Effects of Single and Repeated Oral Doses of Ochratoxin A on the Lipid Peroxidation and Antioxidant Defense Systems in Mouse Kidneys


 and
and
Abstract
:1. Introduction
2. Results
2.1. Water and Food Consumption and Body Weight Change in Mice
2.2. Blood Plasma OTA Content in a Single Oral Dose (24 h), Repeated Daily Oral Dose (72 h), and Repeated Daily Oral Dose (21 Days) Treatments
2.3. Effect of OTA on the Weight of Spleen and Kidney
2.4. Histopathological Analysis of the Renal Cortex
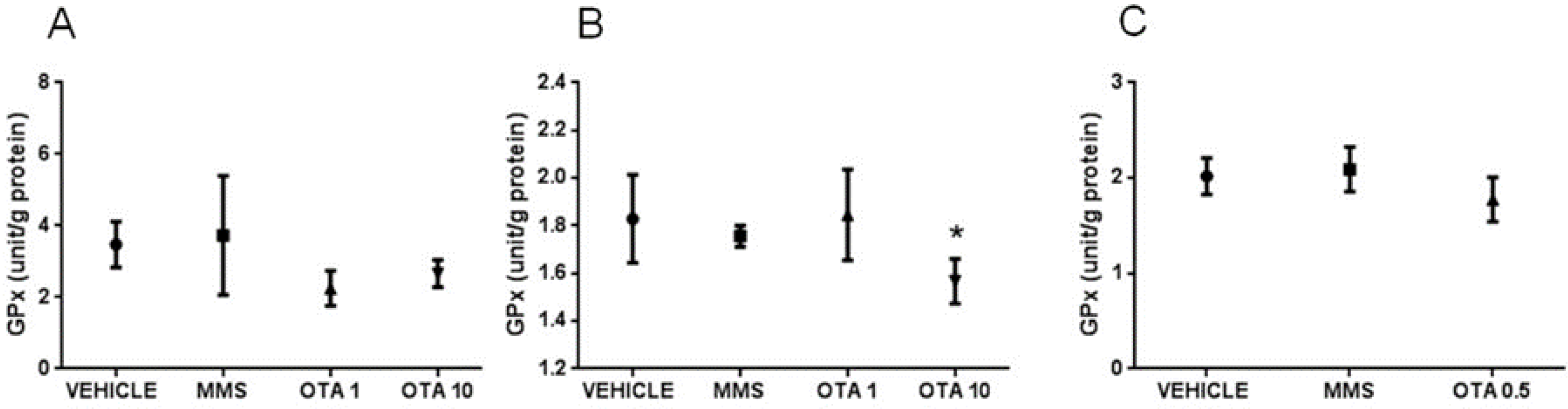
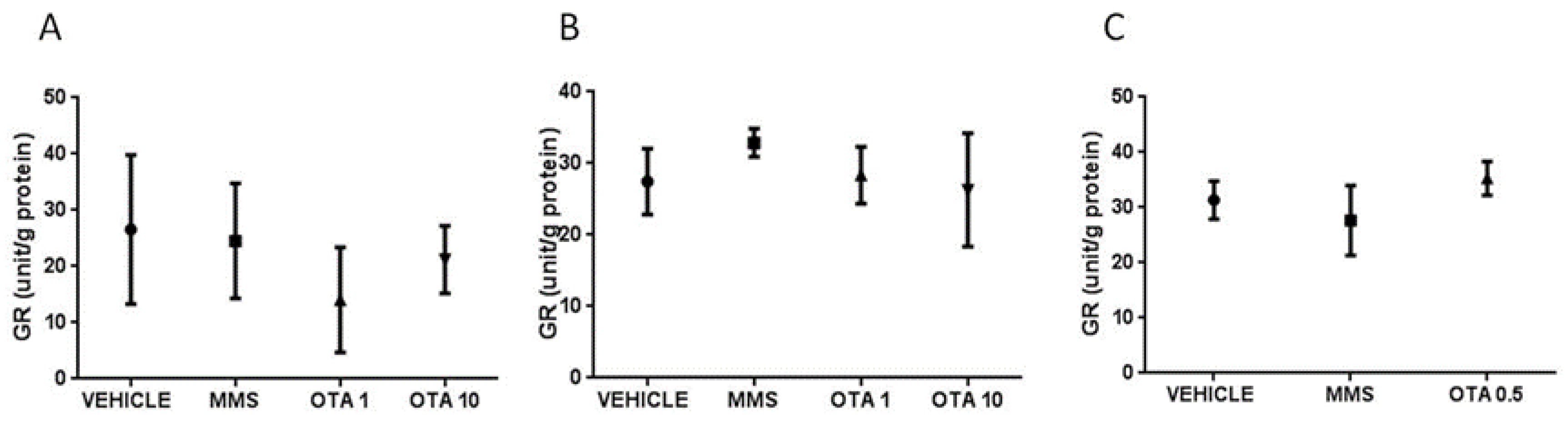
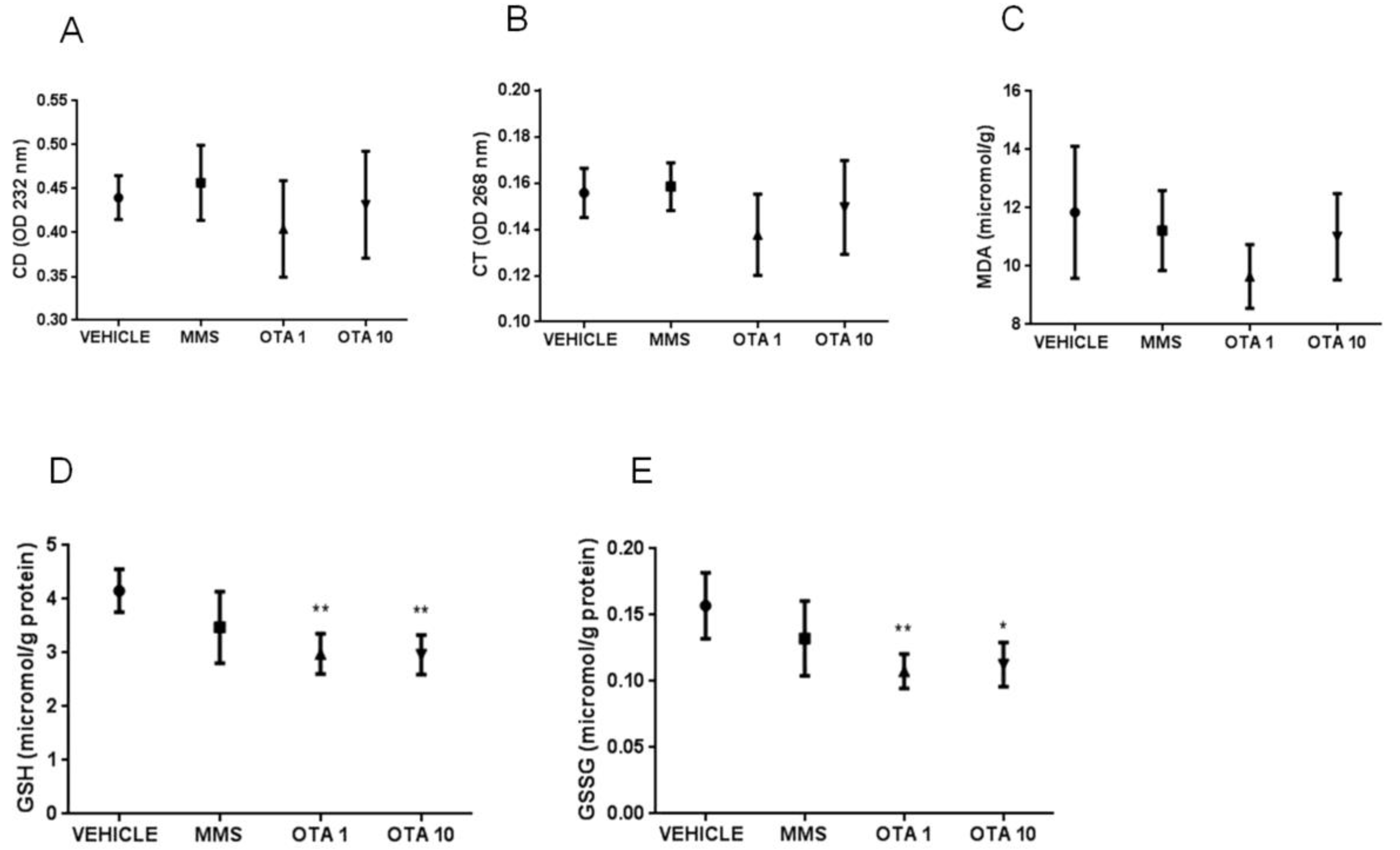
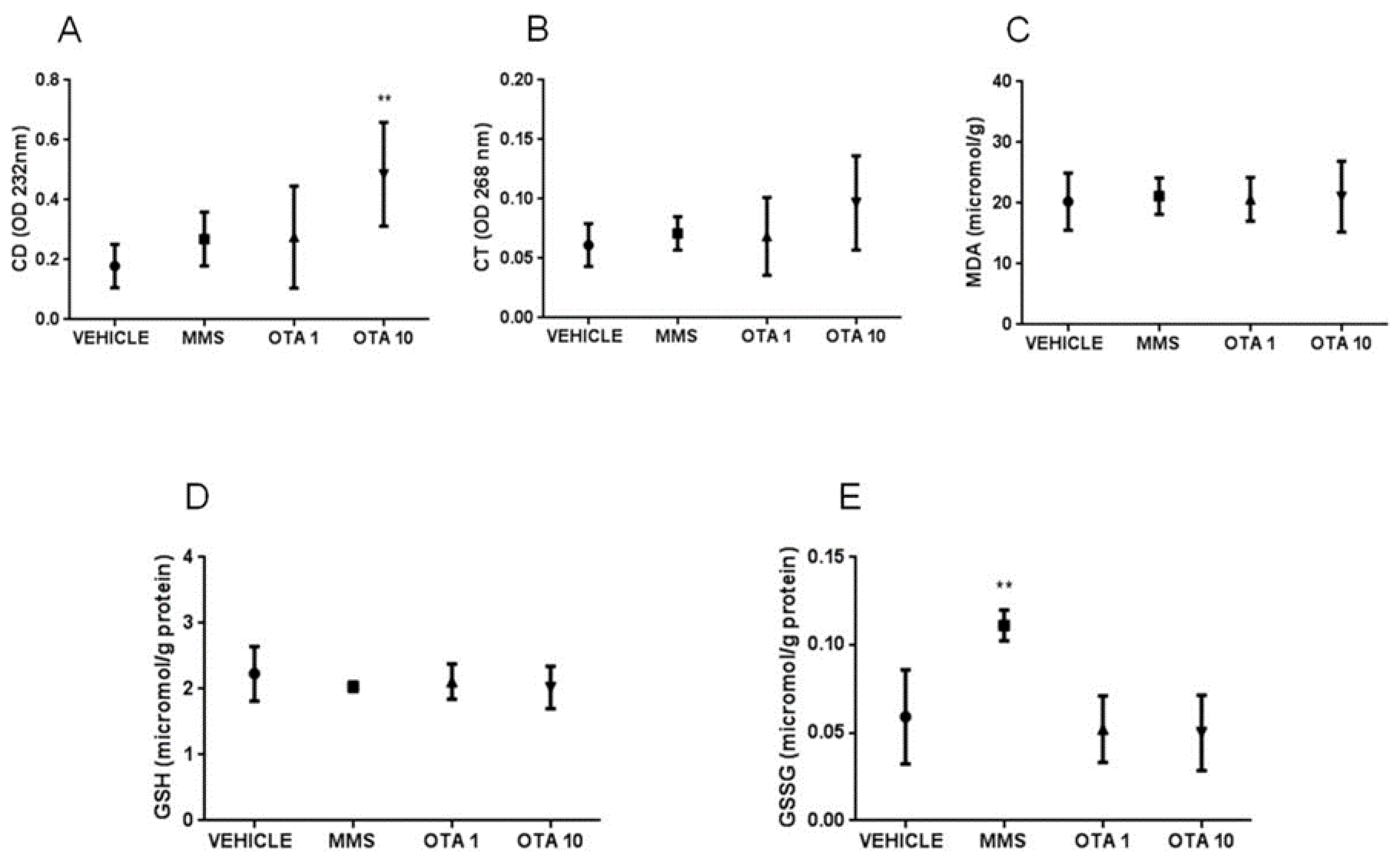
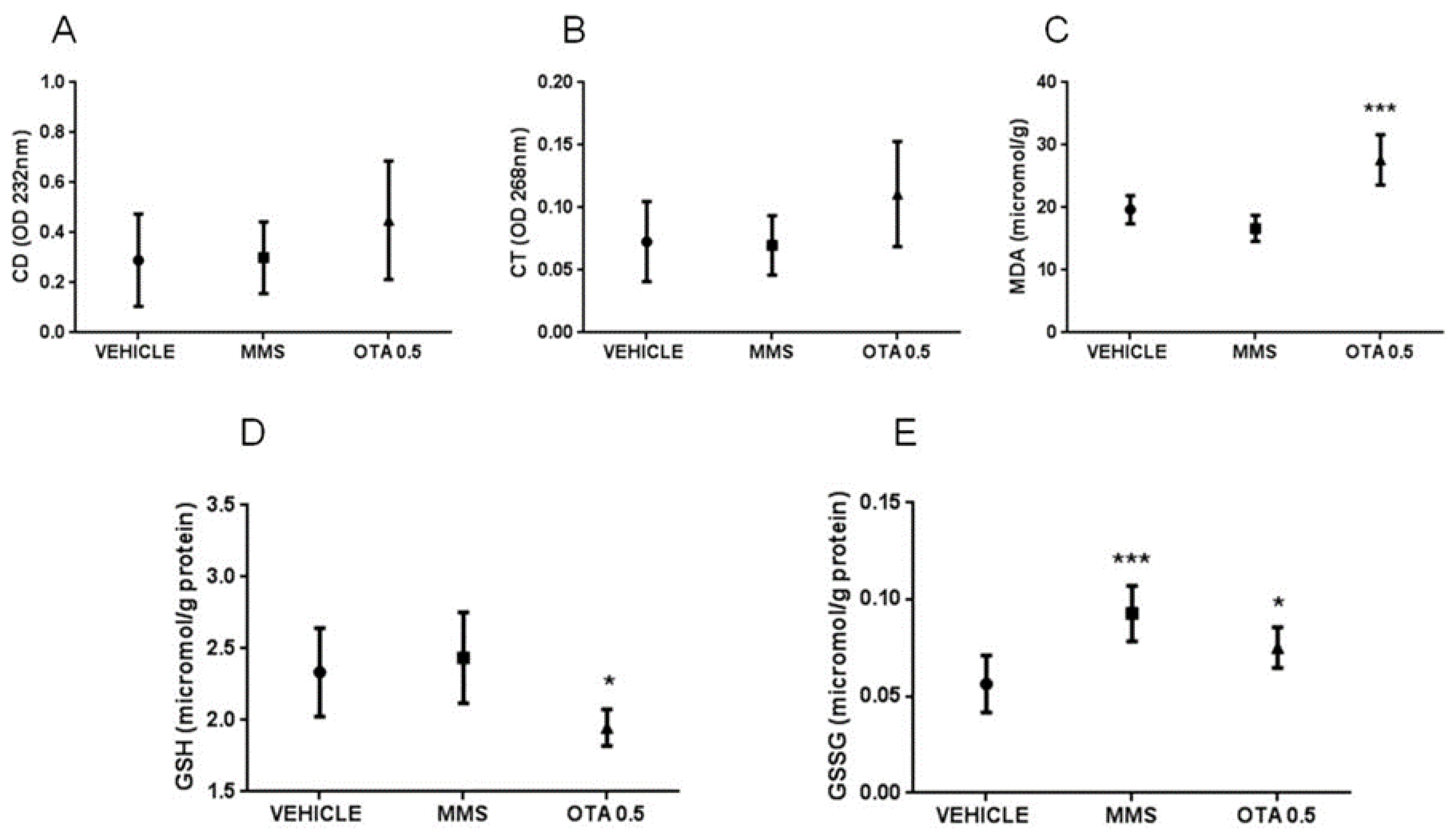
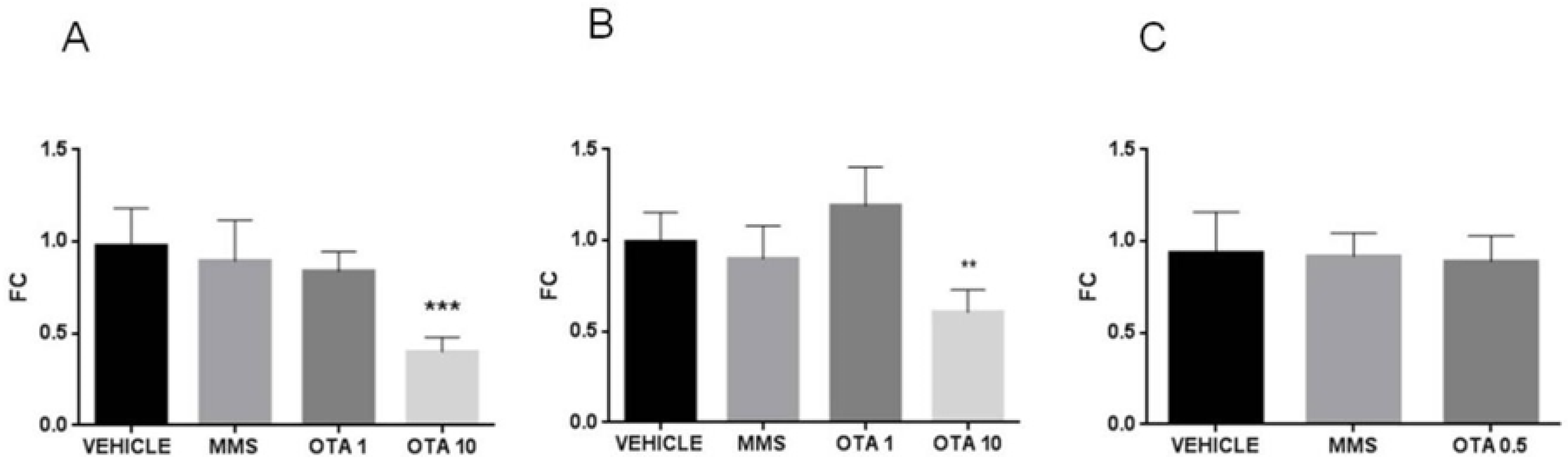
2.5. Changes of Some Parameters of the Glutathione Redox System and Lipid Peroxidation
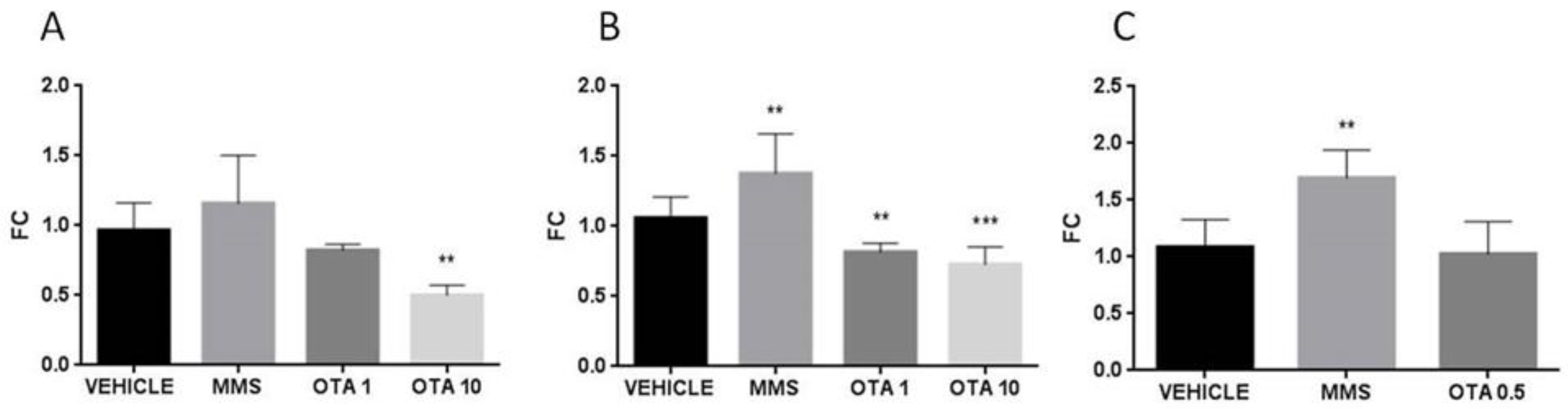
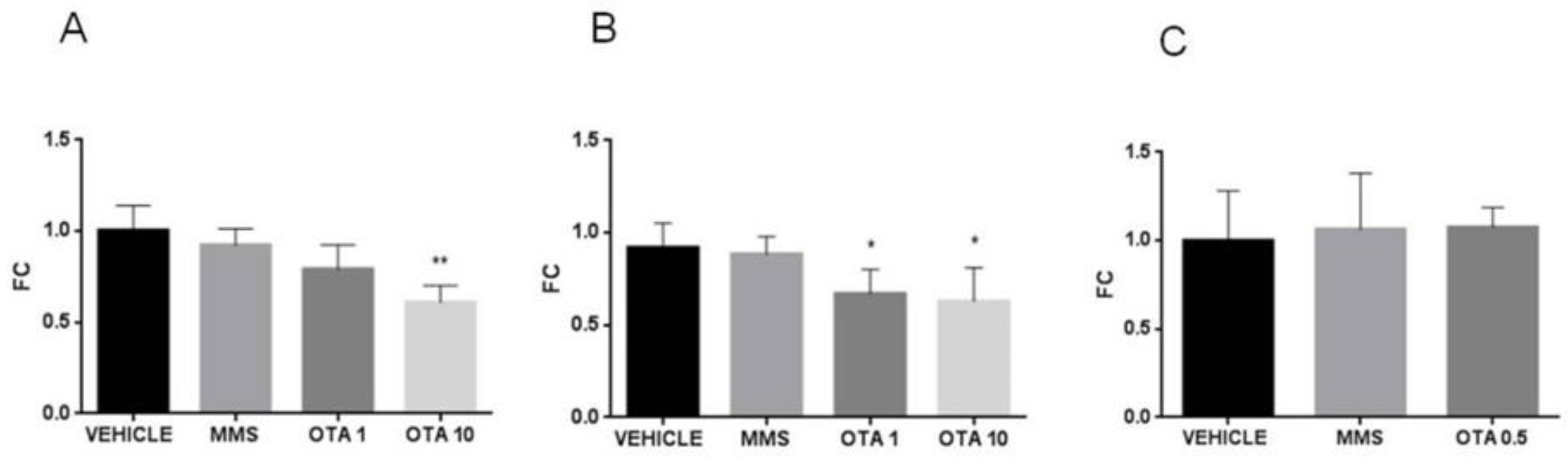
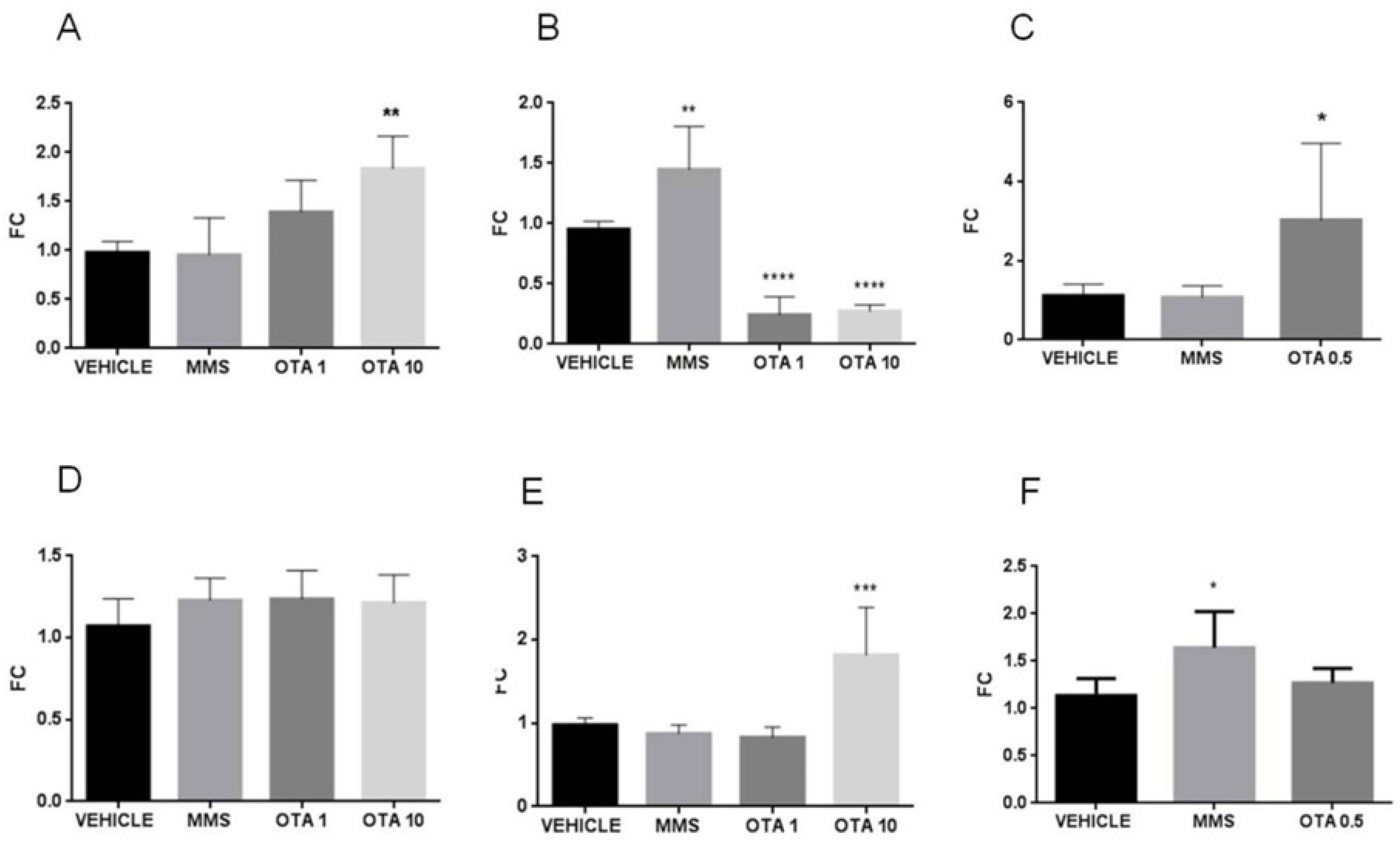
2.6. Gene Expression of Some Parameters of the Glutathione Redox System
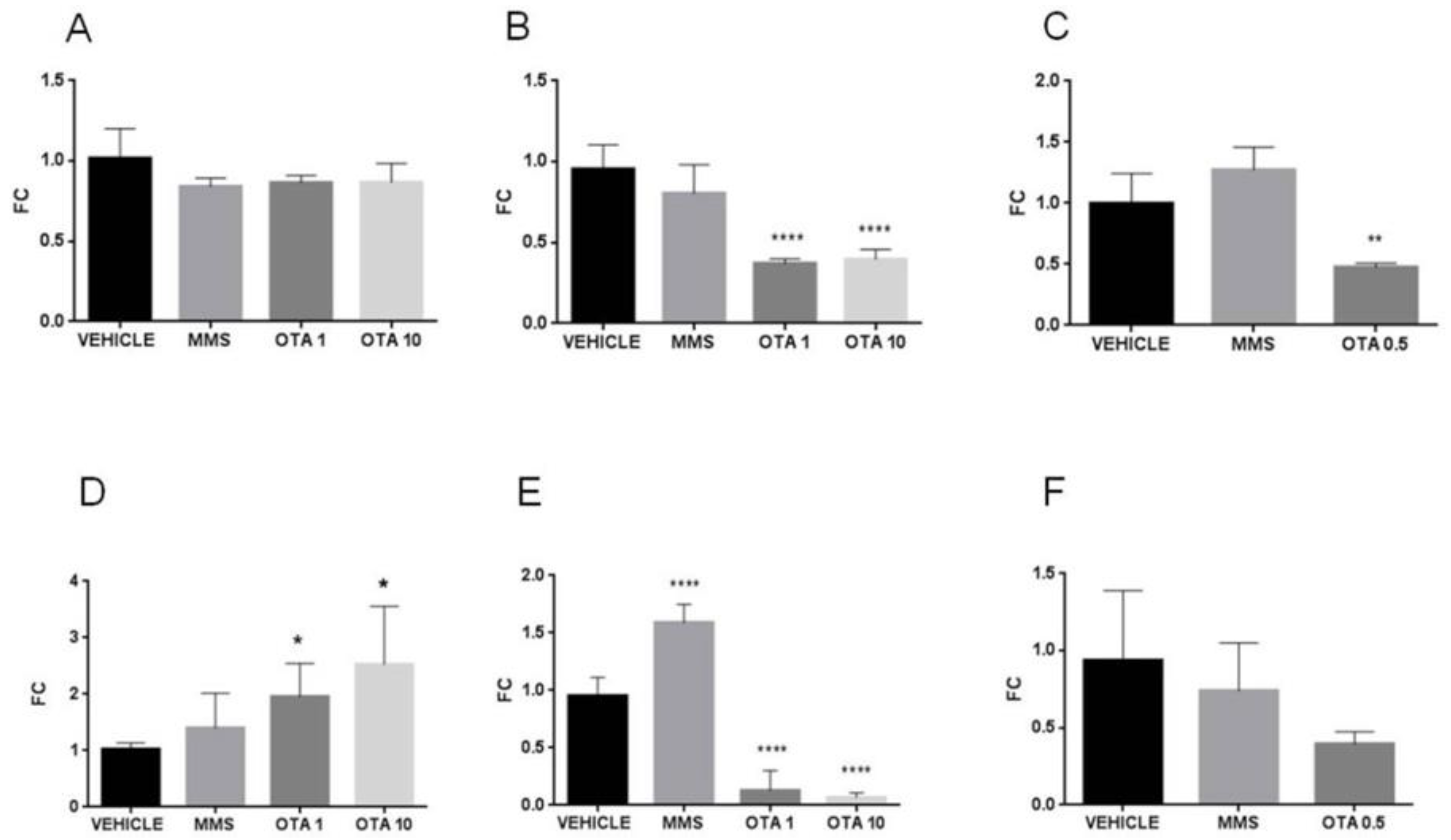
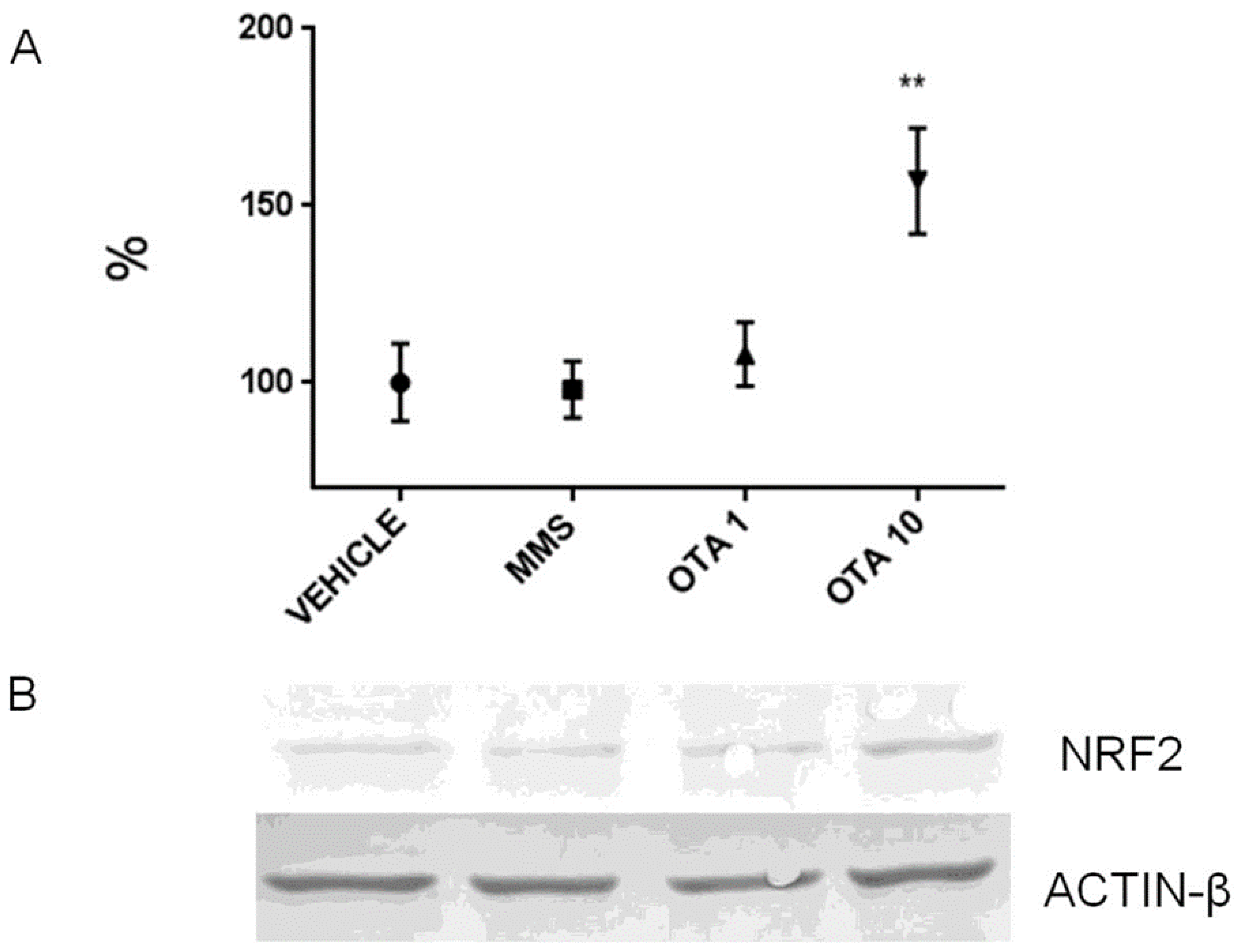
2.7. Expression Alteration at mRNA and Protein Level of Nrf2 and Keap1
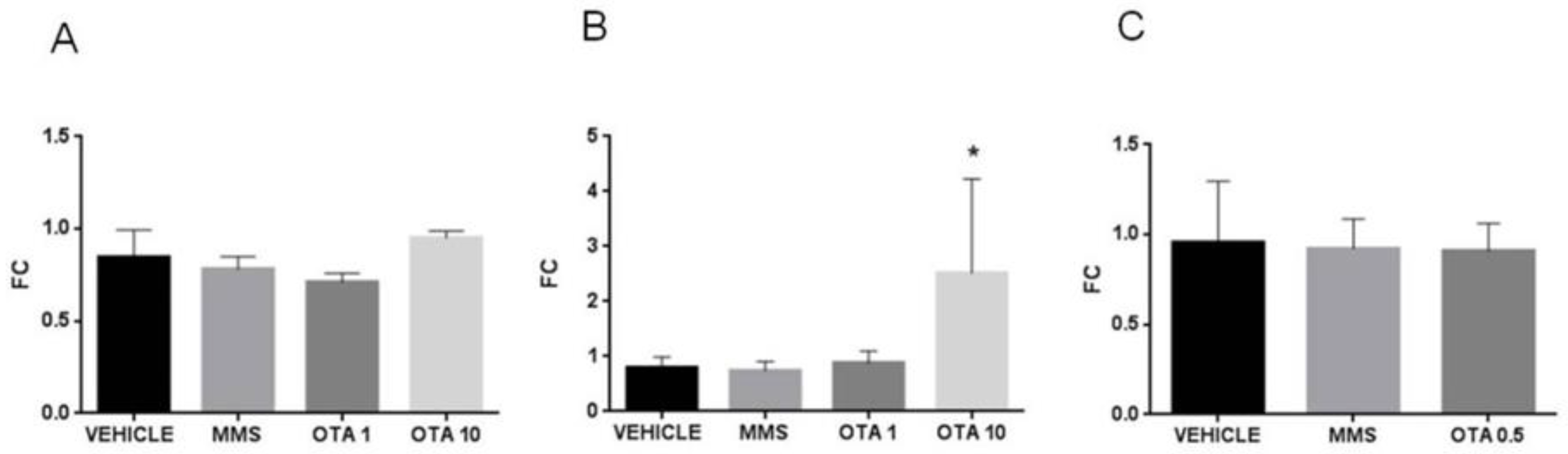
2.8. Gene Expression Alteration of the Antioxidant Response Element (ARE) Responsive Genes
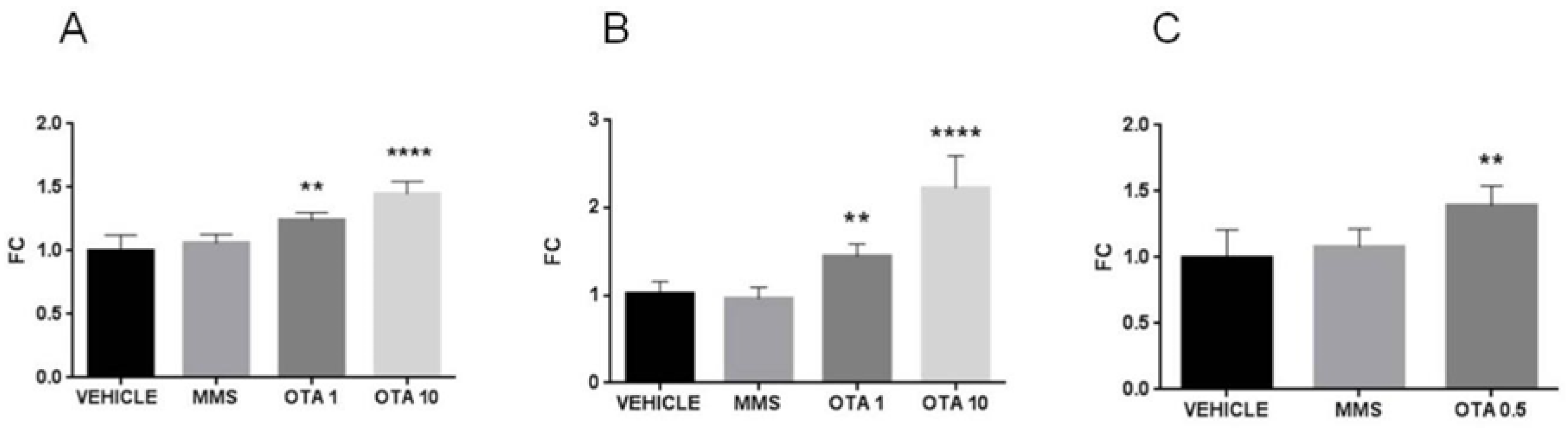
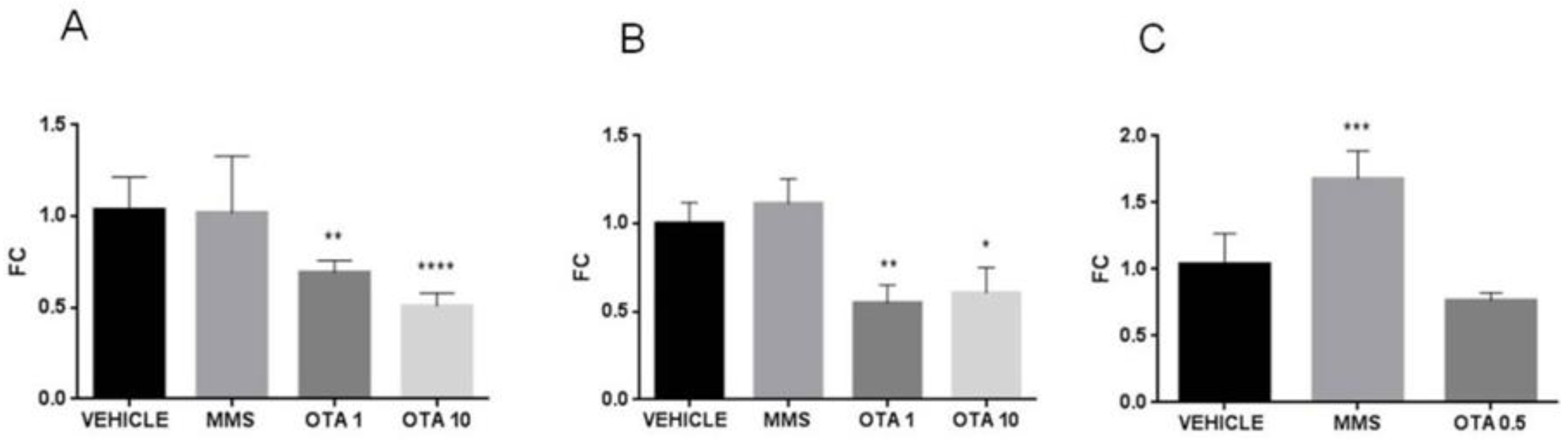
2.9. Expression Changes of the Duperoxide Dismutase (Sod) Enzymes in the Kidney
2.10. Hace1 and Rac1 mRNA Expression Changes by OTA Administration in the Kidney
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Diet
4.3. Measuring of the OTA Concentration in Blood Plasma by ELISA
4.4. Animal Treatment
4.5. Determination of Lipid Peroxidation and Antioxidant Parameters
4.6. Quantitative Real-Time PCR
4.7. Western Hybridization
4.8. Histology
