Aflatoxin Biosynthesis and Genetic Regulation: A Review

 , ,
, ,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Aflatoxin Biosynthetic Pathway
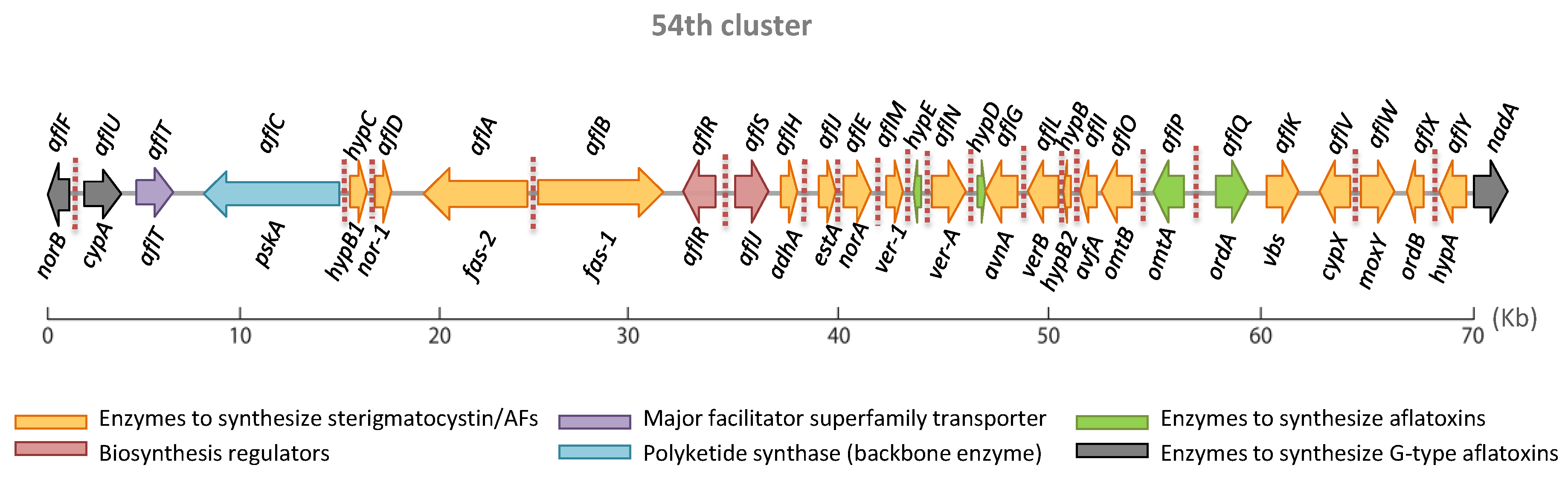
2.1. The Aflatoxin Gene Cluster
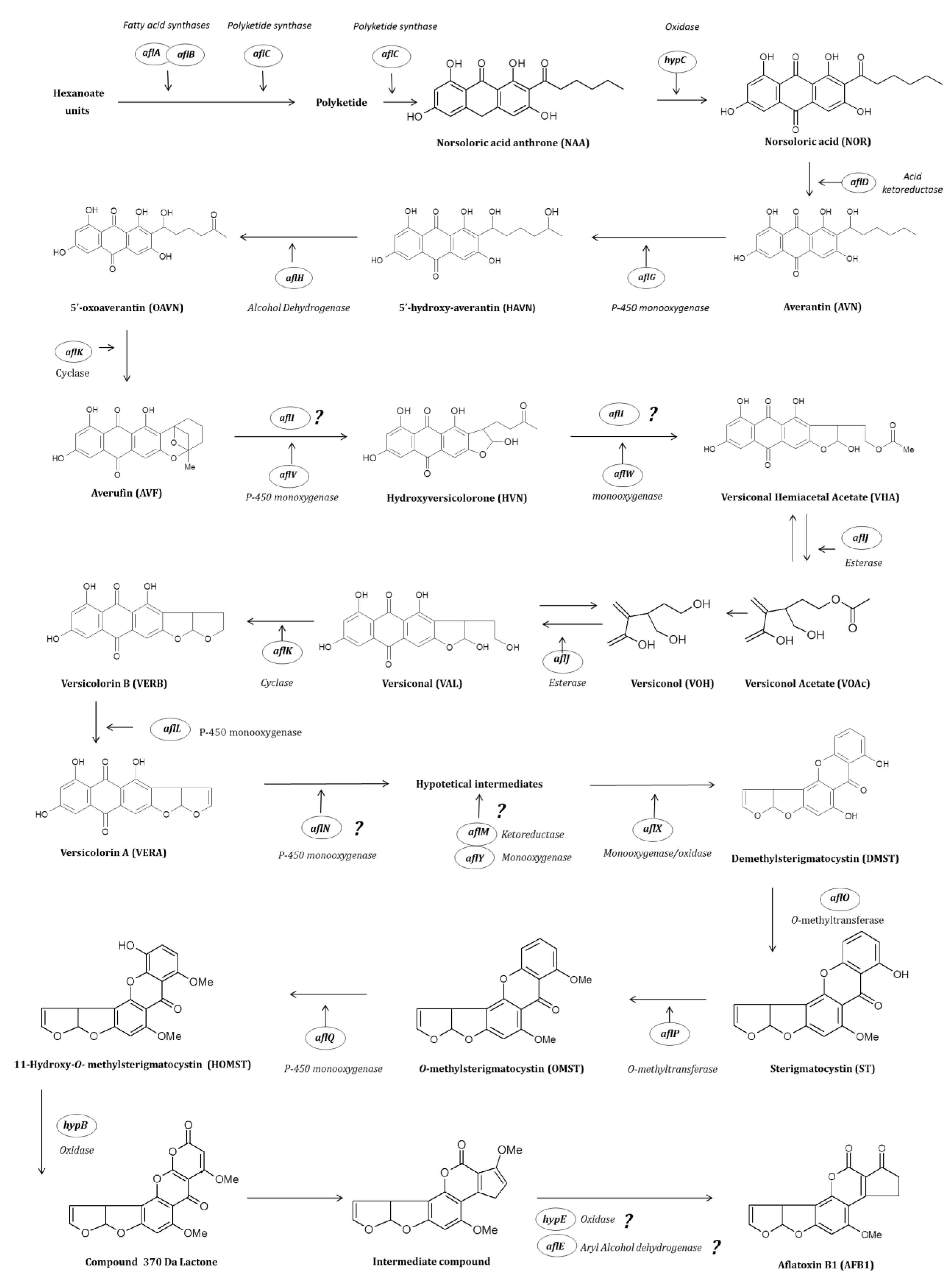
2.2. Enzymatic Cascade Leading to Aflatoxin Synthesis
- (i)
- Monooxygenases: responsible for incorporating one oxygen atom in another being reduced, with nicotinamide adenine dinucleotide phosphate (NADPH) acting as a cofactor.
- (ii)
- Dioxygenases: involved in ring-cleavage reactions.
- (iii)
- Baeyer-Villiger reactions: responsible of inserting oxygen atoms between two carbons.
