Type A Trichothecene Diacetoxyscirpenol-Induced Emesis Corresponds to Secretion of Peptide YY and Serotonin in Mink





Abstract
:1. Introduction
2. Results
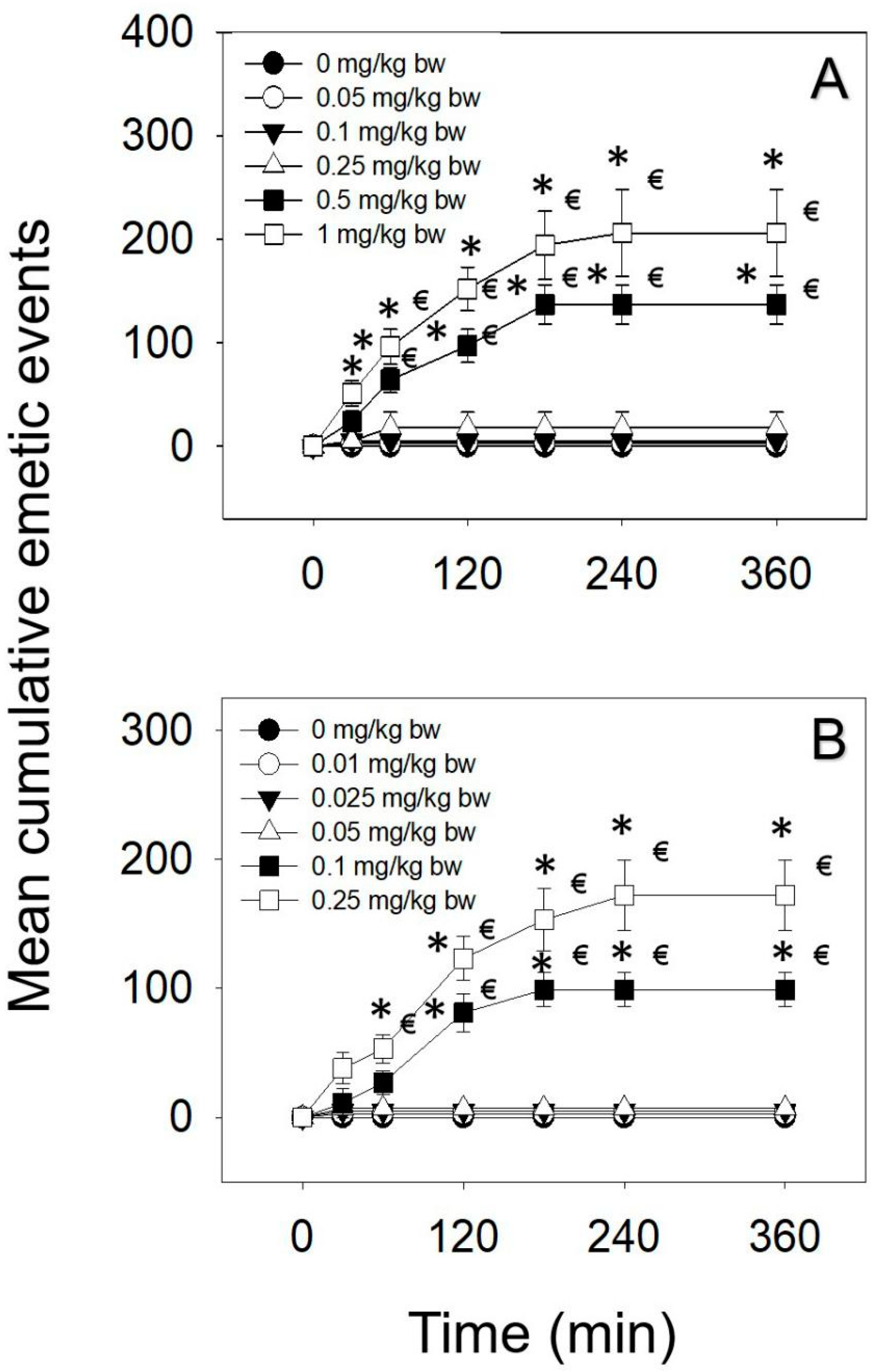
2.1. Emetic Effects of IP and Oral Exposure to DAS
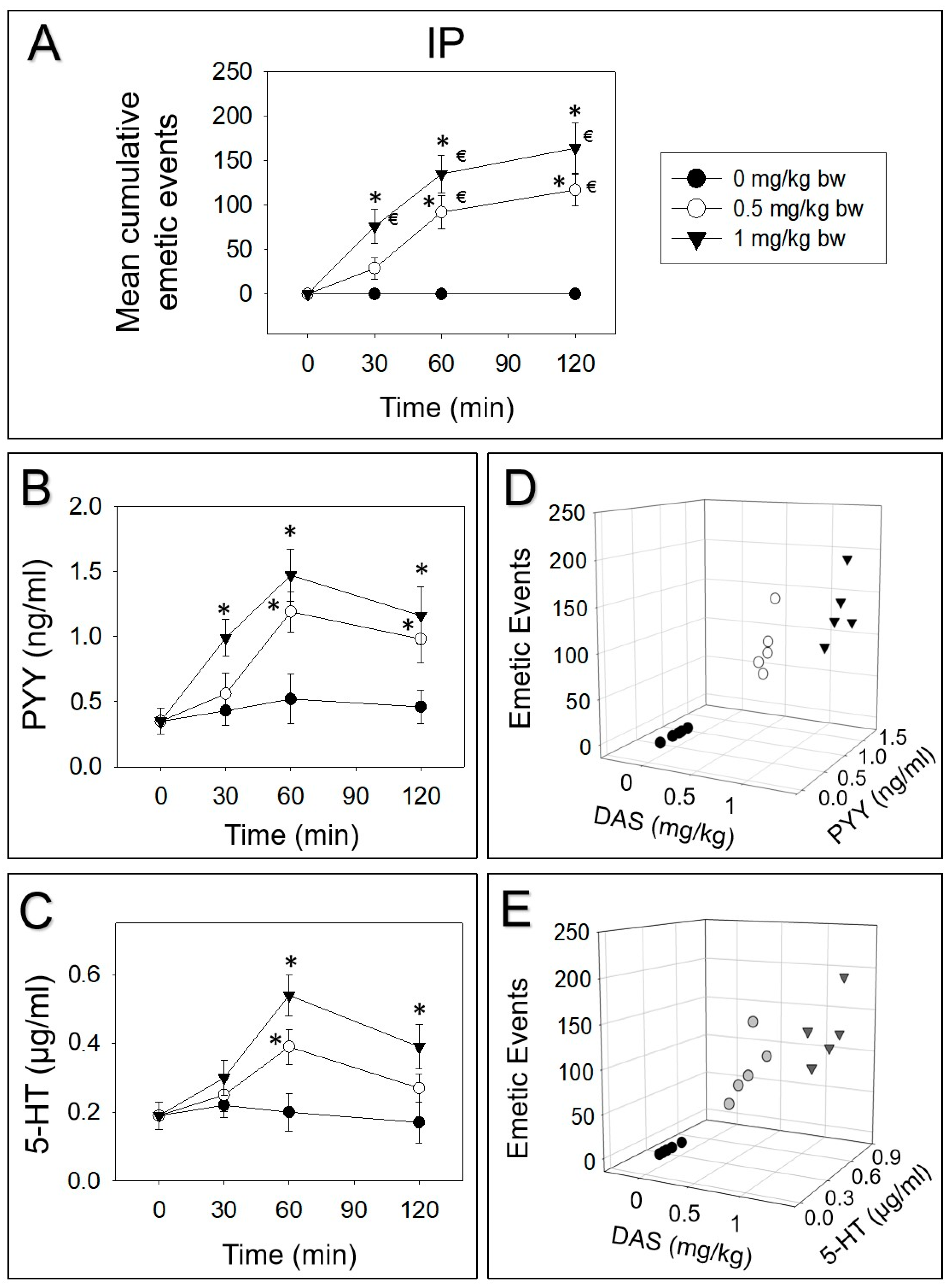
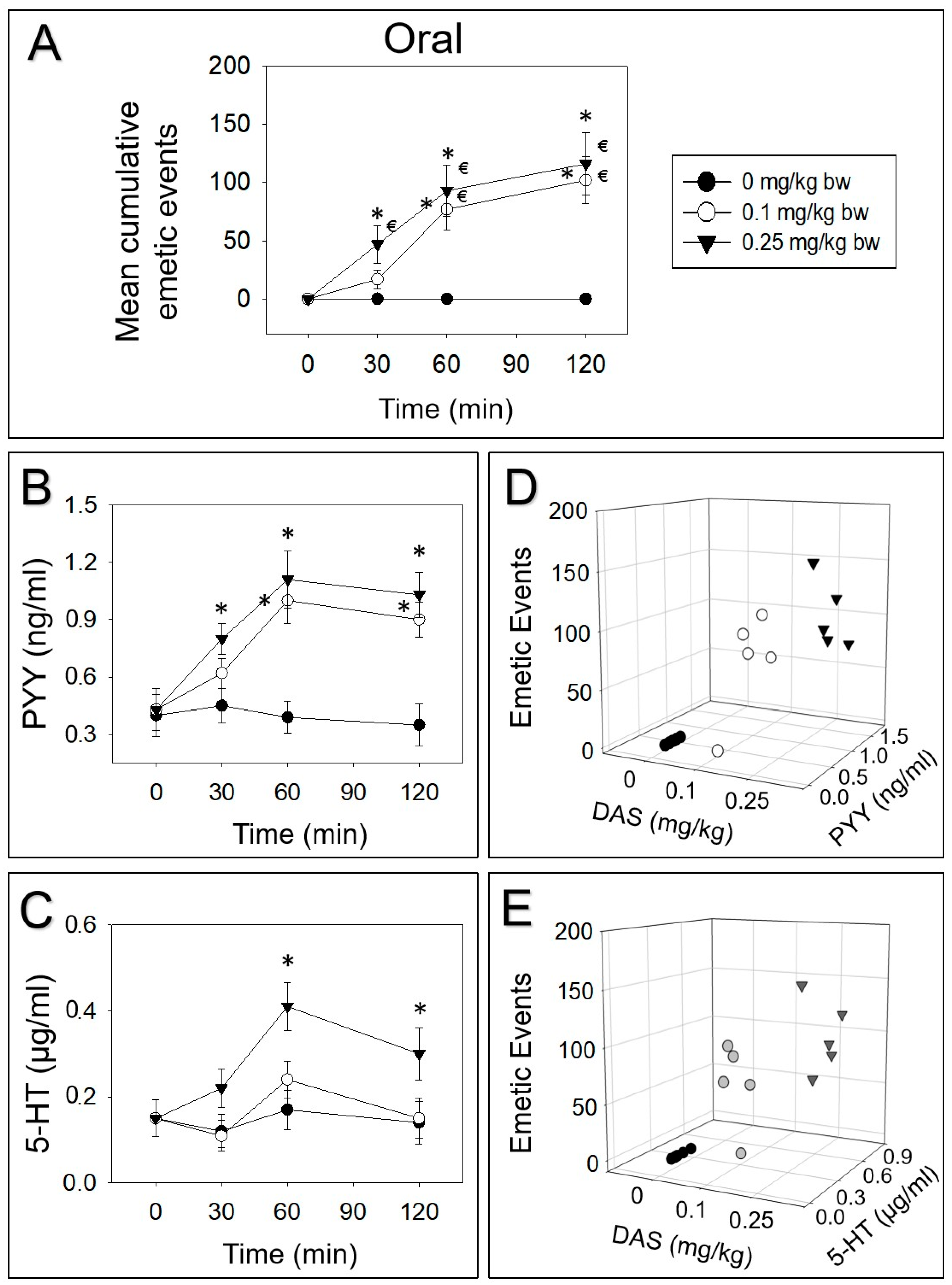
2.2. DAS-Induced Emetic Effect Corresponds to Elevation of Plasma PYY and 5-HT
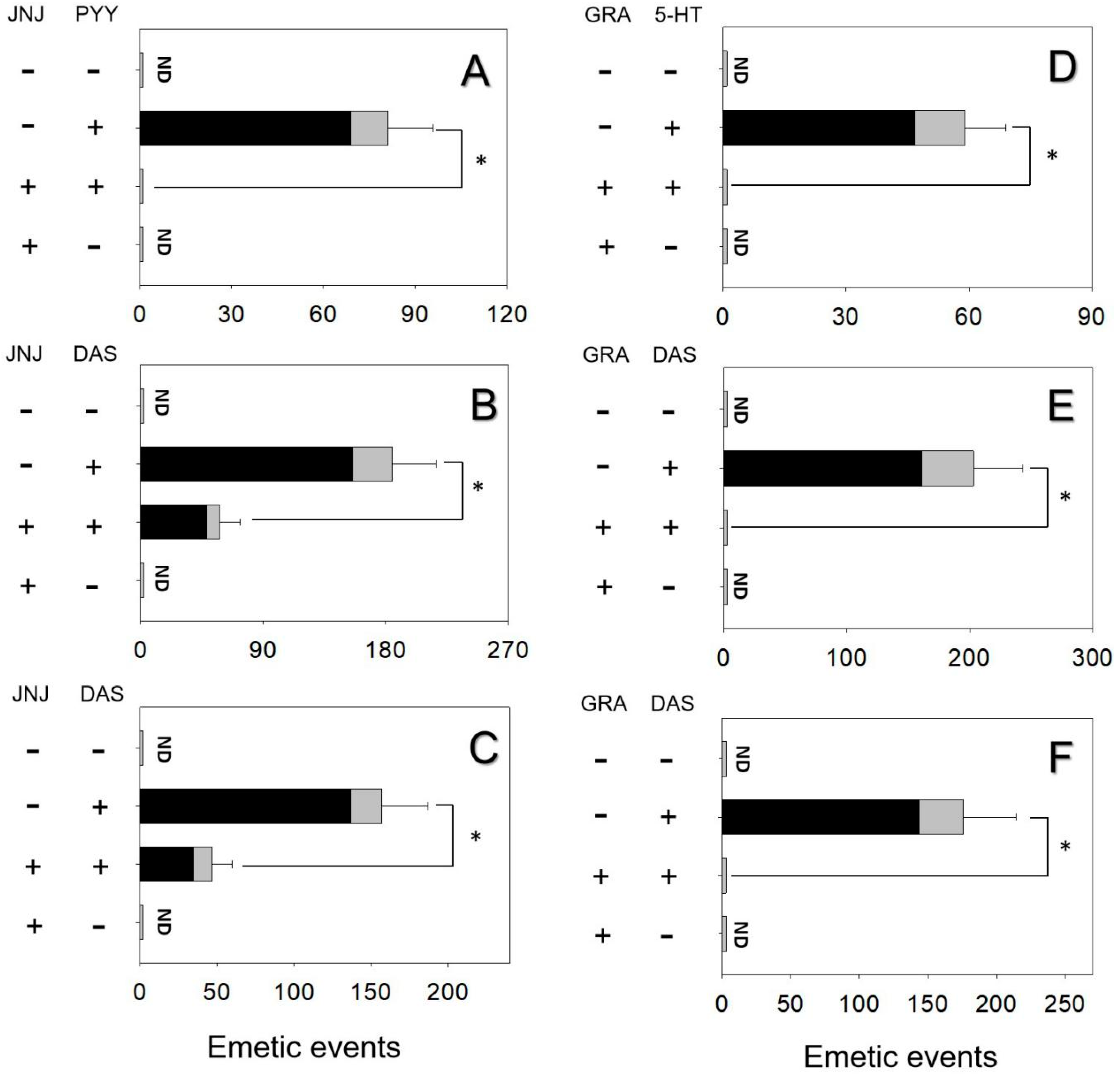
2.3. Effects of NPY2 and 5-HT3 Receptor Antagonists on DAS-Induced Emesis
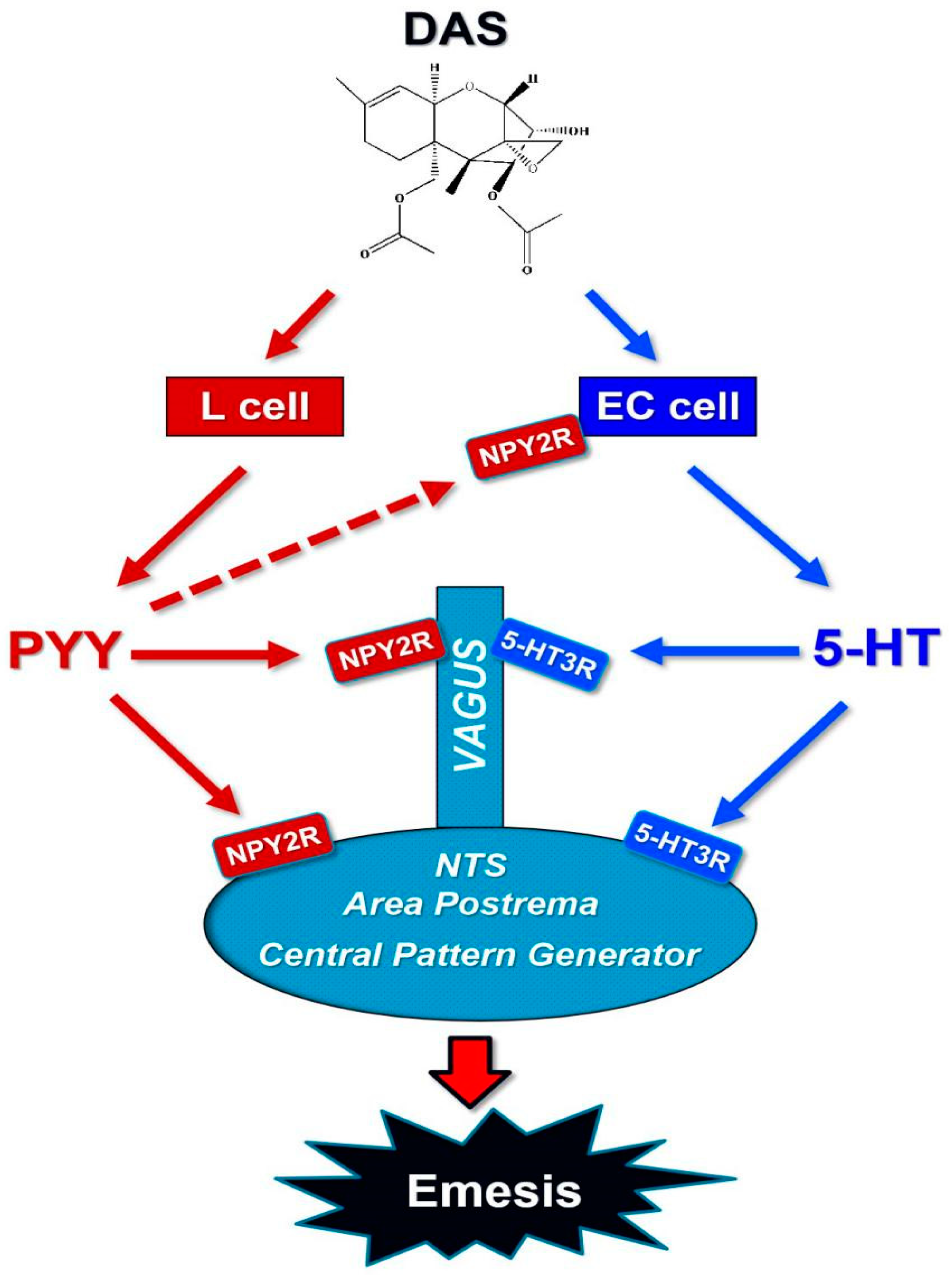
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Animals
5.2. Experimental Design
5.3. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Pestka, J.J. Toxicological mechanisms and potential health effects of deoxynivalenol and nivalenol. World Mycotoxin J. 2010, 3, 323–347. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, H.; Liu, S.; Wu, W.; Zhang, H. Comparison of anorectic potencies of type A Trichothecenes T-2 toxin, HT-2 toxin, Diacetoxyscirpenol, and Neosolaniol. Toxins 2018, 10, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- JECFA. Evaluation of certain contaminants in food: 72nd report of the joint FAO/WHO expert committee on food additives. In Deoxynivalenol; WHO Technical Report Series, No. 959; WHO Press: Geneva, Switzerland, 2011; pp. 37–48. [Google Scholar]
- Vesonder, R.F.; Ciegler, A.; Jensen, A.H. Isolation of the emetic principle from fusarium-infected corn. Appl. Microbiol. 1973, 26, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lysøe, E.; Frandsen, R.J.N.; Divon, H.H.; Terzi, V.; Orrù, L.; Lamontanara, A.; Kolseth, A.