Dual Function of a Novel Bacterium, Slackia sp. D-G6: Detoxifying Deoxynivalenol and Producing the Natural Estrogen Analogue, Equol



Abstract
:1. Introduction
2. Results
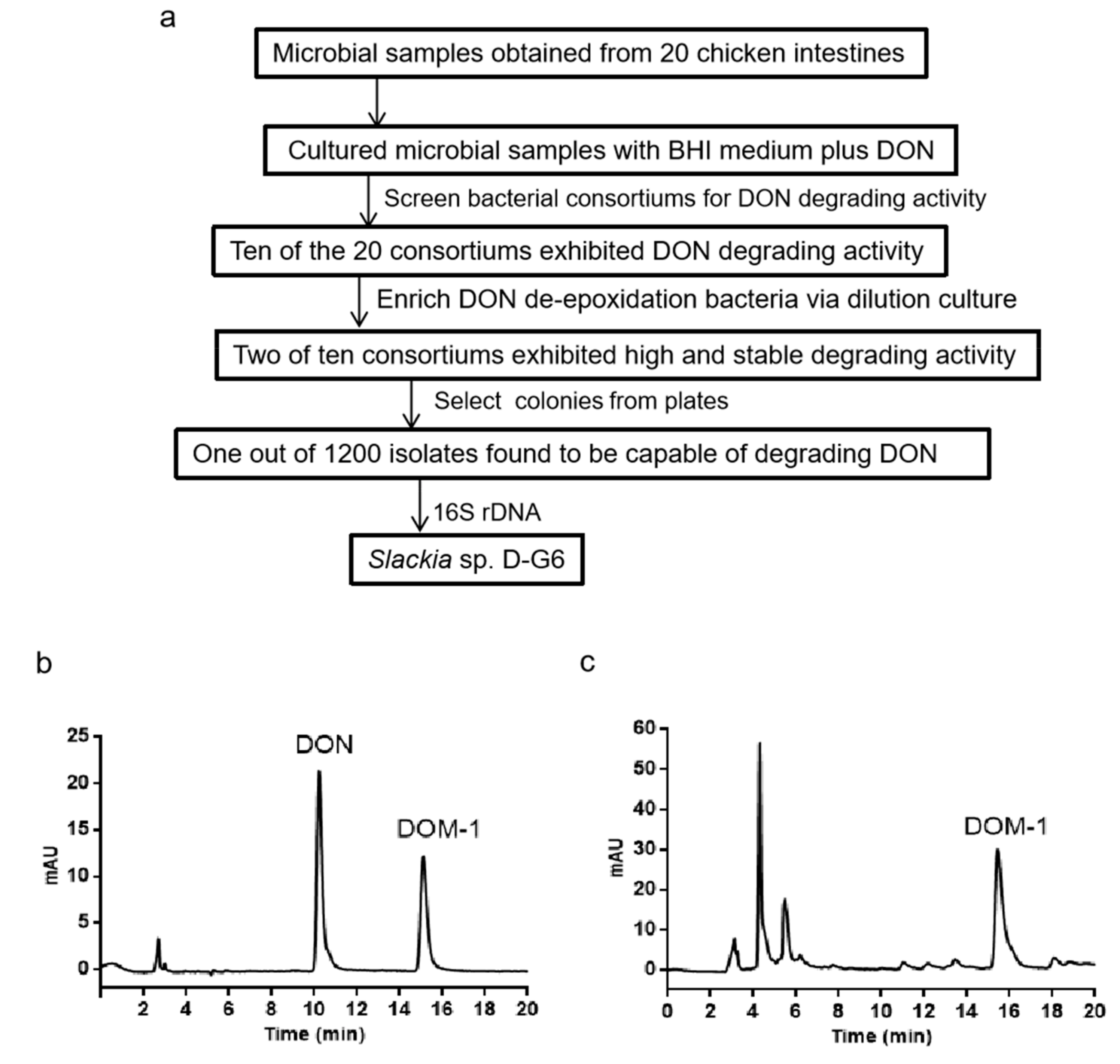
2.1. Isolating the Bacterium Capable of DON De-Epoxidation from Chicken Intestines
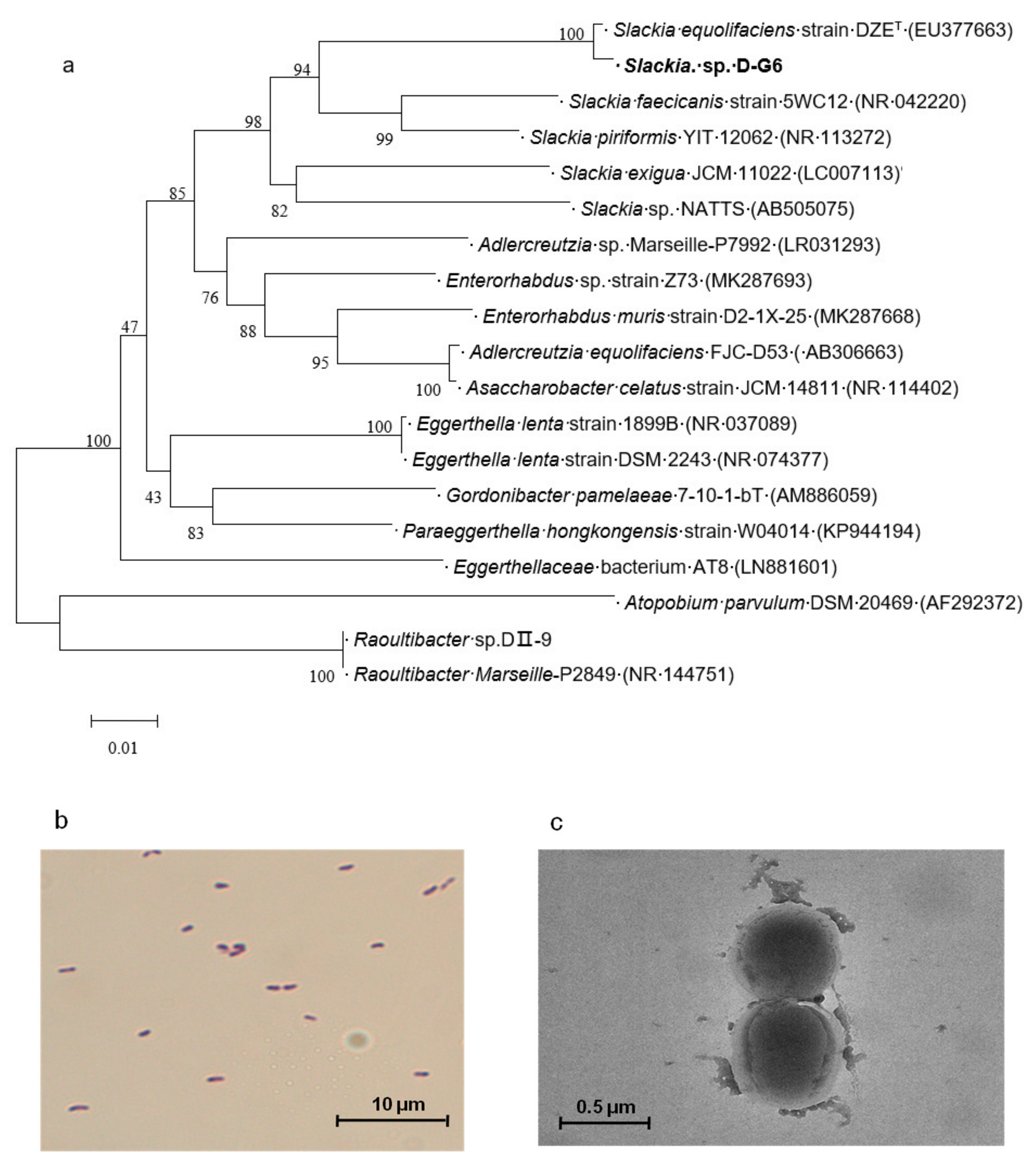
2.2. Identification of the Newly Isolated Bacterium, Slackia sp. D-G6
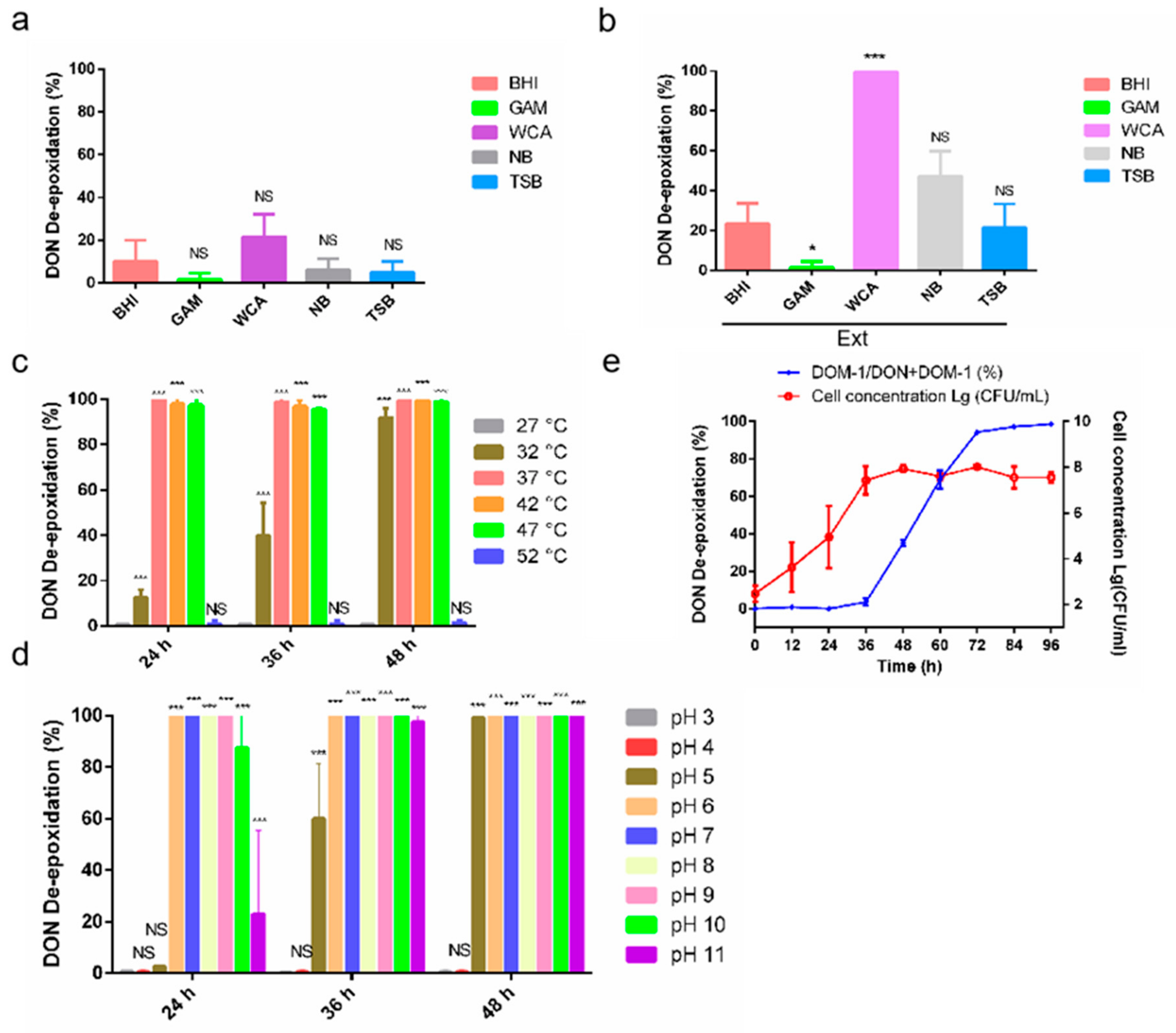
2.3. The Metabolic Characteristics of D-G6
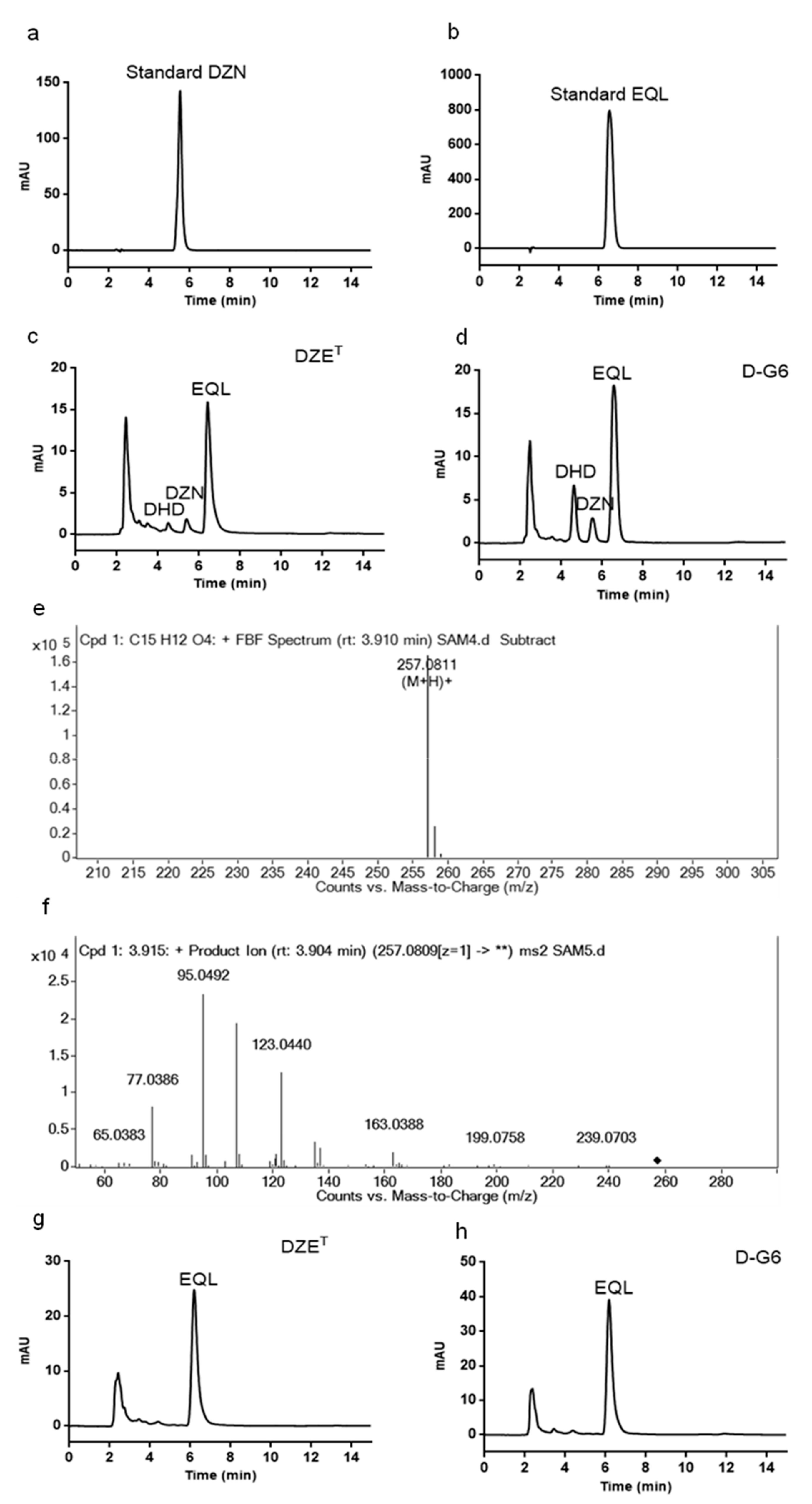
2.4. Slackia sp. D-G6 is Capable of Producing the Natural Estrogen Analogue, EQL, from DZN
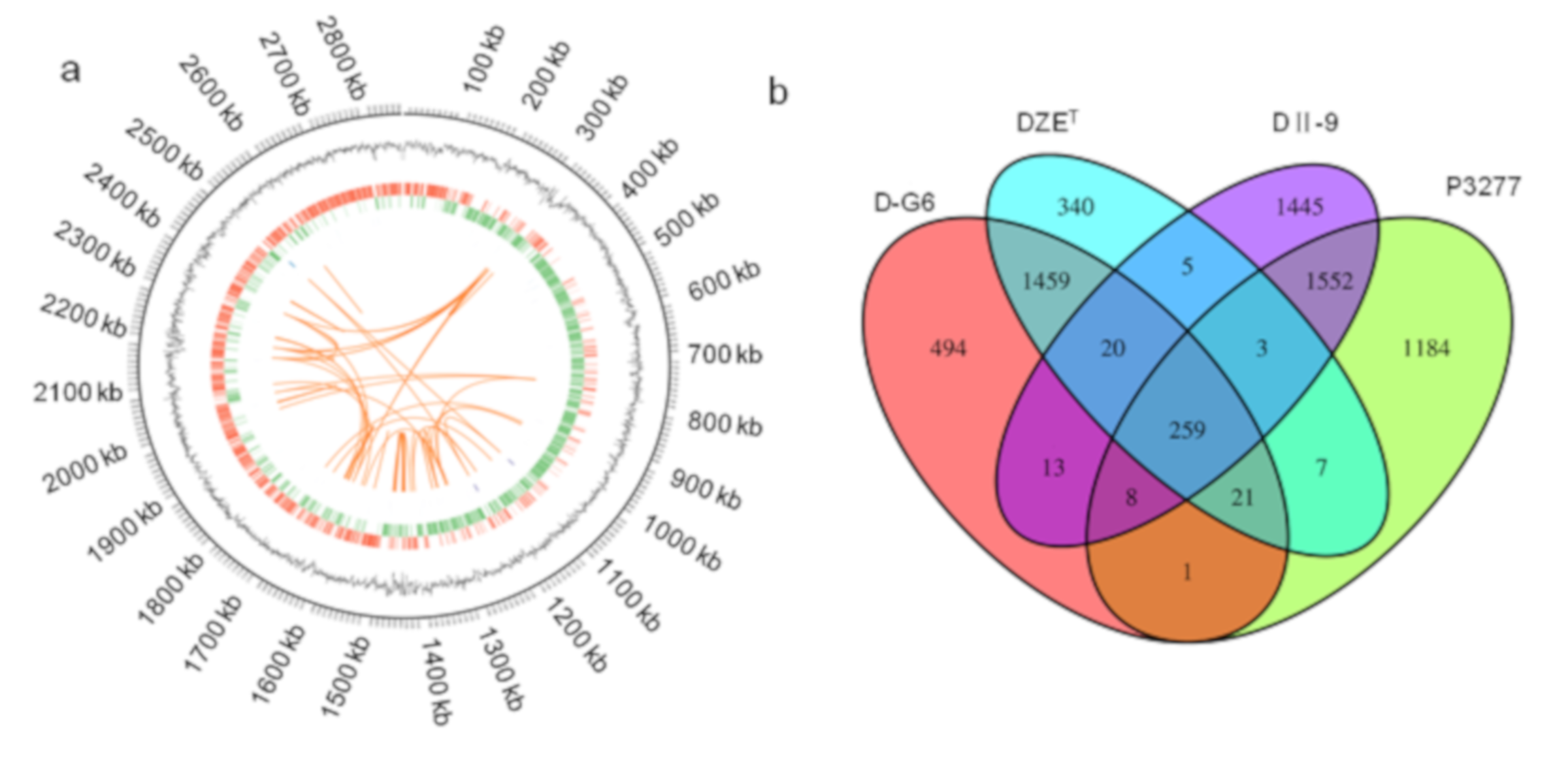
2.5. Characteristics of the D-G6 Genome and Genes Potentially Involved in DON De-Epoxidation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Bacterium Screening Methods
5.3. Extraction and Analysis of DON and DOM-1
5.4. Comparing the Transforming Rate under Different Culture Media, Temperatures and pH
5.5. Phylogenetic Tree Construction
5.6. Morphologic Analysis
5.7. Genome Sequencing
5.8. Drawing the Genome Circle Map
5.9. Comparing the Activity of Slackia sp. D-G6 and Slackia equolifaciens Strain DZET (DZET) on Transforming Daidzein (DZN)
6. Patents
Author Contributions
Funding
Conflicts of Interest
References
- Irakli, M.N.; Skendi, A.; Papageorgiou, M.D. HPLC-DAD-FLD Method for Simultaneous Determination of Mycotoxins in Wheat Bran. J. Chromatogr. Sci. 2017, 55, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: long-term analysis with special focus on Europe and Asia. J. Sci. Food Agr. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Neme, K.; Mohammed, A. Mycotoxin occurrence in grains and the role of postharvest management as a mitigation strategies. A review. Food Control 2017, 78, 412–425. [Google Scholar] [CrossRef]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- Wang, S.; Yang, J.; Zhang, B.; Wu, K.; Yang, A.; Li, C.; Zhang, J.; Zhang, C.; Rajput, S.A.; Zhang, N.; et al. Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets and IPEC-J2 Cells. Toxins 2018, 10, 541. [Google Scholar] [CrossRef] [Green Version]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Inter. J. Env. Res. Pub. Heal. 2017, 14, 632. [Google Scholar] [CrossRef] [Green Version]
- Uzarski, R.L.; Islam, Z.; Pestka, J.J. Potentiation of trichothecene-induced leukocyte cytotoxicity and apoptosis by TNF-alpha and Fas activation. Che. Biol. Interact. 2003, 146, 105–119. [Google Scholar] [CrossRef]
- Pestka, J.J.; Zhou, H.R.; Moon, Y.; Chung, Y.J. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: unraveling a paradox. Toxicol. letters 2004, 153, 61–73. [Google Scholar] [CrossRef]
- Cundliffe, E.; Cannon, M.; Davies, J. Mechanism of inhibition of eukaryotic protein synthesis by trichothecene fungal toxins. PNAS 1974, 71, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Cundliffe, E.; Davies, J.E. Inhibition of initiation, elongation, and termination of eukaryotic protein synthesis by trichothecene fungal toxins. Antimicrob. Agents Chemother. 1977, 11, 491–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.J.; Zhou, H.R.; Pestka, J.J. Transcriptional and posttranscriptional roles for p38 mitogen-activated protein kinase in upregulation of TNF-alpha expression by deoxynivalenol (vomitoxin). Toxicol. Appl. Pharmacol. 2003, 193, 188–201. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol-induced proinflammatory gene expression: mechanisms and pathological sequelae. Toxins 2010, 2, 1300–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, Y.; Pestka, J.J. Vomitoxin-induced cyclooxygenase-2 gene expression in macrophages mediated by activation of ERK and p38 but not JNK mitogen-activated protein kinases. Toxicol. Sci. 2002, 69, 373–382. [Google Scholar] [CrossRef]
