Chemical and Biological Study of Novel Aplysiatoxin Derivatives from the Marine Cyanobacterium Lyngbya sp.

 ,
, 
Abstract
:1. Introduction
2. Results
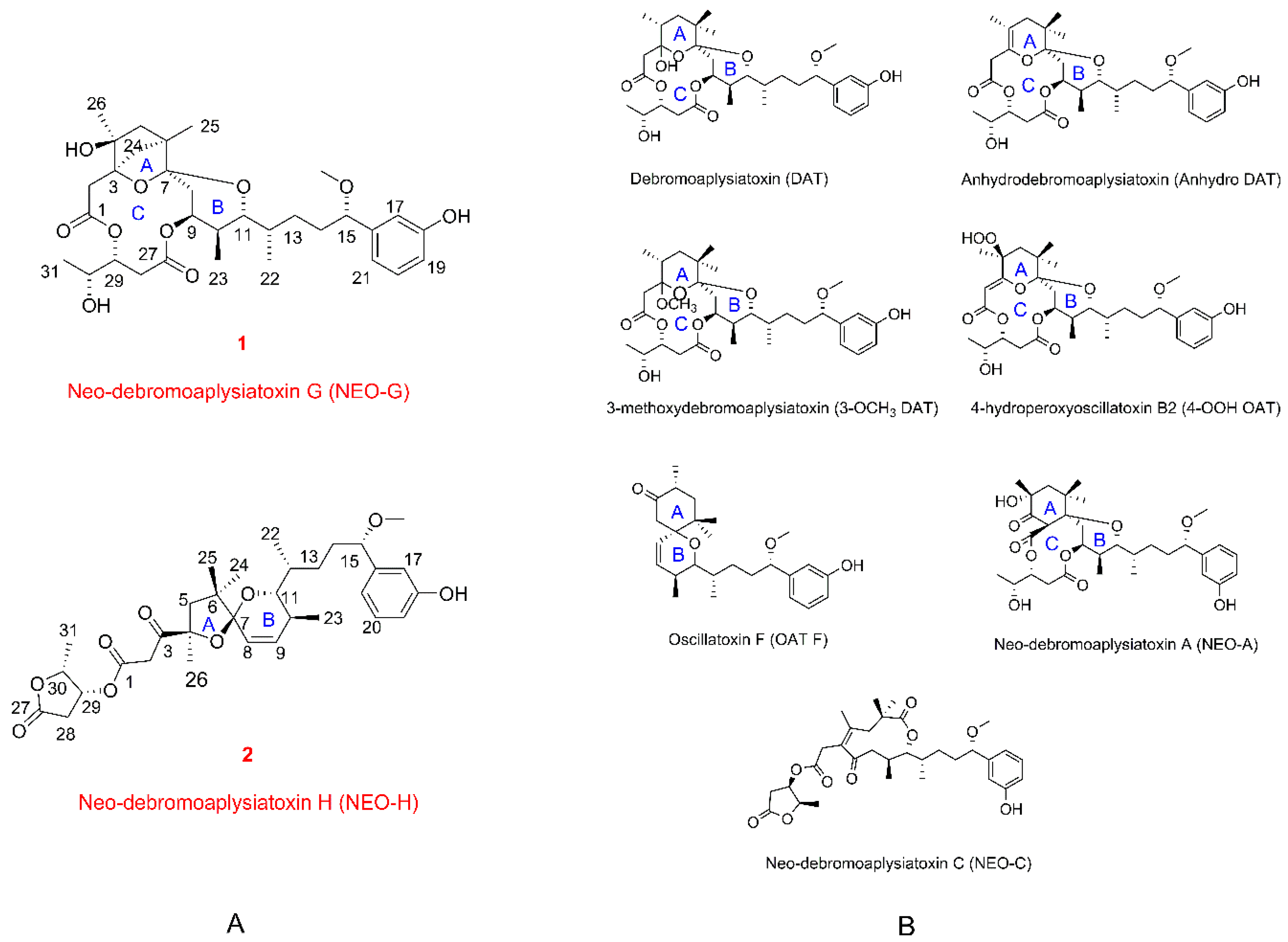
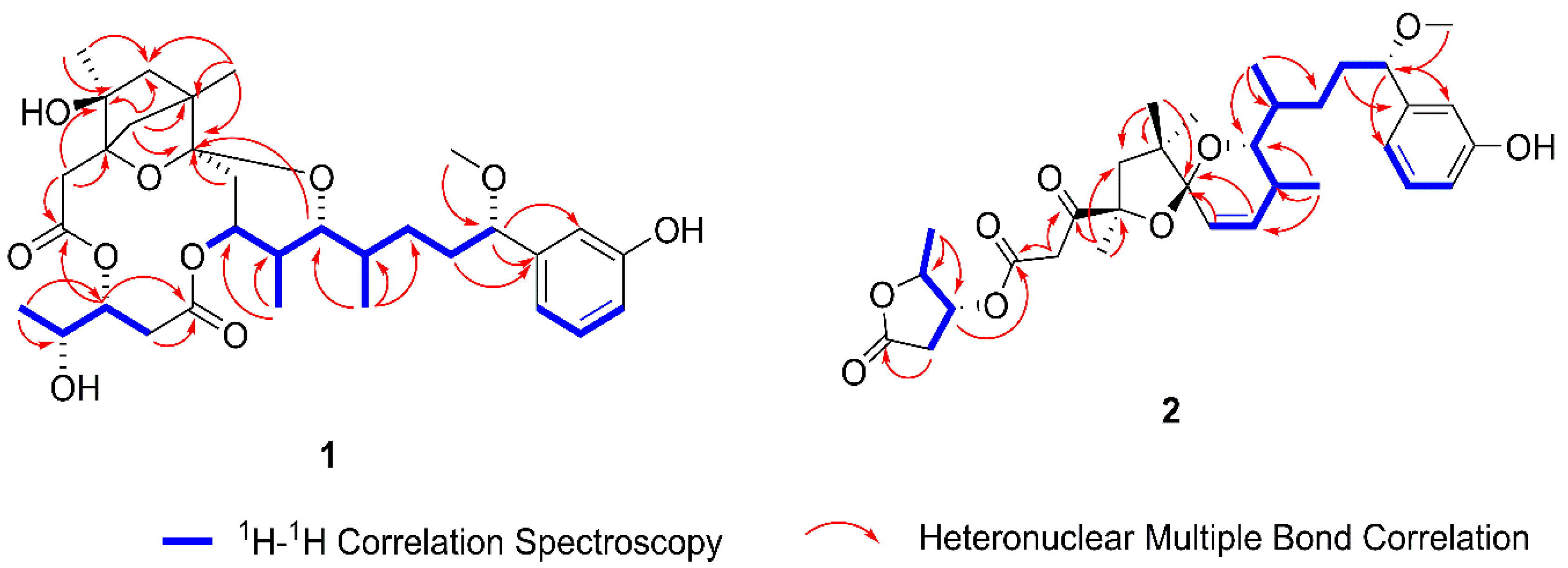
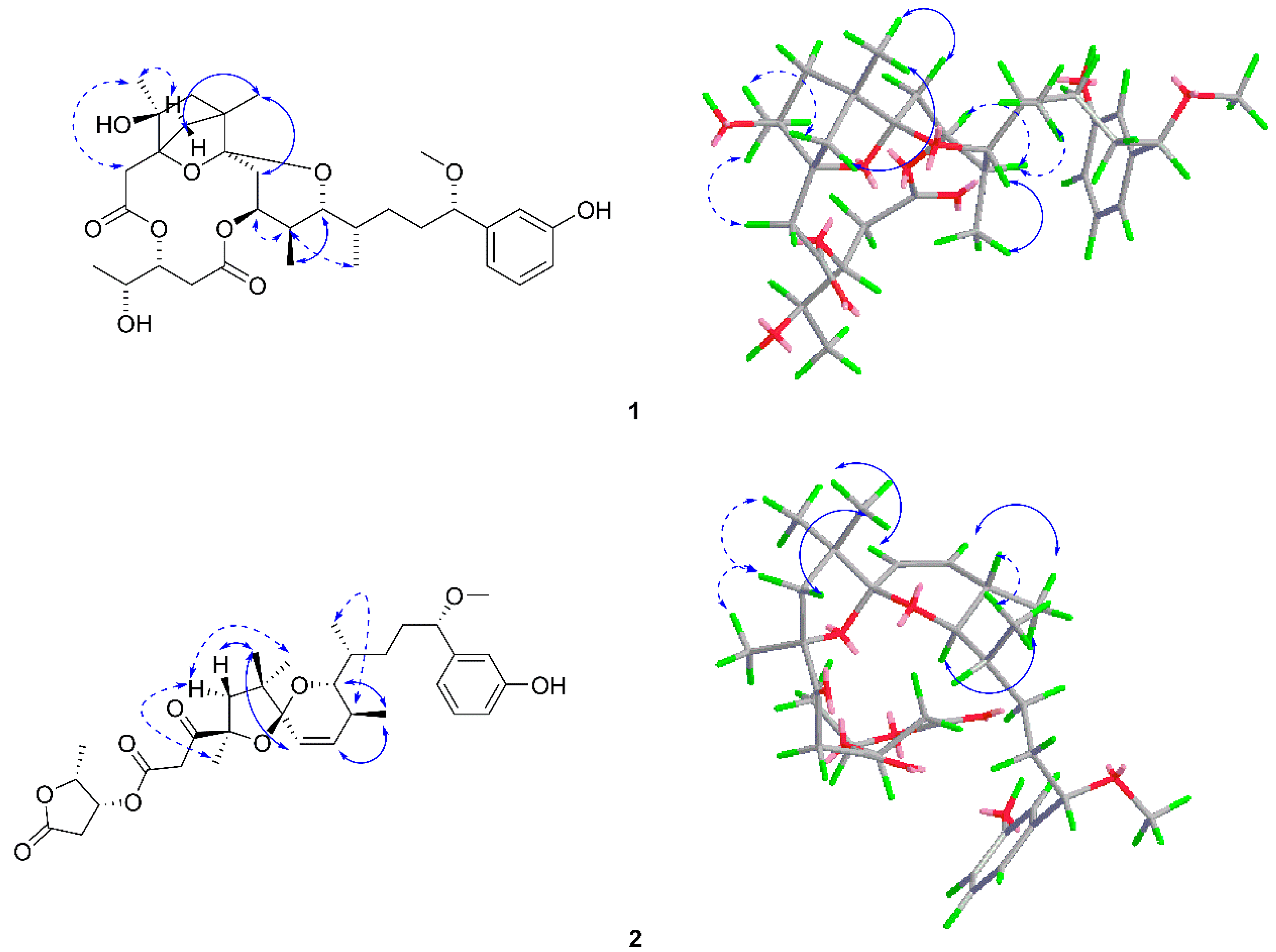
2.1. Structure Elucidation of the New Compounds
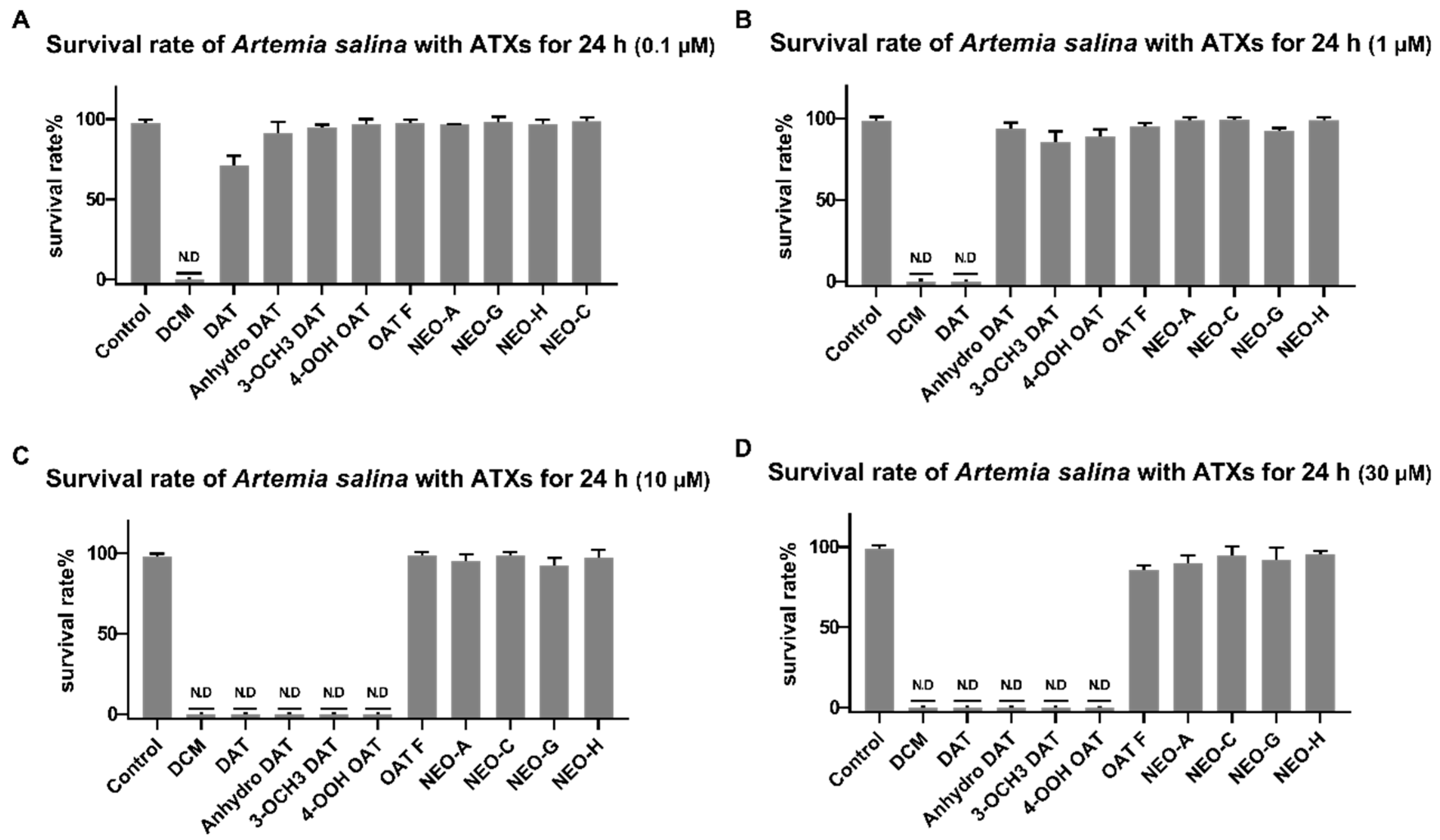
2.2. Biological Activities of the Isolated Compounds
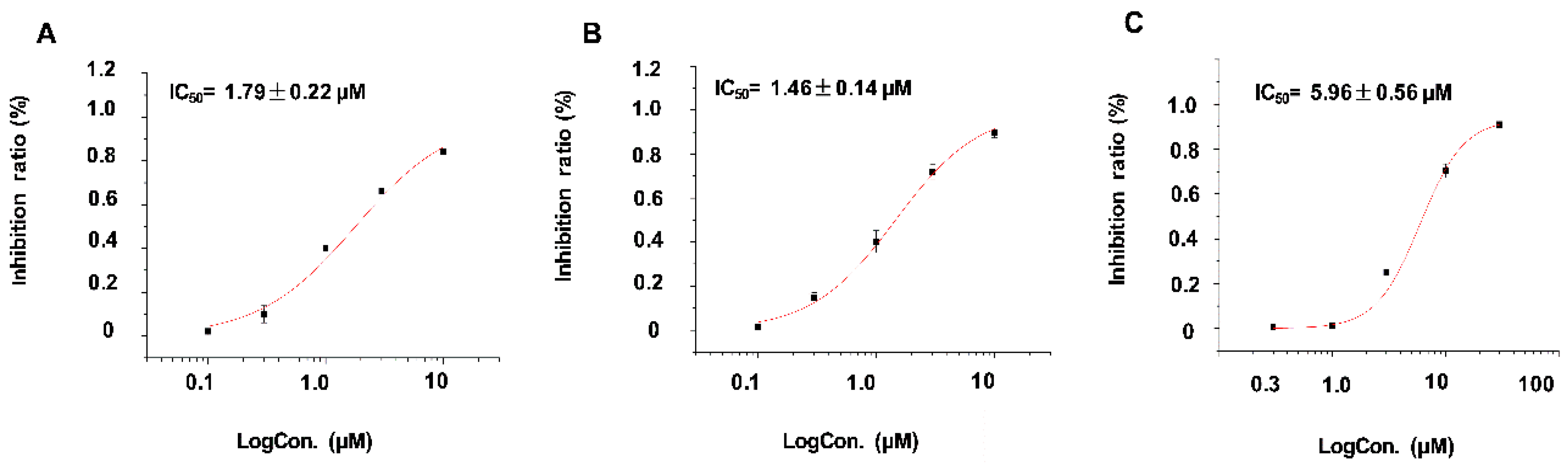
2.2.1. Inhibitory Activities against Kv1.5
2.2.2. Toxicity of Brine Shrimp
3. Discussion
4. Materials and Methods
4.1. Material of Marine Cyanobacterium Lyngbya sp.
4.2. General Experimental Procedures
4.3. Extraction and Isolation
4.4. Ion Channel Inhibitory Experiment
4.5. Brine Shrimp Toxicity Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Niamien-Ebrottie, J.E.; Bhattacharyya, S.; Deep, P.R.; Nayak, B. Cyanobacteria and cyanotoxins in the World: Review. Int. J. Appl. Res. 2015, 1, 563–569. [Google Scholar]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria-a prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar]
- Serdula, M.; Bartolini, G.; Moore, R.E.; Gooch, J.; Wiebenga, N. Seaweed itch on windward Oahu. Hawaii Med. J. 1982, 41, 200–201. [Google Scholar]
- Nagai, H.; Yasumoto, T.; Hokama, Y. Aplysiatoxin and debromoaplysiatoxin as the causative agents of a red alga Gracilaria coronopifolia poisoning in Hawaii. Toxicon Off. J. Int. Soc. Toxinol. 1996, 34, 753–761. [Google Scholar]
- Nagai, H.; Yasumoto, T.; Hokama, Y. Manauealides, Some of the Causative Agents of a Red Alga Gracilaria coronopifolia Poisoning in Hawaii. J. Nat. Prod. 1997, 60, 925–928. [Google Scholar]
