Allergy—A New Role for T Cell Superantigens of Staphylococcus aureus?

 , and
, and
Abstract
:1. Introduction
- (1)
- Provides an overview on the function and diversity of staphylococcal superantigens (SAgs),
- (2)
- Reports on advances in the development of SAg vaccines,
- (3)
- Summarizes recent epidemiological data on the involvement of SAgs in allergy,
- (4)
- Outlines mechanisms by which SAgs could induce or amplify allergic responses,
- (5)
- Elaborates on the evolutionary advantage gained by the production of SAgs, and finally,
- (6)
- Discusses knowledge gaps that should be addressed in future research.
1.1. SAgs are Extremely Potent T Cell Mitogens
1.2. Staphylococcal SAgs are Highly Diverse
1.3. SAgs Can Induce Various Clinical Pictures
2. Vaccination against SAgs
3. SAgs and Allergy
3.1. Epidemiological Evidence for SAg Involvement in Allergy
3.2. Interactions of SAgs with Immune Cells
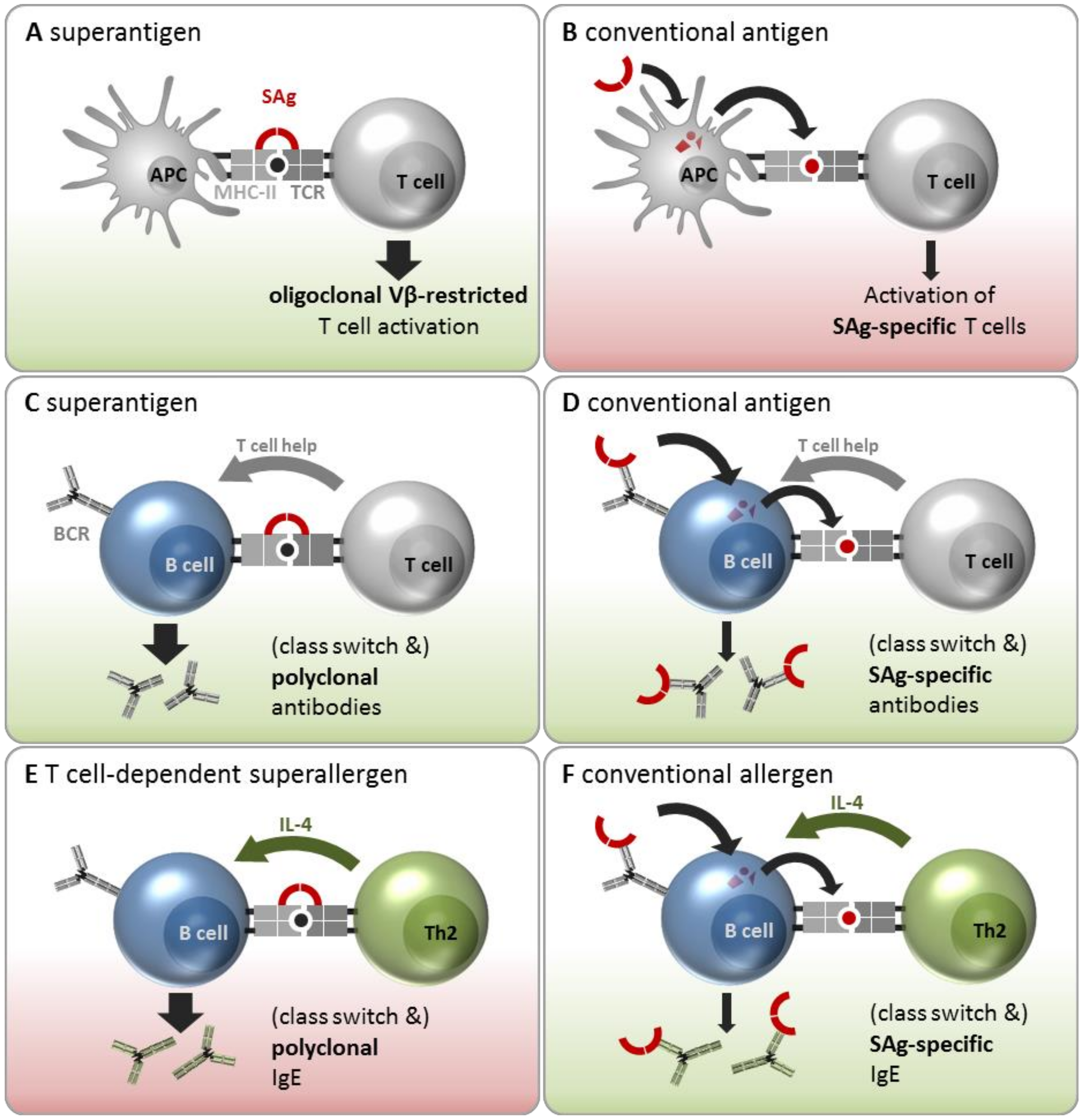
3.2.1. Interaction with T Cells and APCs as SAg and Conventional Antigen
3.2.2. Interactions with B Cells as SAgs and Conventional Antigens
3.2.3. Interaction with B Cells as Superallergens and Conventional Allergens
3.2.4. Interaction with Other Immune Cells
4. How Does S. aureus Benefit from Producing SAgs?
5. Open Questions
- Currently, specific IgE antibodies can be measured against four different SAgs, SEA–SEC, and TSST-1, using commercial tests. To evaluate the potential roles of the other 22 SAgs in allergic diseases the appropriate assays need to be established.
- SAg-specific T cells, i.e., those that specifically recognize a SAg peptide in complex with a MHC-II molecule, have not been studied to date. To understand their role in allergic diseases, however, it is crucial to determine the quality of the natural T cell response against SAgs in healthy individuals and patients. With the availability of SAg toxoids and sophisticated T cell assays, this will now be feasible.
- T cell-dependent superallergens are thought to stimulate B cells of any specificity to form IgE. However, evidence to support this notion of SAg-mediated polysensitization is scarce.
- Considering the high prevalence of anti-SAg antibodies, including SE-IgE, it is likely that most individuals are primed with S. aureus SAgs and have generated an anti-SAg T cell response, which may comprise substantial numbers of Th2 cells. Vaccination with SAg toxoids without adjuvant might enhance this pre-existing Th2 bias in susceptible individuals, thereby exacerbating allergy rather than inducing protection. To avoid this, the SAg-specific T cells responses have to be studied in healthy individuals and patient cohorts.
