A Novel Apilic Antivenom to Treat Massive, Africanized Honeybee Attacks: A Preclinical Study from the Lethality to Some Biochemical and Pharmacological Activities Neutralization

 ,
,  , , ,
, , ,  ,
,  and
and 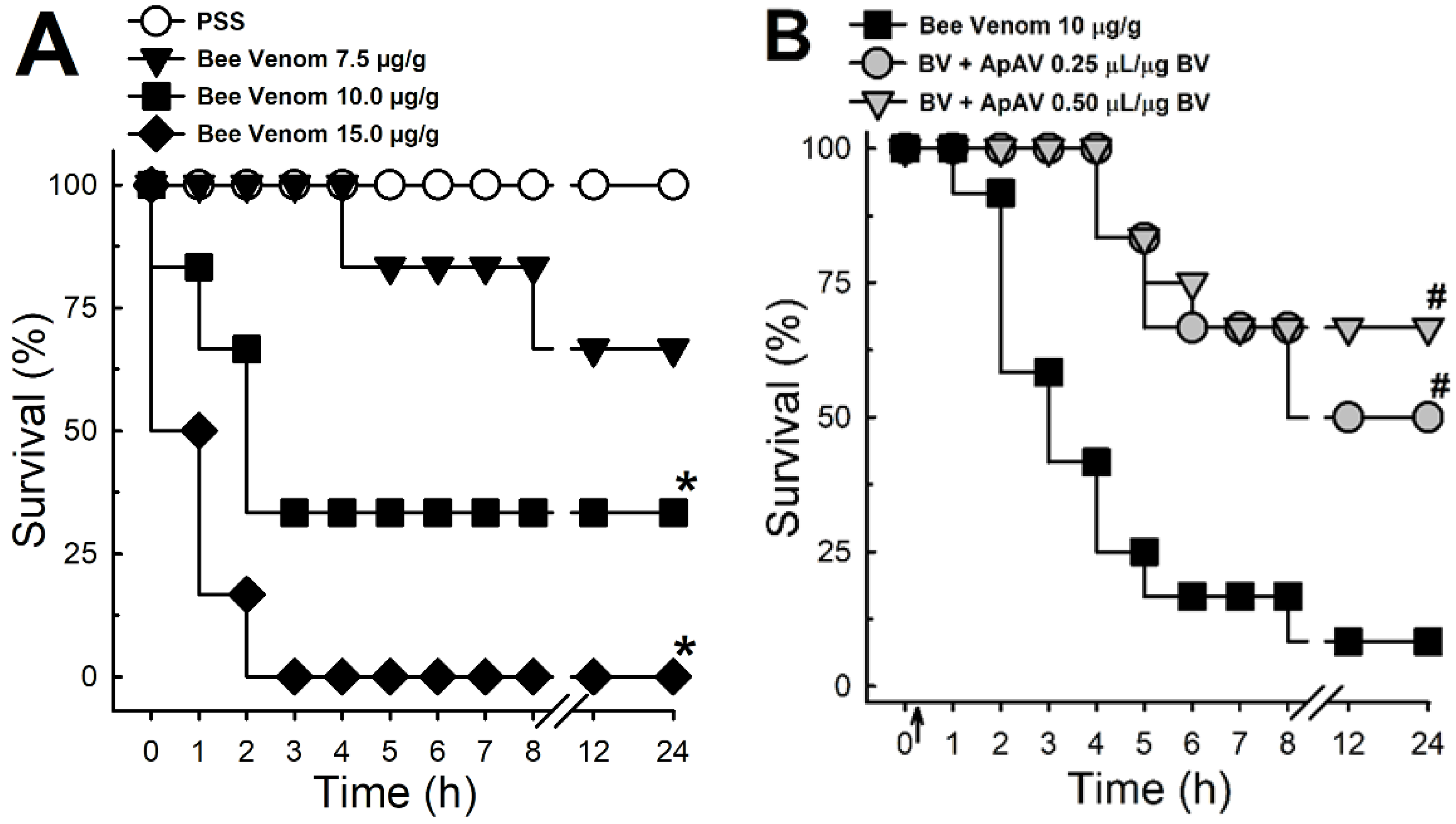{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ApAV