- Nagai, H.; Kan, Y.; Fujita, T.; Sakamoto, B.; Hokama, Y. Manauealide C and Anhydrodebromoaplysiatoxin, Toxic Constituents of the Hawaiian Red Alga, Gracilaria coronopifolia. Biosci. Biotechnol. Biochem. 1998, 62, 1011–1013. [Google Scholar]
- Kato, Y.; Scheuer, P.J. Aplysiatoxin and debromoaplysiatoxin, constituents of the marine mollusk Stylocheilus longicauda (Quoy and Gaimard, 1824). J. Am. Chem. Soc. 1974, 96, 2245–2246. [Google Scholar]
- Watson, M. Midgut gland toxins of Hawaiian sea hares. I. Isolation and preliminary toxicological observations. Toxicon Off. J. Int. Soc. Toxinol. 1973, 11, 259–267. [Google Scholar]
- Mynderse, J.S.; Moore, R.E.; Kashiwagi, M.; Norton, T.R. Antileukemia activity in the Osillatoriaceae: Isolation of Debromoaplysiatoxin from Lyngbya. Science 1977, 196, 538–540. [Google Scholar]
- Luesch, H.; Harrigan, G.G.; Goetz, G.; Horgen, F.D. The cyanobacterial origin of potent anticancer agents originally isolated from sea hares. Curr. Med. Chem. 2002, 9, 1791–1806. [Google Scholar]
- Moore, R.E.; Blackman, A.J.; Cheuk, C.E.; Mynderse, J.S.; Matsumoto, G.K.; Clardy, J.; Woodard, R.W.; Craig, J.C. Cheminform Abstract: Absolute stereochemistries of the aplysiatoxins and oscillatoxin A. Cheminform 1984, 15, 2484–2489. [Google Scholar]
- Chlipala, G.E.; Pham, H.T.; Nguyen, V.H.; Krunic, A.; Shim, S.H.; Soejarto, D.D.; Orjala, J. Nhatrangins A and B, aplysiatoxin-related metabolites from the marine cyanobacterium Lyngbya majuscula from Vietnam. J. Nat. Prod. 2010, 73, 784–787. [Google Scholar] [PubMed] [Green Version]