- After more than 30 years of research, the evolutionary advantages gained from SAgs are still under discussion. Using SAg-knock-out strains or SAg vaccines in animal models could help to scrutinize their role in colonization and infection.
- Since SAgs likely play an important role in allergic disorders, future research should enforce efforts to develop broadly cross-reactive SAg vaccines or SAg antagonists.
6. Closing Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wertheim, H.F.L.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Van Belkum, A.; Verkaik, N.J.; de Vogel, C.P.; Boelens, H.A.; Verveer, J.; Nouwen, J.L.; Verbrugh, H.A.; Wertheim, H.F.L. Reclassification of Staphylococcus aureus nasal carriage types. J. Infect. Dis. 2009, 199, 1820–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Teufelberger, A.R.; Bröker, B.M.; Krysko, D.V.; Bachert, C.; Krysko, O. Staphylococcus aureus orchestrates type 2 airway diseases. Trends Mol. Med. 2019, 25, 696–707. [Google Scholar] [CrossRef] [Green Version]
- Tam, K.; Torres, V.J. Staphylococcus aureus secreted toxins and extracellular enzymes. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Fraser, J.D.; Proft, T. The bacterial superantigen and superantigen-like proteins. Immunol. Rev. 2008, 225, 226–243. [Google Scholar] [CrossRef] [PubMed]
- Marrack, P.; Kappler, J. The staphylococcal enterotoxins and their relatives. Science 1990, 248, 1066. [Google Scholar] [CrossRef]
- Fleischer, B. T lymphocyte-stimulating microbial toxins as “superantigens”. Med. Microbiol. Immunol. 1991, 180, 53–58. [Google Scholar] [CrossRef]
- Bröker, B.M.; Holtfreter, S.; Bekeredjian-Ding, I. Immune control of Staphylococcus aureus—Regulation and counter-regulation of the adaptive immune response. Int. J. Med. Microbiol. 2014, 304, 204–214. [Google Scholar] [CrossRef]
- Spaulding, A.R.; Salgado-Pabon, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.M.; Schlievert, P.M. Staphylococcal and streptococcal superantigen exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef] [Green Version]
- Holtfreter, S.; Bröker, B.M. Staphylococcal superantigens: Do they play a role in sepsis? Arch. Immunol. Ther. Exp. (Warsz) 2005, 53, 13–27. [Google Scholar] [PubMed]
- Grumann, D.; Nübel, U.; Bröker, B.M. Staphylococcus aureus toxins—Their functions and genetics. Infect. Genet. Evol. 2014, 21, 583–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, D.; Borges, A.; Simões, M. Staphylococcus aureus toxins and their molecular activity in infectious diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehindate, K.; Al-Daccak, R.; Damdoumi, F.; Mourad, W. Synergistic effect between CD40 and class II signals overcome the requirement for class II dimerization in superantigen-induced cytokine gene expression. Eur. J. Immunol. 1996, 26, 2075–2080. [Google Scholar] [CrossRef]
- Espel, E.; Garcia-Sanz, J.A.; Aubert, V.; Menoud, V.; Sperisen, P.; Fernández, N.; Spertini, F. Transcriptional and translational control of TNF-alpha gene expression in human monocytes by major histocompatibility complex class II ligands. Eur. J. Immunol. 1996, 26, 2417–2424. [Google Scholar] [CrossRef]
- Noli Truant, S.; de Marzi, M.C.; Sarratea, M.B.; Antonoglou, M.B.; Meo, A.P.; Iannantuono López, L.V.; Fernández Lynch, M.J.; Todone, M.; Malchiodi, E.L.; Fernández, M.M.; et al. egc superantigens impair monocytes/macrophages inducing cell death and inefficient activation. Front. Immunol. 2019, 10, 3008. [Google Scholar] [CrossRef] [Green Version]
- Jarraud, S.; Peyrat, M.A.; Lim, A.; Tristan, A.; Bes, M.; Mougel, C.; Etienne, J.; Vandenesch, F.; Bonneville, M.; Lina, G.; et al. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J. Immunol. 2001, 166, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Holtfreter, S.; Grumann, D.; Schmudde, M.; Nguyen, H.T.T.; Eichler, P.; Strommenger, B.; Kopron, K.; Kolata, J.; Giedrys-Kalemba, S.; Steinmetz, I.; et al. Clonal distribution of superantigen genes in clinical Staphylococcus aureus isolates. J. Clin. Microbiol. 2007, 45, 2669–2680. [Google Scholar] [CrossRef] [Green Version]
- Grumann, D.; Scharf, S.S.; Holtfreter, S.; Kohler, C.; Steil, L.; Engelmann, S.; Hecker, M.; Völker, U.; Bröker, B.M. Immune cell activation by enterotoxin gene cluster (egc)-encoded and non-egc superantigens from Staphylococcus aureus. J. Immunol. 2008, 181, 5054–5061. [Google Scholar] [CrossRef] [Green Version]
- Dauwalder, O.; Thomas, D.; Ferry, T.; Debard, A.-L.; Badiou, C.; Vandenesch, F.; Etienne, J.; Lina, G.; Monneret, G. Comparative inflammatory properties of staphylococcal superantigenic enterotoxins SEA and SEG: Implications for septic shock. J. Leukoc. Biol. 2006, 80, 753–758. [Google Scholar] [CrossRef]
- Holtfreter, S.; Bauer, K.; Thomas, D.; Feig, C.; Lorenz, V.; Roschack, K.; Friebe, E.; Selleng, K.; Lövenich, S.; Greve, T.; et al. egc-Encoded superantigens from Staphylococcus aureus are neutralized by human sera much less efficiently than are classical staphylococcal enterotoxins or toxic shock syndrome toxin. Infect. Immun. 2004, 72, 4061–4071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burian, M.; Grumann, D.; Holtfreter, S.; Wolz, C.; Goerke, C.; Bröker, B.M. Expression of staphylococcal superantigens during nasal colonization is not sufficient to induce a systemic neutralizing antibody response in humans. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Moreno, D.; Wos-Oxley, M.L.; Jáuregui, R.; Medina, E.; Oxley, A.P.; Pieper, D.H. Exploring the transcriptome of Staphylococcus aureus in its natural niche. Sci. Rep. 2016, 6, 33174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.T.; Chung, S.W.; Jung, J.H.; Ha, J.S.; Kang, I.G. Association of T cells and eosinophils with Staphylococcus aureus exotoxin A and toxic shock syndrome toxin 1 in nasal polyps. Am. J. Rhinol. Allergy 2011, 25, 19–24. [Google Scholar] [CrossRef]