2.3. Conversion of Acetate into Norsolorinic Acid
2.4. Conversion of Norsolorinic Acid into Versicolorin A
2.5. Conversion of Versicolorin A into Sterigmatocystin
2.6. Conversion of Sterigmatocystin into Aflatoxin B1
3. Genes Present in the AFB1 Cluster with an Unclear Role in the Aflatoxin Enzymatic Cascade
4. Aflatoxin Cluster-Specific Regulators
4.1. The aflR Transcription Factor
4.2. The aflS Transcription Enhancer
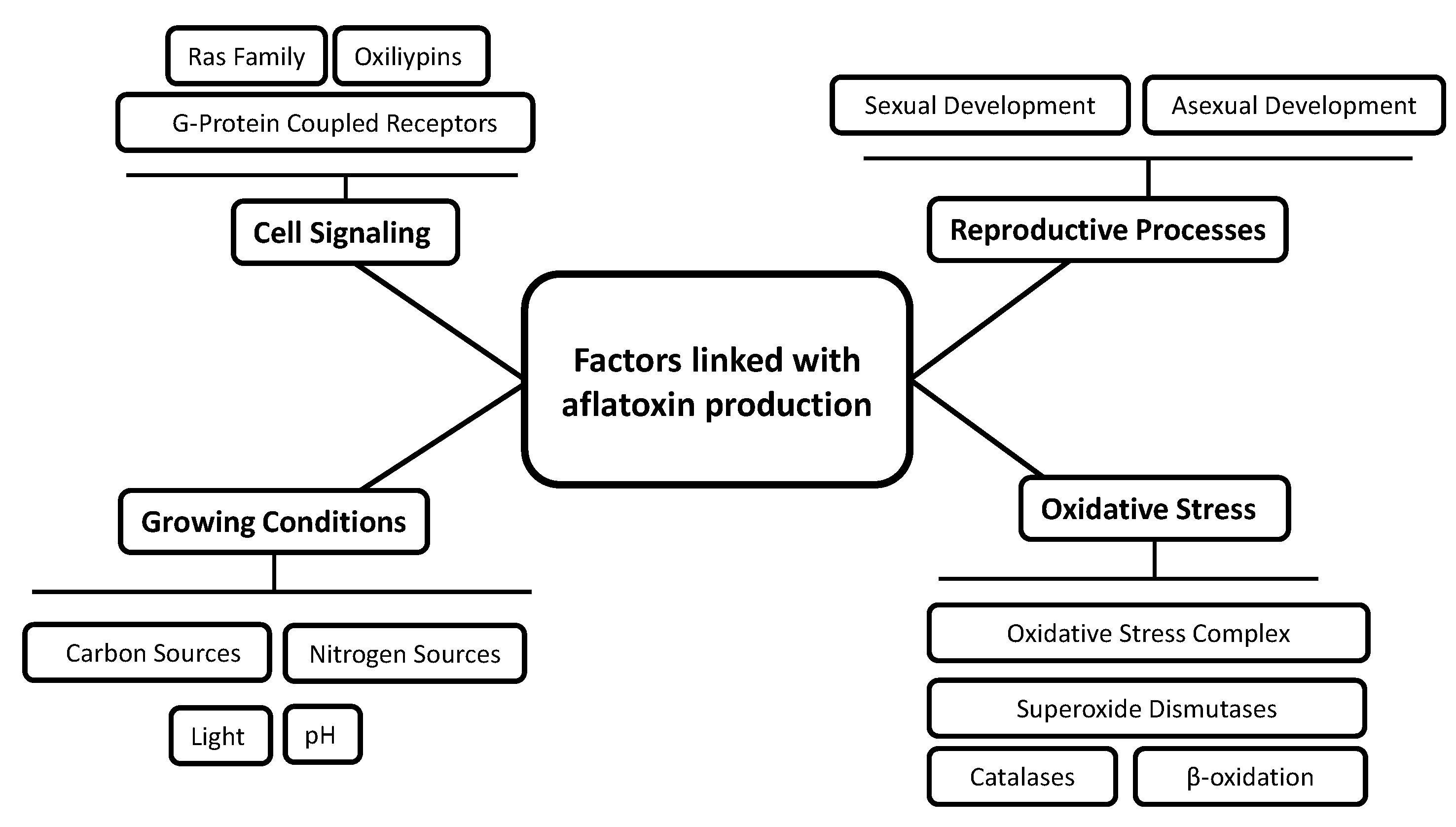
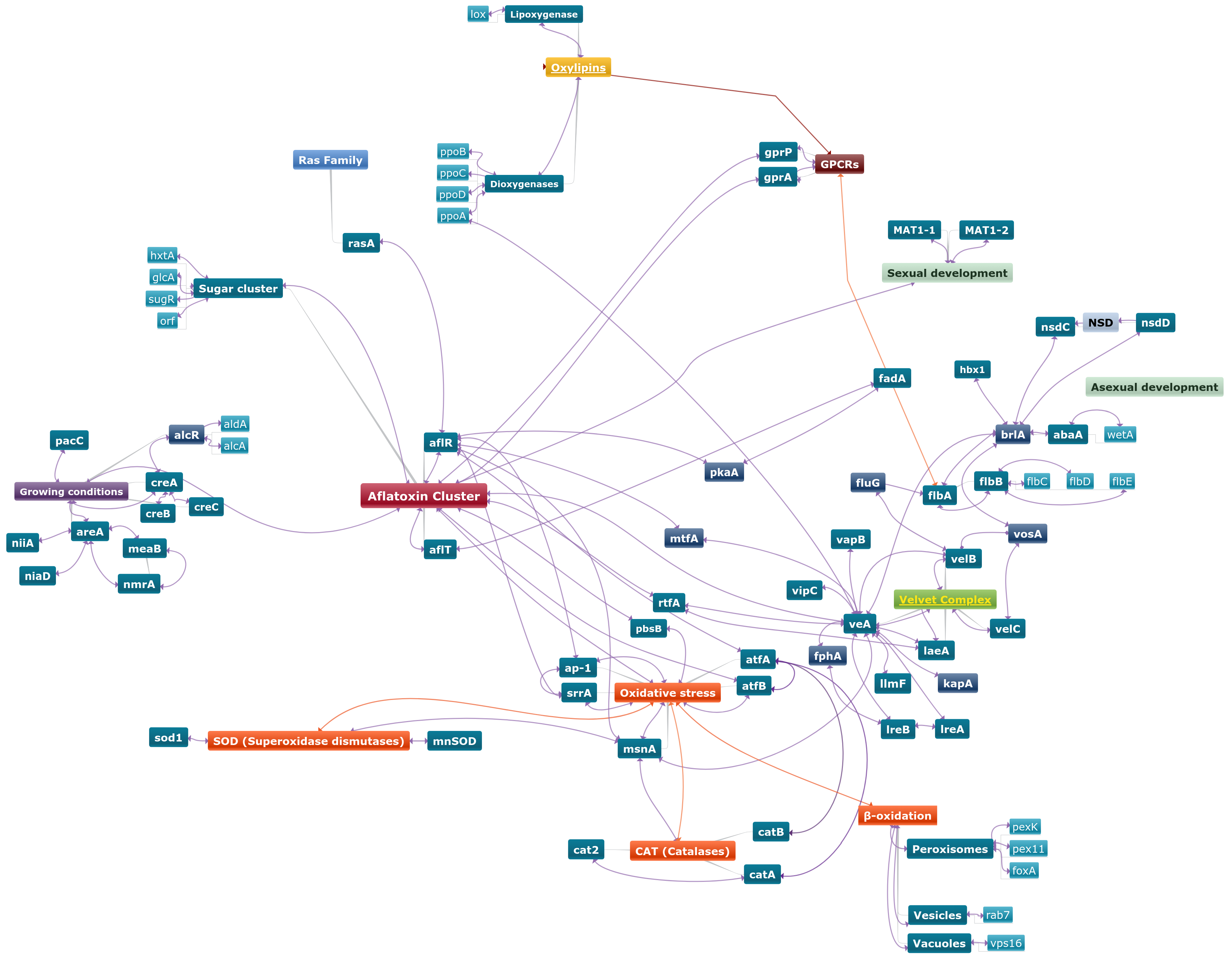
5. Global Regulation of Aflatoxin Production
5.1. Growing Conditions
5.1.1. Nutrient Sources
Carbon Source
Nitrogen Source
5.1.2. pH Conditions
5.1.3. Light
5.2. Reproductive Processes
5.2.1. Sexual Development
5.2.2. Asexual Development
5.3. Oxidative Stress
5.3.1. Superoxide Dismutases and Catalases
5.3.2. β-oxidation
5.4. Cell Signaling
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- Mahato, D.K.; Lee, K.E.; Kamle, M.; Devi, S.; Dewangan, K.N.; Kumar, P.; Kang, S.G. Aflatoxins in Food and Feed: An Overview on Prevalence, Detection and Control Strategies. Front. Microbiol. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Bbosa, G.S.; Kitya, D.; Lubega, A.; Ogwal-Okeng, J.; Anokbonggo, W.W.; Kyegombe, D.B. Review of the Biological and Health Effects of Aflatoxins on Body Organs and Body Systems. In Aflatoxin-Recent Advances and Future Prospects; Razzaghi, A., Ed.; InterchOpen: London, UK, 2013; Volume 12, pp. 239–265. [Google Scholar]
- Gauthier, T.; Duarte-Hospital, C.; Vignard, J.; Boutet-Robinet, E.; Sulyok, M.; Snini, S.P.; Alassane-Kpembi, I.; Lippi, Y.; Puel, S.; Oswald, I.P.; et al. Versicolorin A, a precursor in aflatoxins biosynthesis, is a food contaminant toxic for human intestinal cells. Environ. Int. 2020, 137, 105568. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Rodriguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B1 production. Front. Microbiol. 2014, 5, 348. [Google Scholar] [CrossRef] [PubMed]
- Van der Fels-Klerx, H.J.; Liu, C.; Battilani, P. Modelling climate change impacts on mycotoxin contamination. World Mycotoxin J. 2016, 9, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Bailly, S.; Mahgubi, A.E.; Carvajal, A.; Lorber, S.; Puel, O.; Oswald, I.P.; Bailly, J.D.; Orlando, B. Occurrence and Identification of Aspergillus Section Flavi in the Context of the Emergence of Aflatoxins in French Maize. Toxins 2018, 10, 525. [Google Scholar] [CrossRef] [Green Version]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.; Moretti, A.; Leggieri, M.C.; Rortais, B.A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [Green Version]
- Brankov, T.P.; Jovanovic, M.; Grujic, B. Aflatoxin Standards and Maize Trade. Econ. Agric. 2013, 3, 595–607. [Google Scholar]
- Sakuda, S.; Yoshinari, T.; Furukawa, T.; Jermnak, U.; Takagi, K.; Iimura, K.; Yamamoto, T.; Suzuki, M.; Nagasawa, H. Search for aflatoxin and trichothecene production inhibitors and analysis of their modes of action. Biosci. Biotechnol. Biochem. 2016, 80, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Hu, L.B.; Zhao, Y.; Wang, M.Y.; Zhang, H.; Mo, H.Z. Inhibition of fungal aflatoxin B1 biosynthesis by diverse botanically-derived polyphenols. Trop. J. Pharm. Res. 2015, 14, 605–609. [Google Scholar] [CrossRef] [Green Version]
- Jahanshiri, Z.; Shams-Ghahfarokhi, M.; Allameh, A.; Razzaghi-Abyaneh, M. Effect of curcumin on Aspergillus parasiticus growth and expression of major genes involved in the early and late stages of aflatoxin biosynthesis. Iran. J. Public Health 2012, 41, 72–79. [Google Scholar] [PubMed]
- Moon, Y.-S.; Choi, W.-S.; Park, E.-S.; Bae, I.; Choi, S.-D.; Paek, O.; Kim, S.-H.; Chun, H.; Lee, S.-E. Antifungal and Antiaflatoxigenic Methylenedioxy-Containing Compounds and Piperine-Like Synthetic Compounds. Toxins 2016, 8, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Khoury, R.; Caceres, I.; Puel, O.; Bailly, S.; Atoui, A.; Oswald, I.P.; El Khoury, A.; Bailly, J.D. Identification of the Anti-Aflatoxinogenic Activity of Micromeria graeca and Elucidation of Its Molecular Mechanism in Aspergillus flavus. Toxins 2017, 9, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef]
- Caceres, I.; Snini, S.; Puel, O.; Mathieu, F. Streptomyces roseolus, A Promising Biocontrol Agent Against Aspergillus flavus, the Main Aflatoxin B1 Producer. Toxins 2018, 10, 442. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Xing, F.; Selvaraj, J.N.; Liu, X.; Wang, L.; Hua, H.; Zhou, L.; Zhao, Y.; Wang, Y.; Liu, Y. Inhibitory Effect of Cinnamaldehyde, Citral, and Eugenol on Aflatoxin Biosynthetic Gene Expression and Aflatoxin B1 Biosynthesis in Aspergillus flavus. J. Food Sci. 2015, 80, M2917–M2924. [Google Scholar] [CrossRef]
- Caceres, I.; El Khoury, R.; Medina, Á.; Lippi, Y.; Naylies, C.; Atoui, A.; El Khoury, A.; Oswald, I.P.; Bailly, J.D.; Puel, O. Deciphering the Anti-Aflatoxinogenic Properties of Eugenol Using a Large-Scale q-PCR Approach. Toxins 2016, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Bhatnagar, D.; Ehrlich, K.C. Aflatoxin biosynthesis. Rev. Iberoam. Micol. 2002, 19, 191–200. [Google Scholar]
- Yabe, K.; Nakajima, H. Enzyme reactions and genes in aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2004, 64, 745–755. [Google Scholar] [CrossRef]
- Yu, J.; Ehrlich, K.C. Aflatoxin Biosynthetic Pathway and Pathway Genes. In Aflatoxins–Biochemistry and Molecular Biology; Guevara Gonzalez, R., Ed.; InterchOpen: London, UK, 2011; Volume 3, pp. 41–66. [Google Scholar]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [Green Version]
- Roze, L.V.; Hong, S.-Y.; Linz, J.E. Aflatoxin Biosynthesis: Current Frontiers. Annu. Rev. Food Sci. Technol. 2013, 4, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C. Predicted roles of the uncharacterized clustered genes in aflatoxin biosynthesis. Toxins 2009, 1, 37–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar] [CrossRef]
- Price, M.S.; Yu, J.; Nierman, W.C.; Kim, H.; Pritchard, B.; Jacobus, C.A.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiol. Lett. 2006, 255, 275–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klich, M.A. Environmental and developmental factors influencing aflatoxin production by Aspergillus flavus and Aspergillus parasiticus. Mycoscience 2007, 48, 71–80. [Google Scholar] [CrossRef]
- Montibus, M.; Pinson-Gadais, L.; Richard-Forget, F.; Barreau, C.; Ponts, N. Coupling of transcriptional response to oxidative stress and secondary metabolism regulation in filamentous fungi. Crit. Rev. Microbiol. 2013, 41, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Georgianna, D.R.; Payne, G.A. Genetic regulation of aflatoxin biosynthesis: From gene to genome. Fungal Genet. Biol. 2009, 46, 113–125. [Google Scholar] [CrossRef]
- Affeldt, K.J.; Carrig, J.; Amare, M.; Keller, N.P. Global Survey of Canonical Aspergillus flavus G Protein-Coupled Receptors. Am. Soc. Microbiol. 2014, 5, e01501–e01514. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.-H.; Keller, N. Regulation of secondary metabolism in filamentous fungi. Annu. Rev. Phytopathol. 2005, 43, 437–458. [Google Scholar] [CrossRef]
- Woloshuk, C.P.; Prieto, R. Genetic organization and function of the aflatoxin B1 biosynthetic genes. FEMS Microbiol. Lett. 1998, 160, 169–176. [Google Scholar] [CrossRef]
- Yu, J.; Chang, P.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered Pathway Genes in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trail, F.; Mahanti, N.; Linz, J. Molecular biology of aflatoxin biosynthesis. Microbiology 1995, 141, 755–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, J.H.; Choi, D.-K. Aflatoxins: Detection, Toxicity, and Biosynthesis. Biotechnol. Bioprocess Eng. 2007, 12, 585–593. [Google Scholar] [CrossRef]
- Payne, G.A.; Brown, M.P. Genetics and physiology of aflatoxin biosynthesis. Annu. Rev. Phytopathol. 1998, 36, 329–362. [Google Scholar] [CrossRef]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Yu, J.; Woloshuk, C.P.; Bhatnagar, D.; Cleveland, T.E. Cloning and characterization of avfA and omtB genes involved in aflatoxin biosynthesis in three Aspergillus species. Gene 2000, 248, 157–167. [Google Scholar] [CrossRef]
- Ehrlich, K.C.; Chang, P.-K.; Yu, J.; Cotty, P.J. Aflatoxin Biosynthesis Cluster Gene cypA Is Required for G Aflatoxin Formation. Appl. Environ. Microbiol. 2004, 70, 6518–6524. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, K.C.; Scharfenstein, L.L.; Montalbano, B.G.; Chang, P.K. Are the genes nadA and norB involved in formation of aflatoxin G1? Int. J. Mol. Sci. 2008, 9, 1717–1729. [Google Scholar] [CrossRef]
- Birch, A.L. Biosynthesis of polyketides and related compounds. Science 1967, 156, 202–206. [Google Scholar] [CrossRef]