-K.; Nielsen, K.F.; Thrane, U. Draft genome sequence and chemical profiling of Fusarium langsethiae, an emerging producer of type A trichothecenes. Int. J. Food Microbiol. 2016, 221, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risk to human and animal health related to the presence of 4,15-diacetoxyscirpenol in food and feed. EFSA J. 2018, 16. [Google Scholar] [CrossRef] [Green Version]
- Andrews, P.L.R.; Davis, C.J.; Bingham, S.; Davidson, H.I.M.; Hawthorn, J.; Maskell, L. The abdominal visceral innervation and the emetic reflex: Pathways, pharmacology, and plasticity. Can. J. Physiol. Pharmacol. 1990, 68, 325–345. [Google Scholar] [CrossRef]
- Bauer, J.; Bollwahn, W.; Gareis, M.; Gedek, B.; Heinritzi, K. Kinetic profiles of diacetoxyscirpenol and two of its metabolites in blood serum of pigs. Appl. Environ. Microbiol. 1985, 49, 842–845. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.; Gareis, M.; Gedek, B. Metabolism of the trichothecenes T-2 toxin, diacetoxyscirpenol, and deoxynivalenol by farm animals. In Topics in Secondary Metabolism-Volume 2, Fusarium: Mycotoxins, Taxonomy and Pathogenicity; Chelkowski, J., Ed.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 139–165. [Google Scholar]
- Ueno, Y. General toxicology. In Trichothecenes: Chemical, Biological, and Toxicological Aspects; Ueno, Y., Ed.; Elsevier: New York, NY, USA, 1983; pp. 135–146. [Google Scholar]
- Coppock, R.W.; Gelberg, H.B.; Hoffmann, W.E.; Buck, W.B. The acute toxicopathy of intravenous diacetoxyscirpenol (anguidine) administration in swine. Fundam. Appl. Toxicol. 1985, 5, 1034–1049. [Google Scholar] [CrossRef]