- Seiberling, K.A.; Conley, D.B.; Tripathi, A.; Grammer, L.C.; Shuh, L.; Haines, G.K.; Schleimer, R.; Kern, R.C. Superantigens and chronic rhinosinusitis: Detection of staphylococcal exotoxins in nasal polyps. Laryngoscope 2005, 115, 1580–1585. [Google Scholar] [CrossRef]
- Schmidt, F.; Meyer, T.; Sundaramoorthy, N.; Michalik, S.; Surmann, K.; Depke, M.; Dhople, V.; Gesell Salazar, M.; Holtappels, G.; Zhang, N.; et al. Characterization of human and Staphylococcus aureus proteins in respiratory mucosa by in vivo- and immunoproteomics. J. Proteom. 2017, 155, 31–39. [Google Scholar] [CrossRef]
- Ishii, K.; Adachi, T.; Yasukawa, J.; Suzuki, Y.; Hamamoto, H.; Sekimizu, K. Induction of virulence gene expression in Staphylococcus aureus by pulmonary surfactant. Infect. Immun. 2014, 82, 1500–1510. [Google Scholar] [CrossRef] [Green Version]
- Proft, T.; Fraser, J.D. Bacterial superantigens. Clin. Exp. Immunol. 2003, 133, 299–306. [Google Scholar] [CrossRef]
- Verkaik, N.J.; de Vogel, C.P.; Boelens, H.A.; Grumann, D.; Hoogenboezem, T.; Vink, C.; Hooijkaas, H.; Foster, T.J.; Verbrugh, H.A.; van Belkum, A.; et al. Anti-staphylococcal humoral immune response in persistent nasal carriers and noncarriers of Staphylococcus aureus. J. Infect. Dis. 2009, 199, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.-L.; Nakane, A. Mechanisms of staphylococcal enterotoxin-induced emesis. Eur. J. Pharmacol. 2014, 722, 95–107. [Google Scholar] [CrossRef]
- Papageorgiou, A.C.; Acharya, K.R. Microbial superantigens: From structure to function. Trends Microbiol. 2000, 8, 369–375. [Google Scholar] [CrossRef]
- Petersson, K.; Pettersson, H.; Skartved, N.J.; Walse, B.; Forsberg, G. Staphylococcal enterotoxin H induces V -specific expansion of T cells. J. Immunol. 2003, 170, 4148–4154. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-J.; Hu, D.-L.; Maina, E.K.; Shinagawa, K.; Omoe, K.; Nakane, A. Superantigenic activity of toxic shock syndrome toxin-1 is resistant to heating and digestive enzymes. J. Appl. Microbiol. 2011, 110, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, T. Staphylococcal superantigens: Pyrogenic toxins induce toxic shock. Toxins 2019, 11, 178. [Google Scholar] [CrossRef] [Green Version]
- Holtfreter, S.; Roschack, K.; Eichler, P.; Eske, K.; Holtfreter, B.; Kohler, C.; Engelmann, S.; Hecker, M.; Greinacher, A.; Broker, B.M. Staphylococcus aureus carriers neutralize superantigens by antibodies specific for their colonizing strain: A potential explanation for their improved prognosis in severe sepsis. J. Infect. Dis. 2006, 193, 1275–1278. [Google Scholar] [CrossRef] [Green Version]
- Grumann, D.; Ruotsalainen, E.; Kolata, J.; Kuusela, P.; Järvinen, A.; Kontinen, V.P.; Bröker, B.M.; Holtfreter, S. Characterization of infecting strains and superantigen-neutralizing antibodies in Staphylococcus aureus bacteremia. Clin. Vaccine Immunol. 2011, 18, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Vergeront, J.M.; Stolz, S.J.; Crass, B.A.; Nelson, D.B.; Davis, J.P.; Bergdoll, M.S. Prevalence of serum antibody to staphylococcal enterotoxin F among Wisconsin residents: Implications for toxic-shock syndrome. J. Infect. Dis. 1983, 148, 692–698. [Google Scholar] [CrossRef]
- Jardetzky, T.S.; Brown, J.H.; Gorga, J.C.; Stern, L.J.; Urban, R.G.; Chi, Y.I.; Stauffacher, C.; Strominger, J.L.; Wiley, D.C. Three-dimensional structure of a human class II histocompatibility molecule complexed with superantigen. Nature 1994, 368, 711–718. [Google Scholar] [CrossRef]
- Acharya, K.R.; Passalacqua, E.F.; Jones, E.Y.; Harlos, K.; Stuart, D.I.; Brehm, R.D.; Tranter, H.S. Structural basis of superantigen action inferred from crystal structure of toxic-shock syndrome toxin-1. Nature 1994, 367, 94–97. [Google Scholar] [CrossRef]
- Fields, B.A.; Malchiodi, E.L.; Li, H.; Ysern, X.; Stauffacher, C.V.; Schlievert, P.M.; Karjalainen, K.; Mariuzza, R.A. Crystal structure of a T-cell receptor beta-chain complexed with a superantigen. Nature 1996, 384, 188–192. [Google Scholar] [CrossRef]
- Zeng, C.; Liu, Z.; Han, Z. Structure of staphylococcal enterotoxin N: Implications for binding properties to its cellular proteins. Int. J. Mol. Sci. 2019, 20, 5921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rödström, K.E.J.; Regenthal, P.; Lindkvist-Petersson, K. Structure of staphylococcal enterotoxin E in complex with TCR defines the role of TCR loop positioning in superantigen recognition. PLoS ONE 2015, 10, e0131988. [Google Scholar] [CrossRef] [PubMed]
