Transformations of Selected Fusarium Toxins and Their Modified Forms During Malt Loaf Production

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
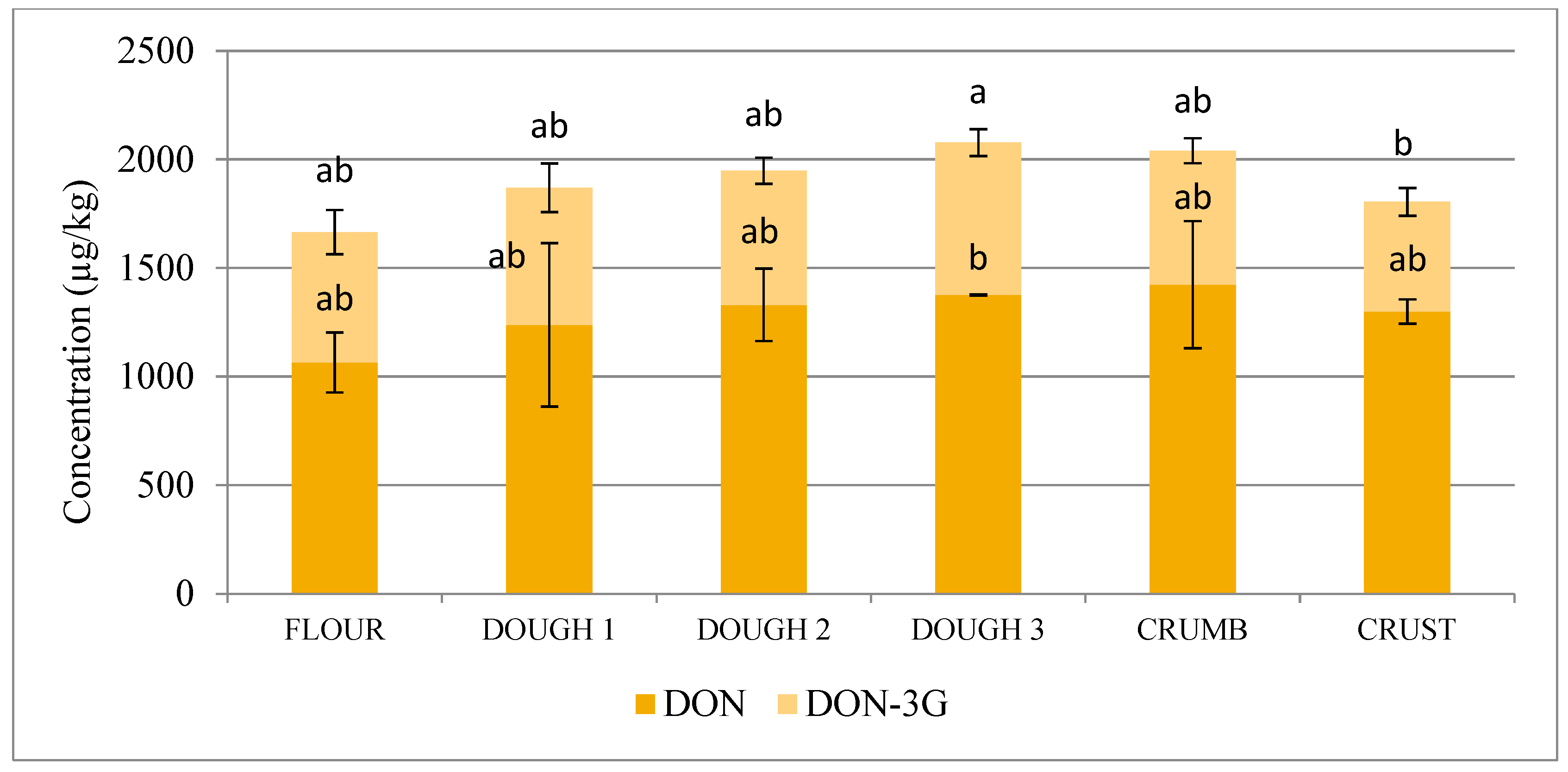
2.1. Trichothecenes and Their Modified Forms
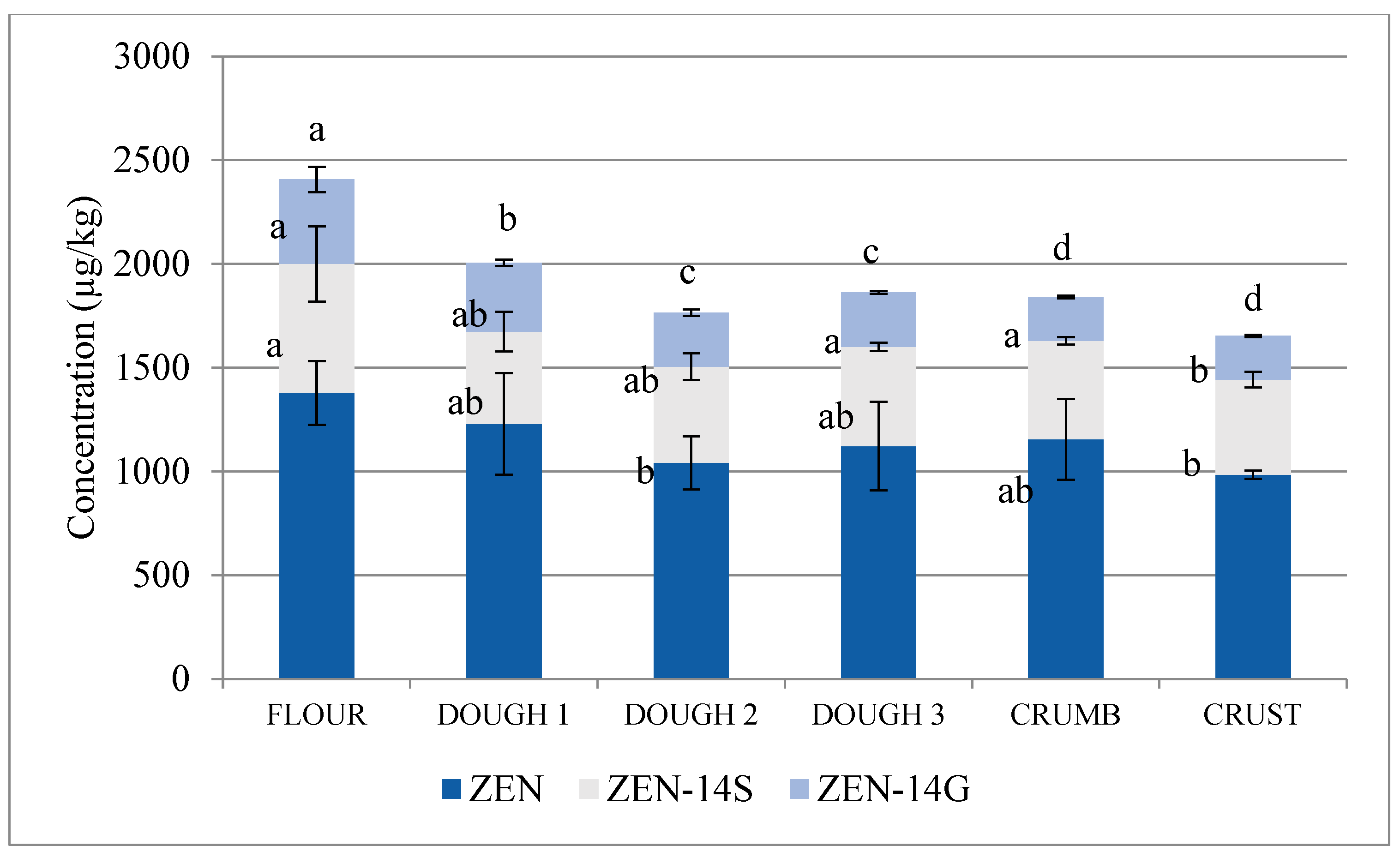
2.2. ZEN and Its Modified Forms
3. Conclusions
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Samples
4.3. Laboratory Baking
4.4. Sample Preparation
4.5. Liquid Chromatography-Mass Spectrometry (LC-TOF-HRMS)
4.6. Statistical Analysis
4.7. Method Validation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pleadin, J.; Vahčić, N.; Perši, N.; Ševelj, D.; Markov, K.; Frece, J. Fusarium mycotoxins’ occurrence in cereals harvested from Croatian fields. Food Control 2013, 32, 49–54. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Walder, F.; Schlaeppi, K.; Wittwer, R.; Held, A.Y.; Vogelgsang, S.; van der Heijden, M.G.A. Community profiling of Fusarium in combination with other plant-associated fungi in different crop species using SMRT sequencing. Front. Plant Sci. 2017, 8, 2019. [Google Scholar] [CrossRef] [PubMed]
- Stenglein, S.A. Fusarium poae: A pathogen that needs more attention. J. Plant Pathol. 2009, 91, 25–36. [Google Scholar]
- Nganje, W.E.; Bangsund, D.A.; Leistritz, F.L.; Wilson, W.W.; Tiapo, N.M. Regional economic impacts of Fusarium head blight in wheat and barley. Appl. Econ. Perspect. Policy 2004, 26, 332–347. [Google Scholar] [CrossRef]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Alshannaq, A.; Yu, J.-H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [Green Version]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [Green Version]
- International Agency for Research on Cancer. Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins; Monograph on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 1993. [Google Scholar]
- Pestka, J. Toxicological mechanisms and potential health effects of deoxynivalenol and nivalenol. World Mycotoxin J. 2010, 3, 323–347. [Google Scholar] [CrossRef]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef]
- Vidal, A.; Cano-Sancho, G.; Marín, S.; Ramos, A.J.; Sanchis, V. Multidetection of urinary ochratoxin A, deoxynivalenol and its metabolites: Pilot time-course study and risk assessment in Catalonia, Spain. World Mycotoxin J. 2016, 9, 597–612. [Google Scholar] [CrossRef]