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a Toxic Metabolite produced by Aspergillus ochraceus Wilh. Nat. Cell Biol. 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Krogh, P.; Hald, B.; Pedersen, E.J. Occurrence of Ochratoxin A and Citrinin in Cereals Associated with Mycotoxic Porcine Nephropathy. Acta Pathol. Microbiol. Scand. Sect. B Microbiol. Immunol. 1973, 81, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Krogh, P. Ochratoxin A residues in tissues of slaughter pigs with nephropathy. Nord. Veter. 1977, 29, 402–405. [Google Scholar]
- Elling, F.; Krogh, P. Fungal Toxins and Balkan (Endemic) Nephropathy. Lancet 1977, 1, 1213. [Google Scholar] [CrossRef]
- Krogh, P.; Hald, B.; Plestina, R.; Ceović, S. Balkan (endemic) nephropathy and foodborn ochratoxin A: Preliminary results of a survey of foodstuffs. Acta Pathol. Microbiol. Scand. Sect. B Microbiol. 1977, 85, 238–240. [Google Scholar] [CrossRef]
- Hald, B. Ochratoxin A in human blood in European countries. IARC Sci. Publ. 1991, 115, 159–164. [Google Scholar]
- Stoev, S.D. The role of ochratoxin A as a possible cause of Balkan endemic nephropathy and its risk evaluation. Vet. Res. Commun. 1998, 40, 352–360. [Google Scholar]
- Stoev, S.D.; Vitanov, S.; Anguelov, G.; Petkova-Bocharova, T.; Creppy, E.E. Experimental mycotoxic nephropathy in pigs provoked by a diet containing ochratoxin A and penicillic acid. Vet. Res. Commun. 2001, 25, 205–223. [Google Scholar] [CrossRef]
- Dirheimer, G.; Creppy, E.E. Mechanism of action of ochratoxin A. IARC Sci. Publ. 1991, 115, 171–186. [Google Scholar]
- Heller, K.; Röschenthaler, R. Inhibition of protein synthesis in Streptococcus faecalis by ochratoxin A. Can. J. Microbiol. 1978, 24, 466–472. [Google Scholar] [CrossRef]
- Castegnaro, M.; Pfohl-Leszkowicz, A.; Bartsch, H.; Tillmann, T.; Mohr, U. Re: Comments on paper by Son et al. Toxicol. Lett. 2005, 156, 315. [Google Scholar] [CrossRef] [PubMed]
- Clark, H.A.; Snedeker, S.M. Ochratoxin a: Its Cancer Risk and Potential for Exposure. J. Toxicol. Environ. Health Part B 2006, 9, 265–296. [Google Scholar] [CrossRef] [PubMed]