- Rödström, K.E.J.; Elbing, K.; Lindkvist-Petersson, K. Structure of the superantigen staphylococcal enterotoxin B in complex with TCR and peptide-MHC demonstrates absence of TCR-peptide contacts. J. Immunol. 2014, 193, 1998–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, M.M.; Cho, S.; de Marzi, M.C.; Kerzic, M.C.; Robinson, H.; Mariuzza, R.A.; Malchiodi, E.L. Crystal structure of staphylococcal enterotoxin G (SEG) in complex with a mouse T-cell receptor {beta} chain. J. Biol. Chem. 2011, 286, 1189–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonventre, P.F.; Heeg, H.; Edwards, C.K.; Cullen, C.M. A mutation at histidine residue 135 of toxic shock syndrome toxin yields an immunogenic protein with minimal toxicity. Infect. Immun. 1995, 63, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Grossman, D.; Van, M.; Mollick, J.A.; Highlander, S.K.; Rich, R.R. Mutation of the disulfide loop in staphylococcal enterotoxin A. Consequences for T cell recognition. J. Immunol. 1991, 147, 3274–3281. [Google Scholar]
- Aman, M.J. Superantigens of a superbug: Major culprits of Staphylococcus aureus disease? Virulence 2017, 8, 607–610. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.H.; Pasetti, M.F.; Adhikari, R.P.; Baughman, H.; Douglas, R.; El-Khorazaty, J.; Greenberg, N.; Holtsberg, F.W.; Liao, G.C.; Reymann, M.K.; et al. Safety and Immunogenicity of a parenterally administered, structure-based rationally modified recombinant staphylococcal enterotoxin B protein vaccine, STEBVax. Clin. Vaccine Immunol. 2016, 23, 918–925. [Google Scholar] [CrossRef] [Green Version]
- Schwameis, M.; Roppenser, B.; Firbas, C.; Gruener, C.S.; Model, N.; Stich, N.; Roetzer, A.; Buchtele, N.; Jilma, B.; Eibl, M.M. Safety, tolerability, and immunogenicity of a recombinant toxic shock syndrome toxin (rTSST)-1 variant vaccine: A randomised, double-blind, adjuvant-controlled, dose escalation first-in-man trial. Lancet Infect. Dis. 2016, 16, 1036–1044. [Google Scholar] [CrossRef]
- Bröker, B.M.; Mrochen, D.; Péton, V. The T cell response to Staphylococcus aureus. Pathogens 2016, 5. [Google Scholar] [CrossRef]
- Bavari, S.; Dyas, B.; Ulrich, R.G. Superantigen vaccines: A comparative study of genetically attenuated receptor-binding mutants of staphylococcal enterotoxin A. J. Infect. Dis. 1996, 174, 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.-L.; Omoe, K.; Sashinami, H.; Shinagawa, K.; Nakane, A. Immunization with a nontoxic mutant of staphylococcal enterotoxin A, SEAD227A, protects against enterotoxin-induced emesis in house musk shrews. J. Infect. Dis. 2009, 199, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.-L.; Omoe, K.; Sasaki, S.; Sashinami, H.; Sakuraba, H.; Yokomizo, Y.; Shinagawa, K.; Nakane, A. Vaccination with nontoxic mutant toxic shock syndrome toxin 1 protects against Staphylococcus aureus infection. J. Infect. Dis. 2003, 188, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Spaulding, A.R.; Salgado-Pabón, W.; Merriman, J.A.; Stach, C.S.; Ji, Y.; Gillman, A.N.; Peterson, M.L.; Schlievert, P.M. Vaccination against Staphylococcus aureus pneumonia. J. Infect. Dis. 2014, 209, 1955–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.-L.; Cui, J.-C.; Omoe, K.; Sashinami, H.; Yokomizo, Y.; Shinagawa, K.; Nakane, A. A mutant of staphylococcal enterotoxin C devoid of bacterial superantigenic activity elicits a Th2 immune response for protection against Staphylococcus aureus infection. Infect. Immun. 2005, 73, 174–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, A.K.; Wang, X.; Scharff, M.D.; MacIntyre, J.; Zollner, R.S.; Kovalenko, O.V.; Martinez, L.R.; Byrne, F.R.; Fries, B.C. Staphylococcal enterotoxin B-specific monoclonal antibody 20B1 successfully treats diverse Staphylococcus aureus infections. J. Infect. Dis. 2013, 208, 2058–2066. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, I.M.; Verdrengh, M.; Ulrich, R.G.; Bavari, S.; Tarkowski, A. Protection against Staphylococcus aureus sepsis by vaccination with recombinant staphylococcal enterotoxin A devoid of superantigenicity. J. Infect. Dis. 1999, 180, 1370–1373. [Google Scholar] [CrossRef]
- Narita, K.; Hu, D.-L.; Asano, K.; Nakane, A. Vaccination with non-toxic mutant toxic shock syndrome toxin-1 induces IL-17-dependent protection against Staphylococcus aureus infection. Pathog. Dis. 2015, 73. [Google Scholar] [CrossRef] [Green Version]
- Bohach, G.A.; Hovde, C.J.; Handley, J.P.; Schlievert, P.M. Cross-neutralization of staphylococcal and streptococcal pyrogenic toxins by monoclonal and polyclonal antibodies. Infect. Immun. 1988, 56, 400–404. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.F.; Miller, L.; Bennett, R.W.; MacMillan, J.D. Development of a monoclonal antibody capable of interacting with five serotypes of Staphylococcus aureus enterotoxin. Appl. Environ. Microbiol. 1984, 47, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Bavari, S.; Ulrich, R.G.; LeClaire, R.D. Cross-reactive antibodies prevent the lethal effects of Staphylococcus aureus superantigens. J. Infect. Dis. 1999, 180, 1365–1369. [Google Scholar] [CrossRef] [PubMed]
- Kum, W.W.; Chow, A.W. Inhibition of staphylococcal enterotoxin A-induced superantigenic and lethal activities by a monoclonal antibody to toxic shock syndrome toxin-1. J. Infect. Dis. 2001, 183, 1739–1748. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, K.; Charles, A.; Bannan, J.; Pugach, P.; Kashfi, K.; Zabriskie, J.B. Inhibition of bacterial superantigens by peptides and antibodies. Infect. Immun. 2001, 69, 875–884. [Google Scholar] [CrossRef] [Green Version]