- Gupta, D.K.; Kaur, P.; Leong, S.T.; Tan, L.T.; Prinsep, M.R.; Chu, J.J. Anti-Chikungunya viral activities of aplysiatoxin-related compounds from the marine cyanobacterium Trichodesmium erythraeum. Mar. Drugs 2014, 12, 115–127. [Google Scholar] [PubMed] [Green Version]
- Nagai, H.; Watanabe, M.; Sato, S.; Kawaguchi, M.; Xiao, Y.Y.; Hayashi, K.; Watanabe, R.; Uchida, H.; Satake, M. New aplysiatoxin derivatives from the Okinawan cyanobacterium Moorea producens. Tetrahedron 2019, 75, 2486–2494. [Google Scholar]
- Yanagita, R.C.; Kamachi, H.; Kikumori, M.; Tokuda, H.; Suzuki, N.; Suenaga, K.; Nagai, H.; Irie, K. Effects of the methoxy group in the side chain of debromoaplysiatoxin on its tumor-promoting and anti-proliferative activities. Bioorg. Med. Chem. Lett. 2013, 23, 4319–4323. [Google Scholar]
- Kikumori, M.; Yanagita, R.C.; Tokuda, H.; Suzuki, N.; Nagai, H.; Suenaga, K.; Irie, K. Structure-activity studies on the spiroketal moiety of a simplified analogue of debromoaplysiatoxin with antiproliferative activity. J. Med. Chem. 2012, 55, 5614–5626. [Google Scholar]
- Kamachi, H.; Tanaka, K.; Yanagita, R.C.; Murakami, A.; Murakami, K.; Tokuda, H.; Suzuki, N.; Nakagawa, Y.; Irie, K. Structure-activity studies on the side chain of a simplified analog of aplysiatoxin (aplog-1) with anti-proliferative activity. Bioorg. Med. Chem. 2013, 21, 2695–2702. [Google Scholar]
- Suganuma, M.; Fujiki, H.; Tahira, T.; Cheuk, C.; Moore, R.E.; Sugimura, T. Estimation of tumor promoting activity and structure-function relationships of aplysiatoxins. Carcinogenesis 1984, 5, 315–318. [Google Scholar]