- Lühe, A.; Hildebrand, H.; Bach, U.; Dingermann, T.; Ahr, H.-J.J. A New Approach to Studying Ochratoxin A (OTA)-Induced Nephrotoxicity: Expression Profiling in Vivo and in Vitro Employing cDNA Microarrays. Toxicol. Sci. 2003, 73, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, G.J.; Nijmeijer, S.; Maas, R.; Roestenberg, P.; De Groene, E.; Fink-Gremmels, J. The role of oxidative stress in the ochratoxin A-mediated toxicity in proximal tubular cells. Biochim. Biophys. ActaMol. Basis Dis. 2002, 1588, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Hoehler, D.; Marquardt, R.R. Influence of Vitamins E and C on the Toxic Effects of Ochratoxin A and T-2 Toxin in Chicks. Poult. Sci. 1996, 75, 1508–1515. [Google Scholar] [CrossRef]
- Taniai, E.; Yafune, A.; Nakajima, M.; Hayashi, S.-M.; Nakane, F.; Itahashi, M.; Shibutani, M. Ochratoxin A induces karyomegaly and cell cycle aberrations in renal tubular cells without relation to induction of oxidative stress responses in rats. Toxicol. Lett. 2014, 224, 64–72. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell Survival Responses to Environmental Stresses Via the Keap1-Nrf2-ARE Pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Holzhäuser, D.; Higgins, L.; Bezençon, C.; Guignard, G.; Junod, S.; Richoz-Payot, J.; Gremaud, E.; et al. Reduction in Antioxidant Defenses may Contribute to Ochratoxin A Toxicity and Carcinogenicity. Toxicol. Sci. 2006, 96, 30–39. [Google Scholar] [CrossRef]
- Boesch-Saadatmandi, C.; Loboda, A.; Jozkowicz, A.; Huebbe, P.; Blank, R.; Wolffram, S.; Dulak, J.; Rimbach, G. Effect of ochratoxin A on redox-regulated transcription factors, antioxidant enzymes and glutathione-S-transferase in cultured kidney tubulus cells. Food Chem. Toxicol. 2008, 46, 2665–2671. [Google Scholar] [CrossRef]