Protection against Africanized A. mellifera Venom Lethality
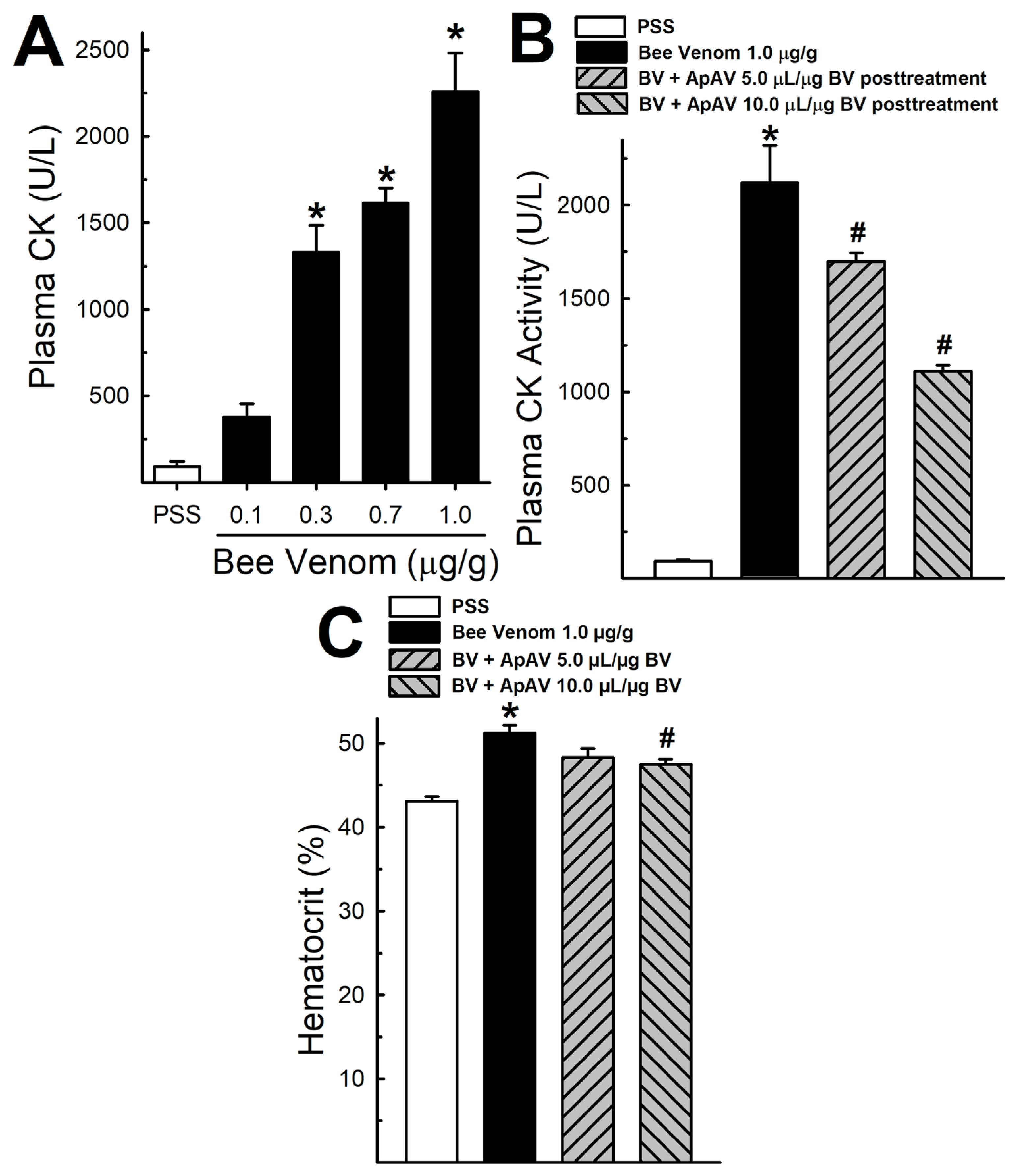
2.2. ApAV Inhibition on In