- Tidgewell, K.; Clark, B.R.; Gerwick, W.H. The natural products chemistry of cyanobacteria. Compr. Nat. Prod. 2010, 2, 141–188. [Google Scholar]
- Han, B.N.; Liang, T.T.; Keen, L.J.; Fan, T.T.; Zhang, X.D.; Xu, L.; Zhao, Q.; Wang, S.P.; Lin, H.W. Two Marine Cyanobacterial Aplysiatoxin Polyketides, Neo-debromoaplysiatoxin A and B, with K+ Channel Inhibition Activity. Org. Lett. 2018, 20, 578–581. [Google Scholar]
- Fan, T.T.; Zhang, H.H.; Tang, Y.H.; Zhang, F.Z.; Han, B.N. Two New Neo-debromoaplysiatoxins-A Pair of Stereoisomers Exhibiting Potent Kv1.5 Ion Channel Inhibition Activities. Mar. Drugs 2019, 17, 652. [Google Scholar]
- Tang, Y.H.; Liang, T.T.; Fan, T.T.; Keen, L.J.; Zhang, X.D.; Xu, L.; Zhao, Q.; Zeng, R.; Han, B.N. Neo-debromoaplysiatoxin C, with new structural rearrangement, derived from debromoaplysiatoxin. Nat. Prod. Res. 2020, 34, 2151–2156. [Google Scholar] [PubMed]
- Nagai, H.; Sato, S.; Iida, K.; Hayashi, K.; Kawaguchi, M.; Uchida, H.; Satake, M. Oscillatoxin I: A New Aplysiatoxin Derivative, from a Marine Cyanobacterium. Toxins 2019, 11, 366. [Google Scholar]
- Kawaguchi, M.; Satake, M.; Zhang, B.T.; Xiao, Y.Y.; Fukuoka, M.; Uchida, H.; Nagai, H. Neo-Aplysiatoxin A Isolated from Okinawan Cyanobacterium Moorea Producens. Molecules 2020, 25, 457. [Google Scholar]
- Taylor, M.S.; Stahl-Timmins, W.; Redshaw, C.H.; Osborne, N.J. Toxic alkaloids in Lyngbya majuscula and related tropical marine cyanobacteria. Harmful Algae 2014, 31, 1–8. [Google Scholar] [PubMed]