- Wild, A.C.C.; Moinova, H.R.; Mulcahy, R.T. Regulation of gamma-glutamylcysteine synthetase subunit gene expression by the transcription factor Nrf2. J. Biol. Chem. 1999, 274, 33627–33636. [Google Scholar] [CrossRef] [Green Version]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; Yamamoto, M. Nrf2–Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc. Natl. Acad. Sci. USA 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slot, J.W.; Geuze, H.J.; A Freeman, B.; Crapo, J.D. Intracellular localization of the copper-zinc and manganese superoxide dismutases in rat liver parenchymal cells. Lab. Investig. 1986, 55, 363–371. [Google Scholar] [PubMed]
- Marklund, S.L.L. Extracellular superoxide dismutase in human tissues and human cell lines. J. Clin. Investig. 1984, 74, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Dansen, T.B.; Wirtz, K.W.A. The Peroxisome in Oxidative Stress. IUBMB Life 2001, 51, 223–230. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, Y.; Xu, W.; Luo, Y.; Hao, J.; Shen, X.L.; Yang, X.; Li, X.; Huang, K. Zinc protects HepG2 cells against the oxidative damage and DNA damage induced by ochratoxin A. Toxicol. Appl. Pharmacol. 2013, 268, 123–131. [Google Scholar] [CrossRef]
- Daugaard, M.; Nitsch, R.; Razaghi, B.; McDonald, L.; Jarrar, A.; Torrino, S.; Castillo-Lluva, S.; Rotblat, B.; Li, L.; Malliri, A.; et al. Hace1 controls ROS generation of vertebrate Rac1-dependent NADPH oxidase complexes. Nat. Commun. 2013, 4, 2180. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Diebold, B.A.; Hughes, Y.; Lambeth, J.D. Nox1-dependent Reactive Oxygen Generation Is Regulated by Rac1. J. Biol. Chem. 2006, 281, 17718–17726. [Google Scholar] [CrossRef] [Green Version]
- Boesch-Saadatmandi, C.; Wagner, A.E.; Graeser, A.C.; Hundhausen, C.; Wolffram, S.; Rimbach, G. Ochratoxin A impairs Nrf2-dependent gene expression in porcine kidney tubulus cells. J. Anim. Physiol. Anim. Nutr. 2009, 93, 547–554. [Google Scholar] [CrossRef]
