The Parotoid Gland Secretion from Peruvian Toad Rhinella horribilis (Wiegmann, 1833): Chemical Composition and Effect on the Proliferation and Migration of Lung Cancer Cells

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Fractionation of Parotoid Gland Secretions of R. horribilis
2.2. 1H NMR Analysis
2.3. HPLC-DAD-Q-TOF-MS/MS Analysis
2.3.1. Compounds Identified: Argininyl Diacids
2.3.2. Alkaloids and Guanidine Derivatives
2.3.3. Bufadienolides
2.3.4. Argininyl Diacids of Bufadienolides
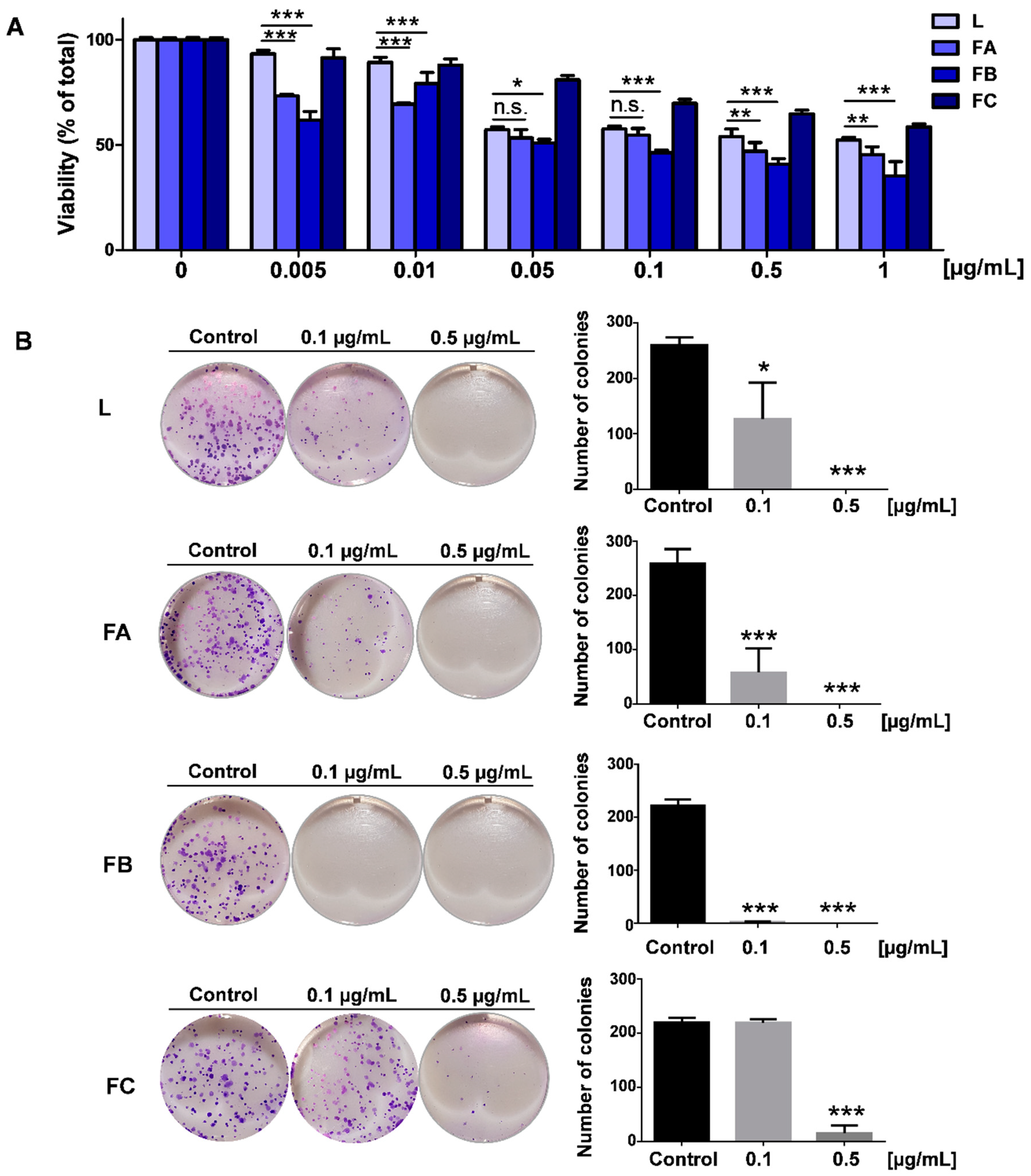
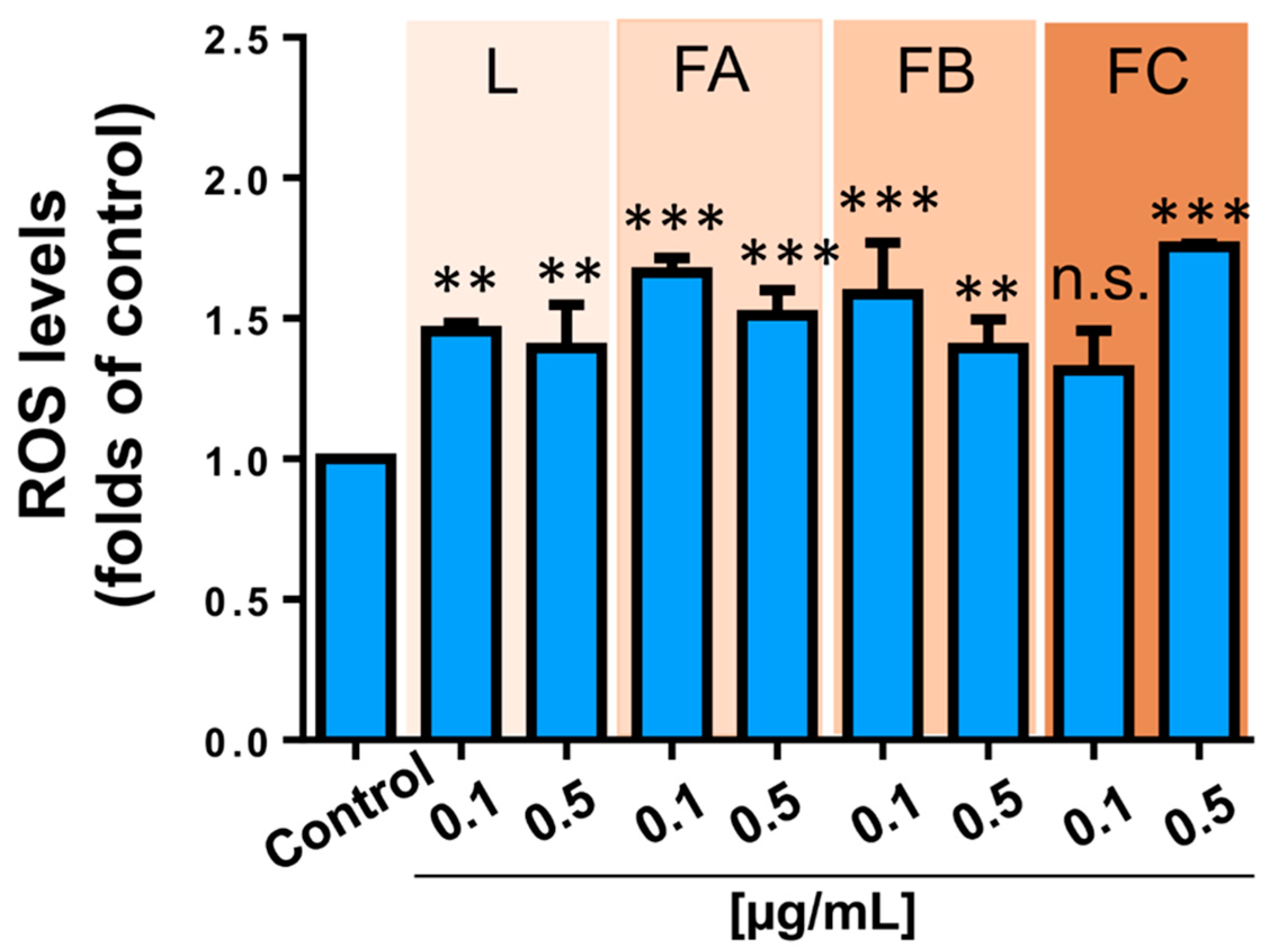
2.4. Effect of R. horribilis PGS on Proliferation, Clonogenic Capacity and Reactive Oxygen Species (ROS) Levels of Lung Cancer Cells
2.5. Effect of R. horribilis PGS and Their Fractions on Etoposide-Induced A549 Cell Death
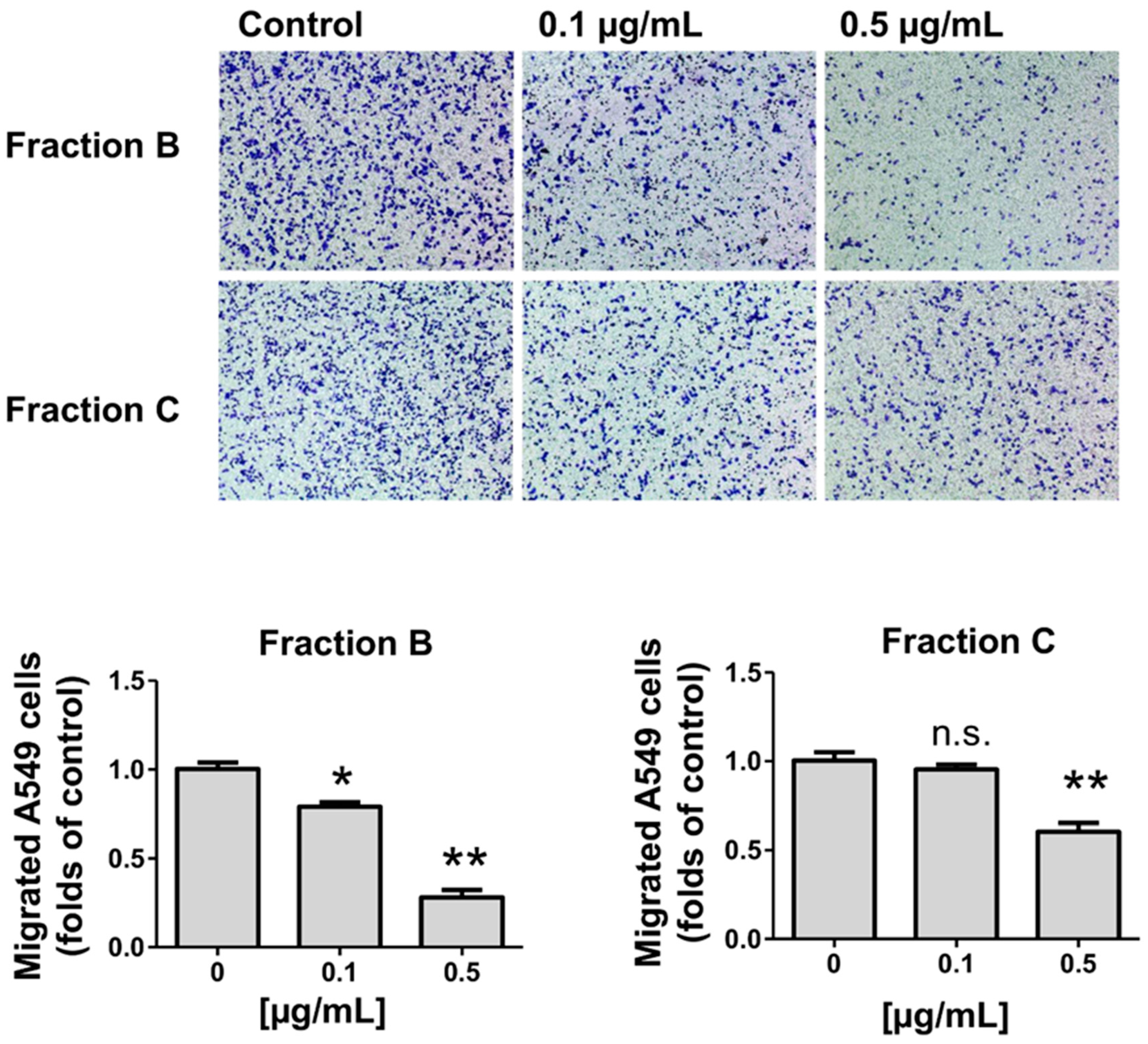
2.6. Effect of Fractions B and C of the R. horribilis PGS on the Migration of Lung Cancer Cells
3. Conclusions
4. Materials and Methods
4.1. Parotid Glands Secretions (PGS) Samples
4.2. Thin-Layer Chromatography Analysis
4.3. HPLC-DAD Analysis
4.4. HPC-DAD-Q-TOF-MS/MS Analysis
4.5. NMR Analysis
4.6. Cell Culture Conditions
4.7. Colony Formation
4.8. Determination of ROS Levels
4.9. Migration Assay
4.10. Annexin V/Propidium Iodide Staining
4.11. Statistics Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Catenazzi, A.; Lehr, E.; May, R.V. The amphibians and reptiles of Manu National Park and its buffer zone, Amazon basin and eastern slopes of the Andes, Peru. Biota Neotrop. 2013, 13, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Moravec, J.; Lehr, E.; Cusi, J.C.; Córdova, J.H.; Gvoždík, V. A new species of the Rhinella margaritifera species group (Anura, Bufonidae) from the montane forest of the Selva Central, Peru. Zookeys 2014, 35–56. [Google Scholar] [CrossRef] [Green Version]
- Pejic, R.; Simic Prskalo, M.; Simic, J. Ocular Hypotonia and Transient Decrease of Vision as a Consequence of Exposure to a Common Toad Poison. Case Rep. Ophthalmol. Med. 2020, 2020, 2983947. [Google Scholar] [CrossRef] [Green Version]
- Isoardi, K.; Lee, S.; Burren, J.M.; George, J.; Goggin, A.; Chan, B.S. Bufadienolide toxicity in a child following cane toad egg ingestion. Emerg. Med. Australas. 2018, 30, 728–729. [Google Scholar] [CrossRef]
- Peterson, M.E.; Roberts, B.K. Toads. In Small Animal Toxicology, 3rd ed.; Peterson, M.E., Talcott, P.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 833–839. [Google Scholar]
- McClelland, D. The Moche Botanical Frog. Arqueol. Iberoam. 2011, 10, 30–42. [Google Scholar] [CrossRef]
- Donnan, C.B. Moche Art of Peru; Museum of Cultural Heritage, University of California: Los Angeles, CA, USA, 1978; p. 205. [Google Scholar]
- Abdelfatah, S.; Lu, X.; Schmeda-Hirschmann, G.; Efferth, T. Cytotoxicity and antimitotic activity of Rhinella schneideri and Rhinella marina venoms. J. Ethnopharmacol. 2019, 242, 112049. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Gomez, C.V.; Rojas de Arias, A.; Burgos-Edwards, A.; Alfonso, J.; Rolon, M.; Brusquetti, F.; Netto, F.; Urra, F.A.; Cárdenas, C. The Paraguayan Rhinella toad venom: Implications in the traditional medicine and proliferation of breast cancer cells. J. Ethnopharmacol. 2017, 199, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Quispe, C.; Theoduloz, C.; de Sousa, P.T., Jr.; Parizotto, C. Antiproliferative activity and new argininyl bufadienolide esters from the “cururú” toad Rhinella (Bufo) schneideri. J. Ethnopharmacol. 2014, 155, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Indini, A.; Rijavec, E.; Bareggi, C.; Grossi, F. Novel treatment strategies for early-stage lung cancer: The oncologist’s perspective. J. Thorac. Dis. 2020, 12, 3390–3398. [Google Scholar] [CrossRef] [PubMed]
- Majem, B.; Nadal, E.; Muñoz-Pinedo, C. Exploiting metabolic vulnerabilities of Non small cell lung carcinoma. Semin. Cell Dev. Biol. 2020, 98, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Rigas, J.R. Taxane-platinum combinations in advanced non-small cell lung cancer: A review. Oncologist 2004, 9 (Suppl. S2), 16–23. [Google Scholar] [CrossRef]
