The Role of Escherichia coli Shiga Toxins in STEC Colonization of Cattle

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Target Cells for Shiga Toxins in Cattle
2.1. Peripheral Lymphocytes
2.1.1. Receptors
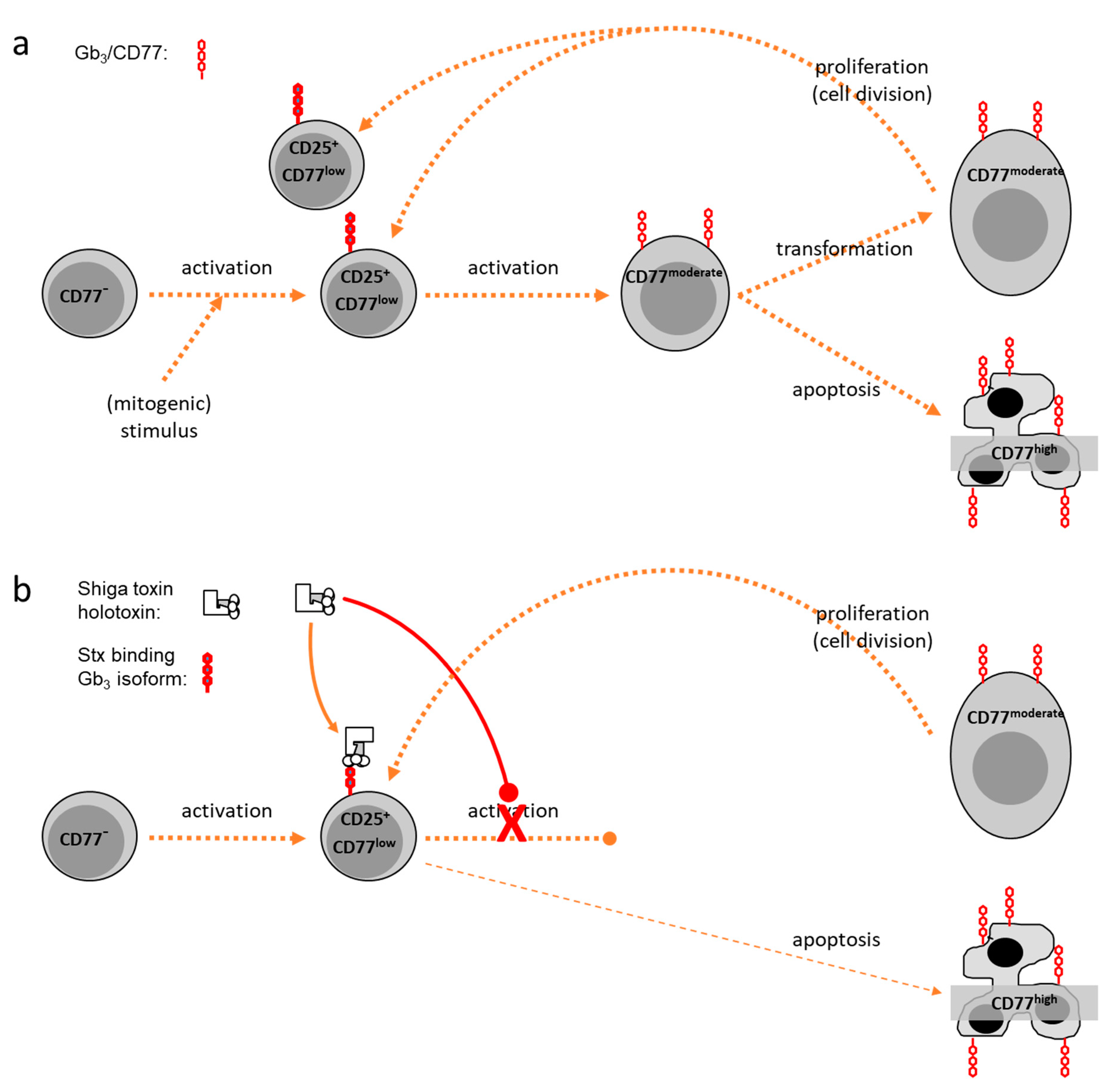
CD77—An Activation-Dependent Surface Antigen
Biochemical Characterisation of Bovine Gb3/CD77
Binding and Internalization of Shiga Toxin and Its B-Subunit
2.1.2. Effects of Shiga Toxins In Vitro
2.2. Intestinal Intraepithelial and Mucosa-Associated Lymphocytes
2.2.1. Stx Receptor Expression by Subsets
2.2.2. Functional Implications of Stx Receptor Expression In Vitro
2.3. Macrophages
2.4. Granulocytes
2.5. Endothelial Cells
2.6. Intestinal Epithelial Cells
3. Intestinal Immunomodulation Upon STEC Infection of Cattle
4. Systemic Effects of Shiga Toxins on the Bovine Immune System
4.1. Impact on the Humoral Immune Response
4.2. Impact on the Cellular Immune Response
5. Concept for the Role of Shiga Toxins in STEC Colonization of Cattle
6. Presumptive Drivers of STEC Preservation and Evolution in Cattle
6.1. Acquisition of Other Virulence-Associated Genes
6.2. Differential Regulation of Virulence Genes, Acquisition, and Loss of Metabolic Traits
6.3. Exchange of Immunodominant Surface Structures
6.4. Antiviral Activity of Shiga Toxins
6.5. Anti-Protozoal Activity of Shiga Toxins
6.6. Horizontal Transfer of STEC and Super-Shedding Events in Cattle Herds
6.7. Vertical Transfer of STEC in Cattle Herds
7. Options for Countermeasures to the Immunomodulatory Effects of Shiga Toxins in Cattle
8. Conclusive Summary
Funding
Acknowledgments
Conflicts of Interest
References
- Hajishengallis, G.; Nawar, H.; Tapping, R.I.; Russell, M.W.; Connell, T.D. The Type II heat-labile enterotoxins LT-IIa and LT-IIb and their respective B pentamers differentially induce and regulate cytokine production in human monocytic cells. Infect. Immun. 2004, 72, 6351–6358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, C.P.; Ghaem-Magami, M.; Petrovska, L.; Lopes, L.; Chain, B.M.; Williams, N.A.; Dougan, G. Immunomodulation using bacterial enterotoxins. Scand. J. Immunol. 2001, 53, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Tapping, R.I.; Martin, M.H.; Nawar, H.; Lyle, E.A.; Russell, M.W.; Connell, T.D. Toll-like receptor 2 mediates cellular activation by the B subunits of type II heat-labile enterotoxins. Infect. Immun. 2005, 73, 1343–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heim, J.B.; Hodnik, V.; Heggelund, J.E.; Anderluh, G.; Krengel, U. Crystal structures of cholera toxin in complex with fucosylated receptors point to importance of secondary binding site. Sci. Rep. 2019, 9, 12243. [Google Scholar] [CrossRef] [Green Version]
- Sixma, T.K.; Stein, P.E.; Hol, W.G.; Read, R.J. Comparison of the B-pentamers of heat-labile enterotoxin and verotoxin-1: Two structures with remarkable similarity and dissimilarity. Biochemistry 1993, 32, 191–198. [Google Scholar] [CrossRef]
- Karmali, M.A. Infection by verocytotoxin-producing Escherichia coli. Clin. Microbiol. Rev. 1989, 2, 15–38. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [Green Version]
- Menge, C. Molecular Biology of Escherichia coli Shiga Toxins’ Effects on Mammalian Cells. Toxins 2020, 12, 345. [Google Scholar] [CrossRef]
- Gannon, V.P.; Gyles, C.L. Characteristics of the Shiga-like toxin produced by Escherichia coli associated with porcine edema disease. Vet. Microbiol. 1990, 24, 89–100. [Google Scholar] [CrossRef]
- Schmidt, H. Shiga-toxin-converting bacteriophages. Res. Microbiol. 2001, 152, 687–695. [Google Scholar] [CrossRef]
- Herold, S.; Karch, H.; Schmidt, H. Shiga toxin-encoding bacteriophages—Genomes in motion. Int. J. Med. Microbiol. 2004, 294, 115–121. [Google Scholar] [CrossRef]
- O’Brien, A.D.; Holmes, R.K. Shiga and Shiga-like toxins. Microbiol. Rev. 1987, 51, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, M.; Yamamoto, M.; Tomiyama-Miyaji, C.; Yuki, Y.; Takeda, Y.; Kiyono, H. Nontoxic Shiga toxin derivatives from Escherichia coli possess adjuvant activity for the augmentation of antigen-specific immune responses via dendritic cell activation. Infect. Immun. 2005, 73, 4088–4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingwood, C.A. Role of verotoxin receptors in pathogenesis. Trends Microbiol. 1996, 4, 147–153. [Google Scholar] [CrossRef]
- Moxley, R.A. Escherichia coli 0157:H7: An update on intestinal colonization and virulence mechanisms. Anim. Health Res. Rev. 2004, 5, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Muhlen, S.; Dersch, P. Treatment Strategies for Infections With Shiga Toxin-Producing Escherichia coli. Front. Cell. Infect. Microbiol. 2020, 10, 169. [Google Scholar] [CrossRef]
- Geue, L.; Segura-Alvarez, M.; Conraths, F.J.; Kuczius, T.; Bockemuhl, J.; Karch, H.; Gallien, P. A long-term study on the prevalence of shiga toxin-producing Escherichia coli (STEC) on four German cattle farms. Epidemiol. Infect. 2002, 129, 173–185. [Google Scholar] [CrossRef]
- Besser, T.E.; Richards, B.L.; Rice, D.H.; Hancock, D.D. Escherichia coli O157:H7 infection of calves: Infectious dose and direct contact transmission. Epidemiol. Infect. 2001, 127, 555–560. [Google Scholar] [CrossRef]
- Grauke, L.J.; Kudva, I.T.; Yoon, J.W.; Hunt, C.W.; Williams, C.J.; Hovde, C.J. Gastrointestinal tract location of Escherichia coli O157:H7 in ruminants. Appl. Environ. Microbiol. 2002, 68, 2269–2277. [Google Scholar] [CrossRef] [Green Version]
- Girard, F.; Dziva, F.; van Diemen, P.; Phillips, A.D.; Stevens, M.P.; Frankel, G. Adherence of enterohemorrhagic Escherichia coli O157, O26, and O111 strains to bovine intestinal explants ex vivo. Appl. Environ. Microbiol. 2007, 73, 3084–3090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naylor, S.W.; Low, J.C.; Besser, T.E.; Mahajan, A.; Gunn, G.J.; Pearce, M.C.; McKendrick, I.J.; Smith, D.G.; Gally, D.L. Lymphoid follicle-dense mucosa at the terminal rectum is the principal site of colonization of enterohemorrhagic Escherichia coli O157:H7 in the bovine host. Infect. Immun. 2003, 71, 1505–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohlenz, J.F.; Dean-Nystrom, E.A. Colonisation of Escherichia coli O157:H7 on squamous epithelial cells at the rectal-anal junction. Vet. Rec. 2004, 155, 248. [Google Scholar] [PubMed]
- Stoffregen, W.C.; Pohlenz, J.F.; Dean-Nystrom, E.A. Escherichia coli O157:H7 in the gallbladders of experimentally infected calves. J. Vet. Diagn. Investig. 2004, 16, 79–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cray, W.C., Jr.; Moon, H.W. Experimental infection of calves and adult cattle with Escherichia coli O157:H7. Appl. Environ. Microbiol. 1995, 61, 1586–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean-Nystrom, E.A.; Stoffregen, W.C.; Bosworth, B.T.; Moon, H.W.; Pohlenz, J.F. Early attachment sites for Shiga-toxigenic Escherichia coli O157:H7 in experimentally inoculated weaned calves. Appl. Environ. Microbiol. 2008, 74, 6378–6384. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, M.E.; Aspan, A.; Eriksson, E.; Vagsholm, I. Persistence of verocytotoxin-producing Escherichia coli O157:H7 in calves kept on pasture and in calves kept indoors during the summer months in a Swedish dairy herd. Int. J. Food Microbiol. 2001, 66, 55–61. [Google Scholar] [CrossRef]
- Liebana, E.; Smith, R.P.; Batchelor, M.; McLaren, I.; Cassar, C.; Clifton-Hadley, F.A.; Paiba, G.A. Persistence of Escherichia coli O157 isolates on bovine farms in England and Wales. J. Clin. Microbiol. 2005, 43, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Pirro, F.; Wieler, L.H.; Failing, K.; Bauerfeind, R.; Baljer, G. Neutralizing antibodies against Shiga-like toxins from Escherichia coli in colostra and sera of cattle. Vet. Microbiol. 1995, 43, 131–141. [Google Scholar] [CrossRef]