- Heyndrickx, E.; Sioen, I.; Huybrechts, B.; Callebaut, A.; De Henauw, S.; De Saeger, S. Human biomonitoring of multiple mycotoxins in the Belgian population: Results of the BIOMYCO study. Environ. Int. 2015, 84, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Murata, H.; Sultana, P.; Shimada, N.; Yoshioka, M. Structure-activity relationships among zearalenone and its derivatives based on bovine neutrophil chemiluminescence. Vet. Hum. Toxicol. 2003, 45, 18–20. [Google Scholar] [PubMed]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2019. [Google Scholar] [CrossRef]
- Bryła, M.; Waśkiewicz, A.; Ksieniewicz-Woźniak, E.; Szymczyk, K.; Jędrzejczak, R. Modified Fusarium mycotoxins in cereals and their products—Metabolism, occurrence, and toxicity: An updated review. Molecules 2018, 23, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freire, L.; Sant’Ana, A.S. Modified mycotoxins: An uptake review on their formation, detection, occurrence, and toxic effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef]
- Pena, R.J. Wheat for bread and other foods. In Bread Wheat: Improvement and Production; Curtis, B.C., Rajaram, S., Macpherson, H.G., Eds.; Food and Agriculture Organization: Rome, Italy, 2002. [Google Scholar]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.C. Understanding Food: Principles and Preparation, 5th ed.; Cengage Learning: Stamford, CT, USA, 2015. [Google Scholar]
- Aehle, W. Industrial enzymes. In Enzymes in Industry: Production and Applications, 3rd ed.; Aehle, W., Ed.; Wiley-VCH: Weinheim, Germany, 2007. [Google Scholar]
- Martorana, A.; Giuffrè, A.M.; Capocasale, M.; Zappia, C.; Sidari, R. Sourdoughs as a source of lactic acid bacteria and yeasts with technological characteristics useful for improved bakery products. Eur. Food Res. Technol. 2018, 244, 1873–1885. [Google Scholar] [CrossRef]
- Reale, A.; Di Renzo, T.; Boscaino, F.; Nazzaro, F.; Fratianni, F.; Aponte, M. Lactic acid bacteria biota and aroma profile of Italian traditional sourdoughs from the Irpinian area in Italy. Front. Microbiol. 2019, 10, 1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachariasova, M.; Vaclavikova, M.; Lacina, O.; Vaclavik, L.; Hajslova, J. Deoxynivalenol oligoglycosides: New “masked” Fusarium toxins occurring in malt, beer, and breadstuff. J. Agric. Food Chem. 2012, 60, 9280–9291. