Toxicity of Recombinant Necrosis and Ethylene-Inducing Proteins (NLPs) from Neofusicoccum parvum

 ,
, 
 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Culture Filtrate of N. parvum on Detached Tomato Leaves
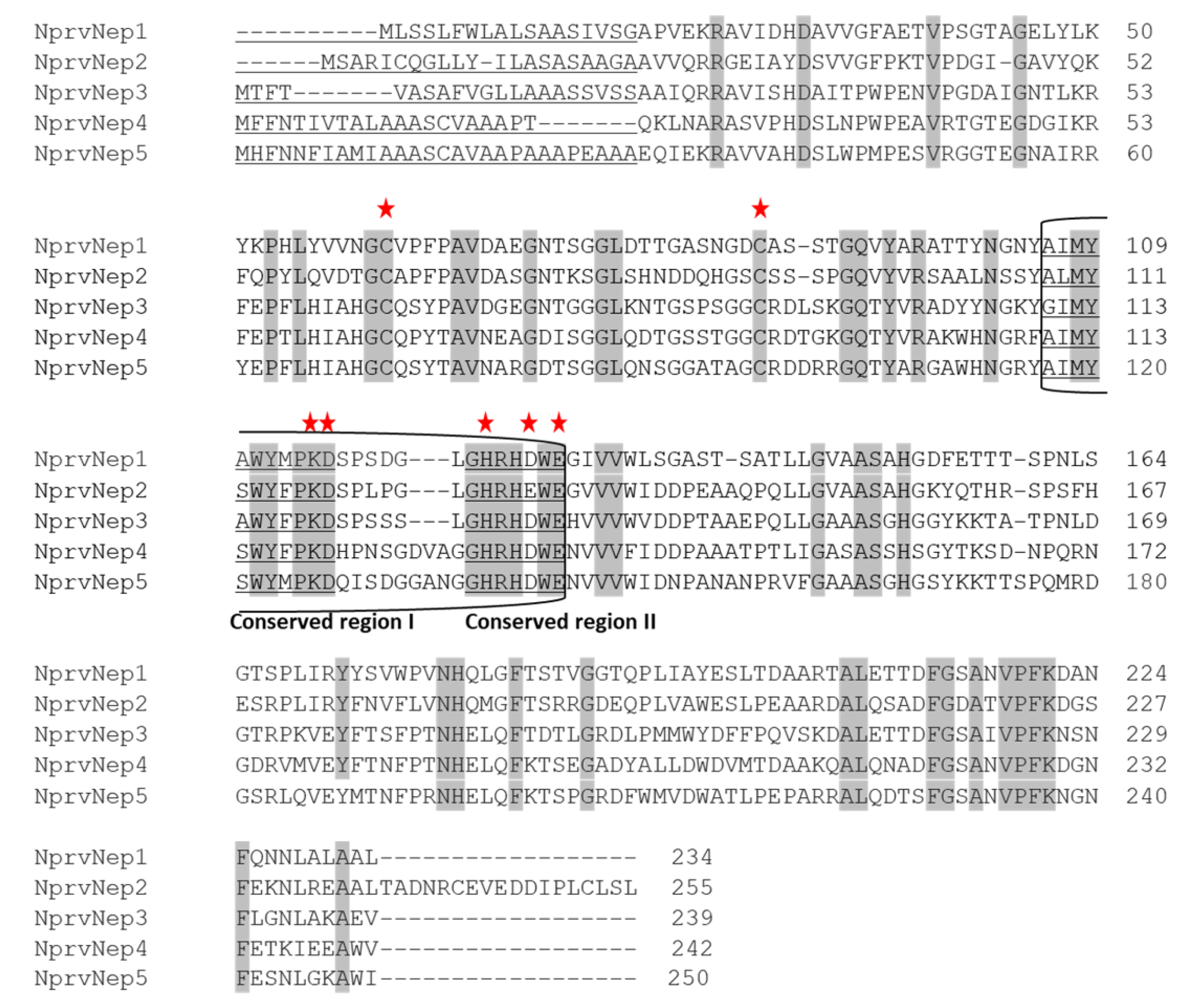
2.2. NprvNep Proteins’ Sequence Analysis
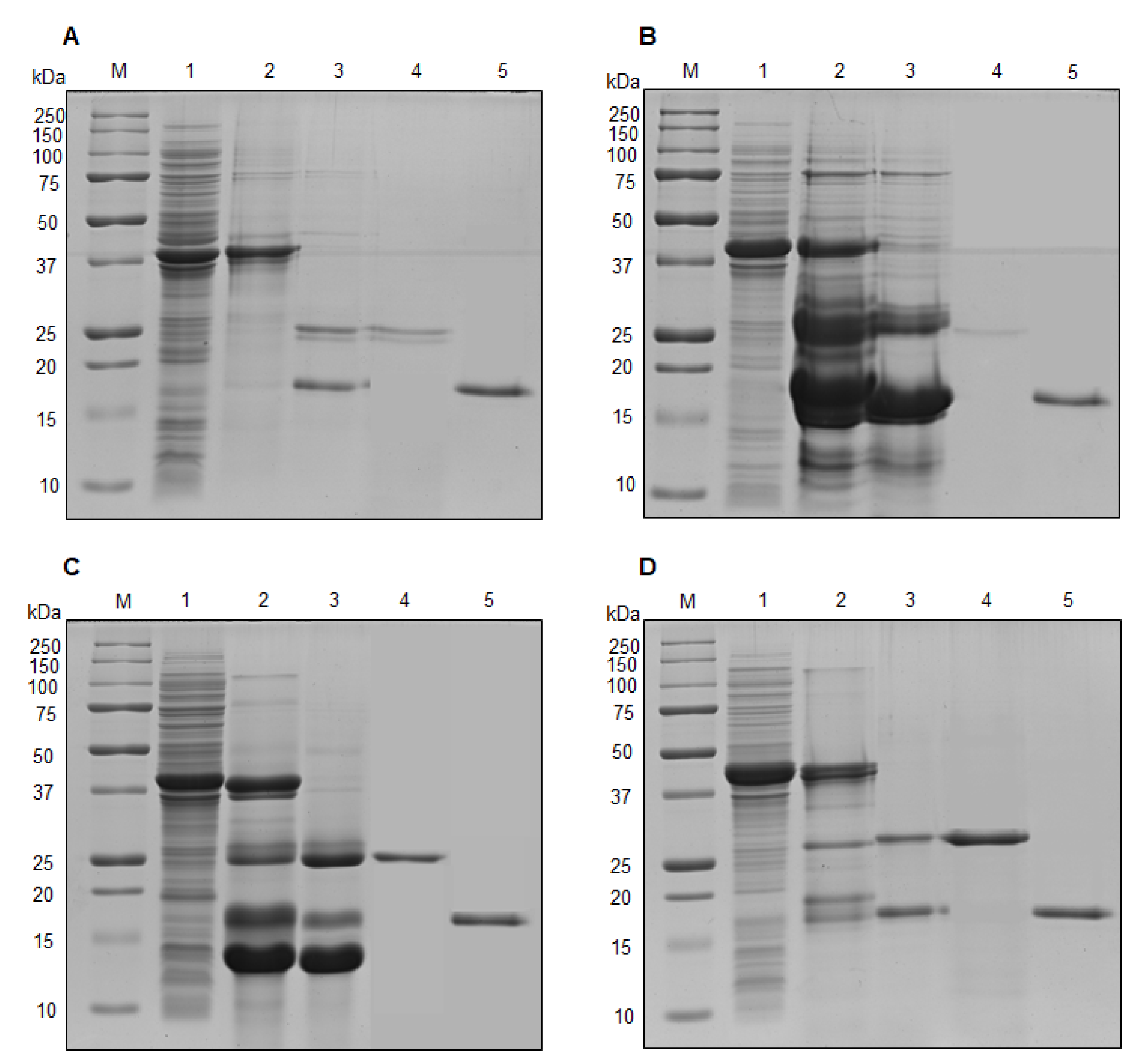
2.3. Cloning, Expression and Purification of NprvNep Proteins
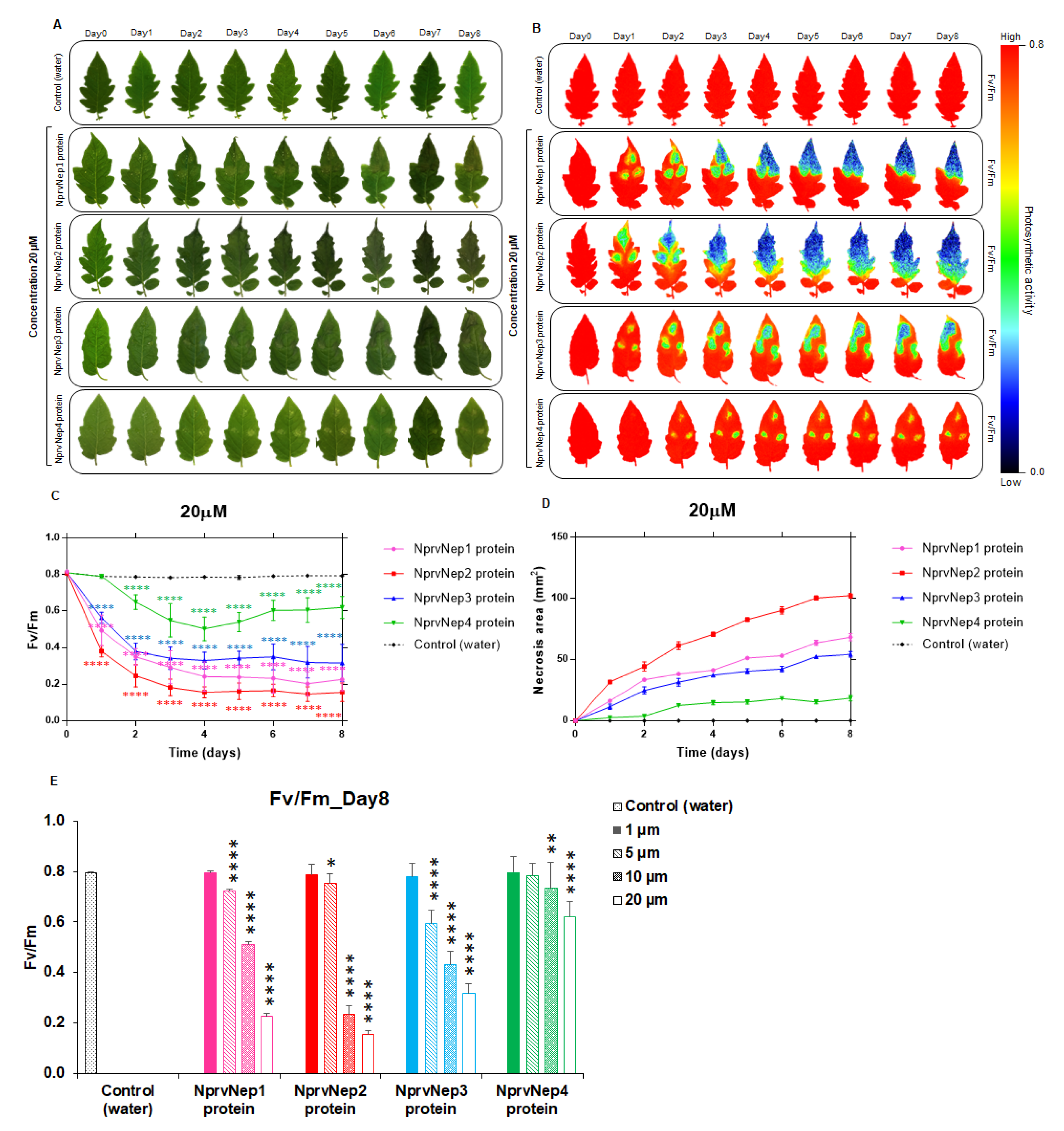
2.4. Activity of NprvNep Proteins—Toxicity to Tomato Leaves
2.5. Activity of NprvNep Proteins—Effect of NprvNep Proteins on Chlorophyll Fluorescence
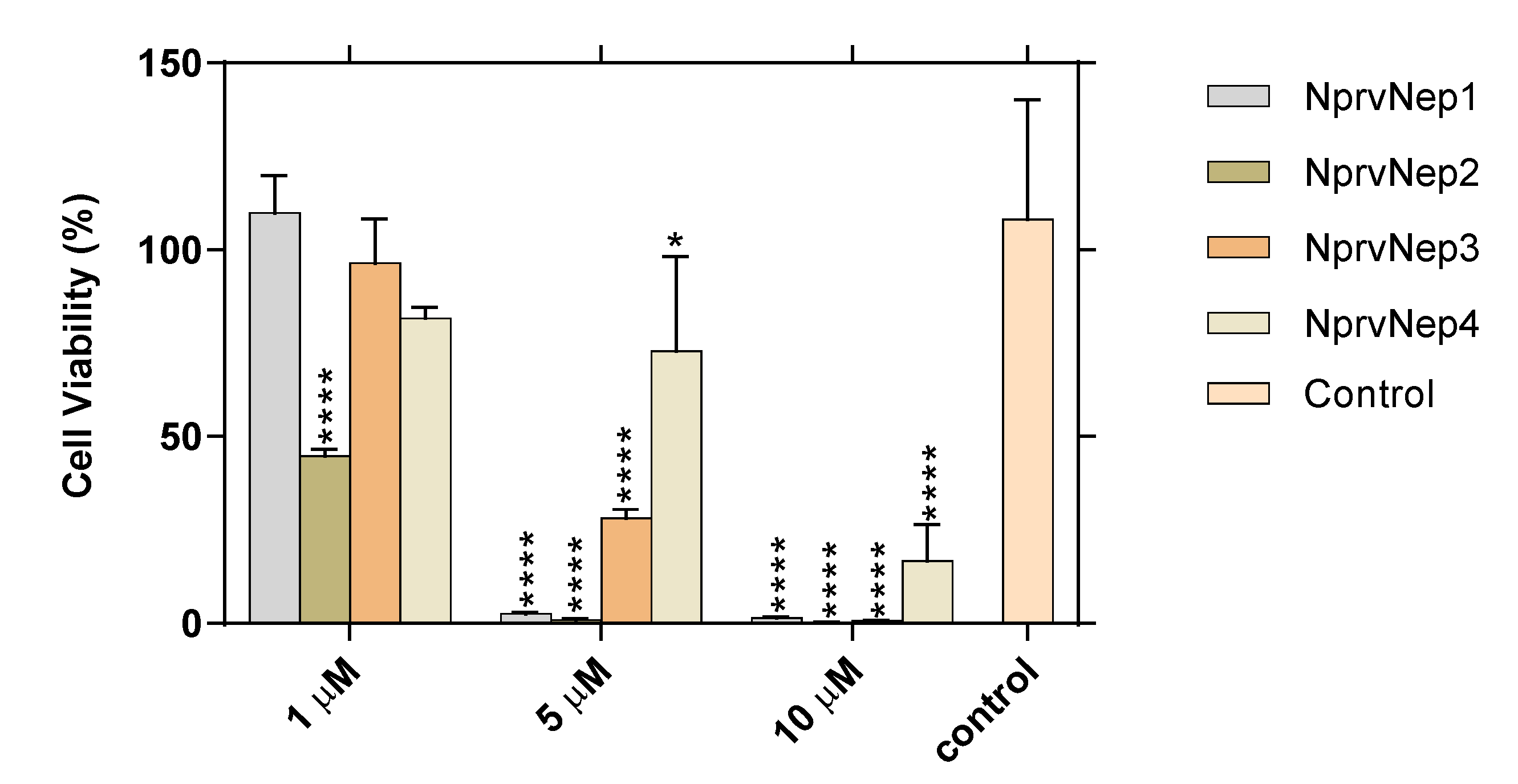
2.6. Activity of NprvNep Proteins—Toxicity to Vero Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strain and Plant Material
5.2. DNA and RNA Extraction and cDNA Synthesis
5.3. Cloning, Expression and Purification of Recombinant NprvNeps
5.4. Protein Concentration
5.5. Phytotoxic Activity
5.6. Chlorophyll Fluorescence Imaging
5.7. Cytotoxicity Assay
5.8. Bioinformatics Analysis
5.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennakoon, K.M.S.; Ridgway, H.J.; Jaspers, M.V.; Langford, G.; Eirian Jones, E. Evaluation of fungicide efficacy against Neofusicoccum species causing dieback disease of blueberries in New Zealand. Australas. Plant Pathol. 2019, 48, 75–84. [Google Scholar] [CrossRef]
- McDonald, V.; Lynch, S.; Eskalen, A. First Report of Neofusicoccum australe, N. luteum, and N. parvum Associated With Avocado Branch Canker in California. Plant Dis. 2009, 93, 967. [Google Scholar] [CrossRef] [PubMed]
- Larignon, P.; Dubos, B. Fungi associated with esca disease in grapevine. Eur. J. Plant Pathol. 1997, 103, 147–157. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (Black Measles) and brown wood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [Green Version]
- Úrbez-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50, S5–S45. [Google Scholar] [CrossRef]
- Andolfi, A.; Mugnai, L.; Luque, J.; Surico, G.; Cimmino, A.; Evidente, A. Phytotoxins produced by fungi associated with grapevine trunk diseases. Toxins 2011, 3, 1569–1605. [Google Scholar] [CrossRef] [Green Version]
- Abou-Mansour, E.; Debieux, J.L.; Ramirez-Suero, M.; Benard-Gellon, M.; Magnin-Robert, M.; Spagnolo, A.; Chong, J.L.; Farine, S.; Bertsch, C.; L’Haridon, F.; et al. Phytotoxic metabolites from Neofusicoccum parvum, a pathogen of Botryosphaeria dieback of grapevine. Phytochemistry 2015, 115, 207–215. [Google Scholar] [CrossRef]