- Katika, M.R.; Hendriksen, P.J.; Shao, J.; van Loveren, H.; Peijnenburg, A. Transcriptome analysis of the human T lymphocyte cell line Jurkat and human peripheral blood mononuclear cells exposed to deoxynivalenol (DON): New mechanistic insights. Toxicol. Appl. Pharmacol. 2012, 264, 51–64. [Google Scholar] [CrossRef]
- Schmeits, P.C.; Katika, M.R.; Peijnenburg, A.A.; van Loveren, H.; Hendriksen, P.J. DON shares a similar mode of action as the ribotoxic stress inducer anisomycin while TBTO shares ER stress patterns with the ER stress inducer thapsigargin based on comparative gene expression profiling in Jurkat T cells. Toxicol. letters 2014, 224, 395–406. [Google Scholar] [CrossRef]
- Bensassi, F.; Gallerne, C.; Sharaf El Dein, O.; Lemaire, C.; Hajlaoui, M.R.; Bacha, H. Involvement of mitochondria-mediated apoptosis in deoxynivalenol cytotoxicity. Food Chem. Toxicol. 2012, 50, 1680–1689. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, A.; Shi, Z.; He, C.; Ding, J.; Wang, X.; Ma, J.; Zhang, H. A mitochondria-mediated apoptotic pathway induced by deoxynivalenol in human colon cancer cells. Toxicol. 2012, 26, 414–420. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Matarese, F.; Sarrocco, S.; Gruber, S.; Seidl-Seiboth, V.; Vannacci, G. Biocontrol of Fusarium head blight: interactions between Trichoderma and mycotoxigenic Fusarium. Microbiology 2012, 158, 98–106. [Google Scholar] [CrossRef]
- Mandala, G.; Tundo, S.; Francesconi, S.; Gevi, F.; Zolla, L.; Ceoloni, C.; D’Ovidio, R. Deoxynivalenol Detoxification in Transgenic Wheat Confers Resistance to Fusarium Head Blight and Crown Rot Diseases. MPMI 2019, 32, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Dall’Asta, C.; Schuhmacher, R.; Lemmens, M.; Adam, G.; Krska, R. Masked