 | 1.12 | 0.81 | −0.47 | chitin synthase, putative |
| AFLA_013690 | 78.45 | 58.64 | −0.42 | chitin synthase C |
| AFLA_091300 | 62.44 | 50.43 | −0.31 | chitin biosynthesis protein (Chs5), putative |
| AFLA_052780 | 5.03 | 3.14 | −0.68 | cell wall glucanase (Scw4), putative |
| AFLA_096680 | 1.53 | 0.41 | −1.89 | glucan endo-1,3-alpha-glucosidase agn1 precursor, putative |
| AFLA_095680 | 1.08 | 0.34 | −1.67 | alpha-1,3-glucanase, putative |
| AFLA_029950 | 6.84 | 2.55 | −1.43 | endo-1,3(4)-beta-glucanase, putative |
| AFLA_045290 | 570.20 | 213.72 | −1.42 | extracellular endoglucanase/cellulase, putative |
| AFLA_102640 | 1.53 | 0.60 | −1.36 | exo-beta-1,3-glucanase, putative |
| AFLA_053390 | 1894.71 | 823.30 | −1.20 | GPI-anchored cell wall beta-1,3-endoglucanase EglC |
| AFLA_068300 | 4467.43 | 2012.10 | −1.15 | 1,3-beta-glucanosyltransferase Bgt1 |
| AFLA_129440 | 704.92 | 365.98 | −0.95 | 1,3-beta-glucanosyltransferase, putative |
| AFLA_034920 | 4.00 | 2.11 | −0.92 | glucan endo-1,3-alpha-glucosidase agn1 precursor, putative |
| AFLA_058480 | 4287.31 | 2388.65 | −0.84 | 1,3-beta-glucanosyltransferase Gel1 |
| AFLA_087870 | 34.41 | 20.02 | −0.78 | Endoglucanase, putative |
| AFLA_111970 | 3.86 | 2.26 | −0.77 | Endoglucanase, putative |
| AFLA_126410 | 4.24 | 2.61 | −0.70 | endoglucanase-1 precursor, putative |
| AFLA_052780 | 5.03 | 3.14 | −0.68 | cell wall glucanase (Scw4), putative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, L.; Chang, P.; Jin, J.; Yang, Q.; Xing, F. Dimethylformamide Inhibits Fungal Growth and Aflatoxin B1 Biosynthesis in Aspergillus flavus by Down-Regulating Glucose Metabolism and Amino Acid Biosynthesis. Toxins 2020, 12, 683. https://doi.org/10.3390/toxins12110683
Pan L, Chang P, Jin J, Yang Q, Xing F. Dimethylformamide Inhibits Fungal Growth and Aflatoxin B1 Biosynthesis in Aspergillus flavus by Down-Regulating Glucose Metabolism and Amino Acid Biosynthesis. Toxins. 2020; 12(11):683. https://doi.org/10.3390/toxins12110683
Chicago/Turabian StylePan, Lin, Peng Chang, Jing Jin, Qingli Yang, and Fuguo Xing. 2020. "Dimethylformamide Inhibits Fungal Growth and Aflatoxin B1 Biosynthesis in Aspergillus flavus by Down-Regulating Glucose Metabolism and Amino Acid Biosynthesis" Toxins 12, no. 11: 683. https://doi.org/10.3390/toxins12110683
APA StylePan, L., Chang, P., Jin, J., Yang, Q., & Xing, F. (2020). Dimethylformamide Inhibits Fungal Growth and Aflatoxin B1 Biosynthesis in Aspergillus flavus by Down-Regulating Glucose Metabolism and Amino Acid Biosynthesis. Toxins, 12(11), 683. https://doi.org/10.3390/toxins12110683