- Benard-Gellon, M.; Farine, S.; Goddard, M.L.; Schmitt, M.; Stempien, E.; Pensec, F.; Laloue, H.; Mazet-Kieffer, F.; Fontaine, F.; Larignon, P.; et al. Toxicity of extracellular proteins from Diplodia seriata and Neofusicoccum parvum involved in grapevine Botryosphaeria dieback. Protoplasma 2015, 252, 679–687. [Google Scholar] [CrossRef]
- Li, G.Q.; Liu, F.F.; Li, J.Q.; Liu, Q.L.; Chen, S.F. Botryosphaeriaceae from Eucalyptus plantations and adjacent plants in China. Persoonia Mol. Phylogeny Evol. Fungi 2018, 40, 63–95. [Google Scholar] [CrossRef] [Green Version]
- Serrato-Diaz, L.M.; Aviles-Noriega, A.; Soto-Bauzó, A.; Rivera-Vargas, L.I.; Goenaga, R.; Bayman, P. Botryosphaeriaceae Fungi as Causal Agents of Dieback and Corky Bark in Rambutan and Longan. Plant Dis. 2019, 104, 105–115. [Google Scholar] [CrossRef]
- Valencia, A.L.; Gil, P.M.; Latorre, B.A.; Rosales, I.M. Characterization and Pathogenicity of Botryosphaeriaceae Species Obtained from Avocado Trees with Branch Canker and Dieback and from Avocado Fruit with Stem End Rot in Chile. Plant Dis. 2019, 103, 996–1005. [Google Scholar] [CrossRef]
- Cobos, R.; Barreiro, C.; Mateos, R.M.; Coque, J.J.R. Cytoplasmic- and extracellular-proteome analysis of Diplodia seriata: A phytopathogenic fungus involved in grapevine decline. Proteome Sci. 2010, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimchuk, Z.; Eulgem, T.; Holt, B.F.; Dangl, J.L. Recognition and response in the plant immune system. Ann. Rev. Genet. 2003, 37, 579–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.; Kim, M.S.; Sicher, R.C.; Bae, H.-J.; Bailey, B.A. Necrosis- and ethylene-inducing peptide from Fusarium oxysporum induces a complex cascade of transcripts associated with signal transduction and cell death in Arabidopsis. Plant Physiol. 2006, 141, 1056–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, B.A. Purification of a protein from culture filtrates of Fusarium oxysporum that induces ethylene and necrosis in leaves of Erythroxylum coca. Phytopathology 1995, 85, 1250–1255. [Google Scholar] [CrossRef]
- Gijzen, M.; Nürnberger, T. Nep1-like proteins from plant pathogens: Recruitment and diversification of the NPP1 domain across taxa. Phytochemistry 2006, 67, 1800–1807. [Google Scholar] [CrossRef]
- Pemberton, C.L.; Salmond, G.P.C. The Nep1-like proteins-a growing family of microbial elicitors of plant necrosis. Mol. Plant Pathol. 2004, 5, 353–359. [Google Scholar] [CrossRef]