mycotoxins: determination of a deoxynivalenol glucoside in artificially and naturally contaminated wheat by liquid chromatography-tandem mass spectrometry. J. Agr. Food Chem. 2005, 53, 3421–3425. [Google Scholar] [CrossRef] [PubMed]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Michlmayr, H.; Malachova, A.; Koutnik, A.; Maloku, I.; Berthiller, F.; Adam, G.; Krska, R.; et al. Biotransformation of the mycotoxin deoxynivalenol in fusarium resistant and susceptible near isogenic wheat lines. PloS One 2015, 10, e0119656. [Google Scholar] [CrossRef] [PubMed]
- Völkl, A.; Vogler, B.; Schollenberger, M.; Karlovsky, P. Microbial detoxifcation of mycotoxin deoxynivalenol. J. Basic. Microbiol. 2014, 44, 147–156. [Google Scholar]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M.; et al. Nocardioides sp. strain WSN05-2, isolated from a wheat feld, degrades deoxynivalenol, producing the novel intermediate 3-epi-deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Zhang, L.; Yi, S.; Tang, X.; Yuan, Q.; Guo, M.; Wu, A.; Qu, B.; Li, H.; Liao, Y. An aldo-keto reductase is responsible for fusarium toxin-degrading activity in a soil sphingomonas strain. Sci. Rep. 2017, 7, 9549. [Google Scholar] [CrossRef] [Green Version]
- Hassan, Y.I.; Lepp, D.; He, J.; Zhou, T. Draf genome sequences of Devosia sp. strain 17-2-E-8 and Devosia ribofavina strain IFO13584. Genome Announc. 2014, 2, e00994-14. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Sato, I.; Ishizaka, M.; Yoshida, S.-I.; Koitabashi, M.; Yoshida, S.; Tsushima, S. Bacterial cytochrome P450 system catabolizing the Fusarium toxin deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Sundstol Eriksen, G.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chemical toxicology: international J. published British Industrial Biol. Res. Assoc. