Evaluation of the Effectiveness of Crotoxin as an Antiseptic against Candida spp. Biofilms

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
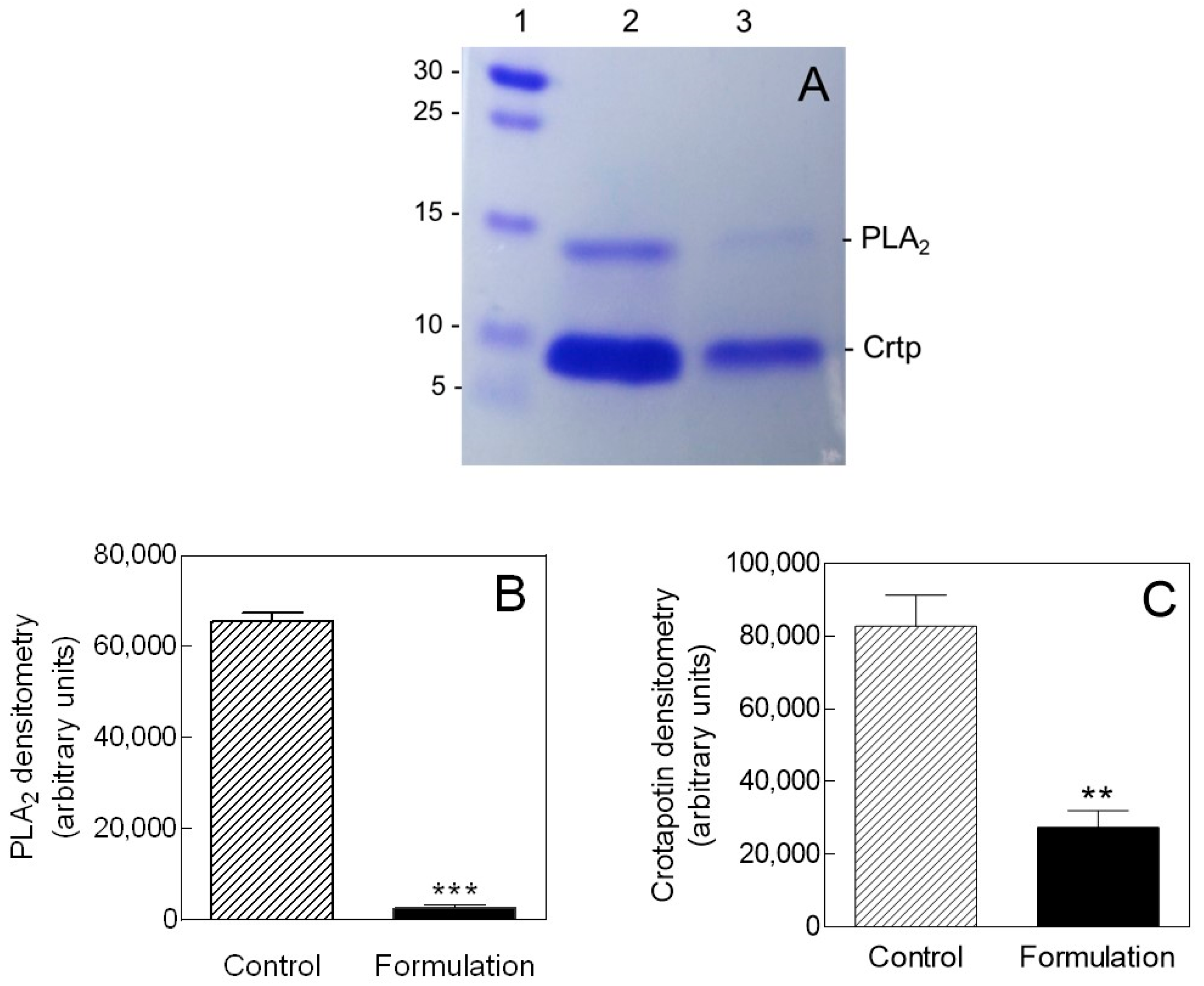
2.1. Isolation and Purification of Crotoxin
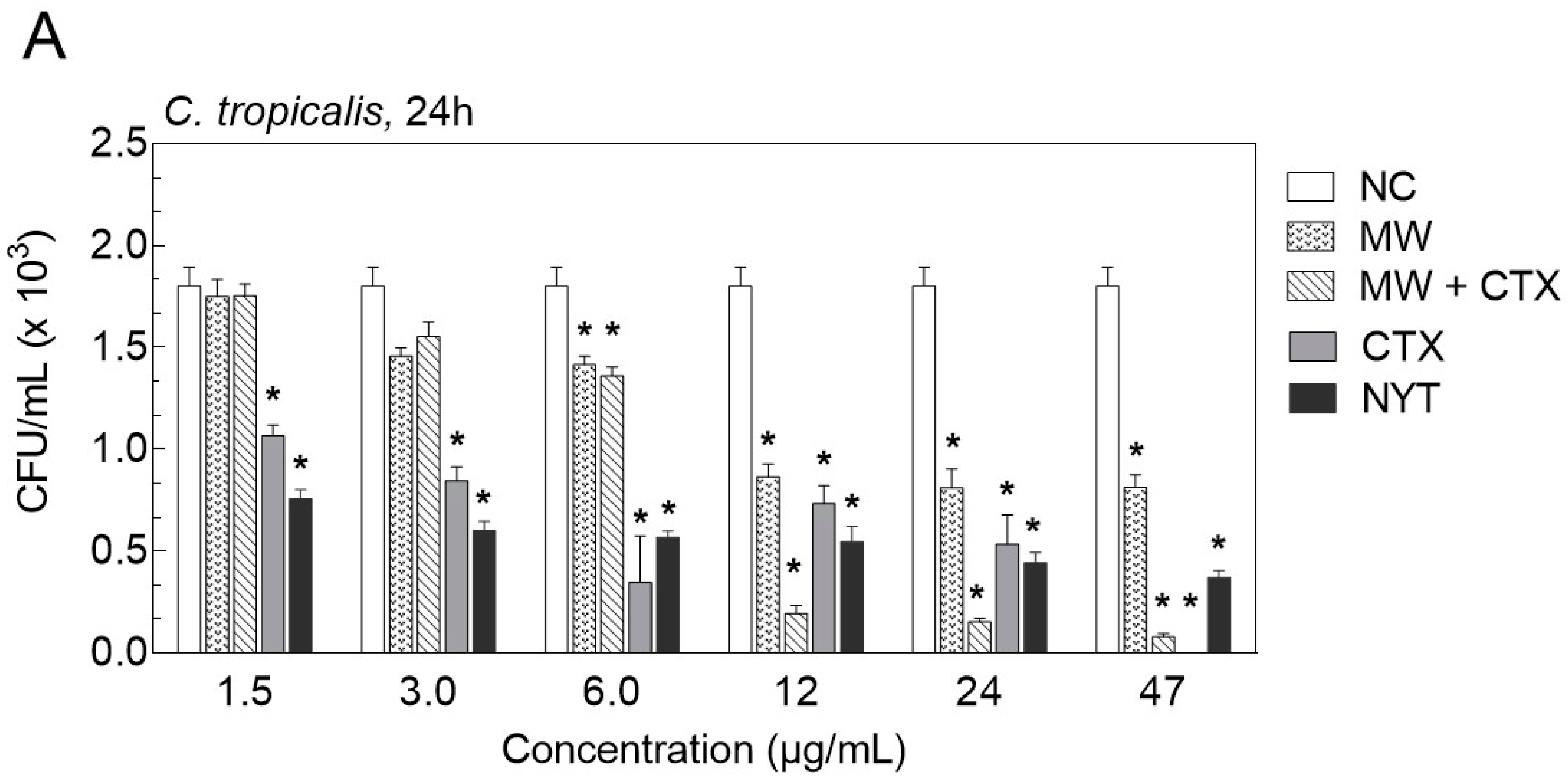
2.2. Antifungal Susceptibility Testing
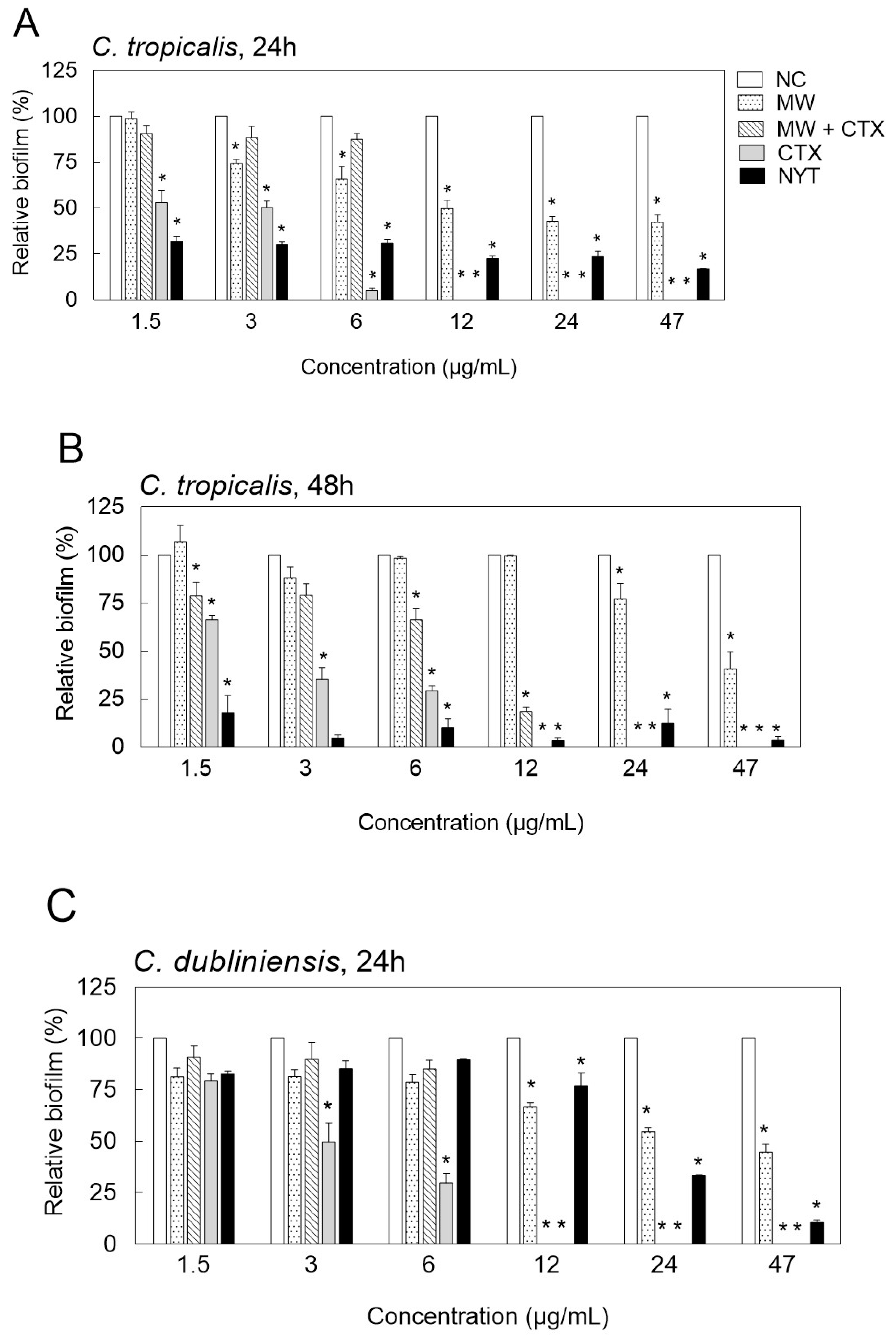
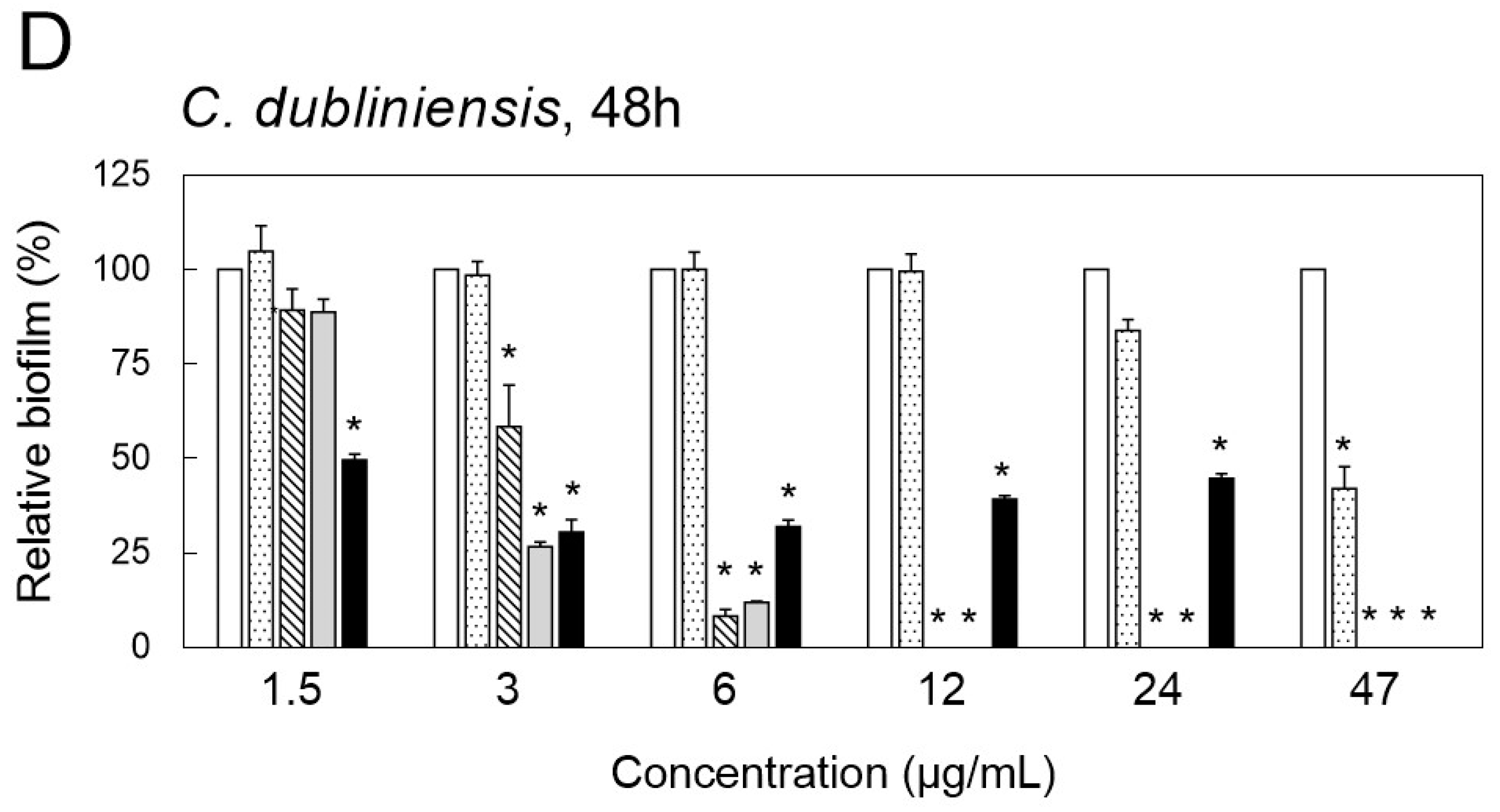
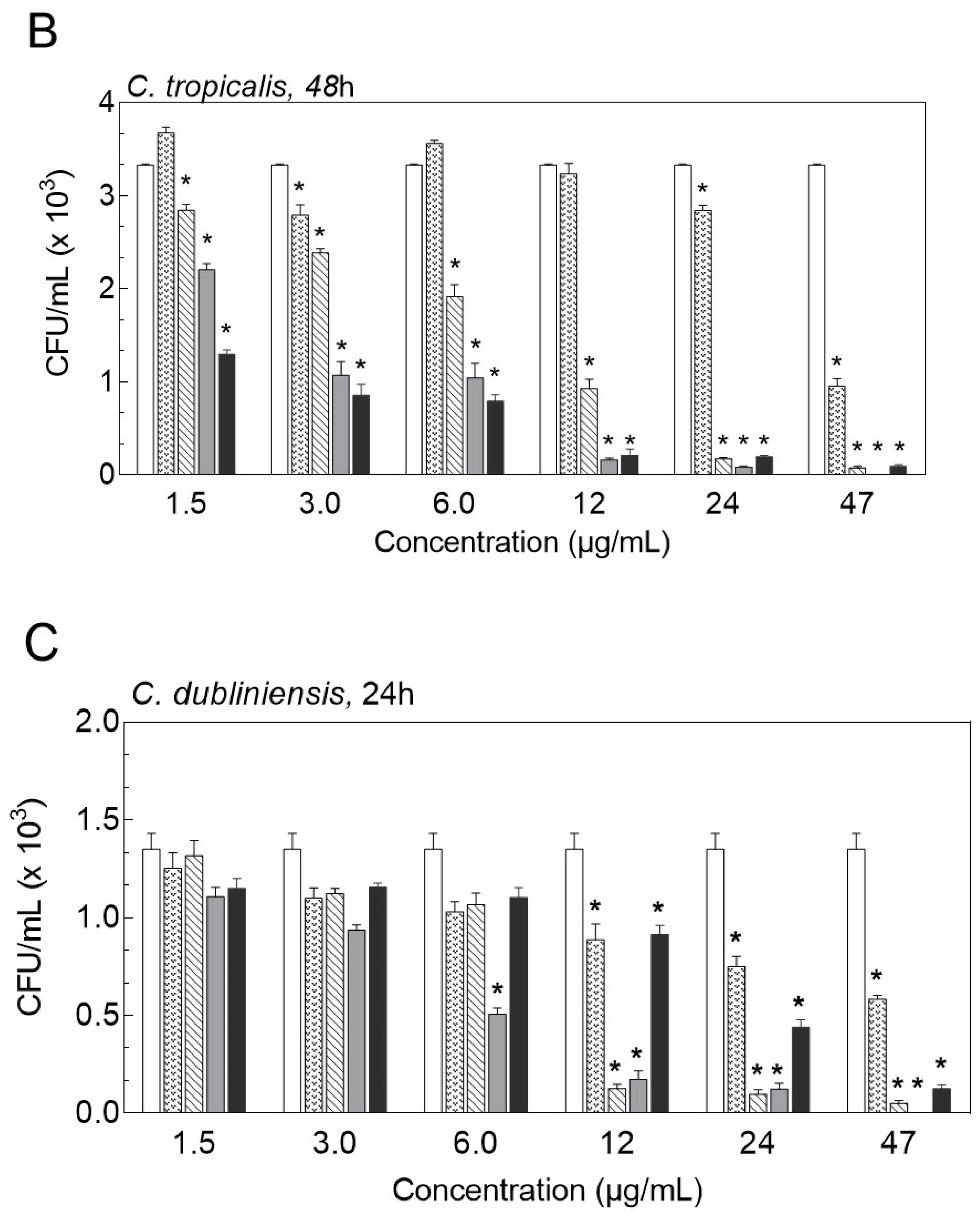
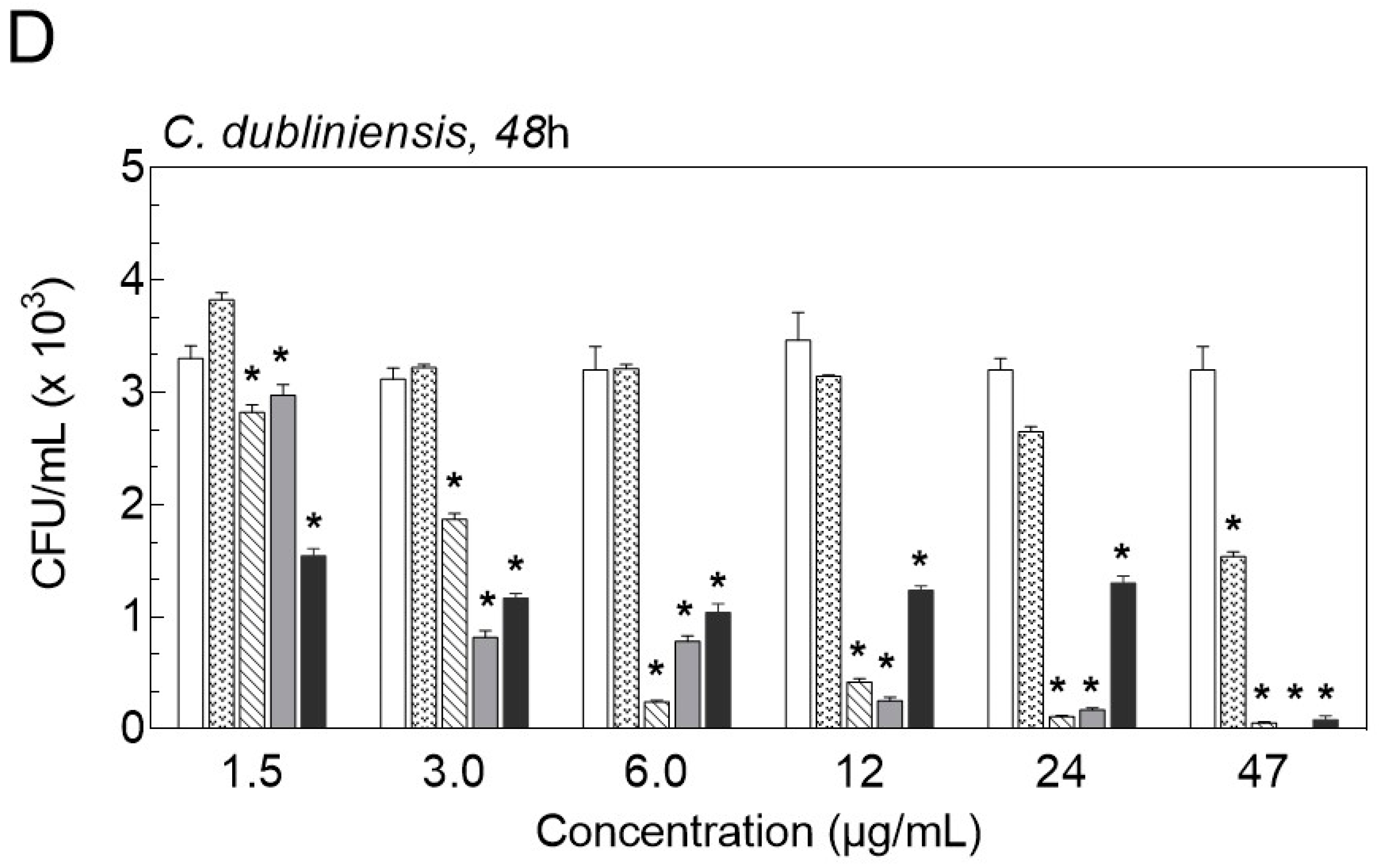
2.3. Inhibition of Biofilm Formation
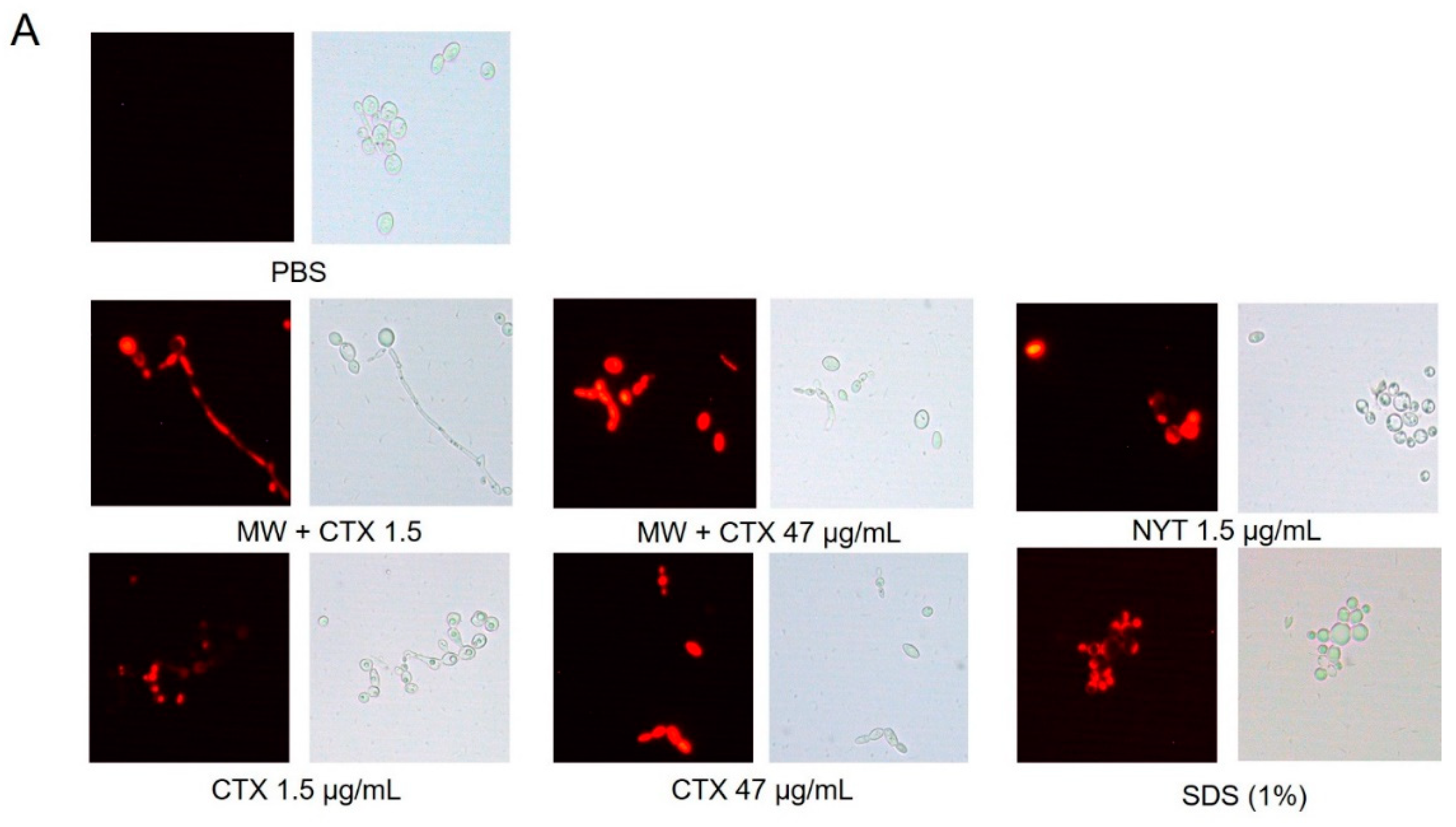
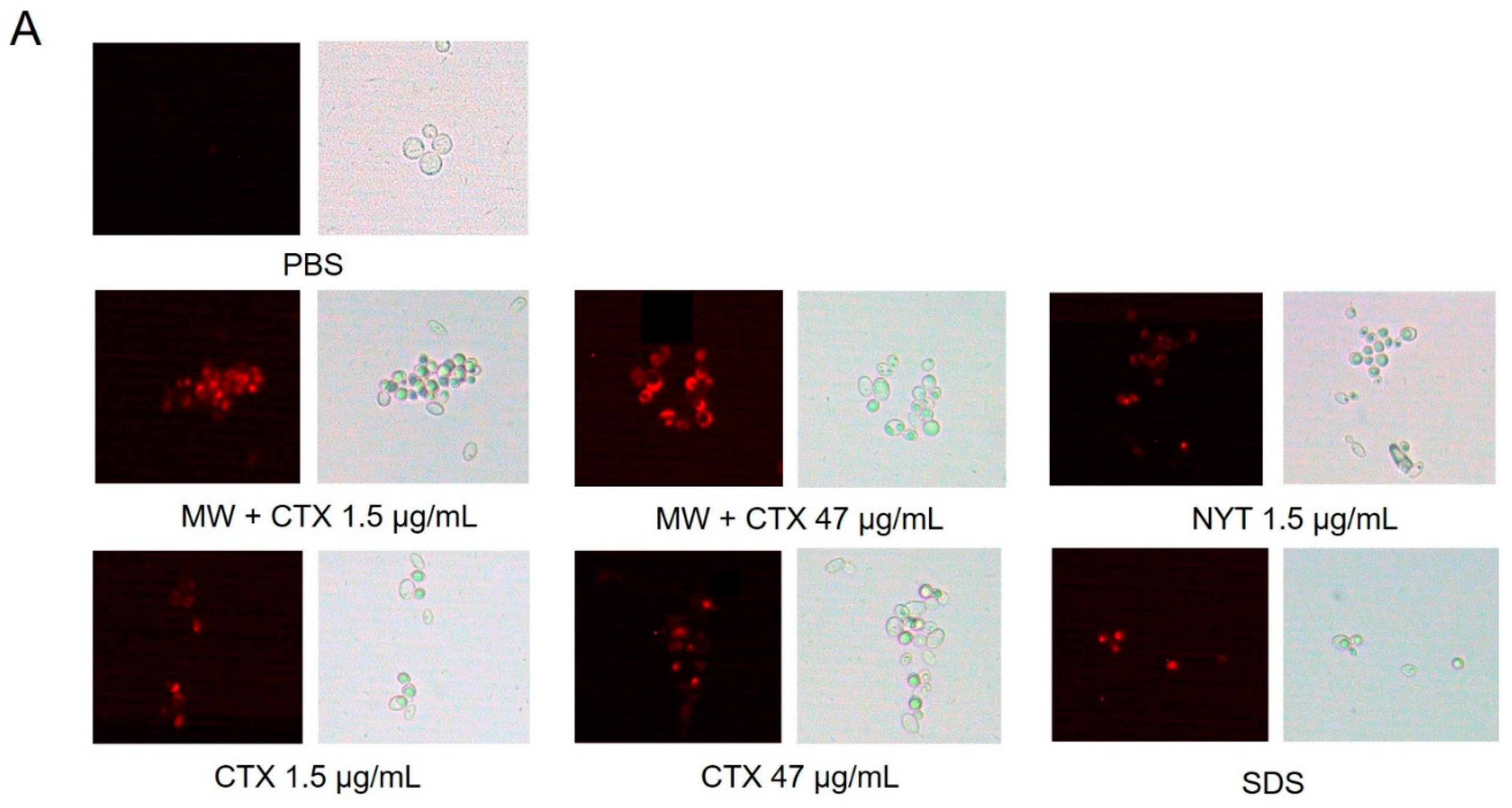
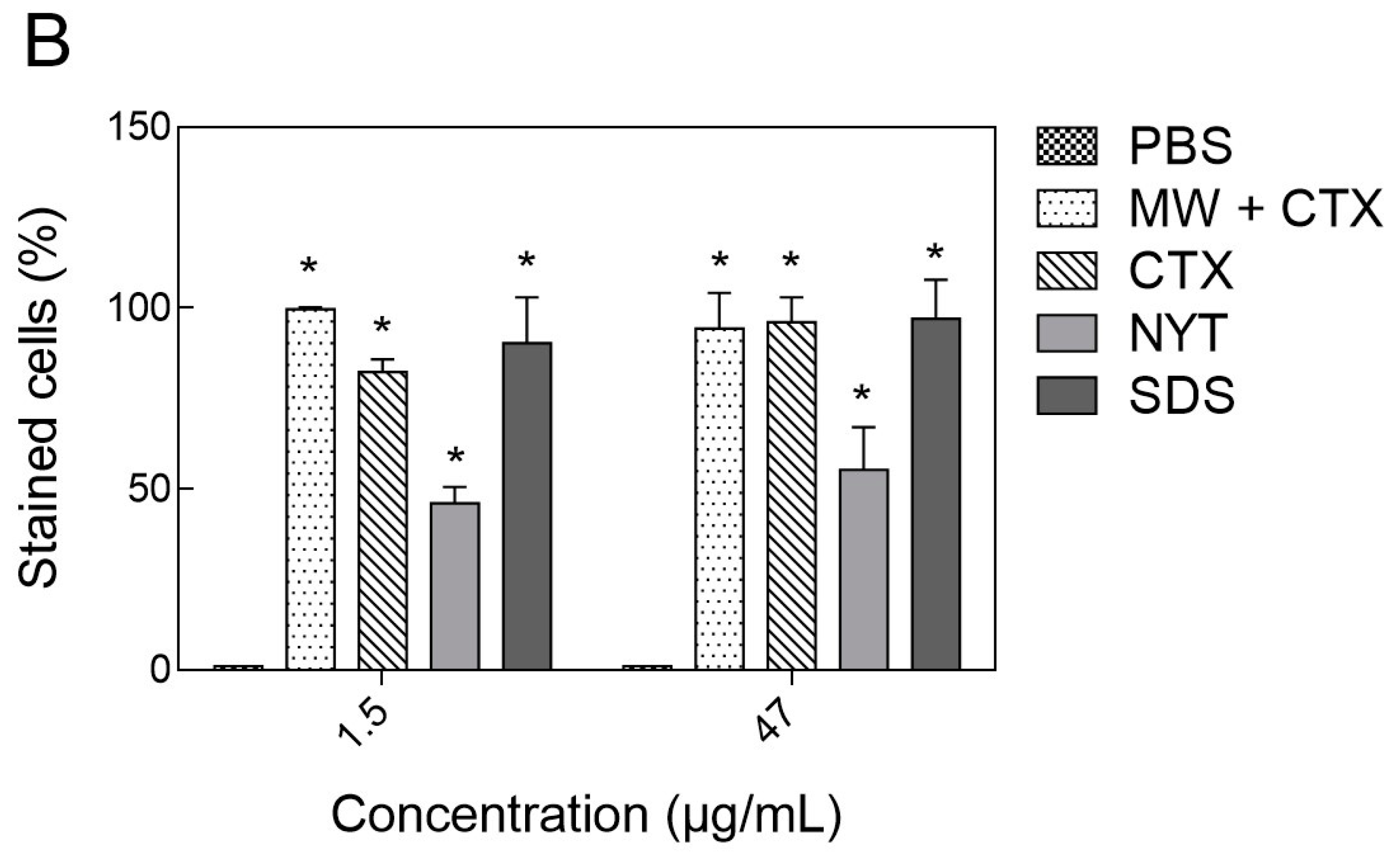
2.4. Plasma Membrane Integrity
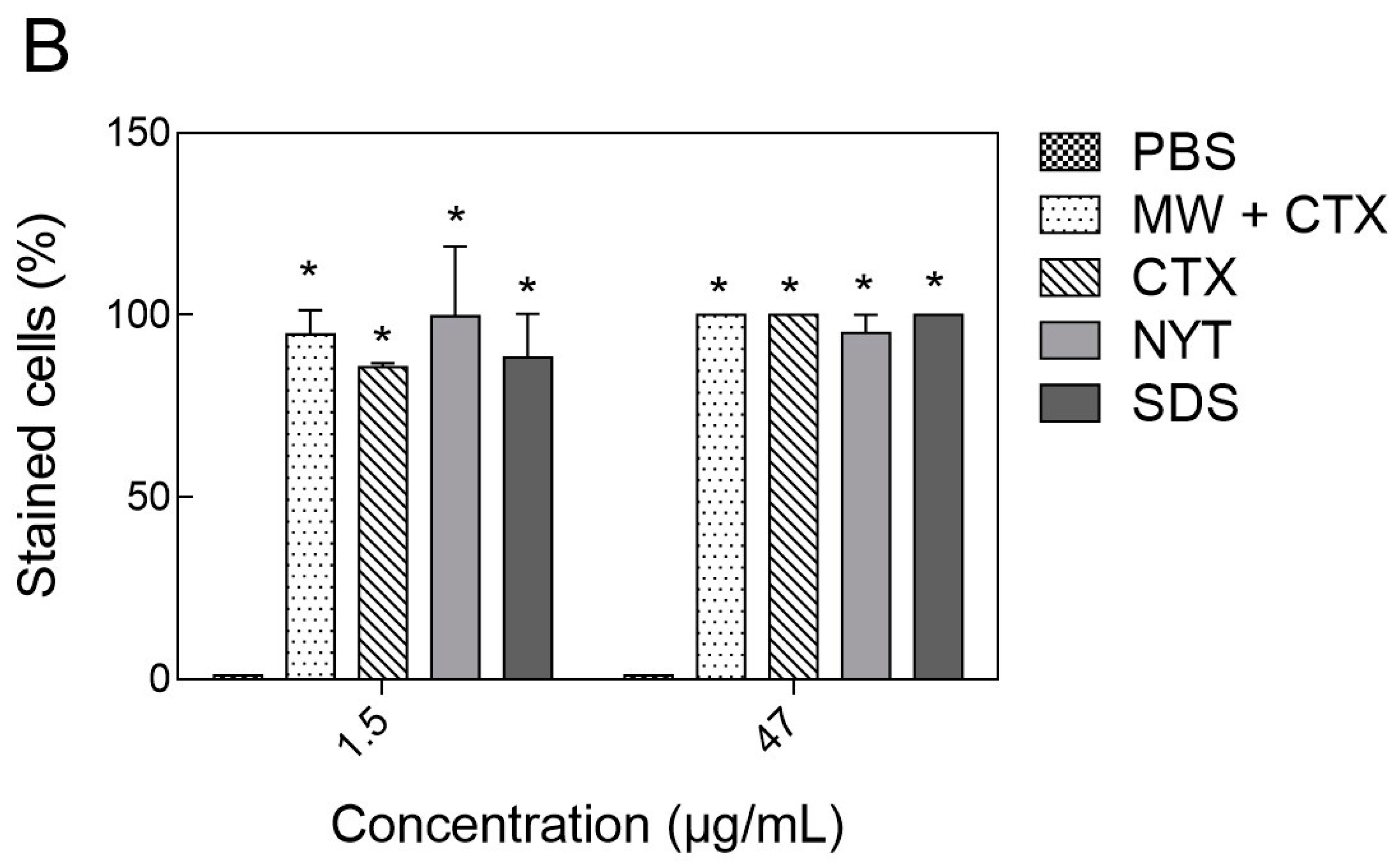
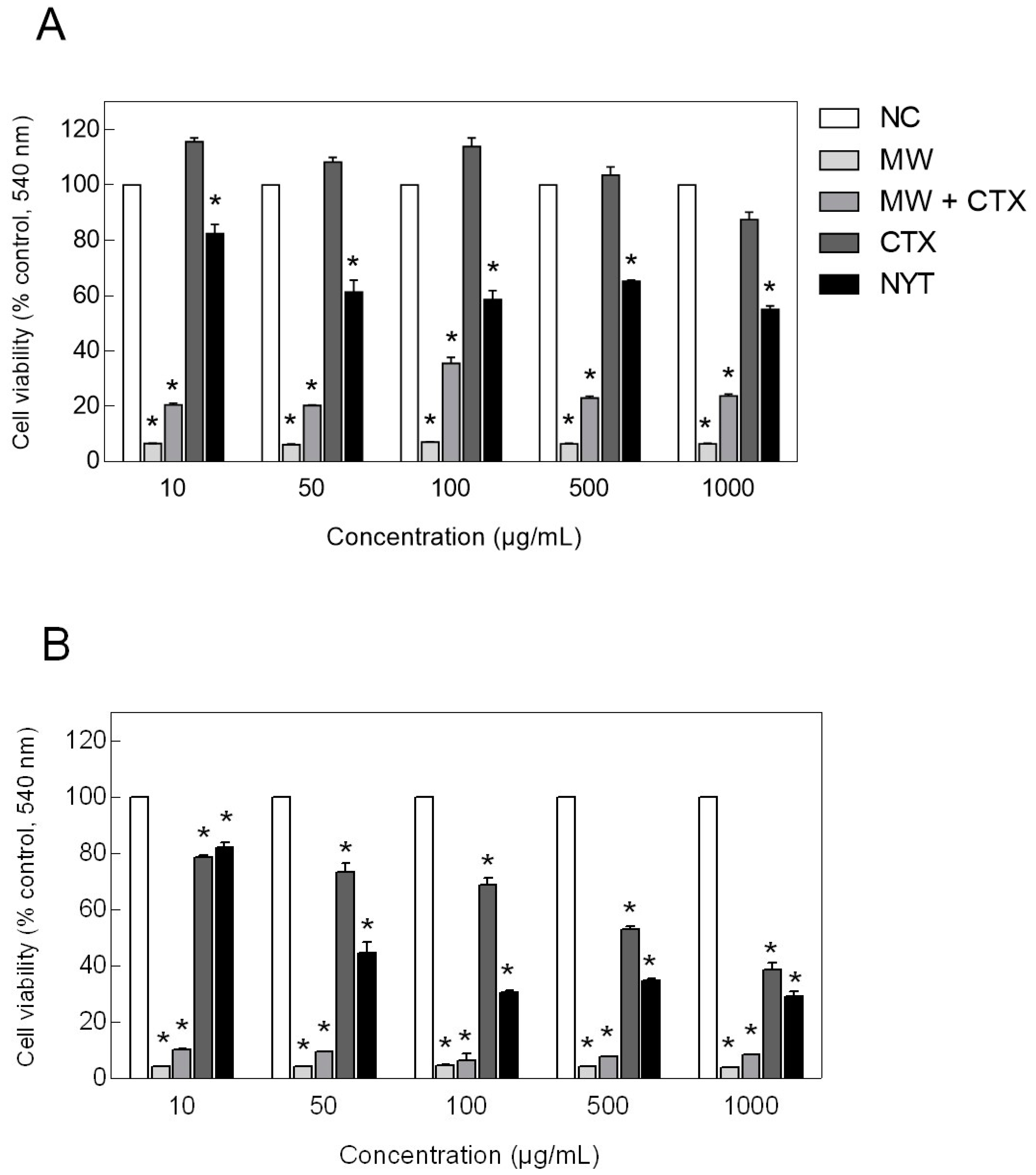
2.5. Cell Viability Assay
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Purification of Crotoxin
5.2. Preparation of a Crotoxin-Containing Mouthwash
5.3. Strains and Culture Conditions
5.4. Antifungal Susceptibility Testing
5.5. Biofilm Growth on Polystyrene and Adherence Inhibition
5.6. Plasma Membrane Permeability
5.7. Cell Viability Assay
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Faure, G.; Saul, F. Crystallographic characterization of functional sites of crotoxin and ammodytoxin, potent β-neurotoxins from Viperidae venom. Toxicon 2012, 60, 531–538. [Google Scholar] [CrossRef]