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Yoshinari, T.; Waśkiewicz, A.; Szymczyk, K. Contamination of wheat cultivated in various regions of Poland during 2017 and 2018 agricultural seasons with selected trichothecenes and their modified forms. Toxins 2019, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Vidal, A.; Marín, S.; Morales, H.; Ramos, A.J.; Sanchis, V. The fate of deoxynivalenol and ochratoxin A during the bread making process, effects of sourdough use and bran content. Food Chem. Toxicol. 2014, 68, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergamini, E.; Catellani, D.; Dall’asta, C.; Galaverna, G.; Dossena, A.; Marchelli, R.; Suman, M. Fate of fusarium mycotoxins in the cereal product supply chain: The deoxynivalenol (DON) case within industrial bread-making technology. Food Addit. Contam. Part A 2010, 27, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, B. Fates of deoxynivalenol and deoxynivalenol-3-glucoside during bread and noodle processing. Food Control 2015, 50, 754–757. [Google Scholar] [CrossRef]
- Neira, M.S.; Pacin, A.M.; Martínez, E.J.; Moltó, G.; Resnik, S.L. The effects of bakery processing on natural deoxynivalenol contamination. Int. J. Food Microbiol. 1997, 37, 21–25. [Google Scholar] [CrossRef]
- Simsek, S.; Burgess, K.; Whitney, K.L.; Gu, Y.; Qian, S.Y. Analysis of deoxynivalenol and deoxynivalenol-3-glucoside in wheat. Food Control 2012, 26, 287–292. [Google Scholar] [CrossRef]
- Generotti, S.; Cirlini, M.; Malachova, A.; Sulyok, M.; Berthiller, F.; Dall’Asta, C.; Suman, M. Deoxynivalenol & deoxynivalenol-3-glucoside mitigation through bakery production strategies: Effective experimental design within industrial rusk-making technology. Toxins 2015, 7, 2773–2790. [Google Scholar]
- Suman, M.; Manzitti, A.; Catellani, D. A design of experiments approach to studying deoxynivalenol and deoxynivalenol-3-glucoside evolution throughout industrial production of whole grain crackers exploiting LC–MS/MS techniques. World Mycotoxin J. 2012, 5, 241–249. [Google Scholar] [CrossRef]
- Vidal, A.; Morales, H.; Sanchis, V.; Ramos, A.J.; Marín, S. Stability of DON and OTA during the breadmaking process and determination of process and performance criteria. Food Control 2014, 40, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Vidal, A.; Ambrosio, A.; Sanchis, V.; Ramos, A.J.; Marín, S. Enzyme bread improvers affect the stability of deoxynivalenol and deoxynivalenol-3-glucoside during breadmaking. Food Chem. 2016, 208, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, Y.; Yoshizawa, T.; Morooka, N. Effect of food additives and heating on the decomposition of zearalenone in wheat flour. Food Hyg. Saf. Sci. (Shokuhin Eiseigaku Zasshi) 1981, 22, 293–298. [Google Scholar] [CrossRef]