- Bailey, B.A.; Bae, H.; Strem, M.D.; de Mayolo, G.A.; Guiltinan, M.J.; Verica, J.A.; Maximova, S.N.; Bowers, J.H. Developmental expression of stress response genes in Theobroma cacao leaves and their response to Nep1 treatment and a compatible infection by Phytophthora megakarya. Plant Physiol. Biochem. 2005, 43, 611–622. [Google Scholar] [CrossRef]
- Fellbrich, G.; Romanski, A.; Varet, A.; Blume, B.; Brunner, F.; Engelhardt, S.; Felix, G.; Kemmerling, B.; Krzymowska, M.; Nürnberger, T. NPP1, a Phytophthora-associated trigger of plant defense in parsley and Arabidopsis. Plant J. 2002, 32, 375–390. [Google Scholar] [CrossRef]
- Jennings, J.C.; Apel-Birkhold, P.C.; Mock, N.M.; Baker, C.J.; Anderson, J.D.; Bailey, B.A. Induction of defense responses in tobacco by the protein Nep1 from Fusarium oxysporum. Plant Sci. 2001, 161, 891–899. [Google Scholar] [CrossRef] [Green Version]
- Keates, S.E.; Kostman, T.A.; Anderson, J.D.; Bailey, B.A. Altered gene expression in three plant species in response to treatment with Nep1, a fungal protein that causes necrosis. Plant Physiol. 2003, 132, 1610–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veit, S.; Wörle, J.M.; Nürnberger, T.; Koch, W.; Seitz, H.U. A Novel protein elicitor (PaNie) from Pythium aphanidermatum induces multiple defense responses in carrot, Arabidopsis, and tobacco. Plant Physiol. 2001, 127, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Verica, J.A.; Maximova, S.N.; Strem, M.D.; Carlson, J.E.; Bailey, B.A.; Guiltinan, M.J. Isolation of ESTs from cacao (Theobroma cacao L.) leaves treated with inducers of the defense response. Plant Cell Rep. 2004, 23, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Qutob, D.; Kemmerling, B.; Brunner, F.; Küfner, I.; Engelhardt, S.; Gust, A.A.; Luberacki, B.; Seitz, H.U.; Stahl, D.; Rauhut, T.; et al. Phytotoxicity and innate immune responses induced by Nep1-like proteins. Plant Cell 2006, 18, 3721–3744. [Google Scholar] [CrossRef] [Green Version]
- Schouten, A.; van Baarlen, P.; van Kan, J.A.L. Phytotoxic Nep1-like proteins from the necrotrophic fungus Botrytis cinerea associate with membranes and the nucleus of plant cells. New Phytol. 2008, 177, 493–505. [Google Scholar] [CrossRef]
- Staats, M.; Van Baarlen, P.; Schouten, A.; Van Kan, J.A.L. Functional analysis of NLP genes from Botrytis elliptica. Mol. Plant Pathol. 2007, 8, 209–214. [Google Scholar] [CrossRef]
- Teh, C.-Y.; Pang, C.-L.; Tor, X.-Y.; Ho, P.-Y.; Lim, Y.-Y.; Namasivayam, P.; Ho, C.-L. Molecular cloning and functional analysis of a necrosis and ethylene inducing protein (NEP) from Ganoderma boninense. Physiol. Mol. Plant Pathol. 2019, 106, 42–48. [Google Scholar] [CrossRef]