- Hoffman, M.A.; Menge, C.; Casey, T.A.; Laegreid, W.; Bosworth, B.T.; Dean-Nystrom, E.A. Bovine immune response to shiga-toxigenic Escherichia coli O157:H7. Clin. Vaccine Immunol. 2006, 13, 1322–1327. [Google Scholar] [CrossRef] [Green Version]
- Magnuson, B.A.; Davis, M.; Hubele, S.; Austin, P.R.; Kudva, I.T.; Williams, C.J.; Hunt, C.W.; Hovde, C.J. Ruminant gastrointestinal cell proliferation and clearance of Escherichia coli O157:H7. Infect. Immun. 2000, 68, 3808–3814. [Google Scholar] [CrossRef] [Green Version]
- Dean-Nystrom, E.A.; Bosworth, B.T.; Moon, H.W.; O’Brien, A.D. Escherichia coli O157:H7 requires intimin for enteropathogenicity in calves. Infect. Immun. 1998, 66, 4560–4563. [Google Scholar] [CrossRef] [PubMed]
- Cornick, N.A.; Booher, S.L.; Moon, H.W. Intimin facilitates colonization by Escherichia coli O157:H7 in adult ruminants. Infect. Immun. 2002, 70, 2704–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziva, F.; van Diemen, P.M.; Stevens, M.P.; Smith, A.J.; Wallis, T.S. Identification of Escherichia coli O157: H7 genes influencing colonization of the bovine gastrointestinal tract using signature-tagged mutagenesis. Microbiology 2004, 150, 3631–3645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Diemen, P.M.; Dziva, F.; Stevens, M.P.; Wallis, T.S. Identification of enterohemorrhagic Escherichia coli O26:H- genes required for intestinal colonization in calves. Infect. Immun. 2005, 73, 1735–1743. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.P.; Marches, O.; Campbell, J.; Huter, V.; Frankel, G.; Phillips, A.D.; Oswald, E.; Wallis, T.S. Intimin, tir, and shiga toxin 1 do not influence enteropathogenic responses to shiga toxin-producing Escherichia coli in bovine ligated intestinal loops. Infect. Immun. 2002, 70, 945–952. [Google Scholar] [CrossRef] [Green Version]
- Shaw, D.J.; Jenkins, C.; Pearce, M.C.; Cheasty, T.; Gunn, G.J.; Dougan, G.; Smith, H.R.; Woolhouse, M.E.; Frankel, G. Shedding patterns of verocytotoxin-producing Escherichia coli strains in a cohort of calves and their dams on a Scottish beef farm. Appl. Environ. Microbiol. 2004, 70, 7456–7465. [Google Scholar] [CrossRef] [Green Version]
- Wieler, L.H.; Bauerfeind, R.; Baljer, G. Characterization of Shiga-like toxin producing Escherichia coli (SLTEC) isolated from calves with and without diarrhoea. Zentralbl. Bakteriol. 1992, 276, 243–253. [Google Scholar] [CrossRef]
- Chanter, N.; Hall, G.A.; Bland, A.P.; Hayle, A.J.; Parsons, K.R. Dysentery in calves caused by an atypical strain of Escherichia coli (S102-9). Vet. Microbiol. 1986, 12, 241–253. [Google Scholar] [CrossRef]
- Dean-Nystrom, E.A.; Bosworth, B.T.; Cray, W.C., Jr.; Moon, H.W. Pathogenicity of Escherichia coli O157:H7 in the intestines of neonatal calves. Infect. Immun. 1997, 65, 1842–1848. [Google Scholar] [CrossRef] [Green Version]
- Janke, B.H.; Francis, D.H.; Collins, J.E.; Libal, M.C.; Zeman, D.H.; Johnson, D.D.; Neiger, R.D. Attaching and effacing Escherichia coli infection as a cause of diarrhea in young calves. J. Am. Vet. Med. Assoc. 1990, 196, 897–901. [Google Scholar]
- Moxley, R.A.; Francis, D.H. Natural and experimental infection with an attaching and effacing strain of Escherichia coli in calves. Infect. Immun. 1986, 53, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.E.; Karmali, M.A.; Becker, L.E.; Smith, C.R. The histopathology of the hemolytic uremic syndrome associated with verocytotoxin-producing Escherichia coli infections. Hum. Pathol. 1988, 19, 1102–1108. [Google Scholar] [CrossRef]
- van de Kar, N.C.; Monnens, L.A.; Karmali, M.A.; van Hinsbergh, V.W. Tumor necrosis factor and interleukin-1 induce expression of the verocytotoxin receptor globotriaosylceramide on human endothelial cells: Implications for the pathogenesis of the hemolytic uremic syndrome. Blood 1992, 80, 2755–2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proulx, F.; Seidman, E.G.; Karpman, D. Pathogenesis of Shiga toxin-associated hemolytic uremic syndrome. Pediatr. Res. 2001, 50, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruimboom-Brees, I.M.; Morgan, T.W.; Ackermann, M.R.; Nystrom, E.D.; Samuel, J.E.; Cornick, N.A.; Moon, H.W. Cattle lack vascular receptors for Escherichia coli O157:H7 Shiga toxins. Proc. Natl. Acad. Sci. USA 2000, 97, 10325–10329. [Google Scholar] [CrossRef] [Green Version]
- Mangeney, M.; Lingwood, C.A.; Taga, S.; Caillou, B.; Tursz, T.; Wiels, J. Apoptosis induced in Burkitt’s lymphoma cells via Gb3/CD77, a glycolipid antigen. Cancer Res. 1993, 53, 5314–5319. [Google Scholar] [PubMed]
- Chark, D.; Nutikka, A.; Trusevych, N.; Kuzmina, J.; Lingwood, C. Differential carbohydrate epitope recognition of globotriaosyl ceramide by verotoxins and a monoclonal antibody. Eur. J. Biochem. 2004, 271, 405–417. [Google Scholar] [CrossRef]
- Schwarting, G.A. Quantitative analysis of neutral glycosphingolipids from human lymphocyte subpopulations. Biochem. J. 1980, 189, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Taga, S.; Tetaud, C.; Mangeney, M.; Tursz, T.; Wiels, J. Sequential changes in glycolipid expression during human B cell differentiation: Enzymatic bases. Biochim. Biophys. Acta 1995, 1254, 56–65. [Google Scholar] [CrossRef]
- Menge, C.; Stamm, I.; Wuhrer, M.; Geyer, R.; Wieler, L.H.; Baljer, G. Globotriaosylceramide (Gb(3)/CD77) is synthesized and surface expressed by bovine lymphocytes upon activation in vitro. Vet. Immunol. Immunopathol. 2001, 83, 19–36. [Google Scholar] [CrossRef]
- Menge, C.; Wieler, L.H.; Schlapp, T.; Baljer, G. Shiga toxin 1 from Escherichia coli blocks activation and proliferation of bovine lymphocyte subpopulations in vitro. Infect. Immun. 1999, 67, 2209–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, A. Immunomodulation by Shiga Toxin 2. Ph.D. Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2010. [Google Scholar]
- Mangeney, M.; Richard, Y.; Coulaud, D.; Tursz, T.; Wiels, J. CD77: An antigen of germinal center B cells entering apoptosis. Eur. J. Immunol 1991, 21, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Mangeney, M.; Rousselet, G.; Taga, S.; Tursz, T.; Wiels, J. The fate of human CD77+ germinal center B lymphocytes after rescue from apoptosis. Mol. Immunol. 1995, 32, 333–339. [Google Scholar] [CrossRef]
- Menge, C.; Stamm, I.; Blessenohl, M.; Wieler, L.H.; Baljer, G. Verotoxin 1 from Escherichia coli affects Gb3/CD77+ bovine lymphocytes independent of interleukin-2, tumor necrosis factor-alpha, and interferon-alpha. Exp. Biol. Med. (Maywood) 2003, 228, 377–386. [Google Scholar] [CrossRef]
- Menge, C.; Blessenohl, M.; Eisenberg, T.; Stamm, I.; Baljer, G. Bovine ileal intraepithelial lymphocytes represent target cells for Shiga toxin 1 from Escherichia coli. Infect. Immun. 2004, 72, 1896–1905. [Google Scholar] [CrossRef] [Green Version]
- Wiels, J.; Fellous, M.; Tursz, T. Monoclonal antibody against a Burkitt lymphoma-associated antigen. Proc. Natl. Acad. Sci. USA 1981, 78, 6485–6488. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Barthelemy, C.; de Bouteiller, O.; Banchereau, J. The differences in survival and phenotype between centroblasts and centrocytes. Adv. Exp. Med. Biol. 1994, 355, 213–218. [Google Scholar] [CrossRef]
- Cohen, A.; Madrid-Marina, V.; Estrov, Z.; Freedman, M.H.; Lingwood, C.A.; Dosch, H.M. Expression of glycolipid receptors to Shiga-like toxin on human B lymphocytes: A mechanism for the failure of long-lived antibody response to dysenteric disease. Int. Immunol. 1990, 2, 1–8. [Google Scholar] [CrossRef]
- Ramegowda, B.; Tesh, V.L. Differentiation-associated toxin receptor modulation, cytokine production, and sensitivity to Shiga-like toxins in human monocytes and monocytic cell lines. Infect. Immun. 1996, 64, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Kniep, B.; Monner, D.A.; Schwulera, U.; Muhlradt, P.F. Glycosphingolipids of the globo-series are associated with the monocytic lineage of human myeloid cells. Eur. J. Biochem. 1985, 149, 187–191. [Google Scholar] [CrossRef]