- El-Desouky, T.A.; May Amer, M.; Naguib, K. Effects of pan bread making on zearalenone levels in artificial contaminated wheat flour. Hung. J. Ind. Chem. 2012, 40, 107–111. [Google Scholar]
- Numanoglu, E.; Yener, S.; Gökmen, V.; Uygun, U.; Koksel, H. Modelling thermal degradation of zearalenone in maize bread during baking. Food Addit. Contam. Part A 2013, 30, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Cano-Sancho, G.; Sanchis, V.; Ramos, A.J.; Marín, S. Effect of food processing on exposure assessment studies with mycotoxins. Food Addit. Contam. Part A 2013, 30, 867–875. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- European Commission. Commission Regulation (EC) No 178/2002 of 28 January 2002 laying down the general principles and requirements of food law, establishing the European Food Safety Authority and laying down procedures in matters of food safety. Off. J. Eur. Communities 2002, 31, 1–24. [Google Scholar]
- Popper, L. Flour treatment. In Future of Flour. A Compendium of Flour Improvement; Popper, L., Schäfer, W., Freund, W., Eds.; Ehrling Verlag: Clenze, Germany, 2006. [Google Scholar]
- Gomaa, A.M. Application of enzymes in brewing. J. Nutr. Food Sci. Forecast 2018, 1, 5. [Google Scholar]
- Kostelanska, M.; Dzuman, Z.; Malachova, A.; Capouchova, I.; Prokinova, E.; Skerikova, A.; Hajslova, J. Effects of milling and baking technologies on levels of deoxynivalenol and its masked form deoxynivalenol-3-glucoside. J. Agric. Food Chem. 2011, 59, 9303–9312. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Sanchis, V.; Ramos, A.J.; Marín, S. Effect of xylanase and α-amylase on DON and its conjugates during the breadmaking process. Food Res. Int. 2017, 101, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Corsetti, A.; Settanni, L. Lactobacilli in sourdough fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Young, J.C.; Fulcher, R.G.; Hayhoe, J.H.; Scott, P.M.; Dexter, J.E. Effect of milling and baking on deoxynivalenol (vomitoxin) content of eastern Canadian wheats. J. Agric. Food Chem. 1984, 32, 659–664. [Google Scholar] [CrossRef]
- Boyacioğlu, D.; Heitiarachchy, N.S.; D’Appolonia, B.D. Additives affect deoxynivalenol (vomi toxin) flour during breadbaking. J. Food Sci. 1993, 58, 416–418. [Google Scholar] [CrossRef]