- Frisone, D.; Friedlaender, A.; Malapelle, U.; Banna, G.; Addeo, A. A BRAF new world. Crit. Rev. Oncol. Hematol. 2020, 152, 103008. [Google Scholar] [CrossRef] [PubMed]
- Gandalovičová, A.; Rosel, D.; Fernandes, M.; Veselý, P.; Heneberg, P.; Čermák, V.; Petruželka, L.; Kumar, S.; Sanz-Moreno, V.; Brábek, J. Migrastatics-Anti-metastatic and Anti-invasion Drugs: Promises and Challenges. Trends Cancer 2017, 3, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čermák, V.; Dostál, V.; Jelínek, M.; Libusová, L.; Kovář, J.; Rösel, D.; Brábek, J. Microtubule-targeting agents and their impact on cancer treatment. Eur. J. Cell Biol. 2020, 99, 151075. [Google Scholar] [CrossRef] [PubMed]
- Sciani, J.M.; Angeli, C.B.; Antoniazzi, M.M.; Jared, C.; Pimenta, D.C. Differences and similarities among parotoid macrogland secretions in South American toads: A preliminary biochemical delineation. Sci. World J. 2013, 2013, 937407. [Google Scholar] [CrossRef] [Green Version]
- Schmeda-Hirschmann, G.; Quispe, C.; Arana, G.V.; Theoduloz, C.; Urra, F.A.; Cárdenas, C. Antiproliferative activity and chemical composition of the venom from the Amazonian toad Rhinella marina (Anura: Bufonidae). Toxicon 2016, 121, 119–129. [Google Scholar] [CrossRef]
- Sousa, L.Q.; Machado, K.D.; Oliveira, S.F.; Araújo, L.D.; Monção-Filho, E.D.; Melo-Cavalcante, A.A.; Vieira-Júnior, G.M.; Ferreira, P.M. Bufadienolides from amphibians: A promising source of anticancer prototypes for radical innovation, apoptosis triggering and Na(+)/K(+)-ATPase inhibition. Toxicon 2017, 127, 63–76. [Google Scholar] [CrossRef]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial peptides as anticancer agents: Functional properties and biological activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef]
- Kunda, N.K. Antimicrobial peptides as novel therapeutics for non-small cell lung cancer. Drug Discov. Today 2020, 25, 238–247. [Google Scholar] [CrossRef]
- Xia, F.; Gao, F.; Yao, H.; Zhang, G.; Gao, B.; Lu, Y.; Wang, X.; Qian, Y. Identification of angiogenesis-inhibiting peptides from Chan Su. Protein Expr. Purif. 2019, 163, 105445. [Google Scholar] [CrossRef]
- Han, Y.; Lu, M.; Zhou, J. Buforin IIb induces androgen-independent prostate cancer cells apoptosis though p53 pathway in vitro. Toxicon 2019, 168, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, A.A.; Lampo, M.; Cipriani, R. The cane or marine toad, Rhinella marina (Anura, Bufonidae): Two genetically and morphologically distinct species. Zootaxa 2016, 4103, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Orihuela-Torres, A.; Ordóñez-Delgado, L.; Brito, J.; López, F.; Mazón, M.; Freile, J.F. Feeding ecology of the Burrowing Owl Athene cunicularia punensis (Strigiformes: Strigidae) in the Jambelí archipelago, El Oro province, southwestern Ecuador. Rev. Peru. Biol. 2018, 25, 123–130. [Google Scholar] [CrossRef]
- de Medeiros, D.S.S.; Rego, T.B.; Dos Santos, A.P.A.; Pontes, A.S.; Moreira-Dill, L.S.; Matos, N.B.; Zuliani, J.P.; Stábeli, R.G.; Teles, C.B.G.; Soares, A.M.; et al. Biochemical and Biological profile of parotoid secretion of the Amazonian Rhinella marina (Anura: Bufonidae). Biomed. Res. Int. 2019, 2019, 2492315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamano, Y.; Nogawa, T.; Yamashita, A.; Pettit, G. The 1H and 13C NMR chemical shift assignments for thirteen bufadienolides isolated from the traditional Chinese drug Ch’an Su. Collect. Czechoslov. Chem. Commun. 2001, 66, 1841–1848. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, J.; Pan, H.; Wang, L. Chemical profile and multicomponent quantitative analysis for the quality evaluation of toad venom from different origins. Molecules 2019, 24, 3595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Guo, D.A. Analysis of bufadienolides in the Chinese drug ChanSu by high-performance liquid chromatography with atmospheric pressure chemical ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 1881–1892. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.L.; Hou, J.J.; Wang, X.; Yu, Y.; Li, H.J.; Li, Z.W.; Feng, Z.J.; Qu, H.; Wu, W.Y.; Guo, D.A. Venenum bufonis: An overview of its traditional use, natural product chemistry, pharmacology, pharmacokinetics and toxicology. J. Ethnopharmacol. 2019, 237, 215–235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jin, H.; Li, X.; Zhao, J.; Guo, X.; Wang, J.; Guo, Z.; Zhang, X.; Tao, Y.; Liu, Y.; et al. Separation and characterization of bufadienolides in toad skin using two-dimensional normal-phase liquid chromatography×reversed-phase liquid chromatography coupled with mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1026, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xiao, Y.; Xue, X.; Zhang, X.; Liang, X. Systematic screening and characterization of novel bufadienolides from toad skin using ultra-performance liquid chromatography/electrospray ionization quadrupole time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Han, L.; Luo, M.; Bian, B.; Guan, M.; Yang, H.; Han, C.; Li, N.; Li, T.; Li, S.; et al. Multi-component identification and target cell-based screening of potential bioactive compounds in toad venom by UPLC coupled with high-resolution LTQ-Orbitrap MS and high-sensitivity Qtrap MS. Anal. Bioanal. Chem. 2018, 410, 4419–4435. [Google Scholar] [CrossRef] [PubMed]
- Bessa-Silva, A.; Vallinoto, M.; Sampaio, I.; Flores-Villela, O.A.; Smith, E.N.; Sequeira, F. The roles of vicariance and dispersal in the differentiation of two species of the Rhinella marina species complex. Mol. Phylogenet. Evol. 2020, 145, 106723. [Google Scholar] [CrossRef] [PubMed]
- Kamano, Y.; Yamashita, A.; Nogawa, T.; Morita, H.; Takeya, K.; Itokawa, H.; Segawa, T.; Yukita, A.; Saito, K.; Katsuyama, M.; et al. QSAR evaluation of the Ch’an Su and related bufadienolides against the colchicine-resistant primary liver carcinoma cell line PLC/PRF/5(1). J. Med. Chem. 2002, 45, 5440–5447. [Google Scholar] [CrossRef] [PubMed]
- Nogawa, T.; Kamano, Y.; Yamashita, A.; Pettit, G.R. Isolation and structure of five new cancer cell growth inhibitory bufadienolides from the Chinese traditional drug Ch’an Su. J. Nat. Prod. 2001, 64, 1148–1152. [Google Scholar] [CrossRef]
- Xu, R.; Xie, H.Q.; Deng, L.L.; Zhang, J.X.; Yang, F.M.; Liu, J.H.; Hao, X.J.; Zhang, Y.H. A new bufadienolide with cytotoxic activity from the Chinese traditional drug Ch’an Su. Chin. J. Nat. Med. 2014, 12, 623–627. [Google Scholar] [CrossRef]
- Kim, N.Y.; Suh, Y.A.; Kim, S.; Lee, C. Bufalin down-regulates Axl expression to inhibit cell proliferation and induce apoptosis in non-small-cell lung cancer cells. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Cai, H.; Liu, Q.; Xia, Y.; Xing, L.; Zuo, Q.; Zhang, Y.; Chen, C.; Xu, K.; Yin, P.; et al. Cinobufacini Inhibits Colon Cancer Invasion and Metastasis via Suppressing Wnt/β-Catenin Signaling Pathway and EMT. Am. J. Chin. Med. 2020, 48, 703–718. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Q.; Zou, G.; Gao, G.; Yue, Q. Arenobufagin, isolated from toad venom, inhibited epithelial-to-mesenchymal transition and suppressed migration and invasion of lung cancer cells via targeting IKKβ/NFκB signal cascade. J. Ethnopharmacol. 2020, 250, 112492. [Google Scholar] [CrossRef]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Jeyapalan, J.; Leake, A.; Ahmed, S.; Saretzki, G.; Tilby, M.; von Zglinicki, T. The role of telomeres in Etoposide induced tumor cell death. Cell Cycle 2004, 3, 1169–1176. [Google Scholar] [CrossRef]
- Jain, C.K.; Roychoudhury, S.; Majumder, H.K. Selective killing of G2 decatenation checkpoint defective colon cancer cells by catalytic topoisomerase II inhibitor. Biochim. Biophys. Acta 2015, 1853, 1195–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, B.; Beljin, M.; Stark, G.R.; Taylor, W.R. G2 arrest in response to topoisomerase II inhibitors: The role of p53. Cancer Res. 2003, 63, 4074–4081. [Google Scholar]