- Coppock, R.W.; Hoffmann, W.E.; Gelberg, H.B.; Bass, D.; Buck, W.B. Hematologic changes induced by intravenous administration of diacetoxyscirpenol in pigs, dogs, and calves. Am. J. Veter-Res. 1989, 50, 411–415. [Google Scholar]
- Hornby, P.J. Central neurocircuitry associated with emesis. Am. J. Med. 2001, 111, 106S–112S. [Google Scholar] [CrossRef]
- Johnston, K.D.; Lu, Z.; Rudd, J.A. Looking beyond 5-HT3 receptors: A review of the wider role of serotonin in the pharmacology of nausea and vomiting. Eur. J. Pharmacol. 2014, 722, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Naylor, R.; Rudd, J.A. Mechanisms of chemotherapy/radiotherapy-induced emesis in animal models. Oncology 1996, 53, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Navari, R.M.; Aapro, M. Antiemetic prophylaxis for chemotherapy-induced nausea and vomiting. N. Engl. J. Med. 2016, 374, 1356–1367. [Google Scholar] [CrossRef]
- Koga, T.; Fukuda, H. Descending pathway from the central pattern generator of vomiting. NeuroReport 1997, 8, 2587–2590. [Google Scholar] [CrossRef]
- Le Roux, C.W.; Borg, C.-M.; Murphy, K.G.; Vincent, R.P.; Ghatei, M.A.; Bloom, S.R. Supraphysiological doses of intravenous PYY3-36 cause nausea, but no additional reduction in food intake. Ann. Clin. Biochem. Int. J. Lab. Med. 2008, 45, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Bates, M.A.; Bursian, S.J.; Flannery, B.; Zhou, H.-R.; Link, J.E.; Zhang, H.; Pestka, J.J. Peptide YY3-36 and 5-hydroxytryptamine mediate emesis induction by trichothecene deoxynivalenol (vomitoxin). Toxicol. Sci. 2013, 133, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Rojas, C.; Raje, M.; Tsukamoto, T.; Slusher, B.S. Molecular mechanisms of 5-HT3 and NK1 receptor antagonists in prevention of emesis. Eur. J. Pharmacol. 2014, 722, 26–37. [Google Scholar] [CrossRef]