- Generotti, S.; Cirlini, M.; Šarkanj, B.; Sulyok, M.; Berthiller, F.; Dall’Asta, C.; Suman, M. Formulation and processing factors affecting trichothecene mycotoxins within industrial biscuit-making. Food Chem. 2017, 229, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.B.; Font, G.; Mañes, J.; Ferrer, E. Development a mitigation strategy of enniatins in pasta under home-cooking conditions. LWT-Food Sci. Technol. 2016, 65, 1017–1024. [Google Scholar] [CrossRef]
- Lancova, K.; Hajslova, J.; Kostelanska, M.; Kohoutkova, J.; Nedelnik, J.; Moravcova, H.; Vanova, M. Fate of trichothecene mycotoxins during the processing: Milling and baking. Food Addit. Contam. Part A 2008, 25, 650–659. [Google Scholar] [CrossRef]
- Banu, I.; Dragoi, L.; Aprodu, I. From wheat to sourdough bread: A laboratory scale study on the fate of deoxynivalenol content. Qual. Assur. Saf. Crops Foods 2014, 6, 53–60. [Google Scholar] [CrossRef]
- Scott, P.M.; Kanhere, S.R.; Dexter, J.E.; Brennan, P.W.; Trenholm, H.L. Distribution of the trichothecene mycotoxin deoxynivalenol (vomitoxin) during the milling of naturally contaminated hard red spring wheat and its fate in baked products. Food Addit. Contam. 1984, 1, 313–323. [Google Scholar] [CrossRef]
- Sugita-Konishi, Y.; Park, B.J.; Kobayashi-Hattori, K.; Tanaka, T.; Chonan, T.; Yoshikawa, K.; Kumagai, S. Effect of cooking process on the deoxynivalenol content and its subsequent cytotoxicity in wheat products. Biosci. Biotechnol. Biochem. 2006, 70, 1764–1768. [Google Scholar] [CrossRef] [Green Version]
- Giménez, I.; Blesa, J.; Herrera, M.; Ariño, A. Effects of bread making and wheat germ addition on the natural deoxynivalenol content in bread. Toxins 2014, 6, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Samar, M.M.; Neira, M.S.; Resnik, S.L.; Pacin, A. Effect of fermentation on naturally occurring deoxynivalenol (DON) in Argentinean bread processing technology. Food Addit. Contam. 2001, 18, 1004–1010. [Google Scholar] [CrossRef]
- Ragab, W.S.M.; Drusch, S.; Kuhlmann, A.; Beyer, M. Characterizing effects of fermentation and baking on the deoxynivalenol content of rolls. J. Appl. Bot. Food Qual. 2005, 79, 197–201. [Google Scholar]
- Dropa, T.; Hajšlová, J.; Lancová, K.; Burešová, I. The effect of bread-making process on contents of key trichothecene mycotoxins: Deoxynivalenol, T-2, and HT-2 toxins. Czech J. Food Sci. 2014, 6, 570–577. [Google Scholar] [CrossRef] [Green Version]