- Pudymaitis, A.; Lingwood, C.A. Susceptibility to verotoxin as a function of the cell cycle. J. Cell Physiol. 1992, 150, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Stamm, I.; Wuhrer, M.; Geyer, R.; Baljer, G.; Menge, C. Bovine lymphocytes express functional receptors for Escherichia coli Shiga toxin 1. Microb. Pathog. 2002, 33, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, A.A.; Brown, J.E.; Stromberg, N.; Westling-Ryd, M.; Schultz, J.E.; Karlsson, K.A. Identification of the carbohydrate receptor for Shiga toxin produced by Shigella dysenteriae type 1. J. Biol. Chem. 1987, 262, 1779–1785. [Google Scholar] [PubMed]
- Pellizzari, A.; Pang, H.; Lingwood, C.A. Binding of verocytotoxin 1 to its receptor is influenced by differences in receptor fatty acid content. Biochemistry 1992, 31, 1363–1370. [Google Scholar] [CrossRef]
- Christopher-Hennings, J.; Willgohs, J.A.; Francis, D.H.; Raman, U.A.; Moxley, R.A.; Hurley, D.J. Immunocompromise in gnotobiotic pigs induced by verotoxin-producing Escherichia coli (O111:NM). Infect. Immun. 1993, 61, 2304–2308. [Google Scholar] [CrossRef] [Green Version]
- Sugatani, J.; Igarashi, T.; Shimura, M.; Yamanaka, T.; Takeda, T.; Miwa, M. Disorders in the immune responses of T- and B-cells in mice administered intravenous verotoxin 2. Life Sci. 2000, 67, 1059–1072. [Google Scholar] [CrossRef]
- Eiklid, K.; Olsnes, S. Interaction of Shigella shigae cytotoxin with receptors on sensitive and insensitive cells. J. Recept. Res. 1980, 1, 199–213. [Google Scholar] [CrossRef]
- Ghislain, J.; Lingwood, C.A.; Fish, E.N. Evidence for glycosphingolipid modification of the type 1 IFN receptor. J. Immunol. 1994, 153, 3655–3663. [Google Scholar]
- Klapproth, J.M.; Donnenberg, M.S.; Abraham, J.M.; Mobley, H.L.; James, S.P. Products of enteropathogenic Escherichia coli inhibit lymphocyte activation and lymphokine production. Infect. Immun. 1995, 63, 2248–2254. [Google Scholar] [CrossRef] [Green Version]
- Klapproth, J.M.; Scaletsky, I.C.; McNamara, B.P.; Lai, L.C.; Malstrom, C.; James, S.P.; Donnenberg, M.S. A large toxin from pathogenic Escherichia coli strains that inhibits lymphocyte activation. Infect. Immun. 2000, 68, 2148–2155. [Google Scholar] [CrossRef] [Green Version]
- Kerner, K.; Bridger, P.S.; Kopf, G.; Frohlich, J.; Barth, S.; Willems, H.; Bauerfeind, R.; Baljer, G.; Menge, C. Evaluation of biological safety in vitro and immunogenicity in vivo of recombinant Escherichia coli Shiga toxoids as candidate vaccines in cattle. Vet. Res. 2015, 46, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obrig, T.G.; Moran, T.P.; Brown, J.E. The mode of action of Shiga toxin on peptide elongation of eukaryotic protein synthesis. Biochem. J. 1987, 244, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, L.M.; van Haaften, W.C.; Tesh, V.L. Regulation of proinflammatory cytokine expression by Shiga toxin 1 and/or lipopolysaccharides in the human monocytic cell line THP-1. Infect. Immun. 2004, 72, 2618–2627. [Google Scholar] [CrossRef] [Green Version]
- Taga, S.; Carlier, K.; Mishal, Z.; Capoulade, C.; Mangeney, M.; Lecluse, Y.; Coulaud, D.; Tetaud, C.; Pritchard, L.L.; Tursz, T.; et al. Intracellular signaling events in CD77-mediated apoptosis of Burkitt’s lymphoma cells. Blood 1997, 90, 2757–2767. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, Y.U.; Kiyokawa, N.; Fujimoto, J. The effect of shiga toxin binding to globotriaosylceramidein rafts of human kidney cells and Burkitt’slymphoma cells. Trends Glycosci. Glycotech. 2001, 13, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Matthews, K.R.; Murdough, P.A.; Bramley, A.J. Invasion of bovine epithelial cells by verocytotoxin-producing Escherichia coli O157:H7. J. Appl. Microbiol. 1997, 82, 197–203. [Google Scholar] [CrossRef]
- Schmidt, N.; Barth, S.A.; Frahm, J.; Meyer, U.; Danicke, S.; Geue, L.; Menge, C. Decreased STEC shedding by cattle following passive and active vaccination based on recombinant Escherichia coli Shiga toxoids. Vet. Res. 2018, 49, 28. [Google Scholar] [CrossRef] [Green Version]
- Waters, W.R.; Harp, J.A.; Nonnecke, B.J. Phenotypic analysis of peripheral blood lymphocytes and intestinal intra-epithelial lymphocytes in calves. Vet. Immunol. Immunopathol. 1995, 48, 249–259. [Google Scholar] [CrossRef]
- Fitzgerald, S.F.; Beckett, A.E.; Palarea-Albaladejo, J.; McAteer, S.; Shaaban, S.; Morgan, J.; Ahmad, N.I.; Young, R.; Mabbott, N.A.; Morrison, L.; et al. Shiga toxin sub-type 2a increases the efficiency of Escherichia coli O157 transmission between animals and restricts epithelial regeneration in bovine enteroids. PLoS Pathog. 2019, 15, e1008003. [Google Scholar] [CrossRef] [Green Version]
- Hoey, D.E.; Currie, C.; Else, R.W.; Nutikka, A.; Lingwood, C.A.; Gally, D.L.; Smith, D.G. Expression of receptors for verotoxin 1 from Escherichia coli O157 on bovine intestinal epithelium. J. Med. Microbiol. 2002, 51, 143–149. [Google Scholar] [CrossRef]
- Moussay, E.; Stamm, I.; Taubert, A.; Baljer, G.; Menge, C. Escherichia coli Shiga toxin 1 enhances il-4 transcripts in bovine ileal intraepithelial lymphocytes. Vet. Immunol. Immunopathol. 2006, 113, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Grogan, J.L.; Mohrs, M.; Harmon, B.; Lacy, D.A.; Sedat, J.W.; Locksley, R.M. Early transcription and silencing of cytokine genes underlie polarization of T helper cell subsets. Immunity 2001, 14, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Winzen, R.; Kracht, M.; Ritter, B.; Wilhelm, A.; Chen, C.Y.; Shyu, A.B.; Muller, M.; Gaestel, M.; Resch, K.; Holtmann, H. The p38 MAP kinase pathway signals for cytokine-induced mRNA stabilization via MAP kinase-activated protein kinase 2 and an AU-rich region-targeted mechanism. EMBO J. 1999, 18, 4969–4980. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, C.M.; Hurley, B.P.; Lincicome, L.L.; Jacewicz, M.S.; Keusch, G.T.; Acheson, D.W. Shiga toxins stimulate secretion of interleukin-8 from intestinal epithelial cells. Infect. Immun. 1999, 67, 5985–5993. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, C.M.; Smith, W.E.; Hurley, B.P.; Acheson, D.W. Shiga toxins induce, superinduce, and stabilize a variety of C-X-C chemokine mRNAs in intestinal epithelial cells, resulting in increased chemokine expression. Infect. Immun. 2001, 69, 6140–6147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, A.; Peltz, S.W. Interrelationships of the pathways of mRNA decay and translation in eukaryotic cells. Annu. Rev. Biochem. 1996, 65, 693–739. [Google Scholar] [CrossRef]
- Rook, G.A.; Hernandez-Pando, R.; Dheda, K.; Teng Seah, G. IL-4 in tuberculosis: Implications for vaccine design. Trends Immunol. 2004, 25, 483–488. [Google Scholar] [CrossRef]
- Bitzan, M.M.; Wang, Y.; Lin, J.; Marsden, P.A. Verotoxin and ricin have novel effects on preproendothelin-1 expression but fail to modify nitric oxide synthase (ecNOS) expression and NO production in vascular endothelium. J. Clin. Investig. 1998, 101, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Stein, G.M.; Pfuller, U.; Schietzel, M.; Bussing, A. Expression of interleukin-4 in apoptotic cells: Stimulation of the type-2 cytokine by different toxins in human peripheral blood mononuclear and tumor cells. Cytometry 2000, 41, 261–270. [Google Scholar] [CrossRef]
- Ebert, E.C.; Roberts, A.I. IL-4 down-regulates the responsiveness of human intraepithelial lymphocytes. Clin. Exp. Immunol. 1996, 105, 556–560. [Google Scholar] [CrossRef]