- Minto, R.E.; Townsend, C.A. Enzymology and Molecular Biology of Aflatoxin Biosynthesis. Chem. Rev. 1997, 97, 2537–2556. [Google Scholar] [CrossRef]
- Dutton, M.F. Enzymes and aflatoxin biosynthesis. Microbiol. Rev. 1988, 52, 274–295. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Progress in tracing the evolutionary paths of cytochrome P450. Biochim. Biophys. Acta 2011, 1814, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Laivenieks, M.; Hong, S.Y.; Wee, J.; Wong, S.S.; Vanos, B.; Awad, D.; Ehrlich, K.C.; Linz, J.E. Aflatoxin biosynthesis is a novel source of reactive oxygen species-A potential redox signal to initiate resistance to oxidative stress? Toxins 2015, 7, 1411–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabe, K.; Ando, Y.; Hamasaki, T. Desaturase Activity in the Branching Step Between Aflatoxins B1 And G1 and Aflatoxins B2 and G2. Agric. Biol. Chem. 1991, 55, 1907–1911. [Google Scholar] [CrossRef]
- Sakuno, E.; Wen, Y.; Hatabayashi, H.; Arai, H.; Aoki, C.; Yabe, K.; Nakajima, H. Aspergillus parasiticus Cyclase Catalyzes Two Dehydration Steps in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2005, 71, 2999–3006. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Hatabayashi, H.; Arai, H.; Kitamoto, H.K.; Yabe, K. Function of the cypX and moxY Genes in Aflatoxin Biosynthesis in Aspergillus parasiticus. Appl. Environ. Microbiol. 2005, 71, 3192–3198. [Google Scholar] [CrossRef] [Green Version]
- Yabe, K.; Chihaya, N.; Hamamatsu, S.; Sakuno, E.; Hamasaki, T.; Nakajima, H.; Bennett, J.W. Enzymatic conversion of averufin to hydroxyversicolorone and elucidation of a novel metabolic grid involved in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2003, 69, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Sakuno, E.; Yabe, K.; Nakajima, H. Involvement of Two Cytosolic Enzymes and a Novel Intermediate, 5′-Oxoaverantin, in the Pathway from 5′-Hydroxyaverantin to Averufin in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2003, 69, 6418–6426. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Yabe, K.; Yu, J. The Aspergillus parasiticus estA-encoded esterase converts versiconal hemiacetal acetate to versiconal and versiconol acetate to versiconol in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 3593–3599. [Google Scholar] [CrossRef] [Green Version]
- Henry, K.M.; Townsend, C.A. Ordering the reductive and cytochrome P450 oxidative steps in demethylsterigmatocystin formation yields general insights into the biosynthesis of aflatoxin and related fungal metabolites. J. Am. Chem. Soc. 2005, 127, 3724–3733. [Google Scholar] [CrossRef]
- Cary, J.W.; Ehrlich, K.C.; Bland, J.M.; Montalbano, B.G. The aflatoxin biosynthesis cluster gene, aflX, encodes an oxidoreductase involved in conversion of versicolorin a to demethylsterigmatocystin. Appl. Environ. Microbiol. 2006, 72, 1096–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, K.C.; Montalbano, B.; Boué, S.M.; Bhatnagar, D. An aflatoxin biosynthesis cluster gene encodes a novel oxidase required for conversion of versicolorin A to sterigmatocystin. Appl. Environ. Microbiol. 2005, 71, 8963–8965. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Hatabayashi, H.; Nakagawa, H.; Cai, J.; Suzuki, R.; Sakuno, E.; Tanaka, T.; Ito, Y.; Ehrlich, K.C.; Nakajima, H.; et al. Conversion of 11-hydroxy-O methylsterigmatocystin to Aflatoxin G1 in Aspergillus parasiticus. Appl. Microbiol. Biotechnol. 2011, 90, 635–650. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Bhatnagar, D.; Cleveland, T.E. Completed sequence of aflatoxin pathway gene cluster in Aspergillus parasiticus. FEBS Lett. 2004, 564, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, K.C.; Li, P.; Scharfenstein, L.; Chang, P.K. HypC, the Anthrone Oxidase Involved in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2010, 76, 3374–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Linz, J.E. Enzymatic function of the Nor-1 protein in aflatoxin biosynthesis in Aspergillus parasiticus. Appl. Environ. Microbiol. 1999, 65, 5639–5641. [Google Scholar] [CrossRef] [Green Version]
- Yabe, K.; Nakamura, Y.; Nakajima, H.; Ando, Y. Enzymatic Conversion of Norsolorinic Acid to Averufin in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 1991, 57, 1340–1345. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chang, P.K.; Bhatnagar, D.; Cleveland, T.E. avnA, Encoding A Putative P-450 Monooxygenase, is Involved in the Conversion of Averantin to Averufin in Aflatoxin Biosynthesis in Aspergillus parasiticus. Appl. Environ. Microbiol. 1997, 63, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Montalbano, B.G.; Ehrlich, K.C. Promoter Elements Involved in The Expression of the Aspergillus parasiticus Aflatoxin Pathway Gene avnA. Biochim. Biophys. Acta 2000, 1491, 7–12. [Google Scholar] [CrossRef]
- Chang, P.K.; Yu, J.; Ehrlich, K.C.; Boue, S.M.; Montalbano, B.G.; Bhatnagar, D.; Cleveland, T.E. adhA in Aspergillus parasiticus is involved in conversion of 5′-hydroxyaverantin to averufin. Appl. Environ. Microbiol. 2000, 66, 4715–4719. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chang, P.K.; Bhatnagar, D.; Cleveland, T.E. Cloning and functional expression of an esterase gene in Aspergillus parasiticus. Mycopathologia 2002, 156, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.K.; Anderson, J.A. Purification and properties of versiconal cyclase from Aspergillus parasiticus. Arch. Biochem. Biophys. 1992, 193, 67–70. [Google Scholar] [CrossRef]
- Kelkar, H.S.; Skloss, T.W.; Haw, J.F.; Keller, N.P.; Adams, T.H. Aspergillus nidulans stcL encodes a putative cytochrome P-450 monooxygenase required for bisfuran desaturation during aflatoxin/sterigmatocystin biosynthesis. J. Biol. Chem. 1997, 272, 1589–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, N.P.; Kantz, N.J.; Adams, T.H. Aspergillus nidulans verA is required for production of the mycotoxin sterigmatocystin. Appl. Environ. Microbiol. 1994, 60, 1444–1450. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.Y.; Roze, L.V.; Wee, J.; Linz, J.E. Evidence that a transcription factor regulatory network coordinates oxidative stress response and secondary metabolism in Aspergilli. Microbiologyopen 2013, 2, 144–160. [Google Scholar] [CrossRef]
- Keller, N.P.; Segner, S.; Bhatnagar, D.; Adams, T.H. stcS, a putative P-450 monooxygenase, is required for the conversion of versicolorin A to sterigmatocystin in Aspergillus nidulans. Appl. Environ. Microbiol. 1995, 61, 3628–3632. [Google Scholar] [CrossRef] [Green Version]
- Yabe, K.; Ando, Y.; Hashimoto, J.; Hamasaki, T. Two distinct O-methyltransferases in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 1989, 55, 2172–2177. [Google Scholar] [CrossRef] [Green Version]
- Scherm, B.; Palomba, M.; Serra, D.; Marcello, A.; Migheli, Q. Detection of transcripts of the aflatoxin genes aflD, aflO, and aflP by reverse transcription-polymerase chain reaction allows differentiation of aflatoxin-producing and non-producing isolates of Aspergillus flavus and Aspergillus parasiticus. Int. J. Food Microbiol. 2005, 98, 201–210. [Google Scholar] [CrossRef]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Montalbano, B.; Dyer, J.M.; Bhatnagar, D.; Cleveland, T.E. Characterization of the Critical Amino Acids of an Aspergillus parasiticus Cytochrome P-450 Monooxygenase Encoded by ordA that is Involved in the Biosynthesis of Aflatoxins B1, G1, B2, and G2. Appl. Environ. Microbiol. 1998, 64, 4834–4841. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Wright, M.; Bhatnagar, D.; Lee, R.; Chu, F.S. Molecular Characterization of an Aspergillus parasiticus dehydrogenase gene, norA, located on the Aflatoxin Biosynthesis Gene. Appl. Environ. Microbiol. 1996, 62, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, K.C.; Chang, P.-K.; Scharfenstein, L.L.; Cary, J.W.; Crawford, J.M.; Townsend, C.A. Absence of the aflatoxin biosynthesis gene, norA, allows accumulation of deoxyaflatoxin B1 in Aspergillus flavus cultures. FEMS Microbiol. Lett. 2010, 305, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Yu, J.; Yu, J.H. aflT, a MFS transporter-encoding gene located in the aflatoxin gene cluster, does not have a significant role in aflatoxin secretion. Fungal Genet. Biol. 2004, 41, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Roze, L.V.; Linz, J.E. A Possible Role for Exocytosis in Aflatoxin Export in Aspergillus parasiticus. Eukaryot. Cell 2010, 9, 1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.-H.; Butchko, R.E.; Fernandes, M.; Keller, N.; Leonard, T.; Adams, T. Conservation of structure and function of the aflatoxin regulatory gene aflR from Aspergillus nidulans and A. flavus. Curr. Genet. 1996, 29, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Hicks, J.K.; Huang, T.P.; Keller, N.P. Pka, Ras and RGS Protein Interactions Regulate Activity of AflR, a Zn(II)2Cys6 Transcription Factor in Aspergillus nidulans. Genetics 2003, 165, 1095–1104. [Google Scholar] [PubMed]