- Stachurska, A.; Ciesla, M.; Kozakowska, M.; Wolffram, S.; Boesch-Saadatmandi, C.; Rimbach, G.; Józkowicz, A.; Dulak, J.; Loboda, A. Cross-talk between microRNAs, nuclear factor E2-related factor 2, and heme oxygenase-1 in ochratoxin A-induced toxic effects in renal proximal tubular epithelial cells. Mol. Nutr. Food Res. 2012, 57, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Gautier, J.-C.; Holzhaeuser, D.; Markovic, J.; Gremaud, E.; Schilter, B.; Turesky, R.J. Oxidative damage and stress response from ochratoxin an exposure in rats. Free Radic. Biol. Med. 2001, 30, 1089–1098. [Google Scholar] [CrossRef]
- Ramyaa, P.; Krishnaswamy, R.; Padma, V.V. Quercetin modulates OTA-induced oxidative stress and redox signalling in HepG2 cells—up regulation of Nrf2 expression and down regulation of NF-κB and COX-2. Biochim. Biophys. Acta 2014, 1840, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Ferenczi, S.; Cserháti, M.; Krifaton, C.; Szoboszlay, S.; Kukolya, J.; Szőke, Z.; Kőszegi, B.; Albert, M.; Barna, T.; Mézes, M.; et al. A New Ochratoxin a Biodegradation Strategy Using Cupriavidus basilensis Őr16 Strain. PLoS ONE 2014, 9, e109817. [Google Scholar] [CrossRef]
- A Nath, K.; Balla, G.; Vercellotti, G.M.; Balla, J.; Jacob, H.S.; Levitt, M.D.; E Rosenberg, M. Induction of heme oxygenase is a rapid, protective response in rhabdomyolysis in the rat. J. Clin. Investig. 1992, 90, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Alam, J. Mechanism of Heme Oxygenase-1 Gene Activation by Cadmium in MCF-7 Mammary Epithelial Cells. Role of OF p38 Kinase and Nrf2 Transcription Factor. J. Biol. Chem. 2000, 275, 27694–27702. [Google Scholar] [CrossRef] [Green Version]
- Sikorski, E.M.; Hock, T.; Hill-Kapturczak, N.; Agarwal, A. The story so far: Molecular regulation of the heme oxygenase-1 gene in renal injury. Am. J. Physiol. Physiol. 2004, 286, F425–F441. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Shi, L.; Huang, K.; Xu, W. Protective effect of N-acetylcysteine against DNA damage and S-phase arrest induced by ochratoxin A in human embryonic kidney cells (HEK-293). Food Chem. Toxicol. 2014, 70, 40–47. [Google Scholar] [CrossRef]