- Venkatasubramaniam, A.; Adhikari, R.P.; Kort, T.; Liao, G.C.; Conley, S.; Abaandou, L.; Kailasan, S.; Onodera, Y.; Krishnan, S.; Djagbare, D.M.; et al. TBA225, a fusion toxoid vaccine for protection and broad neutralization of staphylococcal superantigens. Sci. Rep. 2019, 9, 3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sollid, J.U.E.; Furberg, A.S.; Hanssen, A.M.; Johannessen, M. Staphylococcus aureus: Determinants of human carriage. Infect. Genet. Evol. 2014, 21, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Song, W.-J.; Sintobin, I.; Sohn, K.-H.; Kang, M.-G.; Park, H.-K.; Jo, E.-J.; Lee, S.-E.; Yang, M.-S.; Kim, S.-H.; Kwon, Y.E.; et al. Staphylococcal enterotoxin IgE sensitization in late-onset severe eosinophilic asthma in the elderly. Clin. Exp. Allergy 2016, 46, 411–421. [Google Scholar] [CrossRef]
- Bachert, C.; van Steen, K.; Zhang, N.; Holtappels, G.; Cattaert, T.; Maus, B.; Buhl, R.; Taube, C.; Korn, S.; Kowalski, M.; et al. Specific IgE against Staphylococcus aureus enterotoxins: An independent risk factor for asthma. J. Allergy Clin. Immunol. 2012, 130, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Van Zele, T.; Gevaert, P.; Watelet, J.-B.; Claeys, G.; Holtappels, G.; Claeys, C.; van Cauwenberge, P.; Bachert, C. Staphylococcus aureus colonization and IgE antibody formation to enterotoxins is increased in nasal polyposis. J. Allergy Clin. Immunol. 2004, 114, 981–983. [Google Scholar] [CrossRef]
- Corriveau, M.-N.; Zhang, N.; Holtappels, G.; van Roy, N.; Bachert, C. Detection of Staphylococcus aureus in nasal tissue with peptide nucleic acid-fluorescence in situ hybridization. Am. J. Rhinol. Allergy 2009, 23, 461–465. [Google Scholar] [CrossRef]
- Stentzel, S.; Teufelberger, A.; Nordengrün, M.; Kolata, J.; Schmidt, F.; van Crombruggen, K.; Michalik, S.; Kumpfmüller, J.; Tischer, S.; Schweder, T.; et al. Staphylococcal serine protease-like proteins are pacemakers of allergic airway reactions to Staphylococcus aureus. J. Allergy Clin. Immunol. 2017, 139, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Teufelberger, A.R.; Nordengrün, M.; Braun, H.; Maes, T.; de Grove, K.; Holtappels, G.; O’Brien, C.; Provoost, S.; Hammad, H.; Gonçalves, A.; et al. The IL-33/ST2 axis is crucial in type 2 airway responses induced by Staphylococcus aureus-derived serine protease-like protein D. J. Allergy Clin. Immunol. 2018, 141, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachert, C.; Zhang, N. Chronic rhinosinusitis and asthma: Novel understanding of the role of IgE ‘above atopy’. J. Intern. Med. 2012, 272, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Bachert, C.; Claeys, S.E.M.; Tomassen, P.; van Zele, T.; Zhang, N. Rhinosinusitis and asthma: A link for asthma severity. Curr. Allergy Asthma Rep. 2010, 10, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Tomassen, P.; Vandeplas, G.; van Zele, T.; Cardell, L.-O.; Arebro, J.; Olze, H.; Förster-Ruhrmann, U.; Kowalski, M.L.; Olszewska-Ziąber, A.; Holtappels, G.; et al. Inflammatory endotypes of chronic rhinosinusitis based on cluster analysis of biomarkers. J. Allergy Clin. Immunol. 2016, 137, 1449–1456. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.F.; Ludwig, S.; Brigham, E.P.; McCormack, M.C.; Matsui, E.C. Effect of home exposure to Staphylococcus aureus on asthma in adolescents. J. Allergy Clin. Immunol. 2018, 141, 402–405. [Google Scholar] [CrossRef] [Green Version]
- Dzoro, S.; Mittermann, I.; Resch-Marat, Y.; Vrtala, S.; Nehr, M.; Hirschl, A.M.; Wikberg, G.; Lundeberg, L.; Johansson, C.; Scheynius, A.; et al. House dust mites as potential carriers for IgE sensitization to bacterial antigens. Allergy 2018, 73, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, S.; Jimenez-Bush, I.; Brigham, E.; Bose, S.; Diette, G.; McCormack, M.C.; Matsui, E.C.; Davis, M.F. Analysis of home dust for Staphylococcus aureus and staphylococcal enterotoxin genes using quantitative PCR. Sci. Total Environ. 2017, 581, 750–755. [Google Scholar] [CrossRef] [Green Version]
- Bachert, C.; Humbert, M.; Hanania, N.A.; Zhang, N.; Holgate, S.; Buhl, R.; Bröker, B.M. Staphylococcus aureus and its IgE-inducing enterotoxins in asthma: Current knowledge. Eur. Respir. J. 2020. [Google Scholar] [CrossRef]
- Rha, M.-S.; Kim, S.-W.; Chang, D.-Y.; Lee, J.-K.; Kim, J.; Park, S.-H.; Khamulratova, R.; Lim, H.-S.; Eun, K.M.; Hong, S.-N.; et al. Superantigen-related TH2 CD4+ T cells in nonasthmatic chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Tomassen, P.; Jarvis, D.; Newson, R.; van Ree, R.; Forsberg, B.; Howarth, P.; Janson, C.; Kowalski, M.L.; Krämer, U.; Matricardi, P.M.; et al. Staphylococcus aureus enterotoxin-specific IgE is associated with asthma in the general population: A GA(2)LEN study. Allergy 2013, 68, 1289–1297. [Google Scholar] [CrossRef]
- Kowalski, M.L.; Cieślak, M.; Pérez-Novo, C.A.; Makowska, J.S.; Bachert, C. Clinical and immunological determinants of severe/refractory asthma (SRA): Association with staphylococcal superantigen-specific IgE antibodies. Allergy 2011, 66, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, J.; Mailaender, C. The identify project: Do we know the most prevalent allergens in severe asthma? In A101. Mechanisms and Clinical Features; American Thoracic Society: New York, NY, USA, 2018; p. 2493. [Google Scholar]