- MacPherson, S.; Larochelle, M.; Turcotte, B. A fungal family of transcriptional regulators: The zinc cluster proteins. Microbiol. Mol. Biol. Rev. 2006, 70, 583–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woloshuk, C.P.; Foutz, K.R.; Brewer, J.F.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A. Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Appl. Environ. Microbiol. 1994, 60, 2408–2414. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, K.C.; Montalbano, B.G.; Cary, J.W. Binding of the C6-zinc cluster protein, AFLR, to the promoters of aflatoxin pathway biosynthesis genes in Aspergillus parasiticus. Gene 1999, 230, 249–257. [Google Scholar] [CrossRef]
- Chang, P.-K.; Ehrlich, K.C.; Yu, J.; Bhatnagar, D.; Cleveland, T.E. Increased expression of Aspergillus parasiticus aflR, encoding a sequence- specific DNA-binding protein, relieves nitrate inhibition of aflatoxin biosynthesis. Appl. Environ. Microbiol. 1995, 61, 2372–2377. [Google Scholar] [CrossRef] [Green Version]
- Flaherty, J.E.; Payne, G.A. Overexpression of aflR Leads to Upregulation of Pathway Gene Transcription and Increased Aflatoxin Production in Aspergillus flavus. Appl. Environ. Microbiol. 1997, 63, 3995–4000. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Ehrlich, K.C.; Wright, M.; Chang, P.K.; Bhatnagar, D. Generation of aflR disruption mutants of Aspergillus parasiticus. Appl. Microbiol. Biotechnol. 2000, 53, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Dhanamjayulu, P.; Boga, R.B.; Mehta, A. Inhibition of aflatoxin B1 biosynthesis and down regulation of aflR and aflB genes in presence of benzimidazole derivatives without impairing the growth of Aspergillus flavus. Toxicon 2019, 170, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Jahanshiri, Z.; Shams-Ghahfarokhi, M.; Allameh, A.; Razzaghi-Abyaneh, M. Inhibitory effect of eugenol on aflatoxin B1 production in Aspergillus parasiticus by downregulating the expression of major genes in the toxin biosynthetic pathway. World J. Microbiol. Biotechnol. 2015, 31, 1071–1078. [Google Scholar] [CrossRef]
- Jermnak, U.; Yoshinari, T.; Sugiyama, Y.; Tsuyuki, R.; Nagasawa, H.; Sakuda, S. Isolation of methyl syringate as a specific aflatoxin production inhibitor from the essential oil of Betula alba and aflatoxin production inhibitory activities of its related compounds. Int. J. Food Microbiol. 2012, 153, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Meyers, D.M.; Obrian, G.; Du, W.L.; Bhatnagar, D.; Payne, G.A. Characterization of aflJ, a gene required for conversion of pathway intermediates to aflatoxin. Appl. Environ. Microbiol. 1998, 64, 3713–3717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.; Obrian, G.R.; Payne, G.A. Function and regulation of aflJ in the accumulation of aflatoxin early pathway intermediate in Aspergillus flavus. Food Addit. Contam. 2007, 24, 1043–1050. [Google Scholar] [CrossRef]
- Kong, Q.; Chi, C.; Yu, J.; Shan, S.; Li, Q.; Li, Q.; Guan, B.; Nierman, W.C.; Bennett, J.W. The inhibitory effect of Bacillus megaterium on aflatoxin and cyclopiazonic acid biosynthetic pathway gene expression in Aspergillus flavus. Appl. Environ. Microbiol. 2014, 98, 5161–5172. [Google Scholar] [CrossRef]
- Yoshinari, T.; Akiyama, T.; Nakamura, K.; Kondo, T.; Takahashi, Y.; Muraoka, Y.; Nonomura, Y.; Nagasawa, H.; Sakuda, S. Dioctatin A is a strong inhibitor of aflatoxin production by Aspergillus parasiticus. Microbiology 2007, 153, 2774–2780. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Solfrizzo, M.; Epifani, F.; Panzarini, G.; Perrone, G. Effect of temperature and water activity on gene expression and aflatoxin biosynthesis in Aspergillus flavus on almond medium. Int. J. Food Microbiol. 2016, 217, 162–169. [Google Scholar] [CrossRef]
- Gizachew, D.; Chang, C.H.; Szonyi, B.; De La Torre, S.; Ting, W.E. Aflatoxin B1 (AFB1) production by Aspergillus flavus and Aspergillus parasiticus on ground Nyjer seeds: The effect of water activity and temperature. Int. J. Food Microbiol. 2019, 296, 8–13. [Google Scholar] [CrossRef]
- Deepika, V.B.; Murali, T.S.; Satyamoorthy, K. Modulation of genetic clusters for synthesis of bioactive molecules in fungal endophytes: A review. Microbiol. Res. 2015, 182, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.D.; Diener, U.L. Growth and Aflatoxin Production by Aspergillus parasiticus from Various Carbon Sources. Appl. Microbiol. 1968, 16, 158–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggio-Hall, L.A.; Wilson, R.A.; Keller, N.P. Fundamental contribution of beta-oxidation to polyketide mycotoxin production in planta. Mol. Plant Microbe Interact. 2005, 18, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatnagar, D.; Cary, J.W.; Ehrlich, K.; Yu, J.; Cleveland, T.E. Understanding the genetics of regulation of aflatoxin production and Aspergillus flavus development. Mycopathologia 2006, 162, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between Secondary Metabolism and Fungal Development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sun, L.; Zhang, N.; Zhang, J.; Guo, J.; Li, C.; Rajput, S.A.; Qi, D. Effects of Nutrients in Substrates of Different Grains on Aflatoxin B1 Production by Aspergillus flavus. BioMed Res. Int. 2016, 2016, 1–10. [Google Scholar]
- Zhang, J.-D.; Han, L.; Yan, S.; Liu, C.-M. The non-metabolizable glucose analog D-glucal inhibits aflatoxin biosynthesis and promotes kojic acid production in Aspergillus flavus. BMC Microbiol. 2014, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chang, P.K.; Bhatnagar, D.; Cleveland, T.E. Cloning of a Sugar Utilization Gene Cluster in Aspergillus parasiticus. Biochim. Biophys. Acta 2000, 1493, 211–214. [Google Scholar] [CrossRef]
- Ruijter, G.J.G.; Visser, J. Carbon repression in Aspergilli. FEMS Microbiol. Lett. 1997, 151, 103–114. [Google Scholar] [CrossRef]
- Fasoyin, O.E.; Wang, B.; Qiu, M.; Han, X.; Chung, K.R.; Wang, S. Carbon catabolite repression gene creA regulates morphology, aflatoxin biosynthesis and virulence in Aspergillus flavus. Fungal Genet. Biol. 2018, 115, 41–51. [Google Scholar] [CrossRef]
- Shroff, R.A.; Lockington, R.A.; Kelly, J.M. Analysis of mutations in the creA gene involved in carbon catabolite repression in Aspergillus nidulans. Can. J. Microbiol. 1996, 42, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Ries, L.N.A.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldma, G.H. Diverse Regulation of the CreA Carbon Catabolite Repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowzer, C.E.A.; Kelly, J.M. Cloning of the Crea gene from Aspergillus nidulans–A gene involved in carbon catabolite repression. Curr. Genet. 1989, 15, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhi, Q.-Q.; Li, J.-Y.; Keller, N.; He, Z.-M. The Antioxidant Gallic Acid Inhibits Aflatoxin Formation in Aspergillus flavus by Modulating Transcription Factors FarB and CreA. Toxins 2018, 10, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, J.R.; Yu, J.; Abbas, H.K.; Scheffler, B.E.; Kim, H.S.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E. Aflatoxin formation and gene expression in response to carbon source media shift in Aspergillus parasiticus. Food Addit. Contam. 2007, 24, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Kudla, B.; Caddick, M.X.; Langdon, T.; Martinez-Rossi, N.M.; Bennett, C.F.; Sibley, S.; Davies, R.W.; Arst, H.N. The regulatory gene areA mediating nitrogen metabolite repression in Aspergillus nidulans. Mutations affecting specificity of gene activation alter a loop residue of a putative zinc finger. EMBO J. 1990, 9, 1355–1364. [Google Scholar] [CrossRef]
- Amaike, S.; Affeldt, K.J.; Yin, W.B.; Franke, S.; Choithani, A.; Keller, N.P. The bZIP Protein MeaB Mediates Virulence Attributes in Aspergillus flavus. PLoS ONE 2013, 8, e74030. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Yu, J.H.; Bhatnagar, D.; Cleveland, T.E. Characterization of the Aspergillus parasiticus Major Nitrogen Regulatory Gene, areA. Biochim. Biophys. Acta 2000, 1491, 263–266. [Google Scholar] [CrossRef]