- Menge, C.; Stamm, I.; Van Diemen, P.M.; Sopp, P.; Baljer, G.; Wallis, T.S.; Stevens, M.P. Phenotypic and functional characterization of intraepithelial lymphocytes in a bovine ligated intestinal loop model of enterohaemorrhagic Escherichia coli infection. J. Med. Microbiol. 2004, 53, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Menge, C.; Loos, D.; Bridger, P.S.; Barth, S.; Werling, D.; Baljer, G. Bovine macrophages sense Escherichia coli Shiga toxin 1. Innate Immun. 2015, 21, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, P.; Paton, N.; Smith, D.G. Verotoxin-2 activates mitogen-activated protein kinases in bovine adherent peripheral blood mononuclear cells. J. Comp. Pathol. 2012, 147, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Cameron, P.; Smith, S.J.; Giembycz, M.A.; Rotondo, D.; Plevin, R. Verotoxin activates mitogen-activated protein kinase in human peripheral blood monocytes: Role in apoptosis and proinflammatory cytokine release. Br. J. Pharmacol. 2003, 140, 1320–1330. [Google Scholar] [CrossRef] [Green Version]
- Olsnes, S.; Reisbig, R.; Eiklid, K. Subunit structure of Shigella cytotoxin. J. Biol. Chem. 1981, 256, 8732–8738. [Google Scholar] [PubMed]
- Mosser, D.M. The many faces of macrophage activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Stamm, I.; Mohr, M.; Bridger, P.S.; Schropfer, E.; Konig, M.; Stoffregen, W.C.; Dean-Nystrom, E.A.; Baljer, G.; Menge, C. Epithelial and mesenchymal cells in the bovine colonic mucosa differ in their responsiveness to Escherichia coli Shiga toxin 1. Infect. Immun. 2008, 76, 5381–5391. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Akahoshi, T.; Sasahana, T.; Kitasato, H.; Namai, R.; Sasaki, T.; Inoue, M.; Kondo, H. Inhibition of neutrophil apoptosis by verotoxin 2 derived from Escherichia coli O157:H7. Infect. Immun. 1999, 67, 6203–6205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.J.; Sundaram, S.; Cendoroglo, M.; Acheson, D.W.; Keusch, G.T. Shiga toxin induces superoxide production in polymorphonuclear cells with subsequent impairment of phagocytosis and responsiveness to phorbol esters. J. Infect. Dis. 1999, 179, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, C.; Natori, Y.; Zeng, X.T.; Ohmura, M.; Yamasaki, S.; Takeda, Y. Induction of cytokines in a human colon epithelial cell line by Shiga toxin 1 (Stx1) and Stx2 but not by non-toxic mutant Stx1 which lacks N-glycosidase activity. FEBS Lett. 1999, 442, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Hurley, B.P.; Thorpe, C.M.; Acheson, D.W. Shiga toxin translocation across intestinal epithelial cells is enhanced by neutrophil transmigration. Infect. Immun. 2001, 69, 6148–6155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- te Loo, D.M.; Monnens, L.A.; van Der Velden, T.J.; Vermeer, M.A.; Preyers, F.; Demacker, P.N.; van Den Heuvel, L.P.; van Hinsbergh, V.W. Binding and transfer of verocytotoxin by polymorphonuclear leukocytes in hemolytic uremic syndrome. Blood 2000, 95, 3396–3402. [Google Scholar] [CrossRef] [PubMed]
- Svanborg, C.; Godaly, G.; Hedlund, M. Cytokine responses during mucosal infections: Role in disease pathogenesis and host defence. Curr. Opin. Microbiol. 1999, 2, 99–105. [Google Scholar] [CrossRef]
- Schoonderwoerd, M.; Clarke, R.C.; van Dreumel, A.A.; Rawluk, S.A. Colitis in calves: Natural and experimental infection with a verotoxin-producing strain of Escherichia coli O111:NM. Can. J. Vet. Res. 1988, 52, 484–487. [Google Scholar]
- Diez-Fraile, A.; Meyer, E.; Massart-Leen, A.M.; Burvenich, C. Effect of isoproterenol and dexamethasone on the lipopolysaccharide induced expression of CD11b on bovine neutrophils. Vet. Immunol. Immunopathol. 2000, 76, 151–156. [Google Scholar] [CrossRef]
- Menge, C.; Eisenberg, T.; Stamm, I.; Baljer, G. Comparison of binding and effects of Escherichia coli Shiga toxin 1 on bovine and ovine granulocytes. Vet. Immunol. Immunopathol. 2006, 113, 392–403. [Google Scholar] [CrossRef]
- Mobassaleh, M.; Donohue-Rolfe, A.; Jacewicz, M.; Grand, R.J.; Keusch, G.T. Pathogenesis of shigella diarrhea: Evidence for a developmentally regulated glycolipid receptor for shigella toxin involved in the fluid secretory response of rabbit small intestine. J. Infect. Dis. 1988, 157, 1023–1031. [Google Scholar] [CrossRef]
- Smits, E.; Burvenich, C.; Guidry, A.J.; Heyneman, R.; Massart-Leen, A. Diapedesis across mammary epithelium reduces phagocytic and oxidative burst of bovine neutrophils. Vet. Immunol. Immunopathol. 1999, 68, 169–176. [Google Scholar] [CrossRef]
- Mohr, M. Nachweis und Reaktivität epithelialer und mesenchymaler Zielzellen für Escherichia coli Shigatoxin in den Kolonkrypten des Rindes; [German]; Dissertation Justus-Liebig-Universität Giessen: Giessen, Germany, 2007; Available online: http://geb.uni-giessen.de/geb/volltexte/2007/4685/pdf/MohrMelanie-2007-04-27.pdf (accessed on 20 September 2020).
- Hoey, D.E.; Sharp, L.; Currie, C.; Lingwood, C.A.; Gally, D.L.; Smith, D.G. Verotoxin 1 binding to intestinal crypt epithelial cells results in localization to lysosomes and abrogation of toxicity. Cell. Microbiol. 2003, 5, 85–97. [Google Scholar] [CrossRef]
- Schuller, S.; Frankel, G.; Phillips, A.D. Interaction of Shiga toxin from Escherichia coli with human intestinal epithelial cell lines and explants: Stx2 induces epithelial damage in organ culture. Cell. Microbiol. 2004, 6, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, C.A. Glycolipid receptors for verotoxin and Helicobacter pylori: Role in pathology. Biochim. Biophys. Acta 1999, 1455, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Falguieres, T.; Mallard, F.; Baron, C.; Hanau, D.; Lingwood, C.; Goud, B.; Salamero, J.; Johannes, L. Targeting of Shiga toxin B-subunit to retrograde transport route in association with detergent-resistant membranes. Mol. Biol. Cell 2001, 12, 2453–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesh, V.L.; Ramegowda, B.; Samuel, J.E. Purified Shiga-like toxins induce expression of proinflammatory cytokines from murine peritoneal macrophages. Infect. Immun. 1994, 62, 5085–5094. [Google Scholar] [CrossRef] [Green Version]
- van Setten, P.A.; Monnens, L.A.; Verstraten, R.G.; van den Heuvel, L.P.; van Hinsbergh, V.W. Effects of verocytotoxin-1 on nonadherent human monocytes: Binding characteristics, protein synthesis, and induction of cytokine release. Blood 1996, 88, 174–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlisidou, I.; Lyte, M.; van Diemen, P.M.; Hawes, P.; Monaghan, P.; Wallis, T.S.; Stevens, M.P. The neuroendocrine stress hormone norepinephrine augments Escherichia coli O157:H7-induced enteritis and adherence in a bovine ligated ileal loop model of infection. Infect. Immun. 2004, 72, 5446–5451. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.P.; Roe, A.J.; Vlisidou, I.; van Diemen, P.M.; La Ragione, R.M.; Best, A.; Woodward, M.J.; Gally, D.L.; Wallis, T.S. Mutation of toxB and a truncated version of the efa-1 gene in Escherichia coli O157:H7 influences the expression and secretion of locus of enterocyte effacement-encoded proteins but not intestinal colonization in calves or sheep. Infect. Immun. 2004, 72, 5402–5411. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.P.; van Diemen, P.M.; Frankel, G.; Phillips, A.D.; Wallis, T.S. Efa1 influences colonization of the bovine intestine by shiga toxin-producing Escherichia coli serotypes O5 and O111. Infect. Immun. 2002, 70, 5158–5166. [Google Scholar] [CrossRef] [Green Version]
- Bridger, P.S.; Mohr, M.; Stamm, I.; Frohlich, J.; Follmann, W.; Birkner, S.; Metcalfe, H.; Werling, D.; Baljer, G.; Menge, C. Primary bovine colonic cells: A model to study strain-specific responses to Escherichia coli. Vet. Immunol. Immunopathol. 2010, 137, 54–63. [Google Scholar] [CrossRef]
- Hauf, N.; Chakraborty, T. Suppression of NF-kappa B activation and proinflammatory cytokine expression by Shiga toxin-producing Escherichia coli. J. Immunol. 2003, 170, 2074–2082. [Google Scholar] [CrossRef] [Green Version]