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.; Schatzmayr, G.; He, J.W.; Zhou, T.; Moll, W.D.; et al. Microbial biotransformation of DON: molecular basis for reduced toxicity. Sci. Rep. 2016, 6, 29105. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Zhou, T.; Gong, J.; Young, C.; Su, X.; Li, X.Z.; Zhu, H.; Tsao, R.; Yang, R. Isolation of deoxynivalenol-transforming bacteria from the chicken intestines using the approach of PCR-DGGE guided microbial selection. BMC Microbiol. 2010, 10, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Mu, P.; Wen, J.; Sun, Y.; Chen, Q.; Deng, Y. Detoxification of trichothecene mycotoxins by a novel bacterium, Eggerthella sp. DII-9. Food Chem. Toxicol. 2018, 112, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Birt, D.F.; Hendrich, S.; Wang, W. Dietary agents in cancer prevention: flavonoids and isoflavonoids. Pharmacol. Therap. 2001, 90, 157–177. [Google Scholar] [CrossRef]
- Geller, S.E.; Studee, L. Soy and red clover for mid-life and aging. Climacteric 2006, 9, 245–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C.; Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef]
- Usui, T. Pharmaceutical prospects of phytoestrogens. Endocrine J. 2006, 53, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.S.; Kitahara, M.; Sakamoto, M.; Hattori, M.; Benno, Y. Slackia equolifaciens sp. nov., a human intestinal bacterium capable of producing equol. Int. J. Syst. Evol. Microbiol. 2009, 1721–1724. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, J.; Huang, L.; Chen, H.; Wang, C. Effects of Adding Clostridium sp. WJ06 on Intestinal Morphology and Microbial Diversity of Growing Pigs Fed with Natural Deoxynivalenol Contaminated Wheat. Toxins 2017, 9, 383. [Google Scholar] [CrossRef] [Green Version]