- Chiu, C.C.; Li, C.H.; Ung, M.W.; Fuh, T.S.; Chen, W.L.; Fang, K. Etoposide (VP-16) elicits apoptosis following prolonged G2-M cell arrest in p53-mutated human non-small cell lung cancer cells. Cancer Lett. 2005, 223, 249–258. [Google Scholar] [CrossRef]
- Ceballos-Cancino, G.; Espinosa, M.; Maldonado, V.; Melendez-Zajgla, J. Regulation of mitochondrial Smac/DIABLO-selective release by survivin. Oncogene 2007, 26, 7569–7575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meresse, P.; Dechaux, E.; Monneret, C.; Bertounesque, E. Etoposide: Discovery and medicinal chemistry. Curr. Med. Chem. 2004, 11, 2443–2466. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Song, B.; Jin, H.; Pi, J.; Liu, L.; Jiang, J.; Cai, J. Cinobufacini induced MDA-MB-231 cell apoptosis-associated cell cycle arrest and cytoskeleton function. Bioorg. Med. Chem. Lett. 2012, 22, 1459–1463. [Google Scholar] [CrossRef]
- Hashimoto, S.; Jing, Y.; Kawazoe, N.; Masuda, Y.; Nakajo, S.; Yoshida, T.; Kuroiwa, Y.; Nakaya, K. Bufalin reduces the level of topoisomerase II in human leukemia cells and affects the cytotoxicity of anticancer drugs. Leuk. Res. 1997, 21, 875–883. [Google Scholar] [CrossRef]
- Liu, Y.; Ban, L.Y.; Su, X.; Gao, S.; Liu, J.W.; Cui, X.N. Effects of cinobufacini injection on cell proliferation and the expression of topoisomerases in human HepG-2 hepatocellular carcinoma cells. Mol. Med. Rep. 2015, 12, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.J.; Peng, Q.L.; Wang, L.H.; Xu, J.; Liu, J.S.; Li, Y.J.; Zhuo, Z.J.; Bai, L.L.; Hu, L.P.; Chen, W.M.; et al. Arenobufagin intercalates with DNA leading to G2 cell cycle arrest via ATM/ATR pathway. Oncotarget 2015, 6, 34258–34275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yuan, B.; Takagi, N.; Wang, H.; Zhou, Y.; Si, N.; Yang, J.; Wei, X.; Zhao, H.; Bian, B. Comparative analysis of hydrophilic ingredients in toad skin and toad venom using the UHPLC-HR-MS/MS and UPLC-QqQ-MS/MS methods together with the anti-inflammatory evaluation of indolealkylamines. Molecules 2018, 24, 86. [Google Scholar] [CrossRef] [Green Version]
- Soumoy, L.; Wells, M.; Najem, A.; Krayem, M.; Ghanem, G.; Hambye, S.; Saussez, S.; Blankert, B.; Journe, F. Toad venom antiproliferative activities on metastatic melanoma: Bio-guided fractionation and screening of the compounds of two different venoms. Biology 2020, 9, 218. [Google Scholar] [CrossRef] [PubMed]
- Sinhorin, A.P.; Kerkhoff, J.; Dall’Oglio, E.L.; de Jesus Rodrigues, D.; de Vasconcelos, L.G.; Sinhorin, V.D.G. Chemical profile of the parotoid gland secretion of the Amazonian toad (Rhinella margaritifera). Toxicon 2020, 182, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Wu, H.; Wu, H.; Wang, R.; Luo, C.; Gao, B.; Chen, Z.; Li, Q. Metabolites from Bufo gargarizans (Cantor, 1842): A review of traditional uses, pharmacological activity, toxicity and quality control. J. Ethnopharmacol. 2020, 246, 112178. [Google Scholar] [CrossRef] [PubMed]
- Thirupathi, K.; Chandrakala, G.; Krishna, L.; Bheem Rao, T.; Venkaiah, Y. Antibacterial activity of skin secretion and its extraction from the toad Bufo melanostictus. Eur. J. Pharm. Med Res. 2018, 3, 283–286. [Google Scholar]
- Fuentes-Retamal, S.; Sandoval-Acuna, C.; Peredo-Silva, L.; Guzman-Rivera, D.; Pavani, M.; Torrealba, N.; Truksa, J.; Castro-Castillo, V.; Catalan, M.; Kemmerling, U.; et al. Complex Mitochondrial Dysfunction Induced by TPP(+)-Gentisic Acid and Mitochondrial Translation Inhibition by Doxycycline Evokes Synergistic Lethality in Breast Cancer Cells. Cells 2020, 9, 407. [Google Scholar] [CrossRef] [Green Version]