- Dunning, S.; Rehman, A.U.; Tiebosch, M.H.; Hannivoort, R.A.; Haijer, F.W.; Woudenberg, J.; Heuvel, F.A.V.D.; Buist-Homan, M.; Faber, K.N.; Moshage, H. Glutathione and antioxidant enzymes serve complementary roles in protecting activated hepatic stellate cells against hydrogen peroxide-induced cell death. Biochim. Biophys. Acta 2013, 1832, 2027–2034. [Google Scholar] [CrossRef] [Green Version]
- Marin-Kuan, M.; Nestler, S.; Verguet, C.; Bezencon, C.; Piguet, D.; Mansourian, R.; Holzwarth, J.; Grigorov, M.; Delatour, T.; Mantle, P.; et al. A Toxicogenomics Approach to Identify New Plausible Epigenetic Mechanisms of Ochratoxin a Carcinogenicity in Rat. Toxicol. Sci. 2006, 89, 120–134. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Abdel-Galil, M.M.; El-Lithey, M. Melatonin counteracts oxidative stress in rats fed an ochratoxin A contaminated diet. J. Pineal Res. 2005, 38, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Omar, R.F.; Hasinoff, B.B.; Mejilla, F.; Rahimtula, A.D. Mechanism of ochratoxin a stimulated lipid peroxidation. Biochem. Pharmacol. 1990, 40, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Marí, M.; Cederbaum, A.I. Induction of catalase, alpha, and microsomal glutathione S-transferase in CYP2E1 overexpressing HepG2 cells and protection against short-term oxidative stress. Hepatology 2001, 33, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Petrik, J.; Zanić-Grubisić, T.; Barišić, K.; Pepeljnjak, S.; Radić, B.; Ferencić, Z.; Cepelak, I. Apoptosis and oxidative stress induced by ochratoxin A in rat kidney. Arch. Toxicol. 2003, 77, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Tozlovanu, M.; Canadas, D.; Pfohl-Leszkowicz, A.; Frenette, C.; Paugh, R.J.; Manderville, R.A. Glutathione Conjugates of Ochratoxin A as Biomarkers of exposure. Arch. Ind. Hyg. Toxicol. 2012, 63, 417–427. [Google Scholar] [CrossRef]