- Sintobin, I.; Siroux, V.; Holtappels, G.; Pison, C.; Nadif, R.; Bousquet, J.; Bachert, C. Sensitisation to staphylococcal enterotoxins and asthma severity: A longitudinal study in the EGEA cohort. Eur. Respir. J. 2019, 54. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.N.; Shin, Y.S.; Yoo, H.-S.; Nam, Y.H.; Jin, H.J.; Ye, Y.-M.; Nahm, D.-H.; Park, H.-S. The prevalence of serum specific IgE to superantigens in asthma and allergic rhinitis patients. Allergy Asthma Immunol. Res. 2014, 6, 263–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastacaldi, C.; Lewis, P.; Howarth, P. Staphylococci and staphylococcal superantigens in asthma and rhinitis: A systematic review and meta-analysis. Allergy 2011, 66, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Takishita, T.; Yamamoto, T.; Hattori, H.; Yamashita, Y.; Nishioka, S.; Ogawa, T.; Nishizaki, K. Presence and characterization of sensitization to staphylococcal enterotoxins in patients with allergic Rhinitis. Am. J. Rhinol. 2001, 15, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, M.; Klingenberg, C.; Wickman, M.; Sollid, J.U.E.; Furberg, A.-S.; Bachert, C.; Bousquet, J. Staphylococcus aureus enterotoxin sensitization is associated with allergic poly-sensitization and allergic multimorbidity in adolescents. Allergy 2017, 72, 1548–1555. [Google Scholar] [CrossRef]
- Tsilochristou, O.; Du Toit, G.; Sayre, P.H.; Roberts, G.; Lawson, K.; Sever, M.L.; Bahnson, H.T.; Radulovic, S.; Basting, M.; Plaut, M.; et al. Association of Staphylococcus aureus colonization with food allergy occurs independently of eczema severity. J. Allergy Clin. Immunol. 2019, 144, 494–503. [Google Scholar] [CrossRef]
- Tuffs, S.W.; Haeryfar, S.M.M.; McCormick, J.K. Manipulation of innate and adaptive immunity by staphylococcal superantigens. Pathogens 2018, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.; Dauwalder, O.; Brun, V.; Badiou, C.; Ferry, T.; Etienne, J.; Vandenesch, F.; Lina, G. Staphylococcus aureus superantigens elicit redundant and extensive human Vbeta patterns. Infect. Immun. 2009, 77, 2043–2050. [Google Scholar] [CrossRef] [Green Version]
- Kappler, J.; Kotzin, B.; Herron, L.; Gelfand, E.W.; Bigler, R.D.; Boylston, A.; Carrel, S.; Posnett, D.N.; Choi, Y.; Marrack, P. V beta-specific stimulation of human T cells by staphylococcal toxins. Science 1989, 244, 811–813. [Google Scholar] [CrossRef]
- Herrmann, T.; Maryanski, J.L.; Romero, P.; Fleischer, B.; MacDonald, H.R. Activation of MHC class I-restricted CD8+ CTL by microbial T cell mitogens. Dependence upon MHC class II expression of the target cells and V beta usage of the responder T cells. J. Immunol. 1990, 144, 1181–1186. [Google Scholar] [PubMed]
- Arad, G.; Levy, R.; Nasie, I.; Hillman, D.; Rotfogel, Z.; Barash, U.; Supper, E.; Shpilka, T.; Minis, A.; Kaempfer, R. Binding of superantigen toxins into the CD28 homodimer interface is essential for induction of cytokine genes that mediate lethal shock. PLoS Biol. 2011, 9, e1001149. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.; Rotfogel, Z.; Hillman, D.; Popugailo, A.; Arad, G.; Supper, E.; Osman, F.; Kaempfer, R. Superantigens hyperinduce inflammatory cytokines by enhancing the B7-2/CD28 costimulatory receptor interaction. Proc. Natl. Acad. Sci. USA 2016, 113, E6437–E6446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llewelyn, M.; Sriskandan, S.; Peakman, M.; Ambrozak, D.R.; Douek, D.C.; Kwok, W.W.; Cohen, J.; Altmann, D.M. HLA class II polymorphisms determine responses to bacterial superantigens. J. Immunol. 2004, 172, 1719–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiedemann, R.E.; Fraser, J.D. Cross-linking of MHC class II molecules by staphylococcal enterotoxin A is essential for antigen-presenting cell and T cell activation. J. Immunol. 1996, 157, 3958–3966. [Google Scholar] [PubMed]
- Rocha, N.; Neefjes, J. MHC class II molecules on the move for successful antigen presentation. EMBO J. 2008, 27, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Elgueta, R.; Benson, M.J.; de Vries, V.C.; Wasiuk, A.; Guo, Y.; Noelle, R.J. Molecular mechanism and function of CD40/CD40L engagement in the immune system. Immunol. Rev. 2009, 229, 152–172. [Google Scholar] [CrossRef] [Green Version]
- Takatsu, K. Cytokines involved in B-cell differentiation and their sites of action. Proc. Soc. Exp. Biol. Med. 1997, 215, 121–133. [Google Scholar] [CrossRef]
- Stavnezer, J. Immunoglobulin class switching. Curr. Opin. Immunol. 1996, 8, 199–205. [Google Scholar] [CrossRef]
- Tangye, S.G.; Ferguson, A.; Avery, D.T.; Ma, C.S.; Hodgkin, P.D. Isotype switching by human B cells is division-associated and regulated by cytokines. J. Immunol. 2002, 169, 4298–4306. [Google Scholar] [CrossRef]
- Bergthorsdottir, S.; Gallagher, A.; Jainandunsing, S.; Cockayne, D.; Sutton, J.; Leanderson, T.; Gray, D. Signals that initiate somatic hypermutation of B cells in vitro. J. Immunol. 2001, 166, 2228–2234. [Google Scholar] [CrossRef] [Green Version]