- Stalb, S.; Barth, S.A.; Sobotta, K.; Liebler-Tenorio, E.; Geue, L.; Menge, C. Pro-inflammatory capacity of Escherichia coli O104:H4 outbreak strain during colonization of intestinal epithelial cells from human and cattle. Int. J. Med. Microbiol. 2018, 308, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Barth, S.A.; Menge, C.; Eichhorn, I.; Semmler, T.; Wieler, L.H.; Pickard, D.; Belka, A.; Berens, C.; Geue, L. The Accessory Genome of Shiga Toxin-Producing Escherichia coli Defines a Persistent Colonization Type in Cattle. Appl. Environ. Microbiol. 2016, 82, 5455–5464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, S.; Karve, S.S.; Weiss, A.A.; Hawkins, J.; Poling, H.M.; Helmrath, M.A.; Wells, J.M.; McCauley, H.A. Tissue Responses to Shiga Toxin in Human Intestinal Organoids. Cell Mol. Gastroenterol. Hepatol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferens, W.A.; Hovde, C.J. Antiviral activity of shiga toxin 1: Suppression of bovine leukemia virus-related spontaneous lymphocyte proliferation. Infect. Immun. 2000, 68, 4462–4469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T.A.; Gajewski, T.F.; Danielpour, D.; Chang, E.B.; Beagley, K.W.; Bluestone, J.A. Differential function of intestinal intraepithelial lymphocyte subsets. J. Immunol. 1992, 149, 1124–1130. [Google Scholar] [PubMed]
- Suzuki, R.; Nakao, A.; Kanamaru, Y.; Okumura, K.; Ogawa, H.; Ra, C. Localization of intestinal intraepithelial T lymphocytes involves regulation of alphaEbeta7 expression by transforming growth factor-beta. Int. Immunol. 2002, 14, 339–345. [Google Scholar] [CrossRef]
- Corbishley, A.; Ahmad, N.I.; Hughes, K.; Hutchings, M.R.; McAteer, S.P.; Connelley, T.K.; Brown, H.; Gally, D.L.; McNeilly, T.N. Strain-dependent cellular immune responses in cattle following Escherichia coli O157:H7 colonization. Infect. Immun. 2014, 82, 5117–5131. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Zhang, H.; Wang, H.; Hu, J.; Du, M.; Zhu, M.J. Host inflammatory response inhibits Escherichia coli O157:H7 adhesion to gut epithelium through augmentation of mucin expression. Infect. Immun. 2014, 82, 1921–1930. [Google Scholar] [CrossRef] [Green Version]
- Nart, P.; Naylor, S.W.; Huntley, J.F.; McKendrick, I.J.; Gally, D.L.; Low, J.C. Responses of cattle to gastrointestinal colonization by Escherichia coli O157:H7. Infect. Immun. 2008, 76, 5366–5372. [Google Scholar] [CrossRef] [Green Version]
- Kieckens, E.; Rybarczyk, J.; Li, R.W.; Vanrompay, D.; Cox, E. Potential immunosuppressive effects of Escherichia coli O157:H7 experimental infection on the bovine host. BMC Genom. 2016, 17, 1049. [Google Scholar] [CrossRef] [Green Version]
- Wang, O.; McAllister, T.A.; Plastow, G.; Stanford, K.; Selinger, B.; Guan, L.L. Host mechanisms involved in cattle Escherichia coli O157 shedding: A fundamental understanding for reducing foodborne pathogen in food animal production. Sci. Rep. 2017, 7, 7630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, O.; Liang, G.; McAllister, T.A.; Plastow, G.; Stanford, K.; Selinger, B.; Guan le, L. Comparative Transcriptomic Analysis of Rectal Tissue from Beef Steers Revealed Reduced Host Immunity in Escherichia coli O157:H7 Super-Shedders. PLoS ONE 2016, 11, e0151284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieler, L.H.; Franke, S.; Menge, C.; Rose, M.; Bauerfeind, R.; Karch, H.; Baljer, G. The immune response in edema disease of weaned piglets measured with a recombinant B subunit of shiga-like toxin II. Dtsch. Tierarztl. Wochenschr. 1995, 102, 40–43. [Google Scholar] [PubMed]
- Reymond, D.; Johnson, R.P.; Karmali, M.A.; Petric, M.; Winkler, M.; Johnson, S.; Rahn, K.; Renwick, S.; Wilson, J.; Clarke, R.C.; et al. Neutralizing antibodies to Escherichia coli Vero cytotoxin 1 and antibodies to O157 lipopolysaccharide in healthy farm family members and urban residents. J. Clin. Microbiol. 1996, 34, 2053–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretschneider, G.; Berberov, E.M.; Moxley, R.A. Isotype-specific antibody responses against Escherichia coli O157:H7 locus of enterocyte effacement proteins in adult beef cattle following experimental infection. Vet. Immunol. Immunopathol. 2007, 118, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.P.; Cray, W.C., Jr.; Johnson, S.T. Serum antibody responses of cattle following experimental infection with Escherichia coli O157:H7. Infect. Immun. 1996, 64, 1879–1883. [Google Scholar] [CrossRef] [Green Version]
- Frohlich, J.; Baljer, G.; Menge, C. Maternally and naturally acquired antibodies to Shiga toxins in a cohort of calves shedding Shiga-toxigenic Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 3695–3704. [Google Scholar] [CrossRef] [Green Version]
- Kuribayashi, T.; Seita, T.; Matsumoto, M.; Furuhata, K.; Tagata, K.; Yamamoto, S. Bovine colostral antibody against verotoxin 2 derived from Escherichia coli O157:H7: Resistance to proteases and effects in beagle dogs. Comp. Med. 2009, 59, 163–167. [Google Scholar]
- Tetsurou, S.; Kuribayashi, T.; Fukuyama, M.; Yamaguchi, S.; Yamamoto, S. Inhibition of verotoxin (VT) 2 absorption into systemic blood from intestine by repeated administration of bovine immune colostral antibody against VT2 in mice. J. Microbiol. Immunol. Infect. 2015, 48, 583–587. [Google Scholar] [CrossRef]
- Albanese, A.; Sacerdoti, F.; Seyahian, E.A.; Amaral, M.M.; Fiorentino, G.; Fernandez Brando, R.; Vilte, D.A.; Mercado, E.C.; Palermo, M.S.; Cataldi, A.; et al. Immunization of pregnant cows with Shiga toxin-2 induces high levels of specific colostral antibodies and lactoferrin able to neutralize E. coli O157:H7 pathogenicity. Vaccine 2018, 36, 1728–1735. [Google Scholar] [CrossRef]
- Corbishley, A.; Connelley, T.K.; Wolfson, E.B.; Ballingall, K.; Beckett, A.E.; Gally, D.L.; McNeilly, T.N. Identification of epitopes recognised by mucosal CD4(+) T-cell populations from cattle experimentally colonised with Escherichia coli O157:H7. Vet. Res. 2016, 47, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretschneider, G.; Berberov, E.M.; Moxley, R.A. Enteric mucosal antibodies to Escherichia coli O157:H7 in adult cattle. Vet. Rec. 2008, 163, 218–219. [Google Scholar] [CrossRef] [PubMed]
- Beckett, A.E. Defining the impact of colonisation with Shiga toxin positive E. coli O157 on adaptive immunity in cattle. Ph.D. Thesis, The University of Edinburgh, Edinburgh, UK, 2018. [Google Scholar]
- Smith, D.G.; Naylor, S.W.; Gally, D.L. Consequences of EHEC colonisation in humans and cattle. Int. J. Med. Microbiol. 2002, 292, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, C. Intestinal inflammation: A complex interplay of immune and nonimmune cell interactions. Am. J. Physiol. 1997, 273, G769–G775. [Google Scholar] [CrossRef]
- Wieler, L.H.; Schwanitz, A.; Vieler, E.; Busse, B.; Steinruck, H.; Kaper, J.B.; Baljer, G. Virulence properties of Shiga toxin-producing Escherichia coli (STEC) strains of serogroup O118, a major group of STEC pathogens in calves. J. Clin. Microbiol. 1998, 36, 1604–1607. [Google Scholar] [CrossRef] [Green Version]
- Simmons, C.P.; Clare, S.; Dougan, G. Understanding mucosal responsiveness: Lessons from enteric bacterial pathogens. Semin. Immunol. 2001, 13, 201–209. [Google Scholar] [CrossRef]
- Borman-Eby, H.C.; McEwen, S.A.; Clarke, R.C.; McNab, W.B.; Rahn, K.; Valdivieso-Garcia, A. The seroprevalence of verocytotoxin-producing Escherichia coli in Ontario dairy cows and associations with production and management. Prev. Vet. Med. 1993, 15, 261–274. [Google Scholar] [CrossRef]
- Eisenberg, T. Untersuchungen zur Wirkung von Shigatoxin 1 von Escherichia coli auf Zellen der unspezifischen Immunabwehr bei Rind, Schaf und Ziege; [German]; Dissertation Justus-Liebig-Universität Giessen: Giessen, Germany, 2003; Available online: http://geb.uni-giessen.de/geb/volltexte/2003/1131/pdf/EisenbergTobias-2003-06-05.pdf (accessed on 20 September 2020).