- Lenarčič, T.; Albert, I.; Böhm, H.; Hodnik, V.; Pirc, K.; Zavec, A.B.; Podobnik, M.; Pahovnik, D.; Žagar, E.; Pruitt, R.; et al. Eudicot plant-specific sphingolipids determine host selectivity of microbial NLP cytolysins. Science 2017, 358, 1431–1434. [Google Scholar] [CrossRef] [Green Version]
- Jores, J.; Appel, B.; Lewin, A. Cloning and molecular characterization of a unique hemolysin gene of Vibrio pommerensis sp. nov.: Development of a DNA probe for the detection of the hemolysin gene and its use in identification of related Vibrio spp. from the Baltic Sea. FEMS Microbiol. Lett. 2003, 229, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Oome, S.; Van den Ackerveken, G. Comparative and functional analysis of the widely occurring family of Nep1-like proteins. Mol. Plant Microbe Interact. 2014, 27, 1081–1094. [Google Scholar] [CrossRef] [Green Version]
- Ottmann, C.; Luberacki, B.; Küfner, I.; Koch, W.; Brunner, F.; Weyand, M.; Mattinen, L.; Pirhonen, M.; Anderluh, G.; Seitz, H.U.; et al. A common toxin fold mediates microbial attack and plant defense. Proc. Natl. Acad. Sci. USA 2009, 106, 10359–10364. [Google Scholar] [CrossRef] [Green Version]
- Böhm, H.; Albert, I.; Oome, S.; Raaymakers, T.M.; Van den Ackerveken, G.; Nürnberger, T. A conserved peptide pattern from a widespread microbial virulence factor triggers pattern-induced immunity in Arabidopsis. PLoS Pathog. 2014, 10, e1004491. [Google Scholar] [CrossRef]
- Oome, S.; Raaymakers, T.M.; Cabral, A.; Samwel, S.; Böhm, H.; Albert, I.; Nürnberger, T.; Van den Ackerveken, G. Nep1-like proteins from three kingdoms of life act as a microbe-associated molecular pattern in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 16955–16960. [Google Scholar] [CrossRef] [Green Version]
- Qutob, D.; Kamoun, S.; Gijzen, M. Expression of a Phytophthora sojae necrosis-inducing protein occurs during transition from biotrophy to necrotrophy. Plant J. Cell Mol. Biol. 2002, 32, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, B.A.; Jennings, J.C.; Anderson, J.D. Sensitivity of coca (Erythroxylum coca var. coca) to ethylene and fungal proteins. Weed Sci. 1997, 45, 716–721. [Google Scholar]
- Jennings, J.C.; Apel-Birkhold, P.C.; Bailey, B.A.; Anderson, J.D. Induction of ethylene biosynthesis and necrosis in weed leaves by a Fusarium oxysporum protein. Weed Sci. 2000, 48, 7–14. [Google Scholar]