- Maddaly, R.; Pai, G.; Balaji, S.; Sivaramakrishnan, P.; Srinivasan, L.; Sunder, S.S.; Paul, S.F.D. Receptors and signaling mechanisms for B-lymphocyte activation, proliferation and differentiation—Insights from both in vivo and in vitro approaches. FEBS Lett. 2010, 584, 4883–4894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabara, H.H.; Geha, R.S. The superantigen toxic shock syndrome toxin-1 induces CD40 ligand expression and modulates IgE isotype switching. Int. Immunol. 1996, 8, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.G.; Johnson, H.M. The effect of staphylococcal enterotoxins on the primary in vitro immune response. J. Immunol. 1975, 115, 575–578. [Google Scholar] [PubMed]
- Kawaguchi-Nagata, K.; Okamura, H.; Shoji, K.; Kanagawa, H.; Semma, M.; Shinagawa, K. Immunomodulating activities of staphylococcal enterotoxins. Microbiol. Immunol. 1985, 29, 183–193. [Google Scholar] [CrossRef]
- Poindexter, N.J.; Schlievert, P.M. Suppression of immunoglobulin-secreting cells from human peripheral blood by toxic-shock-syndrome toxin-1. J. Infect. Dis. 1986, 153, 772–779. [Google Scholar] [CrossRef]
- Moseley, A.B.; Huston, D.P. Mechanism of Staphylococcus aureus exotoxin A inhibition of Ig production by human B cells. J. Immunol. 1991, 146, 826–832. [Google Scholar]
- Mourad, W.; Scholl, P.; Diaz, A.; Geha, R.; Chatila, T. The staphylococcal toxic shock syndrome toxin 1 triggers B cell proliferation and differentiation via major histocompatibility complex-unrestricted cognate T/B cell interaction. J. Exp. Med. 1989, 170, 2011–2022. [Google Scholar] [CrossRef] [Green Version]
- Crow, M.K.; Zagon, G.; Chu, Z.; Ravina, B.; Tumang, J.R.; Cole, B.C.; Friedman, S.M. Human B cell differentiation induced by microbial superantigens: Unselected peripheral blood lymphocytes secrete polyclonal immunoglobulin in response to Mycoplasma arthritidis mitogen. Autoimmunity 1992, 14, 23–32. [Google Scholar] [CrossRef]
- Stohl, W.; Elliott, J.E.; Linsley, P.S. Human T cell-dependent B cell differentiation induced by staphylococcal superantigens. J. Immunol. 1994, 153, 117–127. [Google Scholar]
- Takei, S.; Arora, Y.K.; Walker, S.M. Intravenous immunoglobulin contains specific antibodies inhibitory to activation of T cells by staphylococcal toxin superantigens see comment. J. Clin. Investig. 1993, 91, 602–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J. T helper cell differentiation, heterogeneity, and plasticity. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lederman, S.; Yellin, M.J.; Krichevsky, A.; Belko, J.; Lee, J.J.; Chess, L. Identification of a novel surface protein on activated CD4+ T cells that induces contact-dependent B cell differentiation (help). J. Exp. Med. 1992, 175, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Gould, H.J.; Sutton, B.J.; Beavil, A.J.; Beavil, R.L.; McCloskey, N.; Coker, H.A.; Fear, D.; Smurthwaite, L. The biology of IGE and the basis of allergic disease. Annu. Rev. Immunol. 2003, 21, 579–628. [Google Scholar] [CrossRef]
- Hofer, M.F.; Harbeck, R.J.; Schlievert, P.M.; Leung, D.Y. Staphylococcal toxins augment specific IgE responses by atopic patients exposed to allergen. J. Investig. Dermatol. 1999, 112, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Bouvet, J.-P.; Marone, G. Protein Fv: An endogenous immunoglobulin superantigen and superallergen. Chem. Immunol. Allergy 2007, 93, 58–76. [Google Scholar] [CrossRef]
- Marone, G.; Rossi, F.W.; Detoraki, A.; Granata, F.; Marone, G.; Genovese, A.; Spadaro, G. Role of superallergens in allergic disorders. Chem. Immunol. Allergy 2007, 93, 195–213. [Google Scholar] [CrossRef]
- Leung, D.Y.; Harbeck, R.; Bina, P.; Reiser, R.F.; Yang, E.; Norris, D.A.; Hanifin, J.M.; Sampson, H.A. Presence of IgE antibodies to staphylococcal exotoxins on the skin of patients with atopic dermatitis. Evidence for a new group of allergens. J. Clin. Investig. 1993, 92, 1374–1380. [Google Scholar] [CrossRef] [Green Version]
- Bunikowski, R.; Mielke, M.; Skarabis, H.; Herz, U.; Bergmann, R.L.; Wahn, U.; Renz, H. Prevalence and role of serum IgE antibodies to the Staphylococcus aureus—Derived superantigens SEA and SEB in children with atopic dermatitis. J. Allergy Clin. Immunol. 1999, 103, 119–124. [Google Scholar] [CrossRef]
- Ong, P.Y.; Patel, M.; Ferdman, R.M.; Dunaway, T.; Church, J.A. Association of staphylococcal superantigen-specific immunoglobulin e with mild and moderate atopic dermatitis. J. Pediatr. 2008, 153, 803–806. [Google Scholar] [CrossRef] [Green Version]
- Bachert, C.; Gevaert, P.; Holtappels, G.; Johansson, S.G.; van Cauwenberge, P. Total and specific IgE in nasal polyps is related to local eosinophilic inflammation. J. Allergy Clin. Immunol. 2001, 107, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Huvenne, W.; Hellings, P.W.; Bachert, C. Role of staphylococcal superantigens in airway disease. Int. Arch. Allergy Immunol. 2013, 161, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Patou, J.; Gevaert, P.; van Zele, T.; Holtappels, G.; van Cauwenberge, P.; Bachert, C. Staphylococcus aureus enterotoxin B, protein A, and lipoteichoic acid stimulations in nasal polyps. J. Allergy Clin. Immunol. 2008, 121, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Scheuber, P.H.; Golecki, J.R.; Kickhöfen, B.; Scheel, D.; Beck, G.; Hammer, D.K. Skin reactivity of unsensitized monkeys upon challenge with staphylococcal enterotoxin B: A new approach for investigating the site of toxin action. Infect. Immun. 1985, 50, 869–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komisar, J.; Rivera, J.; Vega, A.; Tseng, J. Effects of staphylococcal enterotoxin B on rodent mast cells. Infect. Immun. 1992, 60, 2969–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, H.K.; Hirose, S.; Narita, K.; Sugiyama, M.; Asano, K.; Hu, D.-L.; Nakane, A. Histamine release from intestinal mast cells induced by staphylococcal enterotoxin A (SEA) evokes vomiting reflex in common marmoset. PLoS Pathog. 2019, 15, e1007803. [Google Scholar] [CrossRef] [Green Version]
