The Importance of Allelopathic Picocyanobacterium Synechococcus sp. on the Abundance, Biomass Formation, and Structure of Phytoplankton Assemblages in Three Freshwater Lakes

 , and
, and
Abstract
:1. Introduction
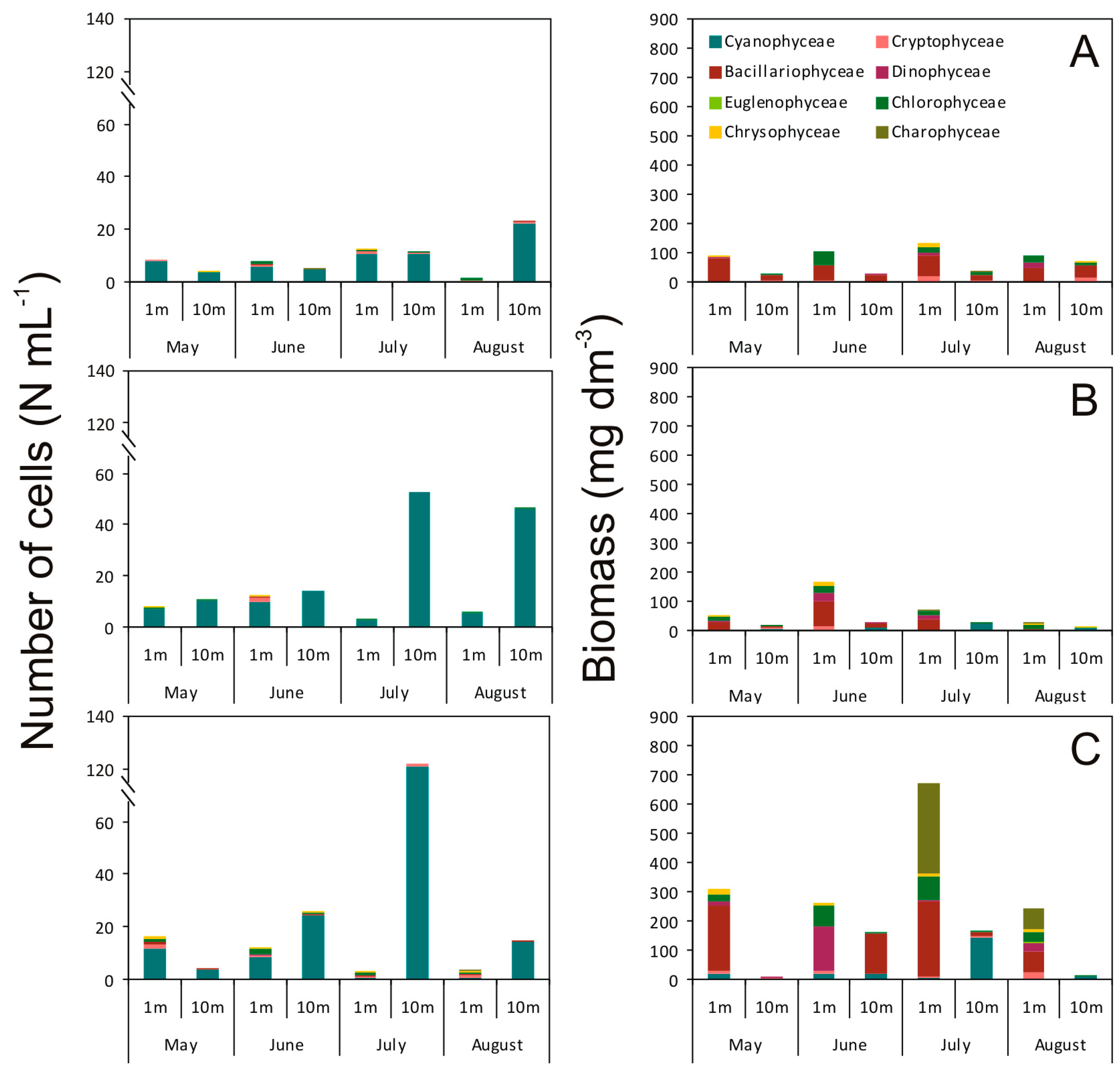
2. Results
2.1. Abundance, Biomass, and Structure of the Phytoplankton Community
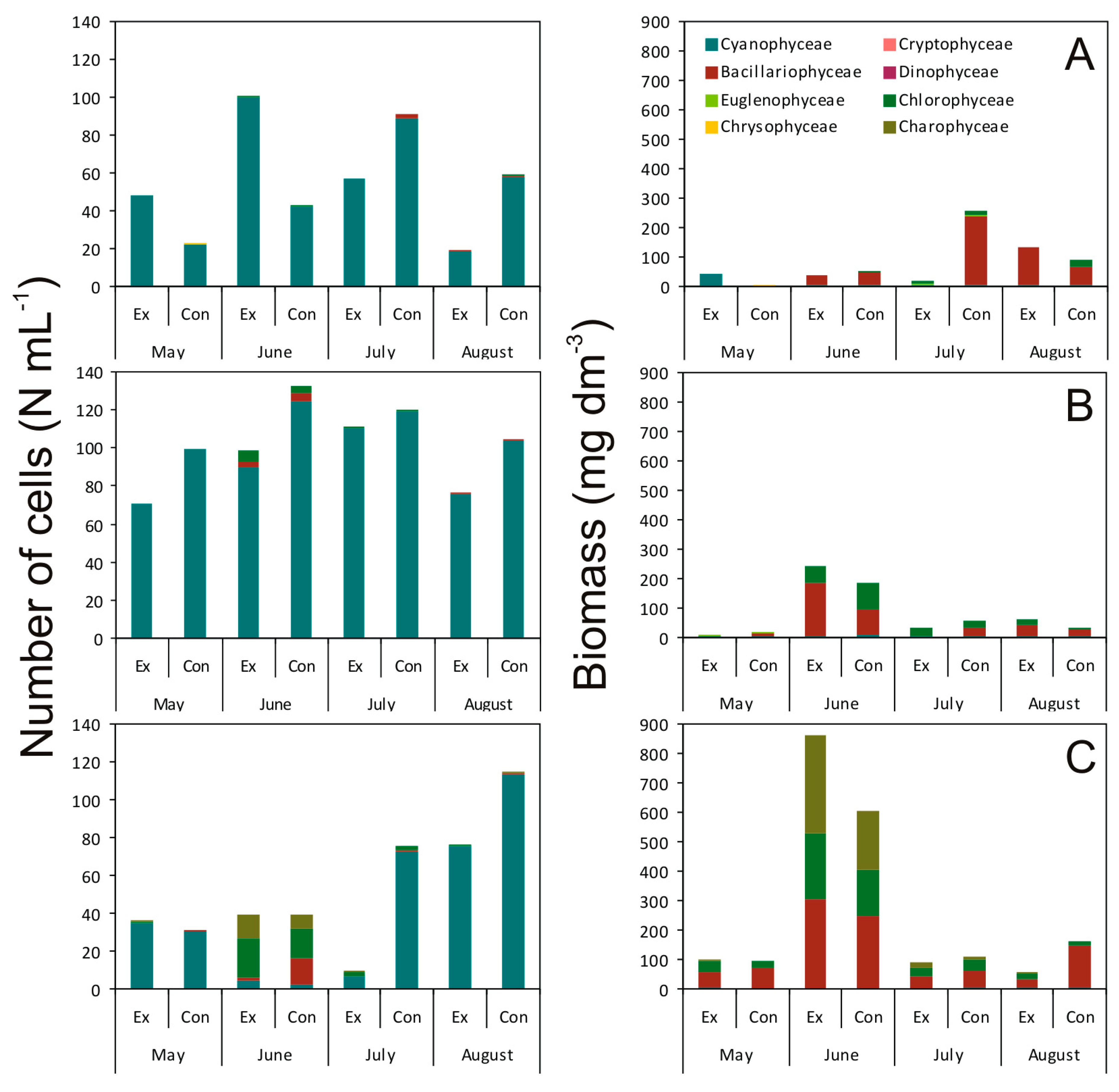
2.2. Effect of Picocyanobacterial Exudates on the Abundance, Biomass, and Structure of Phytoplankton Assemblages
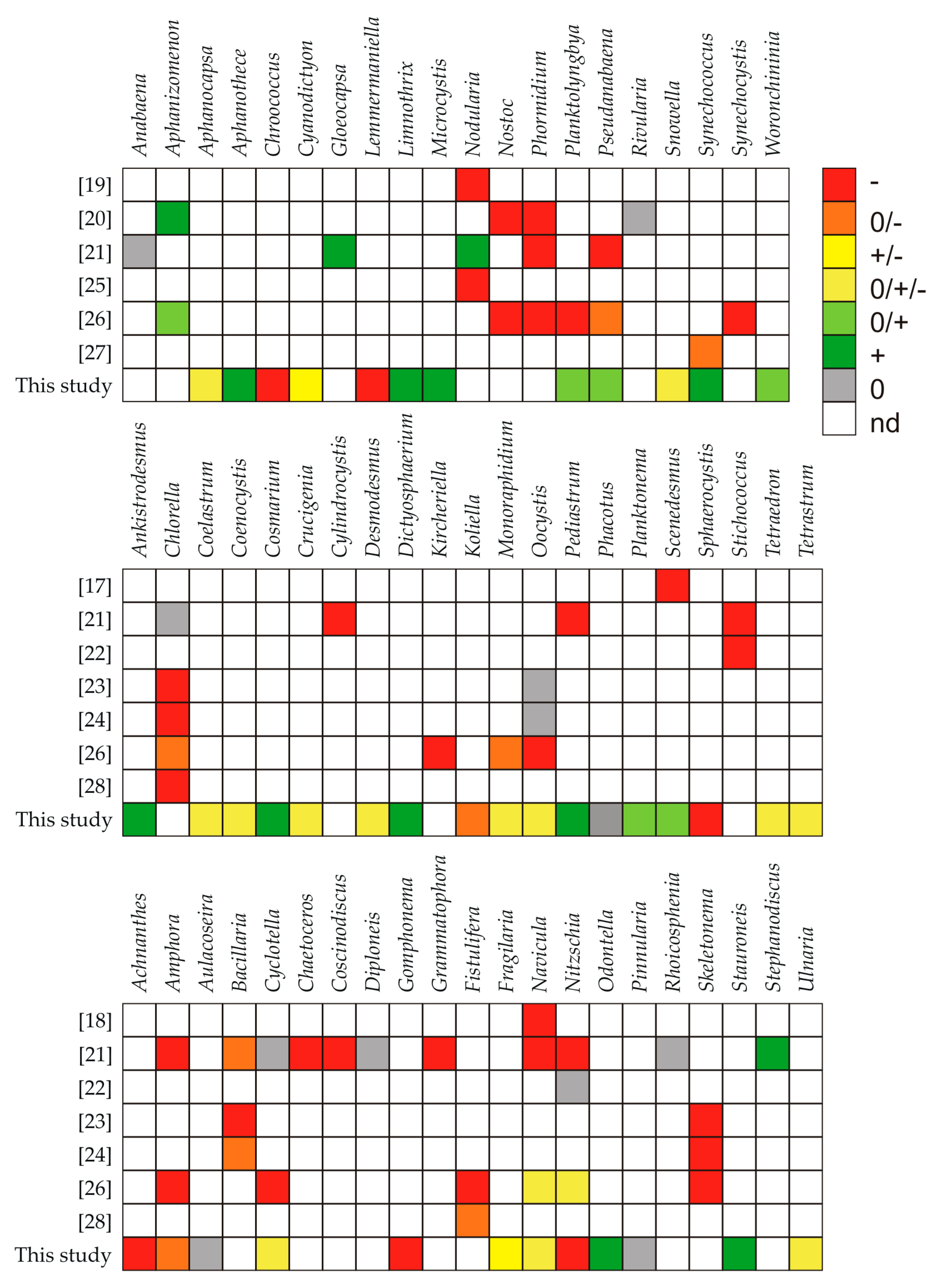
3. Discussion
4. Materials and Methods
4.1. Study Sites