- Fasoyin, O.E.; Yang, K.; Qiu, M.; Wang, B.; Wang, S.; Wang, S. Regulation of Morphology, Aflatoxin Production, and Virulence of Aspergillus flavus by the Major Nitrogen Regulatory Gene areA. Toxins 2019, 11, 718. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, K.C.; Cotty, P.J. Variability in Nitrogen Regulation of Aflatoxin Production by Aspergillus flavus Strains. Microbiol. Biotechnol. 2002, 60, 174–178. [Google Scholar]
- Ehrlich, K.C.; Cary, J.W.; Montalbano, B.G. Characterization of the promoter for the gene encoding the aflatoxin biosynthetic pathway regulatory protein AFLR. Biochim. Biophys. Acta Gene Struct. Expr. 1999, 1444, 412–417. [Google Scholar] [CrossRef]
- Feng, G.H.; Leonard, T.J. Culture Conditions Control Expression Of The Genes For Aflatoxin and Sterigmatocystin Biosynthesis in Aspergillus parasiticus and A. nidulans. Appl. Environ. Microbiol. 1998, 64, 2275–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Ren, H.; Mousa, J.J.; Rangel, D.E.; Zhang, Y.; Bruner, S.D.; Keyhani1ing, N.O. The PacC transcription factor regulates secondary metabolite production, stress response, but has only minor effects on virulence in the insect pathogenic fungus Beauveria bassiana. Environ. Microbiol. 2017, 19, 788–802. [Google Scholar] [CrossRef]
- Selvig, K.; Alspaugh, J.A. pH Response Pathways in Fungi: Adapting to Host-derived and Environmental Signals. Mycobiology 2011, 39, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilburn, J.; Sarkar, S.; Widdick, D.A.; Espeso, E.A.; Orejas, M.; Mungroo, J.; Peñalva, M.A.; Arst, H.N. The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid- and alkaline-expressed genes by ambient pH. EMBO J. 1995, 14, 779–790. [Google Scholar] [CrossRef]
- Cornet, M.; Gaillardin, C. pH signaling in human fungal pathogens: A new target for antifungal strategies. Eukaryot. Cell 2014, 13, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Cotty, P.J. Aflatoxin and Sclerotial Production by Aspergillus flavus: Influence of pH. Physiol. Biochem. 1988, 78, 1250–1253. [Google Scholar]
- Buchanan, R.L.; Ayres, J.C. Effects of initial pH on aflatoxin production. Appl. Microbiol. 1975, 30, 1050–1051. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P.; Nesbitt, C.; Sarr, B.; Phillips, T.D.; Burow, G.B. pH Regulation of Sterigmatocystin and Aflatoxin Biosynthesis in Aspergillus spp. Phytopathology 1997, 87, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Rangel, D.E.N.; Alder-Rangel, A.; Dadachova, E.; Finlay, R.D.; Kupiec, M.; Dijksterhuis, J.; Braga, G.U.L.; Corrochano, L.M.; Hallsworth, J.E. Fungal stress biology: A preface to the Fungal Stress Responses special edition. Curr. Genet. 2015, 61, 231–238. [Google Scholar] [CrossRef]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Purschwitz, J.; Müller, S.; Kastner, C.; Schöser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and Physical Interaction of Blue- and Red-Light Sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.-J.; Keller, N.P.; Yu, J.-H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya-Bayram, O.; Palmer, J.M.; Keller, N.; Braus, G.H.; Bayram, O. One Juliet and four Romeos: VeA and its methyltransferases. Front. Microbiol. 2015, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayram, Ö.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, Y.L.; Gerke, J.; Park, H.S.; Bayram, O.; Neumann, P.; Ni, M.; Dickmanns, A.; Kim, S.C.; Yu, J.H.; Braus, G.H.; et al. The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-kB. PLoS Biol. 2013, 11, e1001750. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.M.; Theisen, J.M.; Duran, R.M.; Grayburn, W.S.; Calvo, A.M.; Keller, N.P. Secondary Metabolism and Development Is Mediated by LlmF Control of VeA Subcellular Localization in Aspergillus nidulans. PLoS Genet. 2013, 9, e1003193. [Google Scholar] [CrossRef] [Green Version]
- Purschwitz, J.; Muller, S.; Fischer, R. Mapping the interaction sites of Aspergillus nidulans phytochrome FphA with the global regulator VeA and the White Collar protein LreB. Mol. Genet. Genom. 2009, 281, 35–42. [Google Scholar] [CrossRef]
- Sarikaya-Bayram, O.; Bayram, O.; Feussner, K.; Kim, J.H.; Kim, H.S.; Kaever, A.; Feussner, I.; Chae, K.S.; Han, D.M.; Han, K.H.; et al. Membrane-bound methyltransferase complex VapA-VipC-VapB guides epigenetic control of fungal development. Dev. Cell 2014, 29, 406–420. [Google Scholar] [CrossRef] [Green Version]
- Duran, R.M.; Cary, J.W.; Calvo, A.M. Production of cyclopiazonic acid, aflatrem, and aflatoxin by Aspergillus flavus is regulated by veA, a gene necessary for sclerotial formation. Appl. Microbiol. Biotechnol. 2007, 73, 1158–1168. [Google Scholar] [CrossRef]
- Duran, R.M.; Cary, J.W.; Calvo, A.M. The Role of veA in Aspergillus flavus Infection of Peanut, Corn and Cotton. Open Mycol. J. 2009, 3, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Amaike, S.; Keller, N.P. Distinct roles for VeA and LaeA in development and pathogenesis of Aspergillus flavus. Eukaryot. Cell 2009, 8, 1051–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.K.; Scharfenstein, L.L.; Ehrlich, K.C.; Wei, Q.; Bhatnagar, D.; Ingber, B.F. Effects of laeA deletion on Aspergillus flavus conidial development and hydrophobicity may contribute to loss of aflatoxin production. Fungal Biol. 2012, 116, 298–307. [Google Scholar] [CrossRef]
- Cary, J.W.; Han, Z.; Yin, Y.; Lohmar, J.M.; Shantappa, S.; Harris-Coward, P.Y.; Mack, B.; Ehrlich, K.C.; Wei, Q.; Arroyo-Manzanares, N.; et al. Transcriptome Analysis of Aspergillus flavus Reveals veA -Dependent Regulation of Secondary Metabolite Gene Clusters, Including the Novel Aflavarin Cluster. Eukaryot. Cell 2015, 14, 983–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, A.M.; Bok, J.W.; Brooks, W.; Keller, N.P. Vea Is Required for Toxin And Sclerotial Production In Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 4733–4739. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R. Developmental biology. Sex and poison in the dark. Science 2008, 320, 1430–1431. [Google Scholar] [CrossRef]
- Kato, N.; Brooks, W.; Calvo, A.M. The Expression of Sterigmatocystin and Penicillin Genes in Aspergillus nidulans is controlled by veA, a gene required for Sexual Development. Eukaryot. Cell 2003, 2, 1178–1186. [Google Scholar] [CrossRef] [Green Version]
- Bok, J.W.; Keller, N.P. LaeA, a Regulator of Secondary Metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Atoui, A.; Kastner, C.; Larey, C.M.; Thokala, R.; Etxebeste, O.; Espeso, E.A.; Fischer, R.; Calvo, A.M. Cross-talk between light and glucose regulation controls toxin production and morphogenesis in Aspergillus nidulans. Fungal Genet. Biol. 2010, 47, 962–972. [Google Scholar] [CrossRef] [Green Version]
- Perrone, G.; Gallo, A.; Logrieco, A.F. Biodiversity of Aspergillus section Flavi in Europe in relation to the management of aflatoxin risk. Front. Microbiol. 2014, 5, 377. [Google Scholar] [CrossRef] [Green Version]
- Horn, B.W.; Moore, G.G.; Carbone, I. Sexual reproduction in Aspergillus flavus. Mycologia 2009, 101, 423–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, B.W.; Sorensen, R.B.; Lamb, M.C.; Sobolev, V.S.; Olarte, R.A.; Worthington, C.J.; Carbone, I. Sexual Reproduction in Aspergillus flavus Sclerotia Naturally produced in Corn. Genet. Resist. 2014, 104, 75–85. [Google Scholar]
- Ramirez-Prado, J.H.; Moore, G.G.; Horn, B.W.; Carbone, I. Characterization and Population Analysis of the Mating-Type Genes in Aspergillus flavus and Aspergillus parasiticus. Fungal Genet. Biol. 2008, 45, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Olarte, R.A.; Horn, B.W.; Dorner, J.W.; Monacell, J.T.; Singh, R.; Stone, E.A.; Carbone, I. Effect of sexual recombination on population diversity in aflatoxin production by Aspergillus flavus and evidence for cryptic heterokaryosis. Mol. Ecol. 2012, 21, 1453–1476. [Google Scholar] [CrossRef]
- Seo, J.A.; Guan, Y.; Yu, J.H. Suppressor mutations bypass the requirement of fluG for asexual sporulation and sterigmatocystin production in Aspergillus nidulans. Genetics 2003, 165, 1083–1093. [Google Scholar]
- Lee, B.N.; Adams, T.H. FluG and flbA function interdependently to initiate conidiophore development in Aspergillus nidulans through brlA beta activation. EMBO J. 1996, 15, 299–309. [Google Scholar] [CrossRef]
- Wieser, J.; Lee, B.N.; Fondon, J.W.; Adams, T.H. Genetic requirements for initiating asexual development in Aspergillus nidulans. Curr. Genet. 1994, 27, 62–69. [Google Scholar] [CrossRef]
- Andrianopoulos, A.; Timberlake, W.E. The Aspergillus nidulans abaA gene encodes a transcriptional activator that acts as a genetic switch to control development. Mol. Cell. Biol. 1994, 14, 2503–2515. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Adams, T.H. Complex control of the developmental regulatory locus brlA in Aspergillus nidulans. Mol. Genet. Genom. 2001, 266, 260–270. [Google Scholar] [CrossRef]