- Valle-Algarra, F.M.; Mateo, E.M.; Medina, Á.; Mateo, F.; Gimeno-Adelantado, J.V.; Jiménez, M. Changes in ochratoxin A and type B trichothecenes contained in wheat flour during dough fermentation and bread-baking. Food Addit. Contam. Part A 2009, 26, 896–906. [Google Scholar] [CrossRef]
- De Angelis, E.; Monaci, L.; Pascale, M.; Visconti, A. Fate of deoxynivalenol, T-2 and HT-2 toxins and their glucoside conjugates from flour to bread: An investigation by high-performance liquid chromatography high-resolution mass spectrometry. Food Addit. Contam. Part A 2013, 30, 345–355. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, L.; Li, W.; Zhang, Y.; Li, J.; Hu, X.; Sun, L.; Du, W.; Wang, B. Conversion of deoxynivalenol-3-glucoside to deoxynivalenol during chinese steamed bread processing. Toxins 2020, 12, 225. [Google Scholar] [CrossRef] [Green Version]
- Stadler, D.; Lambertini, F.; Woelflingseder, L.; Schwartz-Zimmermann, H.; Marko, D.; Suman, M.; Berthiller, F.; Krska, R. The Influence of processing parameters on the mitigation of deoxynivalenol during industrial baking. Toxins 2019, 11, 317. [Google Scholar] [CrossRef] [Green Version]
- Heidari, S.; Milani, J.; Seyed, S.S.; Nazari, J. Effect of the bread making process on zearalenone levels. Food Addit. Contam. Part A 2014, 31, 2047–2054. [Google Scholar] [CrossRef]
- Bol, E.K.; Araujo, L.; Veras, F.F.; Welke, J.E. Estimated exposure to zearalenone, ochratoxin A and aflatoxin B1 through the consume of bakery products and pasta considering effects of food processing. Food Chem. Toxicol. 2016, 89, 85–91. [Google Scholar] [CrossRef]
- EFSA—European Food Safety Authority. Appropriateness to set a group health-based guidance value for ZEN and its modified forms. EFSA J. 2016, 14, 4425. [Google Scholar]
- EFSA—European Food Safety Authority. Risks to human and animal health related to the presence of DON and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, 4718. [Google Scholar]
- Yoshinari, T.; Sakuda, S.; Furihata, K.; Furusawa, H.; Ohnishi, T.; Sugita-Konishi, Y.; Ishizaki, N.; Terajima, J. Structural determination of a nivalenol glucoside and development of an analytical method for the simultaneous determination of nivalenol and deoxynivalenol, and their glucosides, in wheat. J. Agric. Food Chem. 2014, 62, 1174–1180. [Google Scholar] [CrossRef]