- Hesketh, P.J. Chemotherapy-induced nausea and vomiting. N. Engl. J. Med. 2008, 358, 2482–2494. [Google Scholar] [CrossRef]
- Kovac, A.L. Comparative pharmacology and guide to the use of the serotonin 5-HT3 receptor antagonists for postoperative nausea and vomiting. Drugs 2016, 76, 1719–1735. [Google Scholar] [CrossRef]
- Du Bois, A.; Vach, W.; Kiechle, M.; Cramer-Giraud, U.; Meerpohl, H.G. Pathophysiology, Severity, Pattern, and Risk Factors for Carboplatin-induced Emesis. Oncology 1996, 53, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.R.; Rhee, J.; Smith, W.L. Plasma levels of peptide YY correlate with Cisplatin-induced emesis in dogs. J. Pharm. Pharmacol. 1994, 46, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Bates, M.A.; Bursian, S.J.; Link, J.E.; Flannery, B.M.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Comparison of emetic potencies of the 8-ketotrichothecenes deoxynivalenol, 15-acetyldeoxynivalenol, 3-acetyldeoxynivalenol, fusarenon X, and nivalenol. Toxicol. Sci. 2012, 131, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhou, H.-R.; Bursian, S.J.; Link, J.E.; Pestka, J.J. Emetic responses to T-2 toxin, HT-2 toxin and emetine correspond to plasma elevations of peptide YY3–36 and 5-hydroxytryptamine. Arch. Toxicol. 2015, 90, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhou, H.