4.2. Determination of Abundance, Biomass, and Structure of the Phytoplankton Community
4.3. Determination of the Allelopathic Activity of Synechococcus Exudates
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rumin, J.; Nicolau, E.; Junior, R.G.D.O.; Fuentes-Grünewald, C.; Flynn, K.J.; Picot, L. A bibliometric analysis of microalgae research in the world, Europe, and the European Atlantic area. Mar. Drugs 2020, 18, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burchard, L. Key to Identification of Phytoplankton Species in Lakes and Rivers. Guide for Laboratory Classes and Field Research; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2014; pp. 1–179. [Google Scholar]
- Naselli-Flores, L.; Padisak, J.; Albay, M. Shape and size in phytoplankton ecology: Do they matter? Hydrobiologia 2000, 578, 157–161. [Google Scholar] [CrossRef]
- Maberly, S.C.; King, L.; Dent, R.R.; Jones, R.I.; Gibson, C.E. Nutrient limitation of phytoplankton and periphyton growth in upland lakes. Freshw. Biol. 2002, 47, 2136–2152. [Google Scholar] [CrossRef]
- SOlitude Lake Management. Phytoplankton: Get to Know Your Pond Algae. 2017. Available online: https://www.solitudelakemanagement.com/blog/author/solitude-lake-management (accessed on 27 February 2020).