- Urra, F.A.; Munoz, F.; Cordova-Delgado, M.; Ramirez, M.P.; Pena-Ahumada, B.; Rios, M.; Cruz, P.; Ahumada-Castro, U.; Bustos, G.; Silva-Pavez, E.; et al. FR58P1a; a new uncoupler of OXPHOS that inhibits migration in triple-negative breast cancer cells via Sirt1/AMPK/beta1-integrin pathway. Sci. Rep. 2018, 8, 13190. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Rt (min) | Sample | [M+H]+ Detected | [M+H]+ Expected | Error (ppm) | MS/MS Fragmentation | Tentative Identification |
|---|---|---|---|---|---|---|---|
| 1 | 11.4–11.9 | L | 303.1624 | 303.1663 | 12.8 | 303.1624 (<1), 244.1169 (32), 222.1210 (14), 175.1184 (27), 158.0852 (22), 130.1935 (14), 128.0688 (26), 116.0685 (100), 95.0600 (39). | Adipoyl arginine C12H23N4O5+ [28] |
| A1 | 12.8 | L | 176.0685 | 176.0666 | 10.8 | 176.0688 (<1), 159.0660 (17), 143.0720 (9), 133.0536 (16), 117.0560 (52), 115.0533 (100), 115.0533 (100), 105.0673 (31), 89.0368 (17) | Guanidinosuccinic acid C5H10N3O4+ [28] |
| 2 | 15.6–17.0 | A, B, C, L | 317.1804 | 317.1819 | 4.7 | 317.1804 (<1), 273.4732 (40), 194.1169 (52), 158.0895 (65), 125.0623 (87), 112.0849 (100) | Pimeloyl arginine C13H25N4O5+ [28] |
| A2 | 15.7–17.2 | A, B, C, L | 203.1178 | 203.1179 | 0.5 | 203.1178 (<1), 188.0932 (50), 173.0708 (100), 145.0764 (9), 118.0667 (10), 89.0364 (16) | Dehydrobufotenine C12H15N2O+ [28] |
| 3 | 21.8–22.7 | A, B, C, L | 331.1962 | 331.1976 | 4.2 | 331.1980 (20), 272.1486 (100), 172.1057 (86), 125.0603 (48), 112.0879 (46) | Suberoyl arginine C14H27N4O5+ [28] |
| 5 | 25.2–25.5 | A, B, C, L | 359.2260 | 359.2289 | 8.0 | 359.2366 (21), 272.1458 (100), 246.4343 (17), 226.1422 (20), 203.1506 (33), 116.0996 (35) | Sebacyl arginine isomer C16H31N4O5+ [28] |
| 4 | 30.9–31.5 | A, B, C, L | 345.2123 | 345.2132 | 2.6 | 345.2100 (19), 286.1628 (27), 250.1529 (85), 175.1197 (58), 158.0932 (44), 139.0761 (42), 116.0709 (100) | Azelayl arginine C15H29N4O5+ [28] |
| 6 | 34.0–34.1 | A, L | 373.2417 | 373.2445 | 8.3 | 373.2341 (34), 286.1592 (100), 254.1458 (30), 203.1485 (40), 172.1096 (40), 116.0702 (75) | Undecadienoyl arginine C17H33N4O5+ [28] |
| 24 | 48.8–49.3 | A, L | 745.4000 | 745.4018 | 2.4 | 745.3983 (78), 727.3875 (13), 434.7749 (6), 331.1942 (100), 313.1949 (11), 278.1473 (33), 250.1558 (25), 195.1161 (17), 158.0890 (17), 117.0713 (31), 112.0840 (58), 91.0538 (27) | 3-(N-suberoyl argininyl) Hydroxyhellebrigenin isomer 1 C38H57N4O11+ [28,29] |
| 19 | 49.1–49.2 | L | 731.4225 | 731.4226 | 0.1 | 331.1932 (100), 314.1670 (18), 278.1441 (25), 175.1128 (10) | 3-(N-suberoyl argininyl) Hellebrigenol isomer 1 C38H59N4O10+ [28] |
| 14 | 49.3–49.7 | A, L | 715.3888 | 715.3913 | 3.5 | 715.3906 (40), 697.3787 (5), 363.1930 (5), 345.1796 (2), 317.1788 (100), 264.1340 (7), 175.1181 (4), 112.0878 (11) | 3-(N-pimeloyl argininyl) Desacetylcinobufotalin C37H55N4O10+ [32] |
| 10 | 49.8–50.1 | A, L | 687.3952 | 687.3964 | 1.7 | 687.3984 (46), 669.3618 (4), 385.2361 (5), 303.1627 (100), 268.1292 (11), 250.1171 (13), 105.0685 (12) | 3-(N-adipoyl argininyl) Gamabufotalin C36H55N4O9+ [28,32] |
| XIII | 50.6–50.9 | A, B, C, L | 417.2262 | 417.2272 | 2.4 | 417.2286 (100), 399.2198 (18), 381.2053 (50), 363.1946 (34), 345.1840 (33), 209.0929 (62), 165.0687 (62), 103.0526 (89), 91.0544 (94) | Arenobufagin C24H33O6+ [31]. |
| 23 | 50.8–50.9 | L | 731.3845 | 731.3862 | 2.3 | 731.3845 (35), 439.2103 (11), 317.1809 (100), 300.1562 (16), 264.1334 (20), 178.0778 (9), 155.0880 (13) | 3-(N-pimeloyl argininyl) Hydroxyhellebrigenin C37H55N4O11+ [28] |
| 20 | 51.5–51.7 | A, B, L | 701.3741 | 701.3756 | 2.1 | 701.3747 (96), 683.3639 (11), 665.3641 (2), 381.2004 (3), 364.1916 (5), 363.1984 (4), 335.1984 (18), 317.1910 (3), 303.1640 (100), 268.1239 (14), 250.1194 (17), 175.1192 (3), 129.0689 (16), 91.0528 (31) | 3-(N-adipoyl argininyl) Hellebrigenin C36H53N4O10+ [32] |
| 36 | 52.1–52.4 | A, B, L | 743.3854 | 743.3862 | 2.3 | 743.3860 (100), 725.3687 (7), 377.1703 (9), 331.1968 (99), 278.1481 (26), 250.1546 (23), 175.1230 (7), 165.0706 (14), 128.0629 (22), 112.0881 (24), 910533 (25) | 3-(N-suberoyl argininyl) Hydroxybufotalinin isomer 1 C38H55N4O11+ [28] |