- Sampaio, S.C.; Hyslop, S.; Fontes, M.R.; Prado-Franceschi, J.; Zambelli, V.O.; Magro, A.J.; Brigatte, P.; Gutierrez, V.P.; Cury, Y. Crotoxin: Novel activities for a classic beta-neurotoxin. Toxicon 2010, 55, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, N.B.; Sant’Anna, M.B.; Giardini, A.C.; Araujo, L.P.; Fonseca, L.A.; Basso, A.S.; Cury, Y.; Picolo, G. Crotoxin down-modulates pro-inflammatory cells and alleviates pain on the MOG35-55-induced experimental autoimmune encephalomyelitis, an animal model of multiple sclerosis. Brain Behav. Immun. 2020, 84, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.S.; Kam, P.C. Chlorhexidine—Pharmacology and clinical applications. Anaesth. Intensive Care 2008, 36, 502–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorillo, L. Chlorhexidine gel use in the oral district: A systematic review. Gels 2019, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.; Fiehn, N.E. Dental biofilm infections—An update. Apmis 2017, 125, 376–384. [Google Scholar] [CrossRef]
- Bizerra, F.C.; Nakamura, C.V.; de Poersch, C.; Estivalet Svidzinski, T.I.; Borsato Quesada, R.M.; Goldenberg, S.; Krieger, M.A.; Yamada-Ogatta, S.F. Characteristics of biofilm formation by Candida tropicalis and antifungal resistance. FEMS Yeast Res. 2008, 8, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.M.; Jabra-Rizk, M.A.; O’May, G.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial interactions: Impact on pathogenesis and human disease. Clin. Microbiol. Rev. 2012, 25, 193–213. [Google Scholar] [CrossRef] [Green Version]