- Trapani, J.G.; Korn, S.J. Effect of external pH on activation of the Kv1.5 potassium channel. Biophys. J. 2003, 84, 195–204. [Google Scholar] [PubMed] [Green Version]
- Heijman, J.; Dobrev, D. Ion channels as part of macromolecular multiprotein complexes: Clinical significance. Ionenkanäle als Teil makromolekularer Multiproteinkomplexe: Klinische Signifikanz. Herzschrittmacherther. Elektrophysiol. 2018, 29, 30–35. [Google Scholar]
- Paul, V.J.; Thacker, R.W.; Banks, K.; Golubic, S. Benthic cyanobacterial bloom impacts the reefs of South Florida (Broward County, USA). Coral Reefs 2005, 24, 693–697. [Google Scholar]
- Tiling, K.; Proffitt, C.E. Effects of Lyngbya majuscula blooms on the seagrass Halodule wrightii and resident invertebrates. Harmful Algae 2017, 62, 104–112. [Google Scholar]
- Osborne, N.J.; Webb, P.M.; Shaw, G.R. The toxins of Lyngbya majuscula and their human and ecological health effects. Environ. Int. 2001, 27, 381–392. [Google Scholar]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar]
- Ashida, Y.; Yanagita, R.C.; Takahashi, C.; Kawanami, Y.; Irie, K. Binding mode prediction of aplysiatoxin, a potent agonist of protein kinase C, through molecular simulation and structure-activity study on simplified analogs of the receptor-recognition domain. Bioorg. Med. Chem. 2016, 24, 4218–4227. [Google Scholar] [PubMed]
- Tang, Y.H.; Wu, J.; Fan, T.T.; Zhang, H.H.; Gong, X.X.; Cao, Z.Y.; Zhang, J.; Lin, H.W.; Han, B.N. Chemical and biological study of aplysiatoxin derivatives showing inhibition of potassium channel Kv1.5. RSC Adv. 2019, 9, 7594–7600. [Google Scholar]