| 11 | 53.3–54.1 | A, B, L | 701.4062 | 701.4120 | 8.2 | 701.4080 (95), 683.3979 (2), 385.2287 (4), 317.1767 (100), 264.1285 (33), 236.1363 (8), 105.0669 (20) | 3-(N-pimeloyl argininyl) Gamabufotalin C37H57N4O9+ [28] |
| 21 | 54.1–55.0 | A, L | 715.3879 | 715.3913 | 4.8 | 715.3864 (100), 697.3839 (7), 669.3725 (3), 381.2027 (5), 364.1992 (1), 363.1928 (5), 651.3676 (6), 353.2058 (2), 345.1816 (3), 335.2013 (8), 317.1787 (71), 282.1417 (8), 264.1309 (14), 236.1364 (9), 175.1165 (8), 112.0861 (23), 105.0702 (17), 91.0535 (27) | 3-(N-pimeloyl argininyl) Hellebrigenin C37H55N4O10+ [32] |
| 19 | 54.2–54.6 | A, L | 731.4171 | 731.4226 | 7.5 | 731.4143 (72), 363.1946 (12), 331.1891 (100), 278.1446 (35), 250.1450 (15), 175.1177 (17), 112.0873 (23) | 3-(N-suberoyl argininyl) Hellebrigenol isomer 2 C38H59N4O10+ |
| 13 | 54.3–54.8 | A, B, L | 701.3728 | 701.3756 | 4.0 | 701.3721 (100), 684.3766 (1), 610.9012 (1), 399.2135 (6), 381.2023 (2), 353.2069 (2), 335.1959 (1), 303.1635 (89), 286.1387 (9), 268.1256 (15), 250.1156 (17), 175.1180 (11), 116.0692 (16), 112.0878 (15), 105.0691 (13), 91.0537 (10) | 3-(N-adipoyl argininyl) Arenobufagin C36H53N4O10+ [32] |
| 36 | 54.7–54.9 | A, L | 743.3825 | 743.3862 | 4.9 | 743.3829 (65), 349.1888 (8), 331.1935 (100), 296.1560 (9), 278.1480 (25), 250.1528 (55), 233.0955 (4), 179.0855 (11), 112.0869 (34), 91.0555 (22) | 3-(N-suberoyl argininyl) Hydroxybufotalinin isomer 2 C38H55N4O11+ [28] |
| 34 | 55.1–55.4 | A, B, L | 713.3706 | 713.3756 | 7.0 | 713.3715 (100), 695.3598 (3), 667.3739 (4), 317.1781 (88), 300.1509(22), 282.1434(12), 264.1334 (12), 236.1370 (16), 209.0860 (5), 179.0818 (10), 175.1225 (7), 112.0855 (35), 91.0524 (43) | 3-(N-pimeloyl argininyl) Bufotalinin C37H53N4O10+ [28] |
| VI | 56.5–56.7 | A, B, L | 419.2400 | 419.2428 | 6.6 | 419.2400 (5), 401.2318 (30), 365.2088 (63), 347.2036 (36), 319.1615 (33), 213.1616 (27), 91.0531 (51) | Hellebrigenol C24H35O6+ [32] |
| IV or V | 57.0–57.6 | A, B, C, L | 417.2244 | 417.2272 | 6.7 | 417.2226 (100), 399.2122 (35), 371.2176 (4), 353.2056 (3), 335.1972 (5), 255.0976 (2), 175.0722 (4) | Bufarenogin or ψ-Bufarenogin C24H33O6+ [32] |
| 19 | 57.0–57.8 | A, L | 731.4184 | 731.4226 | 5.7 | 731.4189 (55), 713.4048 (5), 683.3909 (10), 665.3843 (4), 331.1919 (100), 278.1468 (10), 250.1475 (9), 175.1179 (4), 112.0854 (20) | 3-(N-suberoyl argininyl) Hellebrigenol isomer 3 C38H59N4O10+ [28] |
| 16 | 57.9–58.2 | A, L | 687.3947 | 687.3964 | 2.4 | 687.3960 (27), 669.3813 (7), 303.1641 (100), 250.1177 (13), 175.1446 (8), 145.0968 (14), 91.0533 (17) | 3-(N-adipoyl argininyl) Telocinobufagin C36H55N4O9+ [28] |
| 24 | 58.1–58.4 | A, L | 745.4001 | 745.4018 | 2.3 | 745.4017 (11), 683.4230 (11), 345.2155 (10), 331.1923 (100), 303.1952 (8), 278.1492 (10), 175.1148 (15), 112.0836 (35) | 3-(N-suberoyl argininyl) Hydroxyhellebrigenin isomer 2 C38H57N4O11+ [28] |
| 30 | 58.3–59.0 | A, B, C, L | 685.3780 | 685.3807 | 3.9 | 685.3757 (28), 669.3790 (5), 667.3653 (3), 365.2086 (3), 349.2058 (4), 303.1615 (100), 268.1240 (12), 250.1140 (12), 175.1140 (6), 105.0678 (13) | 3-(N-adipoyl argininyl) Marinobufagin C36H53N4O9+ [28] |
| 22 | 58.4–60.0 | A, L | 729.4021 | 729.4069 | 6.5 | 729.4010 (100), 711.3899 (8), 693.3760 (2), 683.3970 (2), 363.1912 (4), 331.1926 (68), 278.1459 (16), 250.1516 (16), 175.1188 (8), 112.0861 (27), 91.0534 (27) | 3-(N-suberoyl argininyl) Hellebrigenin C38H57N4O10+ [28,29] |
| XIV | 59.0–59.6 | A, B, C, L | 417.2242 | 417.2272 | 7.2 | 417.2244 (62), 399.2122 (26), 363.1903 (30), 353.2077 (26), 335.1959 (73), 317.1874 (10), 275.1754 (16), 211.1433 (33), 128.0608 (71), 91.0527 (100) | Hellebrigenin C24H33O6+ [28] |
| 31 | 60.5–64.3 | A, L | 699.3980 | 699.3964 | 2.3 | 699.3971 (38), 681.3823 (7), 365.2102 (4), 347.2001 (4), 317.1806 (100), 264.1314 (11), 236.1361 (6), 175.1198 (5), 112.0883 (10), 91.0523 (10) | 3-(N-pimeloyl argininyl) Marinobufagin C37H55N4O9+ [28] |