- Allison, D.L.; Willems, H.M.E.; Jayatilake, J.A.M.S.; Bruno, V.M.; Peters, B.M.; Shirtliff, M.E. Candida-Bacteria Interactions: Their Impact on Human Disease. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.; Seneviratne, C.J.; Koga-Ito, C.Y.; Samaranayake, L.P. Is the oral fungal pathogen Candida albicans a cariogen? Oral Dis. 2018, 24, 518–526. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Morales, P.; González, M.A.; Quindós, G. Candida dubliniensis, a new fungal pathogen. J. Basic Microbiol. 2002, 42, 207–227. [Google Scholar] [CrossRef] [Green Version]
- Gasparoto, T.H.; Dionísio, T.J.; de Oliveira, C.E.; Porto, V.C.; Gelani, V.; Santos, C.F.; Campanelli, A.P.; Lara, V.S. Isolation of Candida dubliniensis from denture wearers. J. Med. Microbiol. 2009, 58, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Meurman, J.H.; Pärnänen, P.; Seneviratne, C.J.; Samaranayake, L.P.; Saarinen, A.M.; Kari, K. Prevalence and antifungal drug sensitivity of non-albicans Candida in oral rinse samples of self-caring elderly. Gerodontology 2010, 28, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Zomorodian, K.; Haghighi, N.N.; Rajaee, N.; Pakshir, K.; Tarazooie, B.; Vojdani, M.; Sedaghat, F.; Vosoghi, M. Assessment of Candida species colonization and denture-related stomatitis in complete denture wearers. Med. Mycol. 