- Nathanail, A.V.; Syvähuoko, J.; Malachová, A.; Jestoi, M.; Varga, E.; Michlmayr, H.; Adam, G.; Sieviläinen, E.; Berthiller, F.; Peltonen, K. Simultaneous determination of major type A and B trichothecenes, zearalenone and certain modified metabolites in Finnish cereal grains with a novel liquid chromatography-tandem mass spectrometric method. Anal. Bioanal. Chem. 2015, 407, 4745–4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]




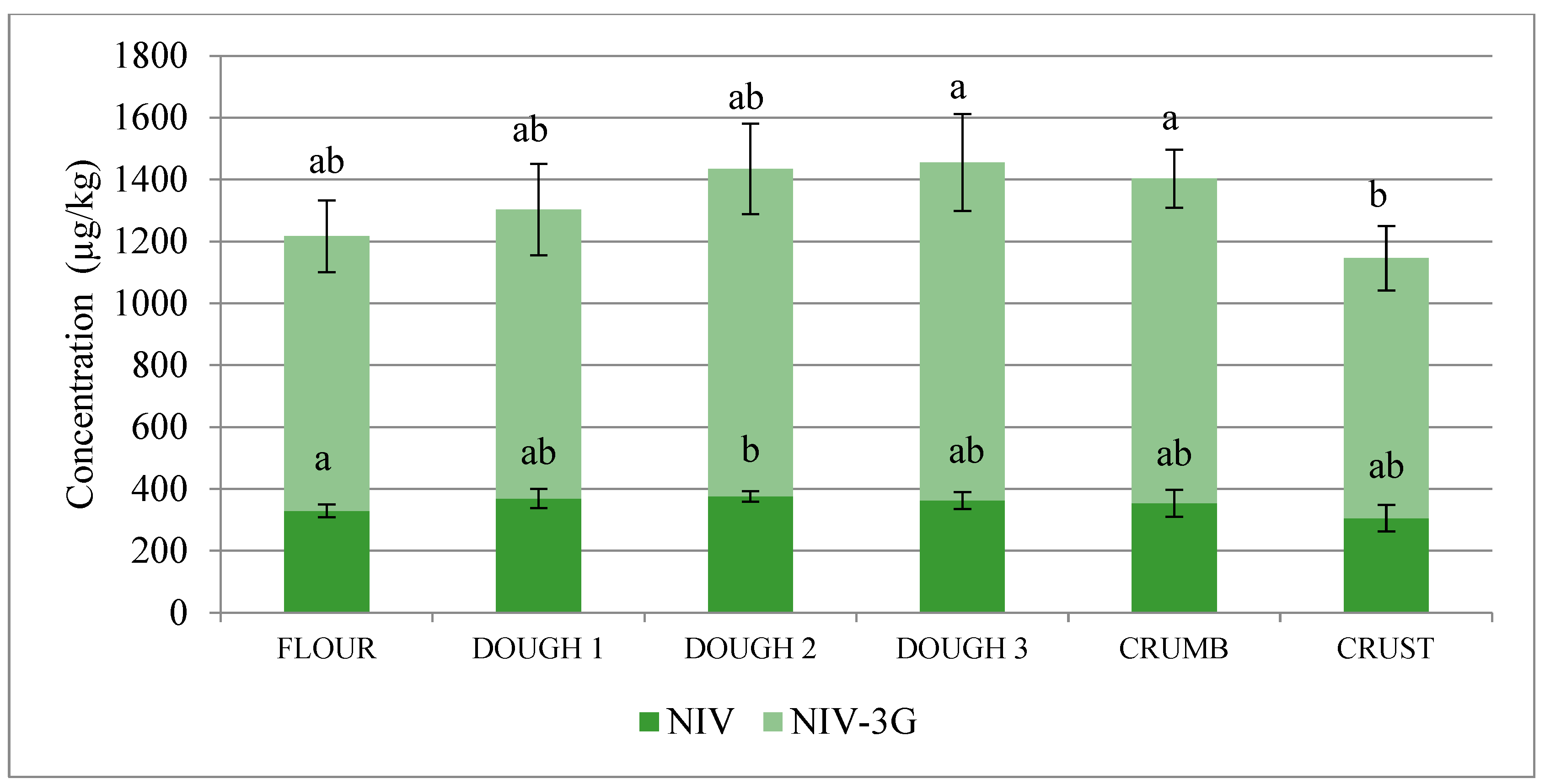{kind=link}
{kind=link}
{kind=link}
| Molar Ratio | Flourp H 5.76 ± 0.09 (n = 3) | Dough 1 pH 5.72 ± 0.10 (n = 3) | Dough 2 pH 5.59 ± 0.08 (n = 3) | Dough 3 pH 5.42 ± 0.12 (n = 3) | Crumb pH 5.37 ± 0.10 (n = 12) | Crust pH 5.27 ± 0.11 (n = 12) |
|---|---|---|---|---|---|---|
| NIV-3G/NIV | 1.77 | 1.66 | 1.85 | 1.98 | 1.95 | 1.80 |
| DON-3G/DON | 0.36 | 0.33 | 0.30 | 0.33 | 0.28 | 0.25 |
| ZEN-14S/ZEN | 0.36 | 0.29 | 0.36 | 0.35 | 0.33 | 0.37 |
| ZEN-14G/ZEN | 0.20 | 0.18 | 0.17 | 0.16 | 0.12 | 0.14 |
| Analyte | Kneading | Fermentation | Baking | Whole Process | |||
|---|---|---|---|---|---|---|---|
| Dough 1/Flour | Dough 2/Dough 1 | Dough 3/Dough 2 | Crumb/Dough 3 | Crust/Dough 3 | Crumb/Flour | Crust/Flour | |
| NIV | +12.1 | +1.6 | −3.5 | −2.5 | −15.7 | +7.3 | −7.3 |
| NIV-3G | +5.3 | +13.3 | +3.2 | −3.9 | −23.2 | +18.3 * | −5.4 |
| NIV+NIV-3G | +7.1 | +10.0 | +1.5 | −3.6 | −21.3 * | +15.3 | −5.9 |
| DON | +16.2 | +7.5 | +3.4 | +3.4 | −5.5 | +33.6 * | 22.1 * |
| DON-3G | +5.0 | −2.2 | +13.8 | −12.1 | −28.1 | +2.7 | −16.0 |
| DON+DON-3G | +12.2 | +4.2 | +6.7 | −1.8 | −13.1 | +22.4 * | +8.3 |
| ZEN | −10.7 | −15.3 | +7.7 | +2.9 | −12.3 | −16.2 | −28.6 * |
| ZEN-14S | −22.2 * | +0.4 | +1.6 | +1.4 | −27.3 | −19.5 * | −42.3 * |
| ZEN-14G | −18.4 | −21.4 | +0.8 | −19.4 | −20.2 | −47.9 | −48.4 * |
| ZEN+ZEN-14S+ZEN-14G | −15.0 | −12.6 | +5.0 | −0.6 | −17.3 | −22.4 * | −35.5 * |
| Analyte | Ion Mass (m/z) | Retention Time (min) | LOD (μg/L) | LOQ (μg/L) | Coefficient of Determination (R2) | |
|---|---|---|---|---|---|---|
| Flour | Bread | |||||
| NIV | 357.21 (M+FA-H)− | 2.38 | 38 | 125 | 0.9155 | 0.9889 |
| NIV-3G | 519.22 (M+FA-H)− | 2.45 | 15 | 50 | 0.9906 | 0.9858 |
| DON | 341.22 (M+FA-H)− | 4.08 | 75 | 250 | 0.9892 | 0.9988 |
| DON-3G | 503.23 (M+FA-H)− | 4.22 | 38 | 125 | 0.9969 | 0.9987 |
| ZEN | 317.10 (M-H)− | 11.65 | 10 | 34 | 0.9958 | 0.9910 |