| XII | 60.8–61.4 | A, B, C, L | 415.2092 | 415.2115 | 5.5 | 415.2085 (22), 379.1865 (8), 361.1792 (12), 351.1927 (30), 333.1820 (26), 237.1619 (28), 165.0669 (30), 128.0609 (64), 115.0530 (79), 91.0534 (100) | Bufotalinin C24H31O6+ [32] |
| 17 | 62.4–64.7 | A, B, L | 701.4097 | 701.4120 | 3.3 | 701.4087 (24), 683.3955 (5), 349.2203 (3), 317.1786 (100), 264.1308 (5), 236.1406 (3), 175.1172 (3), 105.0711 (8) | 3-(N-pimeloyl argininyl) Telocinobufagin C37H57N4O9+ [28] |
| 33 | 62.7–63.7 | A, B, C, L | 683.3682 | 683.3651 | 3.6 | 683.3613 (100), 665.3521 (2), 335.1963 (4), 303.1635 (40), 286.1371 (9), 268.1273 (8), 250.1161 (17), 226.1057 (3), 175.1187 (5), 112.0873 (13) | 3-(N-adipoyl argininyl) Marinobufagin-9,11-ene C36H51N4O9+ |
| II | 62.9–63.4 | A, B, C, L | 403.2453 | 403.2479 | 6.4 | 403.2445 (30), 385.2335 (58), 349.2120 (44), 337.2111 (41), 275.1742 (25), 253.1888 (39), 241.1922 (49), 161.0968 (29), 105.0785 (46), 91.0546 (100) | Gamabufotalin C24H35O5+ [28,32] |
| VIII | 64.0-64.7 | A, B, C, L | 401.2312 | 401.2323 | 2.7 | 401.2295 (34), 383.2866 (3), 365.2081 (15), 347.1970 (20), 337.2165 (14), 319.2024 (10), 257.1163 (21), 239.1059 (32), 211.1096 (27), 183.1154 (24), 128.0626 (86), 105.0700 (85), 91.0547 (100) | Resibufaginol C24H33O5+ [31] |
| 15 | 64.4–66.5 | A, B, C, L | 729.4052 | 729.4069 | 2.3 | 729.4077 (100), 399.2156 (5), 381.2065 (2), 363.1959 (2), 353.2100 (1), 345.1862 (1), 335.1983 (2), 331.1959 (83), 278.1487 (25), 250.1541 (18), 175.1200 (11), 112.0875 (27). | 3-(N-suberoyl argininyl) Desacetylcinobufotalin C38H57N4O10+ [29] |
| 12 | 66.4–67.2 | A, B, C, L | 715.4254 | 715.4277 | 3.2 | 715.4250 (80), 697.4150 (12), 349.2108 (4), 331.1936 (100), 314.1679 (12), 278.1462 (32), 250.1514 (24), 175.1165 (9), 116.0691 (22), 91.0531 (16) | 3-(N-suberoyl argininyl) Gamabufotalin C38H59N4O9+ [28] |
| XI | 66.5–66.7 | B, L | 415.2090 | 415.2115 | 6.0 | 415.2054 (100), 397.1832 (3), 333.1887 (3), 283.1621 (5), 253.0760 (5), 175.0999 (6) | 19-Oxo-desacetylcinobufagin C24H31O6+ [28] |
| 28 | 66.5–69.5 | A, B, C, L | 697.3766 | 697.3807 | 5.9 | 697.3745 (100), 679.3595 (2), 363.1933 (3), 335.1966 (4), 317.1759 (61), 264.1290 (33), 236.1349 (11), 175.1154 (4), | 3-(N-pimeloyl argininyl) Resibufagin C37H53N4O9+ [28] |
| 18 | 68.6–72.0 | A, B, C, L | 715.4265 | 715.4277 | 1.7 | 715.4266 (44), 697.4156 (7), 349.2136 (4), 331.1944 (100), 314.1708 (3), 278.1484 (8), 250.1538 (5), 175.1193 (4), 105.0695 (11), 91.0540 (11) | 3-(N-suberoyl argininyl) Telocinobufagin C38H59N4O9+ [28] |
| 32 | 70.0–72.6 | A, B, C, L | 713.4101 | 713.4120 | 2.6 | 713.4100 (48), 695.3992 (5), 365.2075 (3), 331.1941 (100), 314.1681 (5), 296.1581 (3), 278.1473 (12), 250.1533 (8), 175.1198 (8), 112.0873 (16), 91.0536 (15) | 3-(N-suberoyl argininyl) Marinobufagin C38H57N4O9+ [28] |
| X | 71.8–72.6 | A, B, C, L | 399.2150 | 399.2166 | 4.0 | 399.2150 (<1), 381.2104 (24), 363.2394 (14), 331.1949 (35), 275.1786 (19), 239.1741 (52), 213.1297 (33), 128.0661 (46), 115.0554 (42), 105.0684 (67), 91.0533 (100) | Resibufagin C24H31O5+ [32] |
| 29 | 74.6–78.9 | A, B, C, L | 711.3941 | 711.3964 | 1.1 | 711.3956 (100), 363.1933 (3), 331.1937 (40), 278.1463 (24), 250.1522 (18), 175.1179 (14) | 3-(N-suberoyl argininyl) Resibufagin C38H55N4O9+ [28] |
| 7 | 74.6–75.8 | A, B, L | 671.4014 | 671.4014 | 0.0 | 671.4048 (64), 653.3916 (8), 351.2269 (5), 303.1638 (100), 286.1403 (6), 250.1151 (18), 175.1213 (3), 158.0926 (4), 105.0691 (13), 91.0525 (14) | 3-(N-adipoyl argininyl) bufalin C36H55N4O8+ [28] |
| III | 75.1–76.0 | A, B, C, L | 403.2464 | 403.2479 | 3.7 | 403.2456 (57), 385.2353 (37), 367.2252 (44), 349.2135 (79), 253.1929 (31), 215.1780 (30), 105.0700 (79), 91.0537 (100) | Telocinobufagin C24H35O5+ [29,31] |
| 35 | 76.4–77.5 | A, B, L | 727.3922 | 727.3913 | 4.0 | 727.3917 (100), 711.3898 (6), 397.1995 (9), 331.1962 (100), 278.1494 (35), 250.1532 (29), 175.1180 (21), 112.0885 (34), 91.0540 (23) | 3-(N-suberoyl argininyl) Bufotalinin C38H55N4O10+ [28] |