- Ni, M.; Yu, J.H. A novel regulator couples sporogenesis and trehalose biogenesis in Aspergillus nidulans. PLoS ONE 2007, 2, e970. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Harris-Coward, P.Y.; Ehrlich, K.C.; Mack, B.M.; Kale, S.P.; Larey, C.; Calvo, A.M. NsdC and NsdD affect Aspergillus flavus morphogenesis and aflatoxin production. Eukaryot. Cell 2012, 11, 1104–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, M.K.; Mack, B.M.; Wei, Q.; Bland, J.M.; Bhatnagar, D.; Cary, J.W. RNA sequencing of an nsdC mutant reveals global regulation of secondary metabolic gene clusters in Aspergillus flavus. Microbiol. Res. 2016, 182, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Lohmar, J.M.; Harris-Coward, P.Y.; Cary, J.W.; Dhingra, S.; Calvo, A.M. rtfA, a putative RNA-Pol II transcription elongation factor gene, is necessary for normal morphological and chemical development in Aspergillus flavus. Appl. Microbiol. Biotechnol. 2016, 100, 5029–5041. [Google Scholar] [CrossRef] [PubMed]
- Lohmar, J.M.; Puel, O.; Cary, J.W.; Calvo, A.M. The Aspergillus flavus rtfA gene regulates plant and animal pathogenesis and secondary metabolism. Appl. Environ. Microbiol. 2019, 85, e02446-18. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Harris-Coward, P.; Scharfenstein, L.; Mack, B.M.; Chang, P.K.; Wei, Q.; Lebar, M.; Carter-Wientjes, C.; Majumdar, R.; Mitra, C.; et al. The Aspergillus flavus homeobox gene, hbx1, is required for development and aflatoxin production. Toxins 2017, 9, 315. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Entwistle, S.; Satterlee, T.; Mack, B.M.; Gilbert, M.K.; Chang, P.K.; Scharfenstein, L.; Yin, Y.; Calvo, A.M. The transcriptional regulator Hbx1 affects the expression of thousands of genes in the aflatoxin-producing fungus Aspergillus flavus. G3 Genes Genomes Genet. 2019, 9, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Kale, S.P.; Cary, J.W.; Baker, C.; Walker, D.; Bhatnagar, D.; Bennett, J.W. Genetic Analysis Of Morphological Variants of Aspergillus parasiticus Deficient in Secondary Metabolite Production. Mycol. Res. 2003, 107, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Scharfenstein, L.L.; Mack, B.; Ehrlich, K.C. Deletion of the Aspergillus flavus orthologue of A. nidulans fluG reduces conidiation and promotes production of sclerotia but does not abolish aflatoxin biosynthesis. Appl. Environ. Microbiol. 2012, 78, 7557–7563. [Google Scholar] [CrossRef] [Green Version]
- Hicks, J.K.; Yu, J.H.; Keller, N.P.; Adams, T.H. Aspergillus sporulation and mycotoxin production both require inactivation of the FadA Gα protein-dependent signaling pathway. EMBO J. 1997, 16, 4916–4923. [Google Scholar] [CrossRef]
- Shimizu, K.; Keller, N. Genetic Involvement of a cAMP-dependent Protein Kinase in a G Protein Signaling Pathway Regulating Morphological and Chemical Transitions in Aspergillus nidulans. Genetics 2001, 157, 591–600. [Google Scholar]
- Ramamoorthy, V.; Shantappa, S.; Dhingra, S.; Calvo, A. veA- dependent RNA-pol II transcription elongation factor-like protein, RtfA, is associated with secondary metabolism and morphological development in Aspergillus nidulans. Mol. Microbiol. 2012, 85, 795–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linz, J.E.; Hong, S.Y.; Roze, L.V. Oxidative stress-related transcription factors in the regulation of secondary metabolism. Toxins 2013, 5, 683–702. [Google Scholar]
- Narasaiah, K.V.; Sashidhar, R.B.; Subramanyam, C. Biochemical analysis of oxidative stress in the production of aflatoxin and its precursor intermediates. Mycopathologia 2006, 162, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Grintzalis, K.; Vernardis, S.I.; Klapa, M.I.; Georgiou, C.D. The role of oxidative stress in sclerotial differentiation and aflatoxin B1 biosynthesis in Aspergillus flavus. Appl. Environ. Microbiol. 2014, 80, 5561–5571. [Google Scholar] [CrossRef] [Green Version]
- Gomez del Arco, P.; Martínez-Martínez, S.; Calvo, V.; Armesilla, A.L.; Redondo, J.M. Antioxidants and AP-1 activation: A brief overview. Immunobiology 1997, 198, 273–278. [Google Scholar] [CrossRef]
- Miskei, M.; Karányi, Z.; Pócsi, I. Annotation of stress-response proteins in the Aspergilli. Fungal Genet. Biol. 2009, 46, S105–S120. [Google Scholar] [CrossRef] [Green Version]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef]
- Yang, K.; Qin, Q.; Liu, Y.; Zhang, L.; Liang, L.; Lan, H.; Chen, C.; You, Y.; Zhang, F.; Wang, S. Adenylate Cyclase AcyA Regulates Development, Aflatoxin Biosynthesis and Fungal Virulence in Aspergillus flavus. Front. Cell. Infect. Microbiol. 2016, 6, 190. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Chen, Z.; Guo, Z.; Li, D.; Zhang, F.; Shen, J.; Zhang, Y.; Wang, S.; Zhuang, Z. PbsB regulates morphogenesis, aflatoxin B1 biosynthesis, and pathogenicity of Aspergillus flavus. Front. Cell. Infect. Microbiol. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Scharfenstein, L.L.; Luo, M.; Mahoney, N.; Molyneux, R.J.; Yu, J.; Brown, R.L.; Campbell, B.C. Loss of msnA, a putative stress regulatory gene, in Aspergillus parasiticus and Aspergillus flavus increased production of conidia, aflatoxins and kojic acid. Toxins 2011, 3, 82–104. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Zhao, Y.; Liu, X.; Shang, B.; Xing, F.; Zhou, L.; Wang, Y.; Zhang, C.; Bhatnagar, D.; Liu, Y. The bZIP transcription factor Afap1 mediates the oxidative stress response and aflatoxin biosynthesis in Aspergillus flavus. Rev. Argent. Microbiol. 2019, 51, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Chanda, A.; Wee, J.; Awad, D.; Linz, J.E. Stress-related transcription factor AtfB integrates secondary metabolism with oxidative stress response in Aspergilli. J. Biol. Chem. 2011, 286, 35137–35148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Punelli, F.; Camera, E.; Fabbri, C.; Picardo, M.; Fanelli, C.; Fabbri, A.A. Modulation of antioxidant defense in Aspergillus parasiticus is involved in aflatoxin biosynthesis: A role for the ApyapA gene. Eukaryot. Cell 2008, 7, 988–1000. [Google Scholar] [CrossRef] [Green Version]
- Weydert, J.C.; Cullen, J. Measurement of Superoxide Dismutase, Catalase and Glutathione Peroxidase in cultured cells and tissue. Nat. Protoc. 2011, 5, 51–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.-M.; Price, M.S.; Obrian, G.R.; Georgianna, D.R.; Payne, G.A. Improved protocols for functional analysis in the pathogenic fungus Aspergillus flavus. BMC Microbiol. 2007, 7, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Shang, B.; Wang, L.; Lu, Z.; Liu, Y. Cinnamaldehyde inhibits fungal growth and aflatoxin B1 biosynthesis by modulating the oxidative stress response of Aspergillus flavus. Appl. Microbiol. Biotechnol. 2015, 100, 1355–1364. [Google Scholar] [CrossRef]
- Reverberi, M.; Fabbri, A.A.; Zjalic, S.; Ricelli, A.; Punelli, F.; Fanelli, C. Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl. Microbiol. Biotechnol. 2005, 69, 207–215. [Google Scholar] [CrossRef]
- Leiter, É.; Park, H.-S.; Kwon, N.-J.; Han, K.-H.; Emri, T.; Oláh, V.; Mészáros, I.; Dienes, B.; Vincze, J.; Csernoch, L.; et al. Characterization of the aodA, dnmA, mnSOD and pimA genes in Aspergillus nidulans. Sci. Rep. 2016, 6, 20523. [Google Scholar] [CrossRef] [Green Version]
- Maggio-Hall, L.A.; Keller, N.P. Mitochondrial β-oxidation in Aspergillus nidulans. Mol. Microbiol. 2004, 54, 1173–1185. [Google Scholar] [CrossRef]
- Reverberi, M.; Punelli, M.; Smith, C.A.; Zjalic, S.; Scarpari, M.; Scala, V.; Cardinali, G.; Aspite, N.; Pinzari, F.; Payne, G.A.; et al. How Peroxisomes Affect Aflatoxin Biosynthesis in Aspergillus flavus. PLoS ONE 2012, 7, e48097. [Google Scholar] [CrossRef]
- Chanda, A.; Roze, L.V.; Kang, S.; Artymovich, K.A.; Hicks, G.R.; Raikhel, N.V.; Calvo, A.M.; Linz, J.E. A key role for vesicles in fungal secondary metabolism. Proc. Natl. Acad. Sci. USA 2009, 106, 19533–19538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hynes, M.J.; Murray, S.L.; Khew, G.S.; Davis, M.A. Genetic analysis of the role of peroxisomes in the utilization of acetate and fatty acids in Aspergillus nidulans. Genetics 2008, 178, 1355–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.Y.; Linz, J.E. Functional Expression and Sub-Cellular Localization of the Early Aflatoxin Pathway Enzyme Nor-1 in Aspergillus parasiticus. Mycol. Res. 2009, 113, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.F.; Van Den Berg, M.A.; Ver Loren Van, T.; Liras, P. Sensing and transduction of nutritional and chemical signals in filamentous fungi: Impact on cell development and secondary metabolites biosynthesis. Biotechnol. Adv. 2019, 37, 107392. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Keller, N.P. Oxylipins act as determinants of natural product biosynthesis and seed colonization in Aspergillus nidulans. Mol. Microbiol. 2006, 59, 882–892. [Google Scholar] [CrossRef]