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [Green Version]
- Schindler, D.W. The dilemma of controlling cultural eutrophication of lakes. Proc. R. Soc. 2012, 279, 4322–4333. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W. Mitigating toxic planktonic cyanobacterial blooms in aquatic ecosystems facing increasing anthropogenic and climatic pressures. Toxins 2018, 10, 76. [Google Scholar] [CrossRef] [Green Version]
- They, N.H.; Marques, D.M.; Crossetti, L.O.; Becker, V.; Canterle, E.; Ribeiro Rodrigues, L.; Cardoso, L.; Fragoso, C.R. Júnior. Phytoplankton ecological interactions in freshwater ecosystems—Integrating relationships in subtropical shallow lakes. In Phytoplankton: Biology, Classification and Environmental Impacts, 1st ed.; Sebastiá, M.T., Ed.; NOVA Publishers: New York, NY, USA, 2014; pp. 73–130. [Google Scholar]
- Leão, P.N.; Vasconcelos, M.T.S.D.; Vasconcelos, V.M. Allelopathy in freshwater cyanobacteria. Crit. Rev. Microbiol. 2009, 35, 271–282. [Google Scholar] [CrossRef]
- Brutemark, A.; Vandelannoote, A.; Engström-Öst, J.; Suikkanen, S. A less saline Baltic Sea promotes cyanobacterial growth, hampers intracellular microcystin production, and leads to strain-specific differences in allelopathy. PLoS ONE 2015, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Żak, A.; Kosakowska, A. The influence of extracellular compounds produced by selected Baltic cyanobacteria, diatoms and dinoflagellates on growth of green algae Chlorella vulgaris. Estuar. Coast. Shelf Sci. 2015, 167, 113–118. [Google Scholar] [CrossRef]
- Pichierri, S.; Accoroni, S.; Pezzolesi, L.; Guerrini, F.; Romagnoli, T.; Pistocchi, R.; Totti, C. Allelopathic effects of diatom filtrates on the toxic benthic dinoflagellate Ostreopsis cf. ovata. Mar. Environ. Res. 2017, 131, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Van Meerssche, E.; Pinckney, J.L. The influence of salinity in the domoic acid effect on estuarine phytoplankton communities. Harmful Algae 2017, 69, 65–74. [Google Scholar] [CrossRef]
- Poulin, R.X.; Hogan, S.; Poulson-Ellestad, K.L.; Brown, E.; Fernández, F.M.; Kubanek, J. Karenia brevis allelopathy compromises the lipidome, membrane integrity, and photosynthesis of competitors. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Poulin, R.X.; Poulson-Ellestad, K.L.; Roy, J.S.; Kubanek, J. Variable allelopathy among phytoplankton reflected in red tide metabolome. Harmful Algae 2018, 71, 50–56. [Google Scholar] [CrossRef]
- Kovács, A.W.; Tóth, V.R.; Pálffy, K. The effects of interspecific interactions between bloom forming cyanobacteria and Scenedesmus quadricauda (chlorophyta) on their photophysiology. Acta Biol. Hung. 2018, 69, 210–223. [Google Scholar] [CrossRef] [Green Version]
- Śliwińska-Wilczewska, S.; Pniewski, F.; Latała, A. Allelopathic activity of the picocyanobacterium Synechococcus sp. under varied light, temperature and salinity conditions. Int. Rev. Hydrobiol. 2016, 101, 69–77. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Pniewski, F.; Latała, A. Allelopathic interactions between Synechococcus sp. and Nodularia spumigena under different light conditions. Allelopathy J. 2016, 37, 241–252. [Google Scholar]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Barreiro Felpeto, A.; Vasconcelos, V.; Latała, A. Allelopathic activity of the picocyanobacterium Synechococcus sp. on filamentous cyanobacteria. J. Exp. Mar. Biol. Ecol. 2017, 496, 16–21. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Tuszer, J.; Dobosz, K.; Kalusa, D.; Latała, A. First record of allelopathic activity of the picocyanobacterium Synechococcus sp. on a natural plankton community. Ecohydrol. Hydrobiol. 2017, 17, 227–234. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Barreiro Felpeto, A.; Maculewicz, J.; Sobczyk, A.; Vasconcelos, V.; Latała, A. Allelopathic activity of the picocyanobacterium Synechococcus sp. on unicellular eukaryote planktonic microalgae. Mar. Freshwater Res. 2018, 69, 1472–1479. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Latała, A. Allelopathic activity of the bloom-forming picocyanobacterium Synechococcus sp. on the coexisting microalgae: the role of eutrophication. Int. Rev. Hydrobiol. 2018, 103, 37–47. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Barreiro Felpeto, A.; Możdżeń, K.; Vasconcelos, V.; Latała, A. Physiological Effects on Coexisting Microalgae of the Allelochemicals Produced by the Bloom-Forming Cyanobacteria Synechococcus sp. and Nodularia spumigena. Toxins 2019, 11, 712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreiro Felpeto, A.; Śliwińska-Wilczewska, S.; Złoch, I.; Vasconcelos, V. Light-dependent cytolysis in the allelopathic interaction between picoplanktic and filamentous cyanobacteria. J. Plankton Res. 2018, 40, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Konarzewska, Z.; Śliwińska-Wilczewska, S.; Barreiro Felpeto, A.; Vasconcelos, V.; Latała, A. Assessment of the Allelochemical Activity and Biochemical Profile of Different Phenotypes of Picocyanobacteria from the Genus Synechococcus. Mar. Drugs 2020, 18, 179. [Google Scholar] [CrossRef] [Green Version]
- Paz-Yepes, J.; Brahamsha, B.; Palenik, B. Role of a Microcin-C-like biosynthetic gene cluster in allelopathic interactions in marine Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 12030–12035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreiro Felpeto, A.; Śliwińska-Wilczewska, S.; Klin, M.; Konarzewska, Z.; Vasconcelos, V. Temperature-dependent impacts of allelopathy on growth, pigment, and lipid content between a subpolar strain of Synechocystis sp. CCBA MA-01 and coexisting microalgae. Hydrobiologia 2019, 835, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.F.; Ramos, M.F.; Herfindal, L.; Sousa, J.A.; Skærven, K.; Vasconcelos, V.M. Antimicrobial and cytotoxic assessment of marine cyanobacteria—Synechocystis and Synechococcus. Mar. Drugs 2008, 6, 1–11. [Google Scholar] [CrossRef]
- Frazão, B.; Martins, R.; Vasconcelos, V. Are known cyanotoxins involved in the toxicity of picoplanktonic and filamentous north atlantic marine cyanobacteria? Mar. Drugs 2010, 8, 1908–1919. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.S.; Costa, M.; Ramos, V.; Leão, P.N.; Barreiro, A.; Vasconcelos, V.; Martins, R. Picocyanobacteria from a clade of marine cyanobium revealed bioactive potential against microalgae, bacteria, and marine invertebrates. Toxicol. Environ. Health Part A 2015, 78, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, T.J.; Paz-Yepes, J.; Morrison, R.A.; Palenik, B.; Tresguerres, M. Exposure to bloom-like concentrations of two marine Synechococcus cyanobacteria (strains CC9311 and CC9902) differentially alters fish behaviour. Conserv. Physiol. 2014, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Worden, A.Z.; Wilken, S. A plankton bloom shifts as the ocean warms. Science 2016, 354, 287–288. [Google Scholar] [CrossRef]
- Jasser, I.; Callieri, C. Picocyanobacteria. In Handbook on Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 19–27. [Google Scholar]
- Śliwińska-Wilczewska, S.; Cieszyńska, A.; Maculewicz, J.; Latała, A. Ecophysiological characteristics of red, green, and brown strains of the Baltic picocyanobacterium Synechococcus sp.—A laboratory study. Biogeosciences 2018, 15, 6257–6276. [Google Scholar] [CrossRef] [Green Version]
- Stockner, J.; Callieri, C.; Cronberg, G. Picoplankton and other non-bloom forming cyanobacteria in lakes. In The Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, UK, 2000; pp. 195–238. [Google Scholar]
- Phlips, E.J.; Badylak, S.; Lynch, T.C. Blooms of the picoplanktonic cyanobacterium Synechococcus in Florida Bay, a subtropical inner-shelf lagoon. Limnol. Oceanogr. 1999, 14, 1166–1175. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, P.Y.; Sorokin, Y.I.; Boscolo, R.; Giovanardi, O. Bloom of picocyanobacteria in the Venice lagoon during summer-autumn 2001: Ecological sequences. Hydrobiologia 2004, 523, 71–85. [Google Scholar] [CrossRef]