-R.; Bursian, S.J.; Pan, X.; Link, J.E.; Berthiller, F.; Adam, G.; Krantis, A.; Durst, T.; Pestka, J.J.; et al. Comparison of anorectic and emetic potencies of deoxynivalenol (vomitoxin) to the plant metabolite deoxynivalenol-3-glucoside and synthetic deoxynivalenol derivatives EN139528 and EN139544. Toxicol. Sci. 2014, 142, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Zhou, H.-R.; He, K.; Pan, X.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Role of cholecystokinin in anorexia induction following oral exposure to the 8-ketotrichothecenes deoxynivalenol, 15-acetyldeoxynivalenol, 3-acetyldeoxynivalenol, fusarenon x, and nivalenol. Toxicol. Sci. 2014, 138, 278–289. [Google Scholar] [CrossRef]
- Horn, C.C. Why is the neurobiology of nausea and vomiting so important? Appetite 2007, 50, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Du Sert, N.P.; Holmes, A.M.; Wallis, R.; Andrews, P.L. Predicting the emetic liability of novel chemical entities: A comparative study. Br. J. Pharmacol. 2012, 165, 1848–1867. [Google Scholar] [CrossRef] [Green Version]
- Du Sert, N.P.; Rudd, J.A.; Apfel, C.C.; Andrews, P.L.R. Cisplatin-induced emesis: Systematic review and meta-analysis of the ferret model and the effects of 5-HT3 receptor antagonists. Cancer Chemother. Pharmacol. 2010, 67, 667–686. [Google Scholar] [CrossRef] [Green Version]
- Kottschade, L.; Novotny, P.; Lyss, A.; Mazurczak, M.; Loprinzi, C.; Barton, D.L. Chemotherapy-induced nausea and vomiting: Incidence and characteristics of persistent symptoms and future directions NCCTG N08C3 (Alliance). Support. Care Cancer 2016, 24, 2661–2667. [Google Scholar] [CrossRef] [Green Version]