- Pohlenz, J.F.; Winter, K.R.; Dean-Nystrom, E.A. Shiga-toxigenic Escherichia coli-inoculated neonatal piglets develop kidney lesions that are comparable to those in humans with hemolytic-uremic syndrome. Infect. Immun. 2005, 73, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Winter, K.R.; Stoffregen, W.C.; Dean-Nystrom, E.A. Shiga toxin binding to isolated porcine tissues and peripheral blood leukocytes. Infect. Immun. 2004, 72, 6680–6684. [Google Scholar] [CrossRef] [Green Version]
- Inobe, J.; Slavin, A.J.; Komagata, Y.; Chen, Y.; Liu, L.; Weiner, H.L. IL-4 is a differentiation factor for transforming growth factor-beta secreting Th3 cells and oral administration of IL-4 enhances oral tolerance in experimental allergic encephalomyelitis. Eur. J. Immunol. 1998, 28, 2780–2790. [Google Scholar] [CrossRef]
- Mennechet, F.J.; Kasper, L.H.; Rachinel, N.; Minns, L.A.; Luangsay, S.; Vandewalle, A.; Buzoni-Gatel, D. Intestinal intraepithelial lymphocytes prevent pathogen-driven inflammation and regulate the Smad/T-bet pathway of lamina propria CD4+ T cells. Eur. J. Immunol. 2004, 34, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Naylor, S.W.; Roe, A.J.; Nart, P.; Spears, K.; Smith, D.G.; Low, J.C.; Gally, D.L. Escherichia coli O157: H7 forms attaching and effacing lesions at the terminal rectum of cattle and colonization requires the LEE4 operon. Microbiology 2005, 151, 2773–2781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamm, K.; Barth, S.A.; Stalb, S.; Geue, L.; Liebler-Tenorio, E.; Teifke, J.P.; Lange, E.; Tauscher, K.; Kotterba, G.; Bielaszewska, M.; et al. Experimental Infection of Calves with Escherichia coli O104:H4 outbreak strain. Sci. Rep. 2016, 6, 32812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.P.; van Diemen, P.M.; Dziva, F.; Jones, P.W.; Wallis, T.S. Options for the control of enterohaemorrhagic Escherichia coli in ruminants. Microbiology 2002, 148, 3767–3778. [Google Scholar] [CrossRef] [Green Version]
- Asper, D.J.; Karmali, M.A.; Townsend, H.; Rogan, D.; Potter, A.A. Serological response of Shiga toxin-producing Escherichia coli type III secreted proteins in sera from vaccinated rabbits, naturally infected cattle, and humans. Clin. Vaccine Immunol. 2011, 18, 1052–1057. [Google Scholar] [CrossRef] [Green Version]
- Potter, A.A.; Klashinsky, S.; Li, Y.; Frey, E.; Townsend, H.; Rogan, D.; Erickson, G.; Hinkley, S.; Klopfenstein, T.; Moxley, R.A.; et al. Decreased shedding of Escherichia coli O157:H7 by cattle following vaccination with type III secreted proteins. Vaccine 2004, 22, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.J.; Rogan, D.; Finlay, B.B.; Potter, A.A.; Asper, D.J. Vaccination with type III secreted proteins leads to decreased shedding in calves after experimental infection with Escherichia coli O157. Can. J. Vet. Res. 2011, 75, 98–105. [Google Scholar]
- Sheng, H.; Lim, J.Y.; Knecht, H.J.; Li, J.; Hovde, C.J. Role of Escherichia coli O157:H7 virulence factors in colonization at the bovine terminal rectal mucosa. Infect. Immun. 2006, 74, 4685–4693. [Google Scholar] [CrossRef] [Green Version]
- Cornick, N.A.; Booher, S.L.; Casey, T.A.; Moon, H.W. Persistent colonization of sheep by Escherichia coli O157:H7 and other E. coli pathotypes. Appl. Environ. Microbiol. 2000, 66, 4926–4934. [Google Scholar] [CrossRef] [Green Version]
- Cornick, N.A.; Helgerson, A.F.; Sharma, V. Shiga toxin and Shiga toxin-encoding phage do not facilitate Escherichia coli O157:H7 colonization in sheep. Appl. Environ. Microbiol. 2007, 73, 344–346. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Y.R.; Jokinen, C.C.; Laing, C.R.; Johnson, R.P.; Ziebell, K.; Gannon, V.P.J. Assessing the genomic relatedness and evolutionary rates of persistent verotoxigenic Escherichia coli serotypes within a closed beef herd in Canada. Microb. Genom. 2020, 6. [Google Scholar] [CrossRef]
- Cabal, A.; Geue, L.; Gomez-Barrero, S.; Barth, S.; Barcena, C.; Hamm, K.; Porrero, M.C.; Valverde, A.; Canton, R.; Menge, C.; et al. Detection of virulence-associated genes characteristic of intestinal Escherichia coli pathotypes, including the enterohemorrhagic/enteroaggregative O104:H4, in bovines from Germany and Spain. Microbiol. Immunol. 2015, 59, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; McAteer, S.P.; Paxton, E.; Mahajan, A.; Gally, D.L.; Tree, J.J. Screening of an E. coli O157:H7 Bacterial Artificial Chromosome Library by Comparative Genomic Hybridization to Identify Genomic Regions Contributing to Growth in Bovine Gastrointestinal Mucus and Epithelial Cell Colonization. Front. Microbiol. 2011, 2, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, Y.; Ooka, T.; Iguchi, A.; Toh, H.; Asadulghani, M.; Oshima, K.; Kodama, T.; Abe, H.; Nakayama, K.; Kurokawa, K.; et al. Comparative genomics reveal the mechanism of the parallel evolution of O157 and non-O157 enterohemorrhagic Escherichia coli. Proc. Natl. Acad. Sci. USA 2009, 106, 17939–17944. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Makino, K.; Ohnishi, M.; Kurokawa, K.; Ishii, K.; Yokoyama, K.; Han, C.G.; Ohtsubo, E.; Nakayama, K.; Murata, T.; et al. Complete genome sequence of enterohemorrhagic Escherichia coli O157:H7 and genomic comparison with a laboratory strain K-12. DNA Res. 2001, 8, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, N.T.; Plunkett, G., 3rd; Burland, V.; Mau, B.; Glasner, J.D.; Rose, D.J.; Mayhew, G.F.; Evans, P.S.; Gregor, J.; Kirkpatrick, H.A.; et al. Genome sequence of enterohaemorrhagic Escherichia coli O157:H7. Nature 2001, 409, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Martorelli, L.; Hovde, C.J.; Vilte, D.A.; Albanese, A.; Zotta, E.; Ibarra, C.; Cantet, R.J.; Mercado, E.C.; Cataldi, A. Impact of Infection Dose and Previous Serum Antibodies against the Locus of Enterocyte Effacement Proteins on Escherichia coli O157:H7 Shedding in Calves following Experimental Infection. Biomed. Res. Int. 2015, 2015, 290679. [Google Scholar] [CrossRef] [Green Version]
- Vilte, D.A.; Larzabal, M.; Garbaccio, S.; Gammella, M.; Rabinovitz, B.C.; Elizondo, A.M.; Cantet, R.J.; Delgado, F.; Meikle, V.; Cataldi, A.; et al. Reduced faecal shedding of Escherichia coli O157:H7 in cattle following systemic vaccination with gamma-intimin C(2)(8)(0) and EspB proteins. Vaccine 2011, 29, 3962–3968. [Google Scholar] [CrossRef]
- McNeilly, T.N.; Mitchell, M.C.; Corbishley, A.; Nath, M.; Simmonds, H.; McAteer, S.P.; Mahajan, A.; Low, J.C.; Smith, D.G.; Huntley, J.F.; et al. Optimizing the Protection of Cattle against Escherichia coli O157:H7 Colonization through Immunization with Different Combinations of H7 Flagellin, Tir, Intimin-531 or EspA. PLoS ONE 2015, 10, e0128391. [Google Scholar] [CrossRef]
- Klapproth, J.M.; Donnenberg, M.S.; Abraham, J.M.; James, S.P. Products of enteropathogenic E. coli inhibit lymphokine production by gastrointestinal lymphocytes. Am. J. Physiol. 1996, 271, G841–G848. [Google Scholar] [CrossRef]
- Malstrom, C.; James, S. Inhibition of murine splenic and mucosal lymphocyte function by enteric bacterial products. Infect. Immun. 1998, 66, 3120–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deacon, V.; Dziva, F.; van Diemen, P.M.; Frankel, G.; Stevens, M.P. Efa-1/LifA mediates intestinal colonization of calves by enterohaemorrhagic Escherichia coli O26: H- in a manner independent of glycosyltransferase and cysteine protease motifs or effects on type III secretion. Microbiology 2010, 156, 2527–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassady-Cain, R.L.; Hope, J.C.; Stevens, M.P. Direct Manipulation of T Lymphocytes by Proteins of Gastrointestinal Bacterial Pathogens. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassady-Cain, R.L.; Blackburn, E.A.; Bell, C.R.; Elshina, E.; Hope, J.C.; Stevens, M.P. Inhibition of Antigen-Specific and Nonspecific Stimulation of Bovine T and B Cells by Lymphostatin from Attaching and Effacing Escherichia coli. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Cassady-Cain, R.L.; Blackburn, E.A.; Alsarraf, H.; Dedic, E.; Bease, A.G.; Bottcher, B.; Jorgensen, R.; Wear, M.; Stevens, M.P. Biophysical Characterization and Activity of Lymphostatin, a Multifunctional Virulence Factor of Attaching and Effacing Escherichia coli. J. Biol. Chem. 2016, 291, 5803–5816. [Google Scholar] [CrossRef] [Green Version]
- Higgins, L.M.; Frankel, G.; Douce, G.; Dougan, G.; MacDonald, T.T. Citrobacter rodentium infection in mice elicits a mucosal Th1 cytokine response and lesions similar to those in murine inflammatory bowel disease. Infect. Immun. 1999, 67, 3031–3039. [Google Scholar] [CrossRef] [Green Version]
- Bohle, S.M. Investigations concerning the immunomodulatory effect of the virulence factor “intimin” of enteropathogenic and enterohemorrhagic Escherichia coli in cattle; [German]; Dissertation Justus-Liebig-Universität Giessen: Giessen, Germany, 2006; Available online: http://geb.uni-giessen.de/geb/volltexte/2006/2874/pdf/BohleStefanie-2006-05-24.pdf (accessed on 20 September 2020).