- Sorokin, Y.I.; Zakuskina, O.Y. Features of the Comacchio ecosystem transformed during persistent bloom of picocyanobacteria. J. Oceanogr. 2010, 66, 373–387. [Google Scholar] [CrossRef]
- Li, J.; Chen, Z.; Jing, Z.; Zhou, L.; Li, G.; Ke, Z.; Tan, Y. Synechococcus bloom in the Pearl River Estuary and adjacent coastal area—With special focus on flooding during wet seasons. Sci. Total Environ. 2019, 692, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Ciscar, J.C.; Feyen, L.; Soria, A. Climate impacts in Europe; The JRC PESETA II Project, JRC Scientific and Politicy Reports, EUR 26586EN; EUR—Scientific and Technical Research, Joint Research Centre of the European Commission: Seville, Spain, 2014; p. 155. [Google Scholar]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincon, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.; Lomas, M.W.; Veneziano, D. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [Green Version]
- Dutkiewicz, S.; Morris, J.J.; Follows, M.J.; Scott, J.; Levitan, O.; Dyhrman, S.T.; Berman-Frank, I. Impact of ocean acidification on the structure of future phytoplankton communities. Nat. Clim. Chang. 2015, 5, 1002–1006. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Barreiro Felpeto, A.; Latała, A. Allelopathic and bloom-forming picocyanobacteria in a changing world. Toxins 2018, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Suikkanen, S.; Fistarol, G.O.; Granéli, E. Allelopathic effects of the Baltic cyanobacteria Nodularia spumigena, Aphanizomenon flos-aquae and Anabaena lemmermannii on algal monocultures. J. Exp. Mar. Biol. Ecol. 2004, 308, 85–101. [Google Scholar] [CrossRef]
- Gleason, F.K.; Baxa, C.A. Activity of the natural algicide, cyanobacterin, on eukaryotic microorganisms. FEMS Microbiol. Lett. 1986, 33, 85–88. [Google Scholar] [CrossRef]
- Gleason, F.K. The natural herbicide, cyanobacterin, specifically disrupts thylakoid membrane structure in Euglena gracilis strains Z. FEMS Microbiol. Lett. 1990, 68, 77–82. [Google Scholar] [CrossRef]
- Bagchi, S.N.; Palod, A.; Chauhan, V.S. Algicidal properties of a bloom-forming blue-green-alga, Oscillatoria sp. J. Basic Microbiol. 1990, 30, 21–29. [Google Scholar] [CrossRef]
- Bagchi, S.N.; Chauhan, V.S.; Marwaii, J.B. Effect of an antibiotic from Oscillatoria late-virens on growth, photosynthesis, and toxicity of Microcystis aeruginosa. Curr. Microbiol. 1993, 26, 223–228. [Google Scholar] [CrossRef]
- Issa, A.A. Antibiotic production by the cyanobacteria Oscillatoria angustissima and Calothrix parietina. Environ. Toxicol. Pharmacol. 1999, 8, 33–37. [Google Scholar] [CrossRef]
- Schagerl, M.; Unterrieder, I.; Angeler, D.G. Allelopathy among cyanoprokaryota and other algae originating from Lake Neusiedlersee (Austria). Int. Rev. Hydrobiol. 2002, 87, 365–374. [Google Scholar] [CrossRef]
- Valdor, R.; Aboal, M. Effects of living cyanobacteria, cyanobacterial extracts and pure microcystins on growth and ultrastructure of microalgae and bacteria. Toxicon 2007, 49, 769–779. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B.; Kokociński, M.; Jurczak, T.; Lipski, D.; Wiktorowicz, K. Interspecific allelopathy in cyanobacteria: Cylindrospermopsin and Cylindrospermopsis raciborskii effect on the growth and metabolism of Microcystis aeruginosa. Harmful Algae 2014, 35, 1–8. [Google Scholar] [CrossRef]
- Swift, S.; Bainton, N.J.; Winson, M.K. Gram-negative bacterial communication by N-acyl homoserine lactones: a universal language? Trends Microbiol. 1994, 2, 193–198. [Google Scholar] [CrossRef]
- Pushparaj, B.; Pelosi, E.; Jüttner, F. Toxicological analysis of the marine cyanobacterium Nodularia harveyana. J. Appl. Phycol. 1999, 10, 527–530. [Google Scholar] [CrossRef]