Vivo Myotoxicity and Hemoconcentration Induced by Africanized A. mellifera Venom
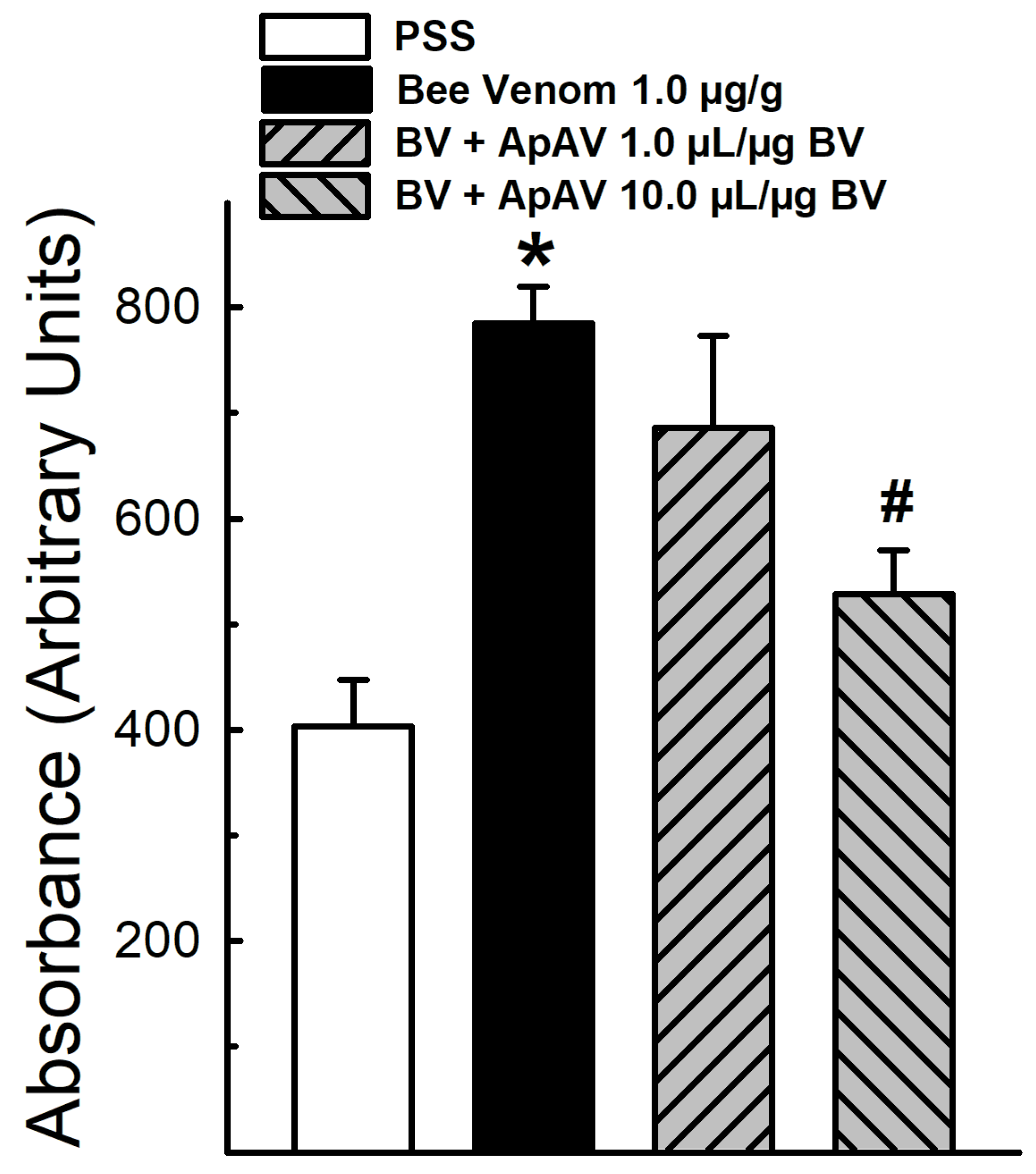
2.3. ApAV Inhibition of