- Rashid, R.A.; Tabata, T.A.; Oatley, M.J.; Besser, T.E.; Tarr, P.I.; Moseley, S.L. Expression of putative virulence factors of Escherichia coli O157:H7 differs in bovine and human infections. Infect. Immun. 2006, 74, 4142–4148. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, J.M.; Wagner, P.L.; Acheson, D.W.; Waldor, M.K. Comparison of Shiga toxin production by hemolytic-uremic syndrome-associated and bovine-associated Shiga toxin-producing Escherichia coli isolates. Appl. Environ. Microbiol. 2003, 69, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Lejeune, J.T.; Abedon, S.T.; Takemura, K.; Christie, N.P.; Sreevatsan, S. Human Escherichia coli O157:H7 genetic marker in isolates of bovine origin. Emerg Infect. Dis. 2004, 10, 1482–1485. [Google Scholar] [CrossRef]
- Lupolova, N.; Dallman, T.J.; Matthews, L.; Bono, J.L.; Gally, D.L. Support vector machine applied to predict the zoonotic potential of E. coli O157 cattle isolates. Proc. Natl. Acad. Sci. USA 2016, 113, 11312–11317. [Google Scholar] [CrossRef] [Green Version]
- Chase-Topping, M.; Gally, D.; Low, C.; Matthews, L.; Woolhouse, M. Super-shedding and the link between human infection and livestock carriage of Escherichia coli O157. Nat Rev Microbiol 2008, 6, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; McAteer, S.P.; Tree, J.J.; Shaw, D.J.; Wolfson, E.B.; Beatson, S.A.; Roe, A.J.; Allison, L.J.; Chase-Topping, M.E.; Mahajan, A.; et al. Lysogeny with Shiga toxin 2-encoding bacteriophages represses type III secretion in enterohemorrhagic Escherichia coli. PLoS Pathog. 2012, 8, e1002672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furniss, R.C.D.; Clements, A. Regulation of the Locus of Enterocyte Effacement in Attaching and Effacing Pathogens. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.T.; Clarke, M.B.; Yamamoto, K.; Rasko, D.A.; Sperandio, V. The QseC adrenergic signaling cascade in Enterohemorrhagic E. coli (EHEC). PLoS Pathog. 2009, 5, e1000553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, C.T.; Russell, R.; Njoroge, J.W.; Jimenez, A.G.; Taussig, R.; Sperandio, V. Genetic and Mechanistic Analyses of the Periplasmic Domain of the Enterohemorrhagic Escherichia coli QseC Histidine Sensor Kinase. J. Bacteriol. 2017, 199. [Google Scholar] [CrossRef] [Green Version]
- Garmendia, J.; Phillips, A.D.; Carlier, M.F.; Chong, Y.; Schuller, S.; Marches, O.; Dahan, S.; Oswald, E.; Shaw, R.K.; Knutton, S.; et al. TccP is an enterohaemorrhagic Escherichia coli O157:H7 type III effector protein that couples Tir to the actin-cytoskeleton. Cell. Microbiol. 2004, 6, 1167–1183. [Google Scholar] [CrossRef]
- Pacheco, A.R.; Curtis, M.M.; Ritchie, J.M.; Munera, D.; Waldor, M.K.; Moreira, C.G.; Sperandio, V. Fucose sensing regulates bacterial intestinal colonization. Nature 2012, 492, 113–117. [Google Scholar] [CrossRef]
- Berger, P.; Kouzel, I.U.; Berger, M.; Haarmann, N.; Dobrindt, U.; Koudelka, G.B.; Mellmann, A. Carriage of Shiga toxin phage profoundly affects Escherichia coli gene expression and carbon source utilization. BMC Genom. 2019, 20, 504. [Google Scholar] [CrossRef]
- Su, L.K.; Lu, C.P.; Wang, Y.; Cao, D.M.; Sun, J.H.; Yan, Y.X. Lysogenic infection of a Shiga toxin 2-converting bacteriophage changes host gene expression, enhances host acid resistance and motility. Mol. Biol. (Mosk) 2010, 44, 60–73. [Google Scholar] [CrossRef]
- Veses-Garcia, M.; Liu, X.; Rigden, D.J.; Kenny, J.G.; McCarthy, A.J.; Allison, H.E. Transcriptomic analysis of Shiga-toxigenic bacteriophage carriage reveals a profound regulatory effect on acid resistance in Escherichia coli. Appl. Environ. Microbiol. 2015, 81, 8118–8125. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Doria, J.D.; Sperandio, V. Bacteriophage Transcription Factor Cro Regulates Virulence Gene Expression in Enterohemorrhagic Escherichia coli. Cell Host Microbe 2018, 23, 607–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, S.A.; Weber, M.; Schaufler, K.; Berens, C.; Geue, L.; Menge, C. Metabolic Traits of Bovine Shiga Toxin-Producing Escherichia coli (STEC) Strains with Different Colonization Properties. Toxins 2020, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P. Role of O-antigen variation in the immune response. Trends Microbiol. 1995, 3, 381–386. [Google Scholar] [CrossRef]
- Bazaka, K.; Crawford, R.J.; Nazarenko, E.L.; Ivanova, E.P. Bacterial extracellular polysaccharides. Adv. Exp. Med. Biol. 2011, 715, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Qu, W.; Reeves, P.R. Sequence analysis of four Shigella boydii O-antigen loci: Implication for Escherichia coli and Shigella relationships. Infect. Immun. 2001, 69, 6923–6930. [Google Scholar] [CrossRef] [Green Version]
- Geue, L.; Menge, C.; Eichhorn, I.; Semmler, T.; Wieler, L.H.; Pickard, D.; Berens, C.; Barth, S.A. Evidence for Contemporary Switching of the O-Antigen Gene Cluster between Shiga Toxin-Producing Escherichia coli Strains Colonizing Cattle. Front. Microbiol. 2017, 8, 424. [Google Scholar] [CrossRef] [Green Version]
- Basu, I.; Ferens, W.A.; Stone, D.M.; Hovde, C.J. Antiviral activity of shiga toxin requires enzymatic activity and is associated with increased permeability of the target cells. Infect. Immun. 2003, 71, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Ferens, W.A.; Hovde, C.J. The non-toxic A subunit of Shiga toxin type 1 prevents replication of bovine immunodeficiency virus in infected cells. Virus Res. 2007, 125, 29–41. [Google Scholar] [CrossRef]
- Ferens, W.A.; Halver, M.; Gustin, K.E.; Ott, T.; Hovde, C.J. Differential sensitivity of viruses to the antiviral activity of Shiga toxin 1 A subunit. Virus Res. 2007, 125, 104–108. [Google Scholar] [CrossRef]
- Ferens, W.A.; Cobbold, R.; Hovde, C.J. Intestinal Shiga toxin-producing Escherichia coli bacteria mitigate bovine leukemia virus infection in experimentally infected sheep. Infect. Immun. 2006, 74, 2906–2916. [Google Scholar] [CrossRef] [Green Version]
- Ferens, W.A.; Haruna, J.; Cobbold, R.; Hovde, C.J. Low numbers of intestinal Shiga toxin-producing E. coli correlate with a poor prognosis in sheep infected with bovine leukemia virus. J. Vet. Sci. 2008, 9, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, K.M.; Levin, B.R. Grazing protozoa and the evolution of the Escherichia coli O157:H7 Shiga toxin-encoding prophage. Proc. Biol. Sci. 2007, 274, 1921–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.E.; Shringi, S.; Besser, T.E. Protozoan Predation of Escherichia coli O157:H7 Is Unaffected by the Carriage of Shiga Toxin-Encoding Bacteriophages. PLoS ONE 2016, 11, e0147270. [Google Scholar] [CrossRef] [Green Version]
- George, A.S.; Rehfuss, M.Y.M.; Parker, C.T.; Brandl, M.T. The transcriptome of Escherichia coli O157: H7 reveals a role for oxidative stress resistance in its survival from predation by Tetrahymena. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef] [PubMed]
- Chekabab, S.M.; Daigle, F.; Charette, S.J.; Dozois, C.M.; Harel, J. Shiga toxins decrease enterohaemorrhagic Escherichia coli survival within Acanthamoeba castellanii. FEMS Microbiol. Lett. 2013, 344, 86–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, L.; McKendrick, I.J.; Ternent, H.; Gunn, G.J.; Synge, B.; Woolhouse, M.E. Super-shedding cattle and the transmission dynamics of Escherichia coli O157. Epidemiol. Infect. 2006, 134, 131–142. [Google Scholar] [CrossRef]
- Robinson, S.E.; Brown, P.E.; Wright, E.J.; Hart, C.A.; French, N.P. Quantifying within- and between-animal variation and uncertainty associated with counts of Escherichia coli O157 occurring in naturally infected cattle faeces. J. R. Soc. Interface 2009, 6, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Cull, C.A.; Paddock, Z.D.; Nagaraja, T.G.; Bello, N.M.; Babcock, A.H.; Renter, D.G. Efficacy of a vaccine and a direct-fed microbial against fecal shedding of Escherichia coli O157:H7 in a randomized pen-level field trial of commercial feedlot cattle. Vaccine 2012, 30, 6210–6215. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.P.; McCabe, E.; Murphy, M.; Buckley, J.F.; Crowley, D.; Fanning, S.; Duffy, G. Longitudinal Study of Two Irish Dairy Herds: Low Numbers of Shiga Toxin-Producing Escherichia coli O157 and O26 Super-Shedders Identified. Front. Microbiol. 2016, 7, 1850. [Google Scholar] [CrossRef] [Green Version]
- Matthews, L.; Reeve, R.; Gally, D.L.; Low, J.C.; Woolhouse, M.E.; McAteer, S.P.; Locking, M.E.; Chase-Topping, M.E.; Haydon, D.T.; Allison, L.J.; et al. Predicting the public health benefit of vaccinating cattle against Escherichia coli O157. Proc. Natl. Acad. Sci. USA 2013, 110, 16265–16270. [Google Scholar] [CrossRef] [Green Version]