- Yanjing, X.; Xiumei, Y. Bioconversion of genistein to (-)-5-hydroxy-equol by a newly isolated cock intestinal anaerobic bacterium. J. Chin. Pharm. Sci. 2015, 24, 442–448. [Google Scholar] [CrossRef]
- Tsuji, H.; Moriyama, K.; Nomoto, K.; Miyanaga, N.; Akaza, H. Isolation and characterization of the equol-producing bacterium Slackia sp. strain NATTS. Arch. Microbiol. 2010, 192, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Matthies, A.; Blaut, M.; Braune, A. Isolation of a human intestinal bacterium capable of daidzein and genistein conversion. Appl. Environ. Microbiol. 2009, 75, 1740–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medjakovic, S.; Mueller, M.; Jungbauer, A. Potential health-modulating effects of isoflavones and metabolites via activation of PPAR and AhR. Nutrients 2010, 2, 241–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyn-Cook, B.D.; Stottman, H.L.; Yan, Y.; Blann, E.; Kadlubar, F.F.; Hammons, G.J. The effects of phytoestrogens on human pancreatic tumor cells in vitro. Cancer letters 1999, 142, 111–119. [Google Scholar] [CrossRef]
- Matthies, A.; Clavel, T.; Gutschow, M.; Engst, W.; Haller, D.; Blaut, M.; Braune, A. Conversion of daidzein and genistein by an anaerobic bacterium newly isolated from the mouse intestine. Appl. Environ. Microbiol. 2008, 74, 4847–4852. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhang, H.; Liu, G.; Sha, W. First report on the bacterial diversity in the distal gut of dholes (Cuon alpinus) by using 16S rRNA gene sequences analysis. J. Appl. Gene. 2016, 57, 275–283. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- McCarthy, A. Third generation DNA sequencing: pacific biosciences’ single molecule real time technology. Chem. Biol. 2010, 17, 675–676. [Google Scholar] [CrossRef] [Green Version]