2011, 4, 208–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothavade, R.J.; Kura, M.M.; Valand, A.G.; Panthaki, M.H. Candida tropicalis: Its prevalence, pathogenicity and increasing resistance to fluconazole. J. Med. Microbiol. 2010, 59, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, A.; Sansinenea, E. The Chemistry of Drugs to Treat Candida albicans. Curr. Top. Med. Chem. 2019, 9, 2554–2566. [Google Scholar] [CrossRef]
- Primon-Barros, M.; Macedo, A.J. Animal Venom Peptides: Potential for new antimicrobial agents. Curr. Top. Med. Chem. 2017, 17, 1119–1156. [Google Scholar] [CrossRef]
- Almeida, J.R.; Palacios, A.L.V.; Patiño, R.S.P.; Mendes, B.; Teixeira, C.A.S.; Gomes, P.; da Silva, S.L. Harnessing snake venom phospholipases A2 to novel approaches for overcoming antibiotic resistance. Drug Dev. Res. 2019, 80, 68–85. [Google Scholar] [CrossRef] [Green Version]
- Naidu, K.T.; Prabhu, N.P. Protein-surfactant interaction: Sodium dodecyl sulfate-induced unfolding of ribonuclease A. J. Phys. Chem. B 2011, 115, 14760–14767. [Google Scholar] [CrossRef]
- Singh, A.; Prasad, T.; Kapoor, K.; Mandal, A.; Roth, M.; Welti, R.; Prasad, R. Phospholipidome of Candida: Each species of Candida has distinctive phospholipid molecular species. OMICS J. Integr. Biol. 2010, 14, 665–677. [Google Scholar] [CrossRef]