- Dos Reis, T.F.; Mellado, L.; Lohmar, J.M.; Lilian, P.S.; Zhou, J.-J.; Calvo, A.M.; Goldman, G.H.; Brown, N.A. GPCR-mediated glucose sensing system regulates light-dependent fungal development and mycotoxin production. PLoS Genet. 2019, 15, e1008419. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Catabolic Repression | |
|---|---|
| Genes | Coding Function |
| creA | Cys2His2 Zinc finger transcription repressor |
| creB | Cys2His2 Zinc finger ubiquitin processing protease |
| creC | Cys2His2 Zinc finger ubiquitin processing protease |
| alcR | Regulon specific transcription factor |
| aldA | Aldehyde dehydrogenase |
| alcA | Alcohol dehydrogenase |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| creA represses the expression of aclR while the latter is a positive regulatory factor for the genes aldA and alcA [101,103]. CreB-CreC form a protein complex that is essential for CreA function and stability [104]. | |
| Demonstration of the connection with AF/ST synthesis | |
| Gene expression in the AF/ST gene cluster is regulated either positively or negatively by CreA depending on the fungal species [31,105]. In A. flavus: ΔcreA strains produced insignificant amounts of aflatoxin in complete medium and their ability to colonize hosts was also impaired [102]. The repression of creA by gallic acid lead to the inhibition of AFB1 production [106]. In A. parasiticus: Several genes in the AFB1 cluster were demonstrated to have CreA-sites near their promoter regions [107]. | |
| Nitrogen Source | |
|---|---|
| Genes | Coding Function |
| areA | Zinc finger mediating nitrogen metabolite repression |
| nmrA | Repressive nitrogen |
| meaB | Regulatory protein |
| niiA | Nitrite reductase |
| niaD | Nitrate reductase |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| areA and meaB are regulatory genes that mediate nitrogen metabolite repression [108,109] AreA regulates nitrate transporters binding at the intergenic regions of niiA and niaD [110] MeaB is a regulatory factor that activates NmrA, which is a repressor of AreA [109]. | |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: areA was recently identified in A. flavus and demonstrated to be partially involved in aflatoxin production. The lack and overexpression of areA resulted in increased/decreased amounts of aflatoxin depending on the nitrogen source media [111]. The direct implication of AreA in aflS expression has been demonstrated in the presence of different sources of nitrate and ammonium [112]. Overexpressing meaB strains lost their capacity to produce AFs on seeds while ΔmeaB produced only a few aflatoxins [109]. niiA is one of the genes that does not belong to the AFB1 gene cluster but that is presumed to be regulated by the major internal AF regulator aflR [26]. In A. parasiticus: AreA binds to the GATA sites of the promoters of aflR/aflS [110,113]. aflC and aflD genes are expressed on ammonium and peptone media but not on nitrate sources [114]. In A. nidulans: Nitrate medium increases the production of sterigmatocystin while ammonium medium does not [114]. | |
| pH Impact | |
|---|---|
| Gene | Coding Function |
| pacC | Cys2His2 (C2-H2)- Zinc finger pH regulator |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| The transcription factor PacC is strongly expressed under alkaline conditions [117]. | |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: An increase in pH in nitrate-based medium results in lower levels of AF, while lower pH (4.0) resulted in a 10-fold increase in production of AF [119]. InA. parasiticus: pH levels below 6.0 promote B-type aflatoxin production while higher levels induce G-type production [120]. A putative PacC binding site was identified in the aflR promoter region, but interestingly, alkaline conditions in which pacC is activated do not support AFB1 production [113]. aflM was higher expressed in acidic media than in neutral and alkaline media. Fungal growth reduced the pH of the medium and increased AF production with time [121]. In A. nidulans: Acidic conditions are more favorable for ST biosynthesis than neutral or alkaline ones [121]. pacC-mutant strains produced 10-fold less ST than the control. The expression level of stcU (aflM homologous) was lower in increased pH media [121]. | |
| Light | |
|---|---|
| Genes | Coding function |
| veA | Global regulator |
| vapB | Methyltransferase |
| vipC | Methyltransferase |
| fphA | Phytochrome-like red light receptor |
| kapA | α transport carrier |
| velB | Velvet-like protein B |
| laeA | Putative methyltransferase |
| velC | Velvet-like protein C |
| lreA | Blue-light sensing protein |
| lreB | Blue-light sensing protein |
| llmF | LaeA-like methyltransferase |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| Vea-LaeA-VelB form a trimeric complex called the velvet complex. VeA interacts with LaeA in the nucleus and with VelB in the cytoplasm and the nucleus. This trimeric complex, together with other light-receptor proteins, perceives light signals and is an essential coordinator of secondary metabolism and fungal development [124,125,126,127,128]. In dark conditions, the nuclear localization of VeA increases its interaction with LaeA to enhance production of secondary metabolites but also with VelB to induce sexual development [129]. FphA interacts with VeA, with LreB and LreA in the nucleus [123,130] and this protein complex is involved in red and blue light sensing [124]. KapA and VeA physically interact in dark conditions. KapA supports the entry of the VeA-VelB complex into the nucleus [126]. Both VelC and VelB form a protein dimer with VosA (involved in spore viability) [126]. VipC and VapB reduce the nuclear accumulation of VeA, thereby reducing secondary metabolism [131]. LlmF interacts with VeA controlling its subcellular location [129]. | |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: veA is essential for AFB1 production [132,133]. In null mutants of veA or laeA, no aflR expression was observed [134]. ΔlaeA reduced aflR, aflD and aflS mRNA levels with no AF production [135]. Overexpression of laeA results in higher levels of AFB1 whereas ΔlaeA strains do not affect aflatoxin production [109]. VeA governs 28 out of the 56 secondary metabolite gene clusters including the AF cluster [136]. In A. parasiticus: Deletion of veA resulted in the loss of the aflatoxin intermediate Versicolorin A. VeA is required for aflR/aflS expression [137]. In A. nidulans: ΔveA or ΔveA/vipC mutants were unable to produce sterigmatocystin but ΔvipC did not affect the mycotoxin production [131]. Both sexual development and sterigmatocystin production were repressed in light conditions [138]. ΔveA strains resulted in no aflR expression and neither ST production [139]. Deletion of the laeA gene inhibits aflR and stcU expression [140]. LreA, LreB and FphA modulate sterigmatocystin biosynthesis depending on light and on the presence of glucose [141]. FphA represses sexual development and ST production while LreA and LreB stimulate both processes. Blue light represses ST production while red light has the opposite effect [124]. LlmF is a negative factor for ST production [129]. | |
| Sexual Development | |
|---|---|
| Genes | Coding Function |
| MAT1-1 | Mating type (alpha) |
| MAT1-2 | Mating type (HMG) |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| Either MAT1-1 or MAT1-2 is expressed in A. flavus and A. parasiticus strains and involved in sexual development [145]. | |
| Demonstration of the connection with AF synthesis | |
| InA. flavus: Strains of sexually developed A. flavus demonstrated that production of AF is highly heritable. In asexual development, non-aflatoxigenic populations are maintained while aflatoxigenicity increases in sexual development [146]. | |
| Asexual Development | |
|---|---|
| Genes | Coding Function |
| fadA | α-subunit of heterotrimeric G-protein |
| fluG | Developmental regulator |
| brlA | C2H2 zinc finger protein transcriptional activator of conidiophore |
| abaA | Transcription factor for conidia formation |
| wetA | Developmental regulatory protein |
| nsdC | Zinc-finger transcription factor |
| nsdD | Zinc-finger transcription factor |
| pkaA | Catalytic subunit of protein kinase A |
| flbA | RGS protein/developmental regulator |
| flbB | bZIP-type transcription factor |