- Myers, E.W.; Sutton, G.G.; Delcher, A.L.; Dew, I.M.; Fasulo, D.P.; Flanigan, M.J.; Kravitz, S.A.; Mobarry, C.M.; Reinert, K.H.; Remington, K.A.; et al. A whole-genome assembly of Drosophila. Science 2000, 287, 2196–2204. [Google Scholar] [CrossRef]
- Kawada, Y.; Yokoyama, S.; Yanase, E.; Niwa, T.; Suzuki, T. The production of S-equol from daidzein is associated with a cluster of three genes in Eggerthella sp. YY7918. Biosci. Microbio. Food Health 2016, 35, 113–121. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DON (μg/mL) | Time (d) | DOM-1/DON+DOM-1 (%) | |
|---|---|---|---|
| D-G6 | DⅡ-9 | ||
| 25 | 1 | 100 | 100 |
| 2 | 100 | 100 | |
| 75 | 1 | 60–70 | 80–100 |
| 2 | >95 | >95 | |
| 250 | 1 | 20–35 | 40–50 |
| 2 | >90 | >95 | |
| DZN (M) | Time (d) | D-G6 | DZET | ||||
|---|---|---|---|---|---|---|---|
| DHD (%) | DZN (%) | EQL (%) | DHD (%) | DZN (%) | EQL (%) | ||
| 0.2 | 2 | 5–30 | 0–10 | 60–100 | 0–18 | 0–6 | 89–100 |
| 3 | 0–5 | 0 | >97 | 0–6 | 0 | >90 | |
| 0.6 | 2 | 10–30 | 30–40 | 32–52 | 8–18 | 10–19 | 60–76 |
| 3 | 4–9 | <5 | >90 | 0 | <5 | >95 | |
| 2 | 2 | 3–15 | 68–80 | 10–19 | 2–10 | 60–78 | 14–25 |
| 3 | 8–15 | 20–35 | 50–60 | 5–10 | 25–40 | 55–65 | |
| D-G6 | DZET | |
|---|---|---|
| Size (Mb) | 2.86 | 2.75 |
| G + C (%) | 60.48 | 59.8 |
| Total number of protein-coding genes | 2312 | 2149 |
| Total number of rRNA genes | 29 | 10 |
| Total number of tRNA genes | 47 | 47 |
| Clusters | Predicted Functions | cds Number of D-G6 | cds Number of D II-9 |
|---|---|---|---|
| Cluster-761 | potassium-transporting ATPase activity | cds-765 | cds-727 |
| Cluster-892 | FAD-binding dehydrogenase | cds-1661 | cds-605 |
| Cluster-1703 | ISL3 family transposase | cds-1137/1145/1147/1172/1175 | cds-1991/1993 |
| Cluster-2351 | IS256 family transposase | cds-761/1023/1046 | cds-1169/1999 |
| Cluster-2392 | ring-opening amidohydrolase | cds-1648 | cds-1300 |