- Raut, J.; Rathod, V.; Karuppayil, S.M. Cell surface hydrophobicity and adhesion: A study on fifty clinical isolates of Candida albicans. Nihon Ishinkin Gakkai Zasshi 2010, 51, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, U.; Khan, M.A. Targeting the GPI biosynthetic pathway. Pathog. Glob. Health 2018, 112, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Magnani, M.; Castro-Gómez, R.J.H. Beta-glucana from Saccharomyces cerevisiae: Constitution, bioactivity and obtaining. Semin. Ciênc. Agrár. 2008, 29, 631–650. [Google Scholar] [CrossRef]
- Faure, G.; Guillaume, J.L.; Camoin, L.; Saliou, B.; Bon, C. Multiplicity of acidic subunit isoforms of crotoxin, the phospholipase A2 neurotoxin from Crotalus durissus terrificus venom, results from posttranslational modifications. Biochemistry 1991, 30, 8074–8083. [Google Scholar] [CrossRef] [PubMed]
- Rudd, C.J.; Viskatis, L.J.; Vidal, J.C.; Etcheverry, M.A. In vitro comparison of cytotoxic effects of crotoxin against three human tumors and a normal human epidermal keratinocyte cell line. Investig. New Drugs 1994, 12, 183–184. [Google Scholar] [CrossRef]
- Cvikl, B.; Lussi, A.; Gruber, R. The in vitro impact of toothpaste extracts on cell viability. Eur. J. Oral Sci. 2015, 123, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Freitas, T.V.; Frézard, F. Encapsulation of native crotoxin in liposomes: A safe approach for the production of antivenom and vaccination against Crotalus durissus terrificus venom. Toxicon 1997, 35, 91–100. [Google Scholar] [CrossRef]