| IX | 78.3–79.2 | A, B, L | 401.2323 | 401.2323 | 0.0 | 401.2335 (23), 383.2235 (10), 365.2105 (51), 347.2012 (32), 337.2162 (17), 319.2068 (19), 269.1890 (20), 253.1938 (44), 251.1797 (31), 239.1061 (22), 183.1157 (25), 128.0639 (74), 105.0710 (81), 91.0543 (100) | Marinobufagin C24H33O5+ [32] |
| 25 | 82.6–86.3 | A, B, C, L | 669.3840 | 669.3858 | 2.7 | 669.3835 (100), 349.2124 (2), 303.1628 (79), 286.1395 (5), 268.1264 (14), 250.1156 (24), 175.1188 (8), 116.0695 (11), 91.0531 (6) | 3-(N-adipoyl argininyl) Resibufogenin C36H53N4O8+ [28] |
| 8 | 83.0–86.2 | A, L | 685.4142 | 685.4171 | 4.2 | 685.4113 (94), 667.4030 (11), 351.2260 (7), 317.1779 (100), 300.1539 (10), 264.1298 (24), 175.1181 (9), 112.0860 (22), 91.0520 (20) | 3-(N-pimeloyl argininyl) bufalin C37H57N4O8+ [28] |
| I | 87.8–88.7 | A, B, C, L | 387.2512 | 387.2530 | 4.6 | 387.2482 (60), 369.2396 (72), 351.2291 (52), 333.2204 (10), 255.2101 (57), 187.1454 (14), 173.1330 (16), 128.0637 (36), 105.0698 (88). 91.0541 (100) | Bufalin C24H35O4+ [31] |
| 26 | 91.4–94.6 | A, L | 683.4000 | 683.4014 | 2.0 | 683.4015 (100), 665.3974 (2), 349.2114 (5), 317.1791 (83), 300.1531 (11), 282.1435 (11), 264.1338 (25), 236.1358 (13), 175.1163 (10), 143.0834 (6), 112.0868 (16), 91.0546 (12). | 3-(N-pimeloyl argininyl) Resibufogenin C37H55N4O8+ [28] |
| 9 | 91.4–95.1 | A, B, C, L | 699.4313 | 699.4327 | 2.0 | 699.4311 (100), 681.4213 (14), 351.2279 (6), 331.1942 (78), 314.16814 (9), 296.1568 (6), 278.1477 (21), 250.1522 (14), 175.1169 (9), 112.0863 (21), 91.0541 (15) | 3-(N-suberoyl argininyl) bufalin C38H59N4O8+ [28] |
| VII | 97.3–98.3 | A, B, C, L | 385.2328 | 385.2373 | 7.7 | 385.2318 (8), 367.2221 (15), 349.2133 (12), 321.2151 (6), 253.1923 (19), 241.1195 (14), 205.0860 (16), 185.0928 (4), 115.0544 (49), 105.0689 (69), 91.0531 (100) | Resibufogenin C24H33O4+ [29,32] |
| 27 | 98.7–103.1 | A, B, C, L | 697.4117 | 697.4171 | 7.7 | 697.4110 (100), 679.4003 (2), 349.2108 (3), 331.1917 (85), 314.1667 (7), 278.1455 (26), 250.1503 (19), 175.1157 (9), 112.0854 (19), 910525 (11) | 3-(N-suberoyl argininyl) Resibufogenin C38H57N4O8+ [28] |
| Time | Crude Extract | Fraction A | Fraction B | Fraction C |
|---|---|---|---|---|
| 24 h | 0.031 ± 0.007 | 0.010 ± 0.001 | 0.007 ± 0.001 | 0.025 ± 0.001 |
| 48 h | 0.015 ± 0.001 a | 0.011 ± 0.003 | 0.006 ± 0.001 | 0.007 ± 0.002 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmeda-Hirschmann, G.; de Andrade, J.P.; Soto-Vasquez, M.R.; Alvarado-García, P.A.A.; Palominos, C.; Fuentes-Retamal, S.; Mellado, M.; Correa, P.; Urra, F.A. The Parotoid Gland Secretion from Peruvian Toad Rhinella horribilis (Wiegmann, 1833): Chemical Composition and Effect on the Proliferation and Migration of Lung Cancer Cells. Toxins 2020, 12, 608. https://doi.org/10.3390/toxins12090608
Schmeda-Hirschmann G, de Andrade JP, Soto-Vasquez MR, Alvarado-García PAA, Palominos C, Fuentes-Retamal S, Mellado M, Correa P, Urra FA. The Parotoid Gland Secretion from Peruvian Toad Rhinella horribilis (Wiegmann, 1833): Chemical Composition and Effect on the Proliferation and Migration of Lung Cancer Cells. Toxins. 2020; 12(9):608. https://doi.org/10.3390/toxins12090608
Chicago/Turabian StyleSchmeda-Hirschmann, Guillermo, Jean Paulo de Andrade, Marilú Roxana Soto-Vasquez, Paul Alan Arkin Alvarado-García, Charlotte Palominos, Sebastián Fuentes-Retamal, Mathias Mellado, Pablo Correa, and Félix A. Urra. 2020. "The Parotoid Gland Secretion from Peruvian Toad Rhinella horribilis (Wiegmann, 1833): Chemical Composition and Effect on the Proliferation and Migration of Lung Cancer Cells" Toxins 12, no. 9: 608. https://doi.org/10.3390/toxins12090608
APA StyleSchmeda-Hirschmann, G., de Andrade, J. P., Soto-Vasquez, M. R., Alvarado-García, P. A. A., Palominos, C., Fuentes-Retamal, S., Mellado, M., Correa, P., & Urra, F. A. (2020). The Parotoid Gland Secretion from Peruvian Toad Rhinella horribilis (Wiegmann, 1833): Chemical Composition and Effect on the Proliferation and Migration of Lung Cancer Cells. Toxins, 12(9), 608. https://doi.org/10.3390/toxins12090608