- Pemberton, C.L.; Whitehead, N.A.; Sebaihia, M.; Bell, K.S.; Hyman, L.J.; Harris, S.J.; Matlin, A.J.; Robson, N.D.; Birch, P.R.J.; Carr, J.P.; et al. Novel quorum-sensing-controlled genes in Erwinia carotovora subsp. carotovora: Identification of a fungal elicitor homologue in a soft-rotting bacterium. Mol. Plant Microbe Interact. 2005, 18, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Garcia, O.; Macedo, J.A.N.; Tibúrcio, R.; Zaparoli, G.; Rincones, J.; Bittencourt, L.M.C.; Ceita, G.O.; Micheli, F.; Gesteira, A.; Mariano, A.C.; et al. Characterization of necrosis and ethylene-inducing proteins (NEP) in the basidiomycete Moniliophthora perniciosa, the causal agent of witches’ broom in Theobroma cacao. Mycol. Res. 2007, 111, 443–455. [Google Scholar] [CrossRef]
- Amsellem, Z.; Cohen, B.A.; Gressel, J. Engineering hypervirulence in a mycoherbicidal fungus for efficient weed control. Nat. Biotechnol. 2002, 20, 1035–1039. [Google Scholar] [CrossRef]
- Santhanam, P.; van Esse, H.P.; Albert, I.; Faino, L.; Nürnberger, T.; Thomma, B.P.H.J. Evidence for functional diversification within a fungal NEP1-like protein family. Mol. Plant Microbe Interact. 2013, 26, 278–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.-L.; Peng, Y.-L.; Fan, J. The Nep1-like protein family of Magnaporthe oryzae is dispensable for the infection of rice plants. Sci. Rep. 2017, 7, 4372. [Google Scholar] [CrossRef] [PubMed]
- Motteram, J.; Küfner, I.; Deller, S.; Brunner, F.; Hammond-Kosack, K.E.; Nürnberger, T.; Rudd, J.J. Molecular characterization and functional analysis of MgNLP, the sole NPP1 domain-containing protein, from the fungal wheat leaf pathogen Mycosphaerella graminicola. Mol. Plant Microbe Interact. 2009, 22, 790–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Félix, C.; Duarte, A.S.; Vitorino, R.; Guerreiro, A.C.L.; Domingues, P.; Correia, A.C.M.; Alves, A.; Esteves, A.C. Temperature modulates the secretome of the phytopathogenic fungus Lasiodiplodia theobromae. Front. Plant Sci. 2016, 7, 1096. [Google Scholar] [CrossRef]
- Félix, C.; Meneses, R.; Gonçalves, M.F.M.; Tilleman, L.; Duarte, A.S.; Jorrín-Novo, J.V.; Van de Peer, Y.; Deforce, D.; Van Nieuwerburgh, F.; Esteves, A.C.; et al. A multi-omics analysis of the grapevine pathogen Lasiodiplodia theobromae reveals that temperature affects the expression of virulence- and pathogenicity-related genes. Sci. Rep. 2019, 9, 13144. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Ulate, B.; Rolshausen, P.; Cantu, D. Draft genome sequence of Neofusicoccum parvum isolate UCR-NP2, a fungal vascular pathogen associated with grapevine cankers. Genome Announc. 2013, 1, e00339-13. [Google Scholar] [CrossRef] [Green Version]