- Magalhães, T.; Proietti Viotti, A.; Teperino Gomes, R.; Viana de Freitas, T. Effect of membrane composition and of co-encapsulation of immunostimulants in a liposome-entrapped crotoxin. Biotechnol. Appl. Biochem. 2001, 33, 61–64. [Google Scholar] [CrossRef]
- Ferguson, E.L.; Richardson, S.C.W.; Duncan, R. Studies on the mechanism of action of dextrin−phospholipase A2 and its suitability for use in combination therapy. Mol. Pharm. 2010, 7, 510–521. [Google Scholar] [CrossRef]
- Bianco, I.D.; Daniele, J.J.; Delgado, C.; Fisher, D.; Francis, G.E.; Fidelio, G.D. Coupling reaction and properties of poly(ethylene glycol)-linked phospholipases A2. Biosci. Biotechnol. Biochem. 2002, 66, 722–729. [Google Scholar] [CrossRef]
- Ye, B.; Xie, Y.; Qin, Z.H.; Wu, J.C.; Han, R.; He, J.K. Anti-tumor activity of CrTX in human lung adenocarcinoma cell line A549. Acta Pharmacol. Sin. 2011, 32, 1397–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqui, A.A.; Horrocks, L.A. Phospholipase A2-generated lipid mediators in the brain: The good, the bad, and the ugly. Neuroscientist 2006, 12, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Taketomi, Y. Secreted phospholipase A2 and mast cells. Allergol. Int. 2015, 64, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartim, M.A.; Menaldo, D.L.; Sampaio, S.V. Immunotherapeutic potential of Crotoxin: Anti-inflammatory and immunosuppressive properties. J. Venom. Anim. Toxins Trop. Dis. 2018, 24, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT cells as a reliable in vitro differentiation model