- Ono, H.K.; Nishizawa, M.; Yamamoto, Y.; Hu, D.-L.; Nakane, A.; Shinagawa, K.; Omoe, K. Submucosal mast cells in the gastrointestinal tract are a target of staphylococcal enterotoxin type A. FEMS Immunol. Med. Microbiol. 2012, 64, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Hayes, S.M.; Biggs, T.C.; Goldie, S.P.; Harries, P.G.; Walls, A.F.; Allan, R.N.; Pender, S.L.F.; Salib, R.J. Staphylococcus aureus internalization in mast cells in nasal polyps: Characterization of interactions and potential mechanisms. J. Allergy Clin. Immunol. 2020, 145, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-B.; James, L.K.; Davies, A.M.; Wu, Y.-C.B.; Rimmer, J.; Lund, V.J.; Chen, J.-H.; McDonnell, J.M.; Chan, Y.-C.; Hutchins, G.H.; et al. Antibodies and superantibodies in patients with chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2017, 139, 1195–1204. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.R.O.; Janik, D.K.; Lee, W.T. Superantigen-induced CD4 memory T cell anergy. I. Staphylococcal enterotoxin B induces Fyn-mediated negative signaling. Cell. Immunol. 2012, 276, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Janik, D.K.; Lee, W.T. Staphylococcal enterotoxin B (SEB) induces memory CD4 T cell anergy in vivo and impairs recall immunity to unrelated antigens. J. Clin. Cell. Immunol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.X.; Kasper, K.J.; Zeppa, J.J.; McCormick, J.K. Superantigens modulate bacterial density during Staphylococcus aureus nasal colonization. Toxins 2015, 7, 1821–1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordengrün, M.; Michalik, S.; Völker, U.; Bröker, B.M.; Gómez-Gascón, L. The quest for bacterial allergens. Int. J. Med. Microbiol. 2018, 308, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-C.; Won, H.-K.; Lee, J.W.; Sohn, K.-H.; Kim, M.-H.; Kim, T.-B.; Chang, Y.-S.; Lee, B.-J.; Cho, S.-H.; Bachert, C.; et al. Staphylococcus aureus nasal colonization and asthma in adults: Systematic review and meta-analysis. J. Allergy Clin. Immunol. Pract. 2019, 7, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Seiti Yamada Yoshikawa, F.; Feitosa de Lima, J.; Notomi Sato, M.; Álefe Leuzzi Ramos, Y.; Aoki, V.; Leao Orfali, R. Exploring the role of Staphylococcus aureus toxins in atopic dermatitis. Toxins 2019, 11, 321. [Google Scholar] [CrossRef] [Green Version]
- Abdurrahman, G.; Bröker, B. The impact of S. aureus and its proteins. In Precision Medicine in Chronic Rhinosinusitis; Zhang, L., Bachert, C., Eds.; Springer: New York, NY, USA, 2020. (in press) [Google Scholar]



{kind=link}
{kind=link}
| Vaccination Type | Target | Name (Company; NCT Number 1) | Study Design | Status and Study Results | Intervention | Duration | Ref. |
|---|---|---|---|---|---|---|---|
| Active | SEB 2 | STEBVax (Integrated BioTherapeu- tics; NCT00974935) | Non-randomized, dose escalation | Phase I, completed. STEBvax was safe, well-tolerated and immunogenic, induced/boosted toxin-neutralizing antibodies | STEBVax vaccine 3 with Alhydrogel adjuvant, six doses (10 ng–20 µg) or 20 µg given in two vaccinations 21 days apart | 02/11- 03/15 | [48] |
| Active | TSST-1 4 | rTSST-1v 5 (Biomedizi- nische Forschungs GmbH; NCT02340338) | Randomized, double-blind, adjuvant- controlled dose escalation | Phase I, completed. rTSST-1v was safe, well-tolerated, and immunogenic, induced/boosted toxin-neutralizing antibodies | rTSST-1 variant 5 with Al(OH)3, six doses in one to two vaccinations (100 ng–30 µg) | 06/14- 06/15 | [49] |
| Active | TSST-1 | rTSST-1v (Biomedizi- nische Forschungs GmbH; NCT02814708) | Randomized, placebo- controlled | Phase II, ongoing | rTSST-1 variant 3 with Al(OH)3, two doses (10, 100 µg) in one to three vaccinations | Since 03/16 | - |
| Passive | SAgs | IVIG 6 (Hospices Civils de Lyon; NCT02219165) | Randomized, placebo- controlled | Phase II, completed | IVIG (single dose, 2 g/kg) | Since 01/15 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdurrahman, G.; Schmiedeke, F.; Bachert, C.; Bröker, B.M.; Holtfreter, S. Allergy—A New Role for T Cell Superantigens of Staphylococcus aureus? Toxins 2020, 12, 176. https://doi.org/10.3390/toxins12030176
Abdurrahman G, Schmiedeke F, Bachert C, Bröker BM, Holtfreter S. Allergy—A New Role for T Cell Superantigens of Staphylococcus aureus? Toxins. 2020; 12(3):176. https://doi.org/10.3390/toxins12030176
Chicago/Turabian StyleAbdurrahman, Goran, Frieder Schmiedeke, Claus Bachert, Barbara M. Bröker, and Silva Holtfreter. 2020. "Allergy—A New Role for T Cell Superantigens of Staphylococcus aureus?" Toxins 12, no. 3: 176. https://doi.org/10.3390/toxins12030176
APA StyleAbdurrahman, G., Schmiedeke, F., Bachert, C., Bröker, B. M., & Holtfreter, S. (2020). Allergy—A New Role for T Cell Superantigens of Staphylococcus aureus? Toxins, 12(3), 176. https://doi.org/10.3390/toxins12030176