- Niture, S.K.; Jain, A.K.; Jaiswal, A.K. Antioxidant-induced modification of INrf2 cysteine 151 and PKC-δ-mediated phosphorylation of Nrf2 serine 40 are both required for stabilization and nuclear translocation of Nrf2 and increased drug resistance. J. Cell Sci. 2009, 122, 4452–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, A.; Kang, M.-I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative Stress Sensor Keap1 Functions as an Adaptor for Cul3-Based E3 Ligase To Regulate Proteasomal Degradation of Nrf2. Mol. Cell. Biol. 2004, 24, 7130–7139. [Google Scholar] [CrossRef] [Green Version]
- Zeljezic, D.; Domijan, A.M.; Peraica, M. DNA damage by ochratoxin A in rat kidney assessed by the alkaline comet assay. Brazilian journal of medical and biological research. Rev. Bras. Pesqui. Med. Biol. 2006, 39, 1563–1568. [Google Scholar]
- Morehouse, L.A.; Tien, M.; Bucher, J.R.; Aust, S.D. Effect of hydrogen peroxide on the initiation of microsomal lipid peroxidation. Biochem. Pharmacol. 1983, 32, 123–127. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, sensitive and specific thiobarbituric acid method for measuring lipid peroxidation in animal tissue, food and feedstuff samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Lawrence, R.A. Reprint of “Glutathione Peroxidase Activity in Selenium-Deficient Rat Liver”. Biochem. Biophys. Res. Commun. 2012, 425, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,5′-dithiobis(2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef]
- Mannervik, B.; Alin, P.; Guthenberg, C.; Jensson, H.; Tahir, M.K.; Warholm, M.; Jornvall, H. Identification of three classes of cytosolic glutathione transferase common to several mammalian species: Correlation between structural data and enzymatic properties. Proc. Natl. Acad. Sci. USA 1985, 82, 7202–7206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Cui, X.; Zhou, J.; Qiu, J.; Johnson, M.R.; Mrug, M. Validation of endogenous internal real-time PCR controls in renal tissues. Am. J. Nephrol. 2009, 30, 413–417. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | 24 h | 24 h | 72 h | 72 h | 21 Days |