to dissect the inflammatory/repair response of human keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Muller, S.P.; Silva, V.A.O.; Silvestrini, A.V.P.; de Macedo, L.H.; Caetano, G.F.; Reis, R.M.; Mazzi, M.V. Crotoxin from Crotalus durissus terrificus venom: In vitro cytotoxic activity of a heterodimeric phospholipase A2 on human cancer-derived cell lines. Toxicon 2018, 156, 13–22. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed.; CLSI standard M27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Marak, M.B.; Dhanashree, B. Antifungal Susceptibility and Biofilm Production of Candida spp. Isolated from Clinical Samples. Int. J. Microbiol. 2018, 2018, 7495218. [Google Scholar] [CrossRef] [Green Version]
- Hafidh, R.R.; Abdulamir, A.S.; Vern, L.S.; Abu Bakar, F.; Abas, F.; Jahanshiri, F.; Sekawi, Z. Inhibition of growth of highly resistant bacterial and fungal pathogens by a natural product. Open Microbiol. J. 2011, 5, 96–106. [Google Scholar] [CrossRef]
- Peters, B.M.; Ward, R.M.; Rane, H.S.; Lee, S.A.; Noverr, M.C. Efficacy of ethanol against Candida albicans and Staphylococcus aureus polymicrobial biofilms. Antimicrob. Agents Chemother. 2013, 57, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Suchodolski, J.; Feder-Kubis, J.; Krasowska, A. Antifungal activity of ionic liquids based on (-)-menthol: A mechanism study. Microbiol. Res. 2017, 197, 56–64. [Google Scholar] [CrossRef]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival: Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Strain | NYT | CTX | MW + CTX | MW | p Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | ||