- Kris, M.G.; Gralla, R.J.; Clark, R.A.; Tyson, L.B.; O’Connell, J.P.; Wertheim, M.S.; Kelsen, D.P. Incidence, course, and severity of delayed nausea and vomiting following the administration of high-dose cisplatin. J. Clin. Oncol. 1985, 3, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Tohmola, N.; Johansson, A.; Sane, T.; Renkonen, R.; Hämäläinen, E.; Itkonen, O. Transient elevation of serum 5-HIAA by dietary serotonin and distribution of 5-HIAA in serum protein fractions. Ann. Clin. Biochem. Int. J. Lab. Med. 2014, 52, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Bearcroft, C.P.; Farthing, M.J.G.; Perrett, D. Determination of 5-hydroxytryptamine, 5-hydroxyindoleacetic acid and tryptophan in plasma and urine by HPLC with fluorimetric detection. Biomed. Chromatogr. 1995, 9, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.-I.; Tohei, A.; Anzai, N. A role for endogenous peptide YY in tachykinin NK2 receptor-triggered 5-HT release from guinea pig isolated colonic mucosa. Br. J. Pharmacol. 2012, 167, 1362–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prelusky, D.B.; Trenholm, H.L. The efficacy of various classes of anti-emetics in preventing deoxynivalenol-induced vomiting in swine. Nat. Toxins 1993, 1, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Fioramonti, J.; Dupuy, C.; Dupuy, J.; Bueno, L. The mycotoxin, deoxynivalenol, delays gastric emptying through serotonin-3 receptors in rodents. J. Pharmacol. Exp. Ther. 1993, 266, 1255–1260. [Google Scholar] [PubMed]