| flbC | Putative C2H2 conidiation transcription factor |
| flbD | MYB family conidiophore development |
| flbE | Developmental regulator |
| vosA | Spore viability/Developmental regulator/Trehalose production |
| rtfA | RNA-pol II transcription elongation factor-like protein |
| hbx1 | Homebox transcription factor |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| fluG activates flbA which in turn represses fadA signaling [147]. fadA up-regulates pkaA [97]. flbA is a regulator of flbB, which regulates flbC, flbD, flbE (flb genes) [148]. flb genes are required with fluG for the correct expression of brlA [149]. brlA is a negative regulator of abaA and abaA is a repressor of wetA [126,150,151]. brlA is regulated by veA [139]. vosA is a repressor of brlA [152]. VosA also forms a protein-complex with VelB and VelC (velvet proteins) [126,127]. nsdC and nsdD are repressors of brlA [153]. | |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: Expression of aflD, aflM, and aflP is strongly reduced in nsdC deleted mutants. Loss of NsdC or NsdD resulted in developmental alterations that impact the ability of AflR to activate the expression of AF biosynthesis genes [153]. In fact, A. flavus nsdC mutants are unable to produce AF or any other secondary metabolites [154]. FadA governs both AF and ST biosynthesis [74,97]. Deleted strains of rtfA greatly reduced AFB1 biosynthesis but interestingly, aflR, aflM and aflP were up-regulated within the 3 first days of incubation and then down-regulated on the 4th and 5th day. Moreover, rtfA controls veA and laeA expression [155] and production of other secondary metabolites [156]. Disrupted hbx1 strains produced null B-type aflatoxin, conidia and sclerotia [157]. Several genes belonging to the aflatoxin cluster were not expressed in hbx1 mutant strains [158]. In A. parasiticus: Mutants defective in conidiation processes also had reduced levels of AF production [159]. FadA is presumed to regulate aflT, the MFS belonging to the AFB1 gene cluster [74]. In A. nidulans: FadA up-regulates pkaA, which down-regulates conidiation. PkaA also inhibits AflR activity by phosphorylation [77]. fluG-deleted strains lost their ability to produce sterigmatocystin [160]. Mutations in flbA and fluG blocked both sterigmatocystin production and sporulation [161]. ΔfadA and ΔpkaA mutants failed in normal processes of conidiation and sterigmatocystin biosynthesis [162]. Co-regulation of brlA and aflR by the fadA signaling pathway genes was reported [139]. Sterigmatocystin production was positively regulated by rtfA [163]. | |
| Oxidative Stress Complex | |
|---|---|
| Genes | Coding Function |
| bZIP transcription factors | |
| ap-1 | bZIP transcription factor |
| atfA | bZIP transcription factor |
| atfB | bZIP transcription factor |
| Stress Response Signaling Pathway | |
| srrA | Transcription factor |
| msnA | Transcription factor |
| acyA | Adenylate Cyclase |
| pbsB | MAP kinase kinase |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| In cell systems, the ap-1 gene is activated under both, pro-oxidant and antioxidant conditions [167]. Together, AtfB, SrrA, Ap-1, PbsB and MsnA form a regulatory network involved in oxidative stress response and secondary metabolite production [67,168]. AtfA may interact with AtfB in response to oxidative stress [67,169]. acyA was shown to regulate AF biosynthesis as well as to intervene in hyperosmotic and oxidative stress [170]. pbsB was reported to be involved in stress responses and AFB1 biosynthesis [171]. | |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: The deleted ΔacyA strains were unable to produce aflatoxin in contrast to the control and some genes in the AF gene cluster (aflR and aflO) were also down-regulated [170]. PbsB positively regulates AFB1 production through aflR, the major regulator of the AFB1 gene cluster, as well as other genes such as aflC, aflD, aflK and aflQ [171]. msnA deletion results in 50% more aflatoxins as well as higher levels of reactive oxygen species (ROS) [172]. ap1 deletion reduces aflatoxin production while the expression of aflM and aflP was down-regulated even though aflR was up-regulated [173]. In A. parasiticus: AtfB binds to promoters of seven genes belonging to the AF gene cluster [174]. ap-1 deletion increases AF production while the Ap-1 protein binds to the promoter region of the aflR gene [175]. | |
| Superoxide Dismutases and Catalases | |
|---|---|
| Genes | Coding Function |
| mnSOD | Manganese superoxide dismutase |
| sod1 | Cu, Zn superoxide dismutase |
| catA | Conidia-specific catalase |
| catB | Mycelial catalase |
| hyr1 | Glutathione peroxidase |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: mnSOD and the genes aflA, aflM, and aflP belonging to the AF gene cluster are co-regulated [67]. Deletion of sod reduced AF production [177]. Increased expression of the genes of catA, cat2 and sod1 as well as an increased CAT enzymatic activity were observed in the presence of the AF inhibitor piperine [15]. The same increase in CAT activity was also observed with another AF inhibitor, cinnamaldehyde [178]. In A. parasiticus: Inhibition of AF production induced by Lentinula edodes increases SOD enzymatic activity within its mechanism of action [179]. In A. nidulans: Deletion of mnSOD increased both glutathione reductase and catalase activities while its overexpression reduced the activity of catalase but increased SOD activity [180]. | |
| β-oxidation | |
|---|---|
| Genes | Coding Function |
| pexK | Existence of peroxisome |
| pex11 | Peroxisome proliferation |
| foxA | Regulation of fatty acid metabolism by β -oxidation |
| rab7 | Vesicle marker |
| vps16 | Vacuole marker |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| In filamentous fungi, peroxisomes are crucial for primary metabolism and play a role in the formation of some secondary metabolites [182]. Aflatoxisomes (aflatoxin vesicles) are partially regulated by VeA [183]. pex mutants are able to grow on acetate medium but their growth is affected by fatty acids, indicating β-oxidation enzymes require a peroxisomal location [184]. | |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: An increase in the number of peroxisomes enhances AFB1 production [182]. InA. parasiticus: An increase in the number of vesicles is positively correlated with AF accumulation/export [183]. Nor-1 (protein involved in the norsolorinic acid biosynthesis) mainly occurs in the cytoplasm and vacuoles [185]. | |
| Cell Signaling | |
|---|---|
| Genes | Coding Function |
| ppoA | (oxylipin) Dioxygenase |
| ppoB | (oxylipin) Dioxygenase |
| ppoC | (oxylipin) Dioxygenase |
| ppoD | (oxylipin) Dioxygenase |
| lox | (oxylipin) Lipoxygenase |
| gprK | GPCR |
| gprA | GPCR |
| gprP | GPCR |
| rasA | GTP-binding protein |
| Relation between genes or corresponding proteins in Aspergilli spp.: | |
| GPCRs are involved in oxylipin response [30]. | |
| Demonstration of the connection with AF/ST synthesis | |
| InA. flavus: The GPCRs were shown to interact with AFB1 synthesis and its precursor, sterigmatocystin (ST). Deletion of gprK and grpA increased AF production compared to the control strain [30]. When all four ppo genes and the lox gene were disrupted simultaneously, the mutant strains showed reduced conidiation and increased AF production on maize and peanut seeds [169]. In A. nidulans: ΔppoA; ΔppoB and ΔppoC mutants are unable to produce ST [187]. RasA has been demonstrated to control aflR activity [77]. GprH and GprM are negative regulators of ST biosynthesis [188]. | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caceres, I.; Al Khoury, A.; El Khoury, R.; Lorber, S.; P. Oswald, I.; El Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.-D. Aflatoxin Biosynthesis and Genetic Regulation: A Review. Toxins 2020, 12, 150. https://doi.org/10.3390/toxins12030150
Caceres I, Al Khoury A, El Khoury R, Lorber S, P. Oswald I, El Khoury A, Atoui A, Puel O, Bailly J-D. Aflatoxin Biosynthesis and Genetic Regulation: A Review. Toxins. 2020; 12(3):150. https://doi.org/10.3390/toxins12030150
Chicago/Turabian StyleCaceres, Isaura, Anthony Al Khoury, Rhoda El Khoury, Sophie Lorber, Isabelle P. Oswald, André El Khoury, Ali Atoui, Olivier Puel, and Jean-Denis Bailly. 2020. "Aflatoxin Biosynthesis and Genetic Regulation: A Review" Toxins 12, no. 3: 150. https://doi.org/10.3390/toxins12030150
APA StyleCaceres, I., Al Khoury, A., El Khoury, R., Lorber, S., P. Oswald, I., El Khoury, A., Atoui, A., Puel, O., & Bailly, J. -D. (2020). Aflatoxin Biosynthesis and Genetic Regulation: A Review. Toxins, 12(3), 150. https://doi.org/10.3390/toxins12030150