| C. tropicalis | 375 | 375 | 47 | 47 | 750 | 750 | 1500 | 1500 | <0.05 |
| C. dubliniensis | 375 | 375 | 47 | 47 | 750 | 3000 | 1500 | 1500 | <0.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canelli, A.P.; dos Santos Rodrigues, T.F.; de Goes, V.F.F.; Caetano, G.F.; Mazzi, M.V. Evaluation of the Effectiveness of Crotoxin as an Antiseptic against Candida spp. Biofilms. Toxins 2020, 12, 532. https://doi.org/10.3390/toxins12090532
Canelli AP, dos Santos Rodrigues TF, de Goes VFF, Caetano GF, Mazzi MV. Evaluation of the Effectiveness of Crotoxin as an Antiseptic against Candida spp. Biofilms. Toxins. 2020; 12(9):532. https://doi.org/10.3390/toxins12090532
Chicago/Turabian StyleCanelli, Amanda Pissinatti, Taís Fernanda dos Santos Rodrigues, Vivian Fernandes Furletti de Goes, Guilherme Ferreira Caetano, and Maurício Ventura Mazzi. 2020. "Evaluation of the Effectiveness of Crotoxin as an Antiseptic against Candida spp. Biofilms" Toxins 12, no. 9: 532. https://doi.org/10.3390/toxins12090532
APA StyleCanelli, A. P., dos Santos Rodrigues, T. F., de Goes, V. F. F., Caetano, G. F., & Mazzi, M. V. (2020). Evaluation of the Effectiveness of Crotoxin as an Antiseptic against Candida spp. Biofilms. Toxins, 12(9), 532. https://doi.org/10.3390/toxins12090532