- Pour, F.N.; Ferreira, V.; Félix, C.; Serôdio, J.; Alves, A.; Duarte, A.S.; Esteves, A.C. Effect of temperature on the phytotoxicity and cytotoxicity of Botryosphaeriaceae fungi. Fungal Biol. 2020, in press. [Google Scholar]
- Cobos, R.; Calvo-Pena, C.; Alvarez-Perez, J.M.; Ibanez, A.; Diez-Galan, A.; Gonzalez-Garcia, S.; Garcia-Angulo, P.; Acebes, J.L.; Coque, J.J.R. Necrotic and cytolytic activity on grapevine leaves produced by Nep1-like proteins of Diplodia seriata. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.-J.; Jia, P.-S.; Gao, F.; Guo, H.-S. Molecular characterization and functional analysis of a necrosis- and ethylene-inducing, protein-encoding gene family from Verticillium dahliae. Mol. Plant Microbe Interact. MPMI 2012, 25, 964–975. [Google Scholar] [CrossRef] [Green Version]
- Küfner, I.; Ottmann, C.; Oecking, C.; Nürnberger, T. Cytolytic toxins as triggers of plant immune response. Plant Signal. Behav. 2009, 4, 977–979. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.R.; Huang, S.X.; Zhang, Y.; Sheng, G.L.; Li, Y.P.; Zhu, F. Identification and functional analysis of the NLP-encoding genes from the phytopathogenic oomycete Phytophthora capsici. Mol. Genet. Genom. 2018, 293, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.M.; Kong, G.H.; Qutob, D.; Yu, X.L.; Tang, J.L.; Kang, J.X.; Dai, T.T.; Wang, H.; Gijzen, M.; Wang, Y.C. The NLP toxin family in Phytophthora sojae includes rapidly evolving groups that lack necrosis-inducing activity. Mol. Plant Microbe Interact. 2012, 25, 896–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, A.; Oome, S.; Sander, N.; Küfner, I.; Nürnberger, T.; Van den Ackerveken, G. Nontoxic Nep1-like proteins of the downy mildew pathogen Hyaloperonospora arabidopsidis: Repression of necrosis-inducing activity by a surface-exposed region. Mol. Plant Microbe Interact. 2012, 25, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-Y.; Cai, Y.; Gou, J.-Y.; Mao, Y.-B.; Xu, Y.-H.; Jiang, W.-H.; Chen, X.-Y. VdNEP, an elicitor from Verticillium dahliae, induces cotton plant wilting. Appl. Environ. Microbiol. 2004, 70, 4989–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta Arenas, Y.; Kalkman, E.R.I.C.; Schouten, A.; Dieho, M.; Vredenbregt, P.; Uwumukiza, B.; Osés Ruiz, M.; van Kan, J.A.L. Functional analysis and mode of action of phytotoxic Nep1-like proteins of Botrytis cinerea. Physiol. Mol. Plant Pathol. 2010, 74, 376–386. [Google Scholar] [CrossRef]