- Horowitz, A.D.; Fujiki, H.; Weinstein, I.B.; Jeffrey, A.; Okin, E.; Moore, R.E.; Sugimura, T. Comparative effects of aplysiatoxin, debromoaplysiatoxin, and teleocidin on receptor binding and phospholipid metabolism. Cancer Res. 1983, 43, 1529–1535. [Google Scholar] [PubMed]
- Yu, J.; Park, M.H.; Jo, S.H. Inhibitory effects of cortisone and hydrocortisone on human Kv1.5 channel currents. Eur. J. Pharmacol. 2015, 746, 158–166. [Google Scholar] [PubMed]
- Jeong, I.; Yoon, S.H.; Hahn, S.J. Effects of dapoxetine on cloned Kv1.5 channels expressed in CHO cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 707–716. [Google Scholar]
- Choi, B.H.; Choi, J.S.; Jeong, S.W.; Hahn, S.J.; Yoon, S.H.; Jo, Y.H.; Kim, M.S. Direct block by bisindolylmaleimide of rat Kv1.5 expressed in Chinese hamster ovary cells. J. Pharmacol. Exp. Ther. 2000, 293, 634–640. [Google Scholar]
- Vanhaecke, P.; Persoone, G.; Claus, C.; Sorgeloos, P. Proposal for a short-term toxicity test with artemia nauplii. Ecotoxicol. Environ. Saf. 1981, 5, 382–387. [Google Scholar]
- Lopes, V.R.; Fernández, N.; Martins, R.F.; Vasconcelos, V. Primary screening of the bioactivity of brackishwater cyanobacteria: Toxicity of crude extracts to Artemia salina larvae and Paracentrotus lividus embryos. Mar. Drugs 2010, 8, 471–482. [Google Scholar]
- Moshafi, M.H.; Sharififar, F.; Dehghan, G.; Ameri, A. Bioassay screening of the essential oil and various extracts of fruits of Heracleum persicum Desf. and Rhizomes of Zingiber officinale Rosc. using brine shrimp cytotoxicity assay. Iran. J. Pharm. Res. 2009, 8, 59–63. [Google Scholar]