In Vivo Vascular Permeability Induced by Africanized A. mellifera Venom
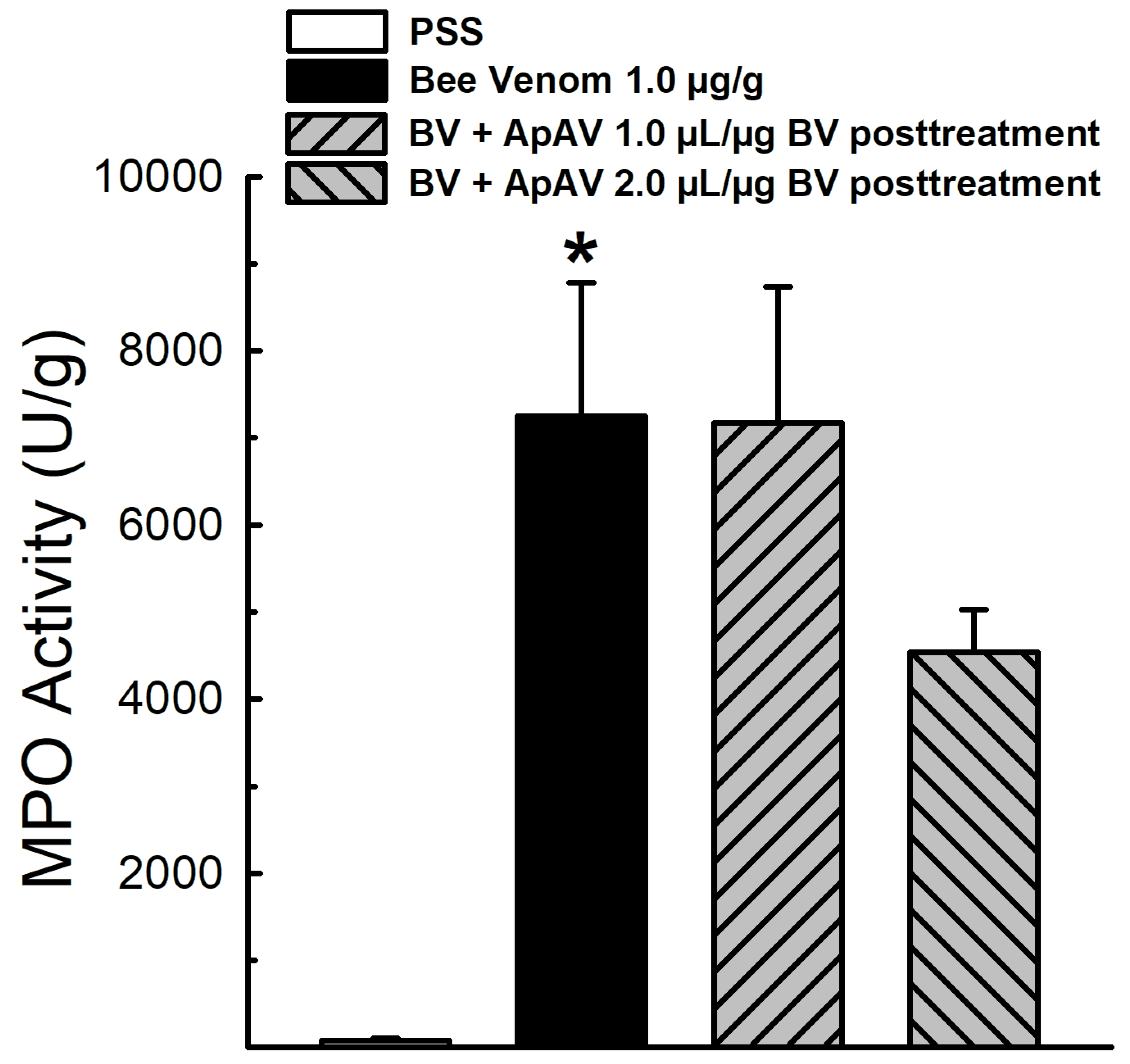
2.4. ApAV Inhibition of In Vivo