- Gross, E.M. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Suikkanen, S.; Fistarol, G.O.; Granéli, E. Effects of cyanobacterial allelochemicals on a natural plankton community. Mar. Ecol. Prog. Ser. 2005, 287, 1–9. [Google Scholar] [CrossRef]
- Żak, A.; Musiewicz, K.; Kosakowska, A. Allelopathic activity of the Baltic cyanobacteria against microalgae. Estuar. Coast. Shelf Sci. 2012, 112, 4–10. [Google Scholar] [CrossRef]
- Wang, L.; Zi, J.; Xu, R.; Hilt, S.; Hou, X.; Chang, X. Allelopathic effects of Microcystis aeruginosa on green algae and a diatom: Evidence from exudates addition and co-culturing. Harmful Algae 2017, 61, 56–62. [Google Scholar] [CrossRef]
- Ma, Z.L.; Fang, T.X.; Thring, R.W.; Li, Y.B.; Yu, H.G.; Zhou, Q.; Zhao, M. Toxic and non-toxic strains of Microcystis aeruginosa induce temperature dependent allelopathy toward growth and photosynthesis of Chlorella vulgaris. Harmful Algae 2015, 48, 21–29. [Google Scholar] [CrossRef]
- Schlegel, I.; Doan, N.T.; de Chazal, N.; Smith, G.D. Antibiotic activity of new cyanobacterial isolates from Australia and Asia against green algae and cyanobacteria. J. Appl. Phycol. 1999, 10, 471–479. [Google Scholar] [CrossRef]
- Song, H.; Lavoie, M.; Fan, X.; Tan, H.; Liu, G.; Xu, P.; Fu, Z.; Paerl, H.W.; Qian, H. Allelopathic interactions of linoleic acid and nitric oxide increase the competitive ability of Microcystis aeruginosa. ISME J. 2017, 11, 1865–1876. [Google Scholar] [CrossRef] [Green Version]
- Lam, C.W.Y.; Silvester, W.B. Growth interactions among blue-green (Anabaena oscillarioides, Microcystis aeruginosa) and green (Chlorella sp.) algae. Hydrobiologia 1979, 63, 135–143. [Google Scholar] [CrossRef]
- Gantar, M.; Berry, J.P.; Thomas, S.; Wang, M.; Perez, R.; Rein, K.S. Allelopathic activity among cyanobacteria and microalgae isolated from Florida freshwater habitats. FEMS Microbiol. Ecol. 2008, 64, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Keating, K.I. Allelopathic Influence on Blue-Green Bloom Sequence in a Eutrophic Lake. Science 1977, 196, 885–887. [Google Scholar] [CrossRef]
- Keating, K.I. Blue-green algal inhibition of diatom growth: Transition from mesotrophic to eutrophic community structure. Science 1978, 199, 971–973. [Google Scholar] [CrossRef]
- Lafforgue, M.; Szeligiewicz, W.; Devaux, J.; Poulin, M. Selective mechanisms controlling algal succession in Aydat Lake. Water Sci. Technol. 1995, 32, 117–127. [Google Scholar] [CrossRef]
- Choiński, A. Limnologia Fizyczna Polski; Wydawnictwo Naukowe UAM: Poznań, Poland, 2007; p. 548. (In Polish) [Google Scholar]
- Szumański, A. Objaśnienia do Szczegółowej Mapy Geologicznej Polski, Arkusz Giżycko (104); Państwowy Instytut Geologiczny: Warszawa, Poland, 2000; p. 33. (In Polish) [Google Scholar]
- Carlson, R.E. A Trophic State Index for Lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Jańczak, J. Atlas jezior Polski. Tom 3, Jeziora Pojezierza Mazurskiego i Polski południowej (The Atlas of Polish Lakes. Volume 3, Masurian Lakes and the Southern Part of Poland); Bogucki Wydaw: Nauk, Poznań, 1999; p. 240, (In Polish, English summary). [Google Scholar]
- SOP for Phytoplankton Analysis—Standard Operating Procedure for Phytoplankton Analysis; Grace Analytical Lab: Chicago, IL, USA, 1994; pp. 367–396.
- Wołowski, K. Taxonomic and Environmental Studies on Euglenophytes of the Kraków-Częstochowa Upland (Southern Poland); W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 1998; pp. 1–192. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2019; Available online: http://www.algaebase.org (accessed on 24 June 2019).
- Brierley, B.; Carvalho, L.; Davies, S.; Krokowski, J. Guidance on the quantitative analysis of phytoplankton in Freshwater Samples. Report to SNIFFER (Project WFD80), Edinburgh, UK, Phytoplankton Counting Guidance_v1_2007 12 05.doc. 2007, p. 1. Available online: http://nora.nerc.ac.uk/id/eprint/5654/1/Phytoplankton_Counting_Guidance_v1_2007_12_05.pdf (accessed on 27 February 2020).