| Cluster-2459 | catalytic activity | iron-sulfur cluster binding | cds-1695 | cds-795 |
| Cluster-3749 | aminoglycoside 3′-phosphotransferase III | cds-1146 | cds-1992 |
| Cluster-3864 | aminoglycoside 3″-adenylyltransferase activity| response to antibiotic | cds-2270 | cds-1167 |
| Cluster-4013 | dimethyladenosine transferase | cds-1144 | cds-1932 |
| Cluster-4328 | protein of unknown function | cds-1907 | cds-1995 |
| Cluster-4382 | hypothetical protein | cds-2273 | cds-1837 |
| Cluster-6654 | isoleucyl-tRNA synthetase | cds-1205 | cds-928 |
| Cluster-6747 | protein of unknown function | cds-2271 | cds-1166 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Mu, P.; Zhu, X.; Chen, X.; Tang, S.; Wu, Y.; Miao, X.; Wang, X.; Wen, J.; Deng, Y. Dual Function of a Novel Bacterium, Slackia sp. D-G6: Detoxifying Deoxynivalenol and Producing the Natural Estrogen Analogue, Equol. Toxins 2020, 12, 85. https://doi.org/10.3390/toxins12020085
Gao X, Mu P, Zhu X, Chen X, Tang S, Wu Y, Miao X, Wang X, Wen J, Deng Y. Dual Function of a Novel Bacterium, Slackia sp. D-G6: Detoxifying Deoxynivalenol and Producing the Natural Estrogen Analogue, Equol. Toxins. 2020; 12(2):85. https://doi.org/10.3390/toxins12020085
Chicago/Turabian StyleGao, Xiaojuan, Peiqiang Mu, Xunhua Zhu, Xiaoxuan Chen, Shulin Tang, Yuting Wu, Xiang Miao, Xiaohan Wang, Jikai Wen, and Yiqun Deng. 2020. "Dual Function of a Novel Bacterium, Slackia sp. D-G6: Detoxifying Deoxynivalenol and Producing the Natural Estrogen Analogue, Equol" Toxins 12, no. 2: 85. https://doi.org/10.3390/toxins12020085
APA StyleGao, X., Mu, P., Zhu, X., Chen, X., Tang, S., Wu, Y., Miao, X., Wang, X., Wen, J., & Deng, Y. (2020). Dual Function of a Novel Bacterium, Slackia sp. D-G6: Detoxifying Deoxynivalenol and Producing the Natural Estrogen Analogue, Equol. Toxins, 12(2), 85. https://doi.org/10.3390/toxins12020085