Myeloperoxidase Activity Induced by Africanized A. mellifera Venom
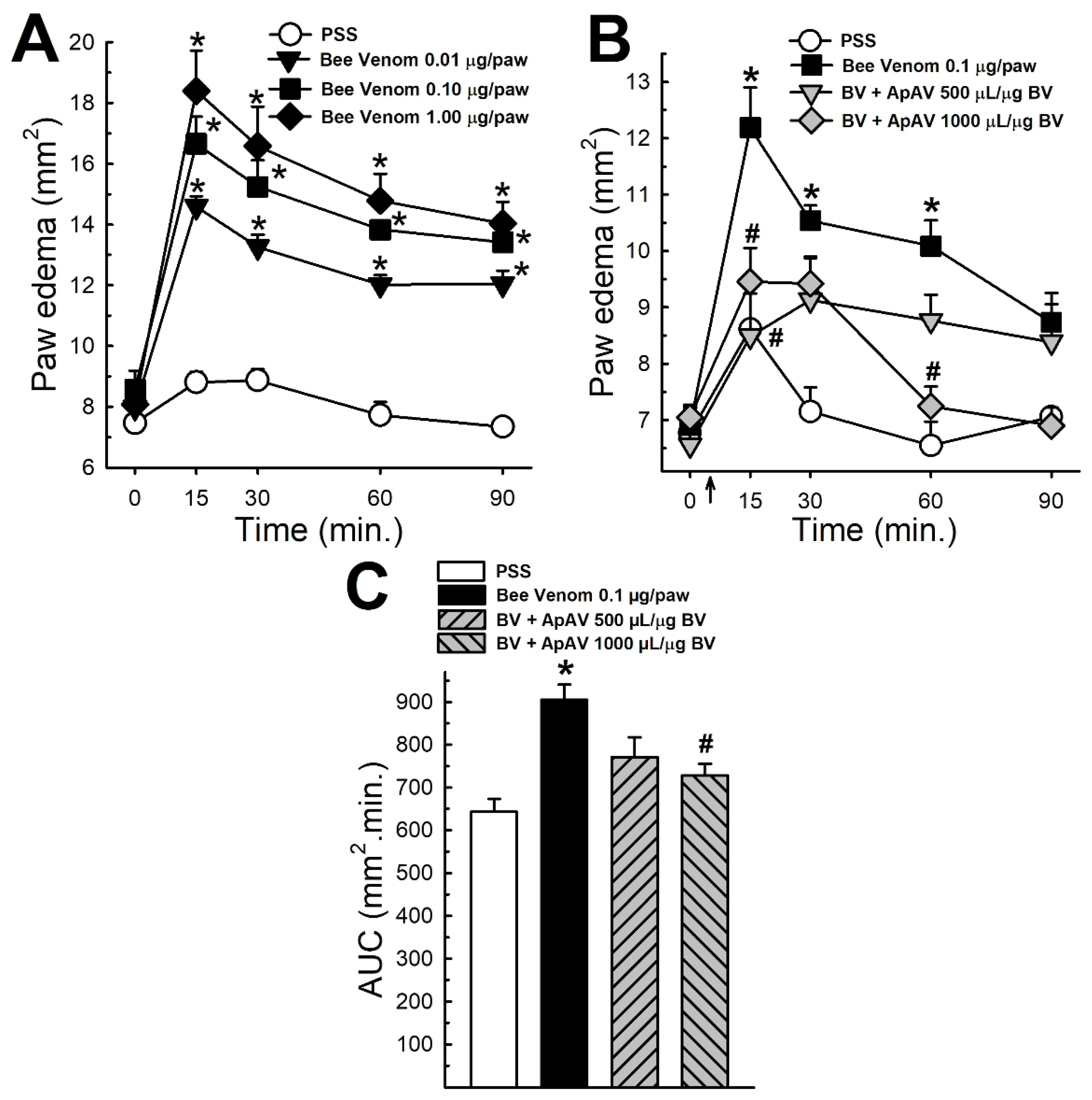
2.5. ApAV Inhibited Paw Edema Induced