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Lake | ||
|---|---|---|---|
| Łazduny | Rzęśniki | Żabińskie | |
| Surface (ha) | 10.6 | 12.0 | 41.6 |
| Volume (tys m2) | 964.6 | 1111.8 | 5072.8 |
| Maximum depth (m) | 22.4 | 26.0 | 44.4 |
| Average depth (m) | 9.1 | 7.8 | 12.2 |
| Maximum length (m) | 790 | 700 | 1073 |
| Maximum width (m) | 210 | 280 | 635 |
| Length of shoreline (m) | 1880 | 1700 | 2846 |
| Shoreline development index | 1.6 | 1.3 | 1.2 |
| Exposure index | 3.4 | 1.8 | 3.4 |
| Hydrological type | outflow lake | flow lake | flow lake |
| Total surface of catchment (km2) | 1.94 | 1.94 | 24.6 |
| Lake | Month | Depth [m] | T (°C) | pH | EC (µS·cm-1) | TP (mg P·dm-3) | TN (mg N·dm-3) |
|---|---|---|---|---|---|---|---|
| Łazduny | May | 1 | 12.9 | 8.5 | 383 | 0.08 | 0.04 |
| 10 | 5.4 | 7.5 | 394 | 0.07 | 0.38 | ||
| June | 1 | 23.3 | 8.5 | 378 | 0.03 | 0.64 | |
| 10 | 5.3 | 7.4 | 397 | 0.06 | 1.41 | ||
| July | 1 | 23.4 | 8.6 | 375 | 0.17 | 0.79 | |
| 10 | 6.4 | 7.6 | 396 | 0.05 | 1.08 | ||
| August | 1 | 22.5 | 8.5 | 369 | 0.10 | 0.62 | |
| 10 | 6.4 | 7.7 | 398 | 0.26 | 0.67 | ||
| Rzęśniki | May | 1 | 13.2 | 8.3 | 392 | 0.03 | 0.41 |
| 10 | 4.7 | 7.5 | 406 | 0.09 | 1.05 | ||
| June | 1 | 23.2 | 8.3 | 384 | 0.01 | 0.78 | |
| 10 | 4.8 | 7.4 | 406 | 0.05 | 0.82 | ||
| July | 1 | 23.8 | 8.4 | 383 | 0.07 | 0.80 | |
| 10 | 5.1 | 7.5 | 406 | 0.29 | 1.21 | ||
| August | 1 | 22.8 | 8.4 | 372 | 0.08 | 0.58 | |
| 10 | 5.3 | 7.6 | 407 | 0.12 | 0.90 | ||
| Żabińskie | May | 1 | 13.0 | 8.8 | 370 | 0.06 | 2.07 |
| 10 | 6.0 | 7.6 | 432 | 0.07 | 1.60 | ||
| June | 1 | - | - | - | 0.09 | 1.24 | |
| 10 | - | - | - | 0.22 | 2.30 | ||
| July | 1 | 23.7 | 8.8 | 325 | 0.06 | 1.14 | |
| 10 | 6.1 | 7.6 | 436 | 0.18 | 2.39 | ||
| August | 1 | 22.7 | 8.7 | 340 | 0.16 | 0.59 | |
| 10 | 6.1 | 7.6 | 438 | 0.23 | 2.04 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bubak, I.; Śliwińska-Wilczewska, S.; Głowacka, P.; Szczerba, A.; Możdżeń, K. The Importance of Allelopathic Picocyanobacterium Synechococcus sp. on the Abundance, Biomass Formation, and Structure of Phytoplankton Assemblages in Three Freshwater Lakes. Toxins 2020, 12, 259. https://doi.org/10.3390/toxins12040259
Bubak I, Śliwińska-Wilczewska S, Głowacka P, Szczerba A, Możdżeń K. The Importance of Allelopathic Picocyanobacterium Synechococcus sp. on the Abundance, Biomass Formation, and Structure of Phytoplankton Assemblages in Three Freshwater Lakes. Toxins. 2020; 12(4):259. https://doi.org/10.3390/toxins12040259
Chicago/Turabian StyleBubak, Iwona, Sylwia Śliwińska-Wilczewska, Paulina Głowacka, Agnieszka Szczerba, and Katarzyna Możdżeń. 2020. "The Importance of Allelopathic Picocyanobacterium Synechococcus sp. on the Abundance, Biomass Formation, and Structure of Phytoplankton Assemblages in Three Freshwater Lakes" Toxins 12, no. 4: 259. https://doi.org/10.3390/toxins12040259
APA StyleBubak, I., Śliwińska-Wilczewska, S., Głowacka, P., Szczerba, A., & Możdżeń, K. (2020). The Importance of Allelopathic Picocyanobacterium Synechococcus sp. on the Abundance, Biomass Formation, and Structure of Phytoplankton Assemblages in Three Freshwater Lakes. Toxins, 12(4), 259. https://doi.org/10.3390/toxins12040259