- Meyer, S.; Saccardy-Adji, K.; Rizza, F.; Genty, B. Inhibition of photosynthesis by Colletotrichum lindemuthianum in bean leaves determined by chlorophyll fluorescence imaging. Plant Cell Environ. 2001, 24, 947–956. [Google Scholar] [CrossRef]
- Berger, S.; Papadopoulos, M.; Schreiber, U.; Kaiser, W.; Roitsch, T. Complex regulation of gene expression, photosynthesis and sugar levels by pathogen infection in tomato. Physiol. Plant 2004, 122, 419–428. [Google Scholar] [CrossRef]
- Durães, F.O.M.; Gama, E.E.G.; Magalhães, P.C.; Marriel, I.E.; Casela, C.R.; Oliveira, A.C.; Junior, A.L.; Shanahan, J.F. The usefulness of chlorophyll fluorescence in screening for disease resistance, water stress tolerance, aluminium toxicity tolerance, and use efficiency in maize. In Proceedings of the Seventh Eastern and Southern Africa Regional Maize Conference. KARI|CIMMYT, Nairobi, Kenya; 2002; pp. 356–360. [Google Scholar]
- Tung, J.; Goodwin, P.H.; Hsiang, T. Chlorophyll fluorescence for quantification of fungal foliar infection and assessment of the effectiveness of an induced systemic resistance activator. Eur. J. Plant Pathol. 2013, 136, 301–315. [Google Scholar] [CrossRef]
- Santos, E.D.O.; Alves, N., Jr.; Dias, G.M.; Mazotto, A.M.; Vermelho, A.; Vora, G.J.; Wilson, B.; Beltran, V.H.; Bourne, D.G.; Le Roux, F.; et al. Genomic and proteomic analyses of the coral pathogen Vibrio coralliilyticus reveal a diverse virulence repertoire. ISME J. 2011, 5, 1471–1483. [Google Scholar] [CrossRef]
- Yang, J.C.; Madupu, R.; Durkin, A.S.; Ekborg, N.A.; Pedamallu, C.S.; Hostetler, J.B.; Radune, D.; Toms, B.S.; Henrissat, B.; Coutinho, P.M.; et al. The complete genome of Teredinibacter turnerae T7901: An intracellular endosymbiont of marine wood-boring bivalves (shipworms). PLoS ONE 2009, 4, e6085. [Google Scholar] [CrossRef]
- Barradas, C.; Phillips, A.; Correia, A.; Diogo, E.; Bragança, H.; Alves, A. Diversity and potential impact of Botryosphaeriaceae species associated with Eucalyptus globulus plantations in Portugal. Eur. J. Plant Pathol. 2006, 146, 245–257. [Google Scholar] [CrossRef]
- Alves, A.; Correia, A.; Luque, J.; Phillips, A. Botryosphaeria corticola, sp nov on Quercus species, with notes and description of Botryosphaeria stevensii and its anamorph, Diplodia mutila. Mycologia 2004, 96, 598–613. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Scott, J.W. Tomato. In Genetic Resources, Chromosome Engineering, and Crop Improvement: Vegetable Crops; Singh, R.J., Ed.; CRC Press: Boca Raton, FL, USA, 2019; Volume 3, pp. 59–113. [Google Scholar]