by Africanized A. mellifera Venom
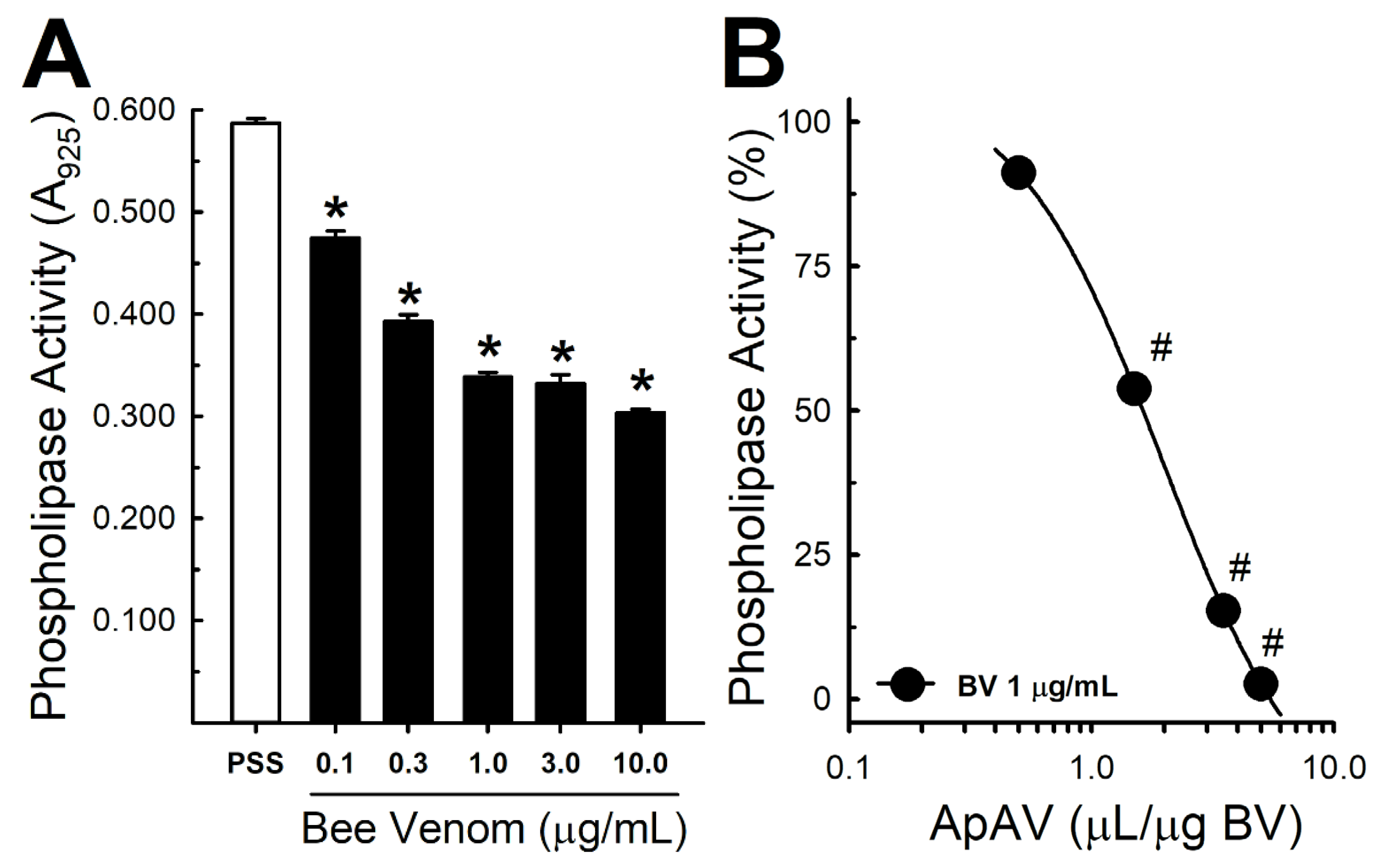
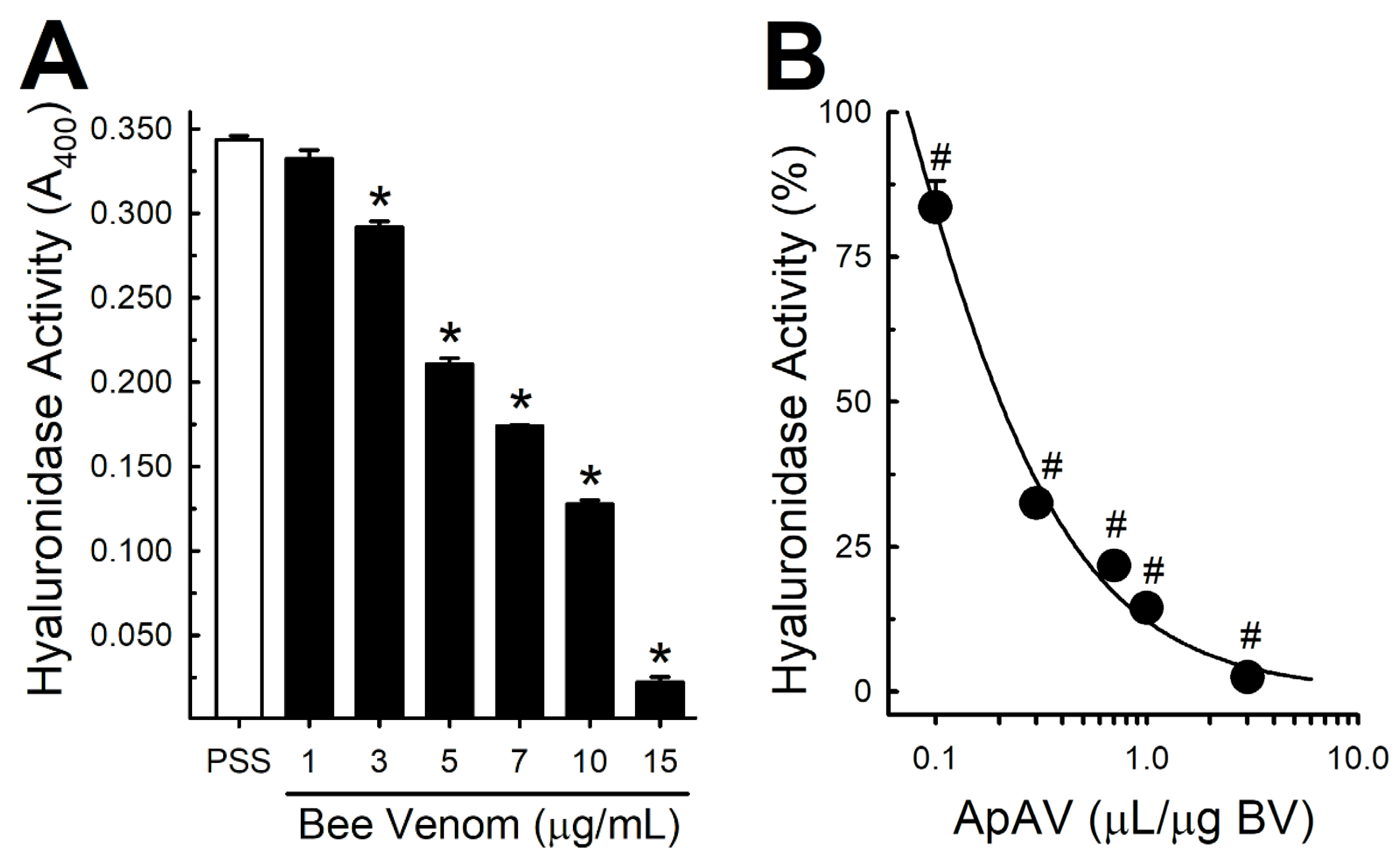
2.6. ApAV Inhibited Phospholipase