- Manilal, A.; Sujith, S.; Kiran, G.S.; Selvin, J.; Shakir, C. Cytotoxic potentials of red alga, Laurencia brandenii collected from the Indian coast. Glob. J. Pharmacol. 2009, 3, 90–94. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pos. | 1 | 2 | ||
|---|---|---|---|---|
| δH (J in Hz) | δC | δH (J in Hz) | δC | |
| 1 | 172.1 | 167.4 | ||
| 2 | a 2.72, d (12.6) b 2.69, d (12.6) | 34.8 | a 4.10, d (16.5) b 3.45, d (16.5) | 43.5 |
| 3 | 85 | 206.5 | ||
| 4 | 79.2 | 86.2 | ||
| 5 | a 2.02, d (10.5) b 1.84, d (10.5, 2.9) | 47.1 | a 2.59, d (12.7) b 1.68, d (12.7) | 46.4 |
| 6 | 48.7 | 47,3 | ||
| 7 | 105.5 | 106.8 | ||
| 8 | a 2.24, dd (14.3, 3.1) b 1.59, dd (14.3, 3.2) | 34 | 5.60, dd (10.2, 2.7) | 122.6 |
| 9 | 4.89, m | 74.1 | 5.86, dd (10.2, 2.7) | 138.1 |
| 10 | 1.69, m | 34 | 2.16, m | 30.0 |
| 11 | 3.86, d (10.7) | 74 | 3.21, overlap | 78.7 |
| 12 | 1.52, m | 33.9 | 1.60, overlap | 34.6 |
| 13 | a 1.52, m b 1.41, m | 31.3 | 1.27 | 29.4 |
| 14 | a 1.79, m b 1.62, m | 37.5 | a 1.85, m b 1.60, overlap | 36.3 |
| 15 | 4.00, dd (8.2, 5.0) | 84.6 | 3.94, t-like (6.7) | 83.5 |
| 16 | 144.7 | 144 | ||
| 17 | 6.79, t-like (2.0) | 113.5 | 6.77, overlap | 113.9 |
| 18 | 156.3 | 156.1 | ||
| 19 | 6.76, ddd (8.0, 2.6, 1.2) | 114.7 | 6.77, overlap | 114.6 |
| 20 | 7.22, t-like (8.0) | 129.8 | 7.21, t-like (7.7) | 129.7 |
| 21 | 6.86, dt (8.0, 1.2) | 118.2 | 6.83, d (7.4) | 118.5 |
| 22 | 0.81, d (6.4) | 12.1 | 0.81, d (6.8) | 13.3 |
| 23 | 0.78, d (6.9) | 13.6 | 0.90, d (6.4) | 17.0 |
| 24 | a 2.47, dd (12.9, 2.9) b 1.31, d (12.9) | 47.2 | 0.89, s | 23.0 |
| 25 | 1.01, s | 15.7 | 1.09, s | 26.3 |