- Halatchev, I.G.; Cone, R.D. Peripheral administration of PYY3–36 produces conditioned taste aversion in mice. Cell Metab. 2005, 1, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Jia, H.; Wang, Q.; Wu, W.; Zhang, H. Role of peptide YY3-36 and glucose-dependent insulinotropic polypeptide in anorexia induction by trichothecences t-2 toxin, ht-2 toxin, diacetoxyscirpenol, and neosolaniol. Toxicol. Sci. 2017, 159, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Sheng, K.; Wu, W.; Zhang, H. Anorectic responses to T-2 toxin, HT-2 toxin, diacetoxyscirpenol and neosolaniol correspond to plasma elevations of neurotransmitters 5-hydroxytryptamine and substance P. Ecotoxicol. Environ. Saf. 2018, 161, 451–458. [Google Scholar] [CrossRef]
- Sheng, K.; Zhang, H.; Yue, J.; Gu, W.; Gu, C.; Zhang, H.; Wu, W. Anorectic response to the trichothecene T-2 toxin correspond to plasma elevations of the satiety hormone glucose-dependent insulinotropic polypeptide and peptide YY 3-36. Toxicology 2018, 28–36. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Route | Dose (mg/kg bw) | Incidence (Responding/Tested) | Latency (min) A,B | Duration (min) A,B | Retch (Times) | Vomit (Times) | Total (Times) |
|---|---|---|---|---|---|---|---|
| IP | 0 | 0/5 | - | - | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 0.05 | 0/5 | - | - | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| 0.1 | 0/5 | - | - | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| 0.25 | 1/5 | 42 ± 0 a | 3 ± 0 a | 7 ± 7 | 2 ± 2 | 9 ± 9 | |
| 0.5* | 5/5 | 27 ± 4 a | 120 ± 13 b | 122 ± 11 | 15 ± 8 | 137 ± 19 | |
| 1* | 5/5 | 18 ± 5 a | 189 ± 19 b | 181 ± 31 | 25 ± 11 | 206 ± 42 | |