- Serôdio, J.; Ezequiel, J.; Frommlet, J.; Laviale, M.; Lavaud, J. A method for the rapid generation of nonsequential light-response curves of chlorophyll fluorescence. Plant Physiol. 2013, 163, 1089–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serôdio, J.; Schmidt, W.; Frankenbach, S. A chlorophyll fluorescence-based method for the integrated characterization of the photophysiological response to light stress. J. Exp. Bot. 2017, 68, 1123–1135. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Cruz, A.; Areias, D.; Duarte, A.; Correia, A.; Suzuki, S.; Mendo, S. Aeromonas molluscorum Av27 is a potential tributyltin (TBT) bioremediator: Phenotypic and genotypic characterization indicates its safe application. Antonie Leeuwenhoek 2013, 104, 385–396. [Google Scholar] [CrossRef]
- Duarte, A.S.; Cavaleiro, E.; Pereira, C.; Merino, S.; Esteves, A.C.; Duarte, E.P.; Tomas, J.M.; Correia, A.C. Aeromonas piscicola AH-3 expresses an extracellular collagenase with cytotoxic properties. Lett. Appl. Microbiol. 2015, 60, 288–297. [Google Scholar] [CrossRef]
- Ammerman, N.C.; Beier-Sexton, M.; Azad, A.F. Growth and maintenance of Vero cell lines. Curr. Protoc. Microbiol. 2008, 11, A.4E.1–A.4E.7. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nature Meth. 2011, 8, 785–786. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazar Pour, F.; Cobos, R.; Rubio Coque, J.J.; Serôdio, J.; Alves, A.; Félix, C.; Ferreira, V.; Esteves, A.C.; Duarte, A.S. Toxicity of Recombinant Necrosis and Ethylene-Inducing Proteins (NLPs) from Neofusicoccum parvum. Toxins 2020, 12, 235. https://doi.org/10.3390/toxins12040235
Nazar Pour F, Cobos R, Rubio Coque JJ, Serôdio J, Alves A, Félix C, Ferreira V, Esteves AC, Duarte AS. Toxicity of Recombinant Necrosis and Ethylene-Inducing Proteins (NLPs) from Neofusicoccum parvum. Toxins. 2020; 12(4):235. https://doi.org/10.3390/toxins12040235
Chicago/Turabian StyleNazar Pour, Forough, Rebeca Cobos, Juan José Rubio Coque, João Serôdio, Artur Alves, Carina Félix, Vanessa Ferreira, Ana Cristina Esteves, and Ana Sofia Duarte. 2020. "Toxicity of Recombinant Necrosis and Ethylene-Inducing Proteins (NLPs) from Neofusicoccum parvum" Toxins 12, no. 4: 235. https://doi.org/10.3390/toxins12040235
APA StyleNazar Pour, F., Cobos, R., Rubio Coque, J. J., Serôdio, J., Alves, A., Félix, C., Ferreira, V., Esteves, A. C., & Duarte, A. S. (2020). Toxicity of Recombinant Necrosis and Ethylene-Inducing Proteins (NLPs) from Neofusicoccum parvum. Toxins, 12(4), 235. https://doi.org/10.3390/toxins12040235