- Cobbold, R.; Desmarchelier, P. Horizontal transmission of Shiga toxin-producing Escherichia coli within groups of dairy calves. Appl. Environ. Microbiol. 2002, 68, 4148–4152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Diemen, P.M.; Dziva, F.; Abu-Median, A.; Wallis, T.S.; van den Bosch, H.; Dougan, G.; Chanter, N.; Frankel, G.; Stevens, M.P. Subunit vaccines based on intimin and Efa-1 polypeptides induce humoral immunity in cattle but do not protect against intestinal colonisation by enterohaemorrhagic Escherichia coli O157:H7 or O26:H. Vet. Immunol. Immunopathol. 2007, 116, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Dziva, F.; Vlisidou, I.; Crepin, V.F.; Wallis, T.S.; Frankel, G.; Stevens, M.P. Vaccination of calves with EspA, a key colonisation factor of Escherichia coli O157:H7, induces antigen-specific humoral responses but does not confer protection against intestinal colonisation. Vet. Microbiol. 2007, 123, 254–261. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, T.N.; Naylor, S.W.; Mahajan, A.; Mitchell, M.C.; McAteer, S.; Deane, D.; Smith, D.G.; Low, J.C.; Gally, D.L.; Huntley, J.F. Escherichia coli O157:H7 colonization in cattle following systemic and mucosal immunization with purified H7 flagellin. Infect. Immun. 2008, 76, 2594–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieckens, E.; Rybarczyk, J.; Barth, S.A.; Menge, C.; Cox, E.; Vanrompay, D. Effect of lactoferrin on release and bioactivity of Shiga toxins from different Escherichia coli O157:H7 strains. Vet. Microbiol. 2017, 202, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Rybarczyk, J.; Khalenkow, D.; Kieckens, E.; Skirtach, A.G.; Cox, E.; Vanrompay, D. Lactoferrin translocates to the nucleus of bovine rectal epithelial cells in the presence of Escherichia coli O157:H7. Vet. Res. 2019, 50, 75. [Google Scholar] [CrossRef] [Green Version]
- Kieckens, E.; Rybarczyk, J.; De Zutter, L.; Duchateau, L.; Vanrompay, D.; Cox, E. Clearance of Escherichia coli O157:H7 infection in calves by rectal administration of bovine lactoferrin. Appl. Environ. Microbiol. 2015, 81, 1644–1651. [Google Scholar] [CrossRef] [Green Version]
- Kuribayashi, T.; Seita, T.; Fukuyama, M.; Furuhata, K.; Honda, M.; Matsumoto, M.; Seguchi, H.; Yamamoto, S. Neutralizing activity of bovine colostral antibody against verotoxin derived from enterohemorrhagic Escherichia coli O157:H7 in mice. J. Infect. Chemother. 2006, 12, 251–256. [Google Scholar] [CrossRef]
- Martorelli, L.; Garimano, N.; Fiorentino, G.A.; Vilte, D.A.; Garbaccio, S.G.; Barth, S.A.; Menge, C.; Ibarra, C.; Palermo, M.S.; Cataldi, A. Efficacy of a recombinant Intimin, EspB and Shiga toxin 2B vaccine in calves experimentally challenged with Escherichia coli O157:H7. Vaccine 2018, 36, 3949–3959. [Google Scholar] [CrossRef]
- MacLeod, D.L.; Gyles, C.L. Immunization of pigs with a purified Shiga-like toxin II variant toxoid. Vet. Microbiol. 1991, 29, 309–318. [Google Scholar] [CrossRef]
- Hovde, C.J.; Calderwood, S.B.; Mekalanos, J.J.; Collier, R.J. Evidence that glutamic acid 167 is an active-site residue of Shiga-like toxin I. Proc. Natl. Acad. Sci. USA 1988, 85, 2568–2572. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, S.; Furutani, M.; Ito, K.; Igarashi, K.; Nishibuchi, M.; Takeda, Y. Importance of arginine at position 170 of the A subunit of Vero toxin 1 produced by enterohemorrhagic Escherichia coli for toxin activity. Microb. Pathog. 1991, 11, 1–9. [Google Scholar] [CrossRef]
- Makino, S.; Watarai, M.; Tabuchi, H.; Shirahata, T.; Furuoka, H.; Kobayashi, Y.; Takeda, Y. Genetically modified Shiga toxin 2e (Stx2e) producing Escherichia coli is a vaccine candidate for porcine edema disease. Microb. Pathog. 2001, 31, 1–8. [Google Scholar] [CrossRef]
- Ohmura-Hoshino, M.; Yamamoto, M.; Yuki, Y.; Takeda, Y.; Kiyono, H. Non-toxic Stx derivatives from Escherichia coli possess adjuvant activity for mucosal immunity. Vaccine 2004, 22, 3751–3761. [Google Scholar] [CrossRef]
- Ishikawa, S.; Kawahara, K.; Kagami, Y.; Isshiki, Y.; Kaneko, A.; Matsui, H.; Okada, N.; Danbara, H. Protection against Shiga toxin 1 challenge by immunization of mice with purified mutant Shiga toxin 1. Infect. Immun. 2003, 71, 3235–3239. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Mateo, K.; Zhao, M.; Lin, J.; Zhang, W.; Francis, D.H. Avirulent K88 (F4)+ Escherichia coli strains constructed to express modified enterotoxins protect young piglets from challenge with a virulent enterotoxigenic Escherichia coli strain that expresses the same adhesion and enterotoxins. Vet. Microbiol. 2012, 159, 337–342. [Google Scholar] [CrossRef]
- Mayer, C.L.; Leibowitz, C.S.; Kurosawa, S.; Stearns-Kurosawa, D.J. Shiga toxins and the pathophysiology of hemolytic uremic syndrome in humans and animals. Toxins 2012, 4, 1261–1287. [Google Scholar] [CrossRef] [Green Version]
- Cobbold, R.; Desmarchelier, P. A longitudinal study of Shiga-toxigenic Escherichia coli (STEC) prevalence in three Australian diary herds. Vet. Microbiol. 2000, 71, 125–137. [Google Scholar] [CrossRef]
- Martorelli, L.; Albanese, A.; Vilte, D.; Cantet, R.; Bentancor, A.; Zolezzi, G.; Chinen, I.; Ibarra, C.; Rivas, M.; Mercado, E.C.; et al. Shiga toxin-producing Escherichia coli (STEC) O22:H8 isolated from cattle reduces E. coli O157:H7 adherence in vitro and in vivo. Vet. Microbiol. 2017, 208, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Carter, M.Q. Decoding the Ecological Function of Accessory Genome. Trends Microbiol. 2017, 25, 6–8. [Google Scholar] [CrossRef]
- Leopold, S.R.; Magrini, V.; Holt, N.J.; Shaikh, N.; Mardis, E.R.; Cagno, J.; Ogura, Y.; Iguchi, A.; Hayashi, T.; Mellmann, A.; et al. A precise reconstruction of the emergence and constrained radiations of Escherichia coli O157 portrayed by backbone concatenomic analysis. Proc. Natl. Acad. Sci. USA 2009, 106, 8713–8718. [Google Scholar] [CrossRef] [Green Version]
- Caprioli, A.; Morabito, S.; Brugereb, H.; Oswald, E. Enterohaemorrhagic Escherichia coli: Emerging issues on virulence and modes of transmission. Vet. Res. 2005, 36, 289–311. [Google Scholar] [CrossRef] [Green Version]
- Naylor, S.W.; Gally, D.L.; Low, J.C. Enterohaemorrhagic E. coli in veterinary medicine. Int. J. Med. Microbiol. 2005, 295, 419–441. [Google Scholar] [CrossRef]
- Callaway, T.R.; Anderson, R.C.; Edrington, T.S.; Genovese, K.J.; Harvey, R.B.; Poole, T.L.; Nisbet, D.J. Recent pre-harvest supplementation strategies to reduce carriage and shedding of zoonotic enteric bacterial pathogens in food animals. Anim. Health Res. Rev. 2004, 5, 35–47. [Google Scholar] [CrossRef]
- Callaway, T.R.; Carr, M.A.; Edrington, T.S.; Anderson, R.C.; Nisbet, D.J. Diet, Escherichia coli O157:H7, and cattle: A review after 10 years. Curr. Issues Mol. Biol. 2009, 11, 67–79. [Google Scholar]
- Farrokh, C.; Jordan, K.; Auvray, F.; Glass, K.; Oppegaard, H.; Raynaud, S.; Thevenot, D.; Condron, R.; De Reu, K.; Govaris, A.; et al. Review of Shiga-toxin-producing Escherichia coli (STEC) and their significance in dairy production. Int. J. Food. Microbiol. 2013, 162, 190–212. [Google Scholar] [CrossRef]
- Vande Walle, K.; Vanrompay, D.; Cox, E. Bovine innate and adaptive immune responses against Escherichia coli O157:H7 and vaccination strategies to reduce faecal shedding in ruminants. Vet. Immunol. Immunopathol. 2013, 152, 109–120. [Google Scholar] [CrossRef]
- Snedeker, K.G.; Campbell, M.; Sargeant, J.M. A systematic review of vaccinations to reduce the shedding of Escherichia coli O157 in the faeces of domestic ruminants. Zoonoses Public Health 2012, 59, 126–138. [Google Scholar] [CrossRef]
- Van Donkersgoed, J.; Hancock, D.; Rogan, D.; Potter, A.A. Escherichia coli O157:H7 vaccine field trial in 9 feedlots in Alberta and Saskatchewan. Can. Vet. J. 2005, 46, 724–728. [Google Scholar]
- Mahajan, A.; Currie, C.G.; Mackie, S.; Tree, J.; McAteer, S.; McKendrick, I.; McNeilly, T.N.; Roe, A.; La Ragione, R.M.; Woodward, M.J.; et al. An investigation of the expression and adhesin function of H7 flagella in the interaction of Escherichia coli O157: H7 with bovine intestinal epithelium. Cell. Microbiol. 2009, 11, 121–137. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menge, C. The Role of Escherichia coli Shiga Toxins in STEC Colonization of Cattle. Toxins 2020, 12, 607. https://doi.org/10.3390/toxins12090607
Menge C. The Role of Escherichia coli Shiga Toxins in STEC Colonization of Cattle. Toxins. 2020; 12(9):607. https://doi.org/10.3390/toxins12090607
Chicago/Turabian StyleMenge, Christian. 2020. "The Role of Escherichia coli Shiga Toxins in STEC Colonization of Cattle" Toxins 12, no. 9: 607. https://doi.org/10.3390/toxins12090607
APA StyleMenge, C. (2020). The Role of Escherichia coli Shiga Toxins in STEC Colonization of Cattle. Toxins, 12(9), 607. https://doi.org/10.3390/toxins12090607