| Oral | 0 | 0/5 | - | - | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 0.01 | 0/5 | - | - | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| 0.025 | 0/5 | - | - | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| 0.05 | 0/5 | - | - | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| 0.1* | 4/5 | 29 ± 5 a | 97 ± 11 a | 87 ± 9 | 12 ± 4 | 99 ± 13 | |
| 0.25* | 5/5 | 20 ± 2 a | 141 ± 23 a | 153 ± 16 | 19 ± 11 | 172 ± 27 |
| Toxin (mg/kg BW) | IP | Oral | ||||
|---|---|---|---|---|---|---|
| NOAELa | LOAELb | ED50c | NOAELa | LOAELb | ED50c | |
| T-2d | 0.01 | 0.05 | 0.05 | 0.005 | 0.05 | 0.02 |
| HT-2d | 0.01 | 0.05 | 0.05 | 0.005 | 0.05 | 0.02 |
| DAS | 0.1 | 0.25 | 0.3 | 0.05 | 0.1 | 0.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Kuca, K.; Nepovimova, E.; Wu, W. Type A Trichothecene Diacetoxyscirpenol-Induced Emesis Corresponds to Secretion of Peptide YY and Serotonin in Mink. Toxins 2020, 12, 419. https://doi.org/10.3390/toxins12060419
Wu Q, Kuca K, Nepovimova E, Wu W. Type A Trichothecene Diacetoxyscirpenol-Induced Emesis Corresponds to Secretion of Peptide YY and Serotonin in Mink. Toxins. 2020; 12(6):419. https://doi.org/10.3390/toxins12060419
Chicago/Turabian StyleWu, Qinghua, Kamil Kuca, Eugenie Nepovimova, and Wenda Wu. 2020. "Type A Trichothecene Diacetoxyscirpenol-Induced Emesis Corresponds to Secretion of Peptide YY and Serotonin in Mink" Toxins 12, no. 6: 419. https://doi.org/10.3390/toxins12060419
APA StyleWu, Q., Kuca, K., Nepovimova, E., & Wu, W. (2020). Type A Trichothecene Diacetoxyscirpenol-Induced Emesis Corresponds to Secretion of Peptide YY and Serotonin in Mink. Toxins, 12(6), 419. https://doi.org/10.3390/toxins12060419