| 26 | 1.41, s | 22.3 | 1.43, s | 25.7 |
| 27 | 171 | 174.3 | ||
| 28 | a 2.95, dd (14.8, 4.7) b 2.56, d (14.8, 7.4) | 36.3 | a 2.92, dd (18.3, 6.1) b 2.70, d (18.3) | 36.7 |
| 29 | 4.81, m | 77.1 | 5.51, t-like (5.1) | 72.1 |
| 30 | 4.22, m | 68.6 | 4.73, m | 79.1 |
| 31 | 1.23, d (6.4) | 18.5 | 1.39, d (6.6) | 14.1 |
| 15-OCH3 | 3.26, s | 57.2 | 3.21, overlap | 56.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-H.; Zhang, X.-K.; Si, R.-R.; Shen, S.-C.; Liang, T.-T.; Fan, T.-T.; Chen, W.; Xu, L.-H.; Han, B.-N. Chemical and Biological Study of Novel Aplysiatoxin Derivatives from the Marine Cyanobacterium Lyngbya sp. Toxins 2020, 12, 733. https://doi.org/10.3390/toxins12110733
Zhang H-H, Zhang X-K, Si R-R, Shen S-C, Liang T-T, Fan T-T, Chen W, Xu L-H, Han B-N. Chemical and Biological Study of Novel Aplysiatoxin Derivatives from the Marine Cyanobacterium Lyngbya sp. Toxins. 2020; 12(11):733. https://doi.org/10.3390/toxins12110733
Chicago/Turabian StyleZhang, Hui-Hui, Xin-Kai Zhang, Ran-Ran Si, Si-Cheng Shen, Ting-Ting Liang, Ting-Ting Fan, Wei Chen, Lian-Hua Xu, and Bing-Nan Han. 2020. "Chemical and Biological Study of Novel Aplysiatoxin Derivatives from the Marine Cyanobacterium Lyngbya sp." Toxins 12, no. 11: 733. https://doi.org/10.3390/toxins12110733
APA StyleZhang, H. -H., Zhang, X. -K., Si, R. -R., Shen, S. -C., Liang, T. -T., Fan, T. -T., Chen, W., Xu, L. -H., & Han, B. -N. (2020). Chemical and Biological Study of Novel Aplysiatoxin Derivatives from the Marine Cyanobacterium Lyngbya sp. Toxins, 12(11), 733. https://doi.org/10.3390/toxins12110733