|---|---|---|---|---|---|
| Dose | single | single | repeated | repeated | repeated |
| 1 mg/kg bw | 10 mg/kg bw | 1 mg/kg bw | 10 mg/kg bw | 0.5 mg/kg bw | |
| OTA level (ng/mL) | 843.02 ± 285.16 | 2717.88 ± 391.52 | 269.73 ± 60.6 | 1969.28 ± 654.6 | 231.35 ± 50.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferenczi, S.; Kuti, D.; Cserháti, M.; Krifaton, C.; Szoboszlay, S.; Kukolya, J.; Szőke, Z.; Albert, M.; Kriszt, B.; Kovács, K.J.; et al. Effects of Single and Repeated Oral Doses of Ochratoxin A on the Lipid Peroxidation and Antioxidant Defense Systems in Mouse Kidneys. Toxins 2020, 12, 732. https://doi.org/10.3390/toxins12110732
Ferenczi S, Kuti D, Cserháti M, Krifaton C, Szoboszlay S, Kukolya J, Szőke Z, Albert M, Kriszt B, Kovács KJ, et al. Effects of Single and Repeated Oral Doses of Ochratoxin A on the Lipid Peroxidation and Antioxidant Defense Systems in Mouse Kidneys. Toxins. 2020; 12(11):732. https://doi.org/10.3390/toxins12110732
Chicago/Turabian StyleFerenczi, Szilamér, Dániel Kuti, Mátyás Cserháti, Csilla Krifaton, Sándor Szoboszlay, József Kukolya, Zsuzsanna Szőke, Mihály Albert, Balázs Kriszt, Krisztina J. Kovács, and et al. 2020. "Effects of Single and Repeated Oral Doses of Ochratoxin A on the Lipid Peroxidation and Antioxidant Defense Systems in Mouse Kidneys" Toxins 12, no. 11: 732. https://doi.org/10.3390/toxins12110732
APA StyleFerenczi, S., Kuti, D., Cserháti, M., Krifaton, C., Szoboszlay, S., Kukolya, J., Szőke, Z., Albert, M., Kriszt, B., Kovács, K. J., Mézes, M., & Balogh, K. (2020). Effects of Single and Repeated Oral Doses of Ochratoxin A on the Lipid Peroxidation and Antioxidant Defense Systems in Mouse Kidneys. Toxins, 12(11), 732. https://doi.org/10.3390/toxins12110732