| ZEN-14G | 525.11 (M+FA-H)− | 6.63 | 20 | 68 | 0.9981 | 0.9887 |
| ZEN-14S | 397.09 (M-H)− | 7.25 | 20 | 68 | 0.9970 | 0.9941 |
| Mycotoxin | Bread | Flour | ||||
|---|---|---|---|---|---|---|
| Fortification (μg/kg) | RSD (%) | R (%) | Fortification (μg/kg) | RSD (%) | R (%) | |
| NIV | 125 | 10.5 | 87.3 | 250 | 10.5 | 81.9 |
| 250 | 3.5 | 76 | 500 | 13.9 | 90.9 | |
| 500 | 5.3 | 78 | 1000 | 19.2 | 94.1 | |
| NIV-3G | 50 | 11.9 | 79.1 | 100 | 16.1 | 91.4 |
| 100 | 15.2 | 99 | 200 | 8.1 | 98.2 | |
| 200 | 6.9 | 77 | 400 | 11.3 | 100 | |
| DON | 1250 | 4.6 | 94.3 | 2500 | 4.8 | 99.9 |
| 2500 | 5.2 | 84 | 5000 | 4.6 | 98.9 | |
| 5000 | 4.6 | 78 | 10,000 | 10.2 | 111.1 | |
| DON-3G | 400 | 13.8 | 102 | 800 | 13.2 | 100.8 |
| 800 | 15.6 | 74 | 1600 | 8.1 | 99.7 | |
| 1600 | 16.7 | 81 | 3200 | 11.2 | 109.7 | |
| ZEN | 320 | 3.8 | 79.3 | 320 | 14.1 | 109 |
| 630 | 4.1 | 81 | 630 | 6.4 | 97 | |
| 1260 | 4.5 | 87 | 1260 | 8.5 | 97 | |
| ZEN-14G | 64 | 8.1 | 80 | 64 | 12.3 | 102 |
| 126 | 4.3 | 83 | 126 | 8.2 | 111 | |
| 252 | 7 | 86 | 252 | 8.5 | 103 | |
| ZEN-14S | 64 | 9.5 | 84.4 | 64 | 18.5 | 103 |
| 126 | 7.1 | 80 | 126 | 16.3 | 109 | |
| 252 | 4.8 | 81 | 252 | 10.6 | 90 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryła, M.; Ksieniewicz-Woźniak, E.; Waśkiewicz, A.; Yoshinari, T.; Szymczyk, K.; Podolska, G.; Gwiazdowski, R.; Kubiak, K. Transformations of Selected Fusarium Toxins and Their Modified Forms During Malt Loaf Production. Toxins 2020, 12, 385. https://doi.org/10.3390/toxins12060385
Bryła M, Ksieniewicz-Woźniak E, Waśkiewicz A, Yoshinari T, Szymczyk K, Podolska G, Gwiazdowski R, Kubiak K. Transformations of Selected Fusarium Toxins and Their Modified Forms During Malt Loaf Production. Toxins. 2020; 12(6):385. https://doi.org/10.3390/toxins12060385
Chicago/Turabian StyleBryła, Marcin, Edyta Ksieniewicz-Woźniak, Agnieszka Waśkiewicz, Tomoya Yoshinari, Krystyna Szymczyk, Grażyna Podolska, Romuald Gwiazdowski, and Krzysztof Kubiak. 2020. "Transformations of Selected Fusarium Toxins and Their Modified Forms During Malt Loaf Production" Toxins 12, no. 6: 385. https://doi.org/10.3390/toxins12060385
APA StyleBryła, M., Ksieniewicz-Woźniak, E., Waśkiewicz, A., Yoshinari, T., Szymczyk, K., Podolska, G., Gwiazdowski, R., & Kubiak, K. (2020). Transformations of Selected Fusarium Toxins and Their Modified Forms During Malt Loaf Production. Toxins, 12(6), 385. https://doi.org/10.3390/toxins12060385