and Hyaluronidase Activity of Africanized A. mellifera Venom
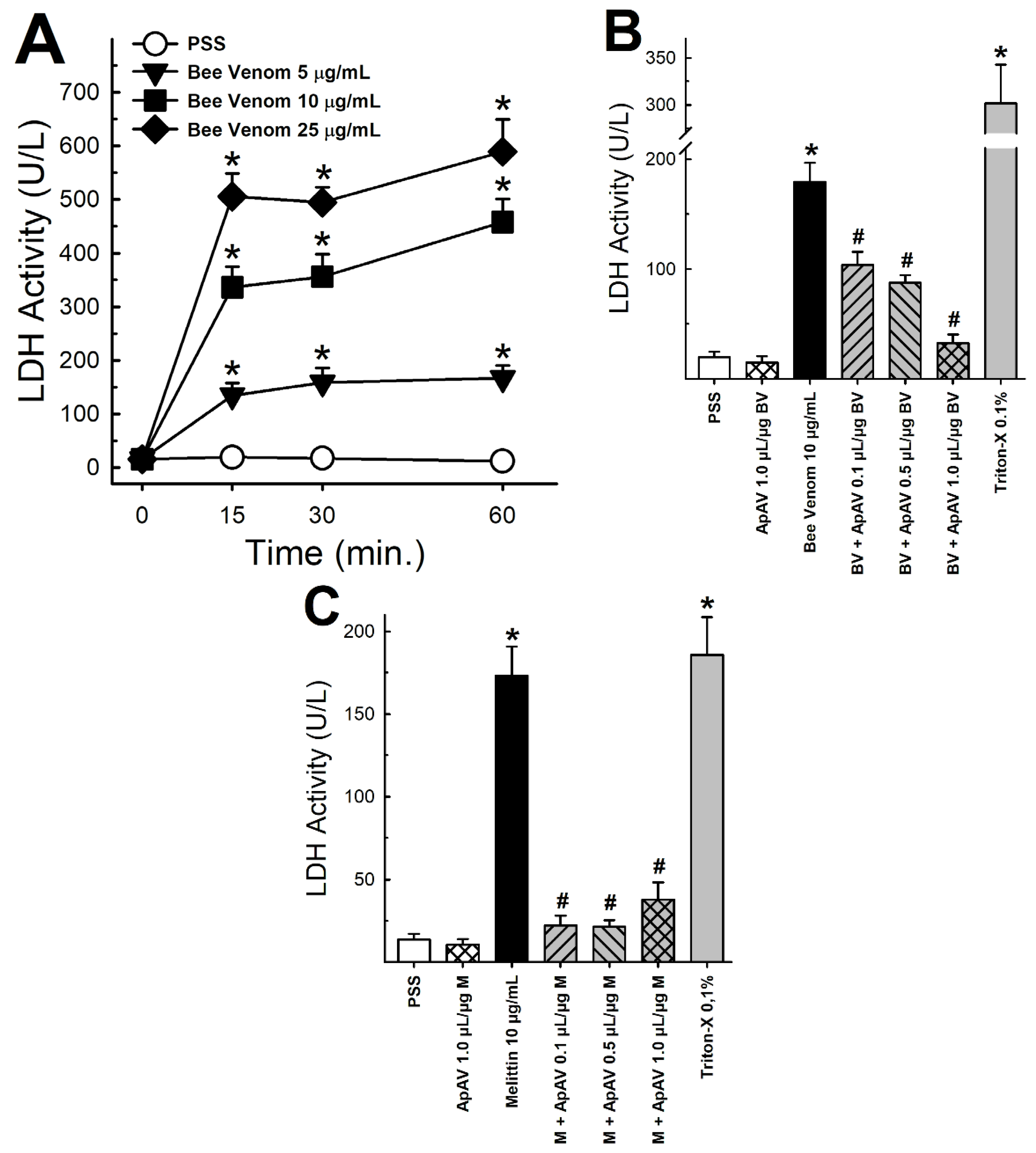
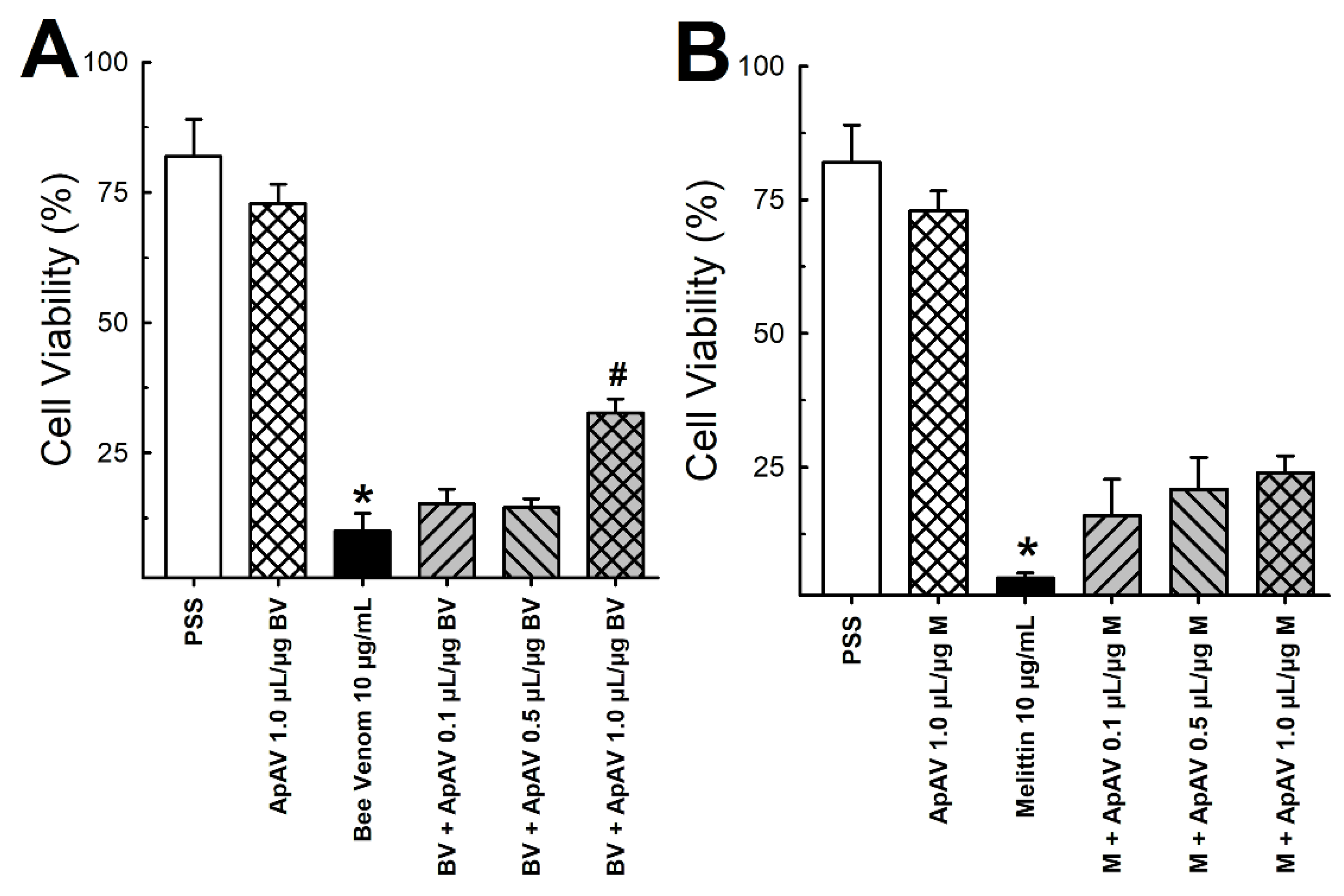
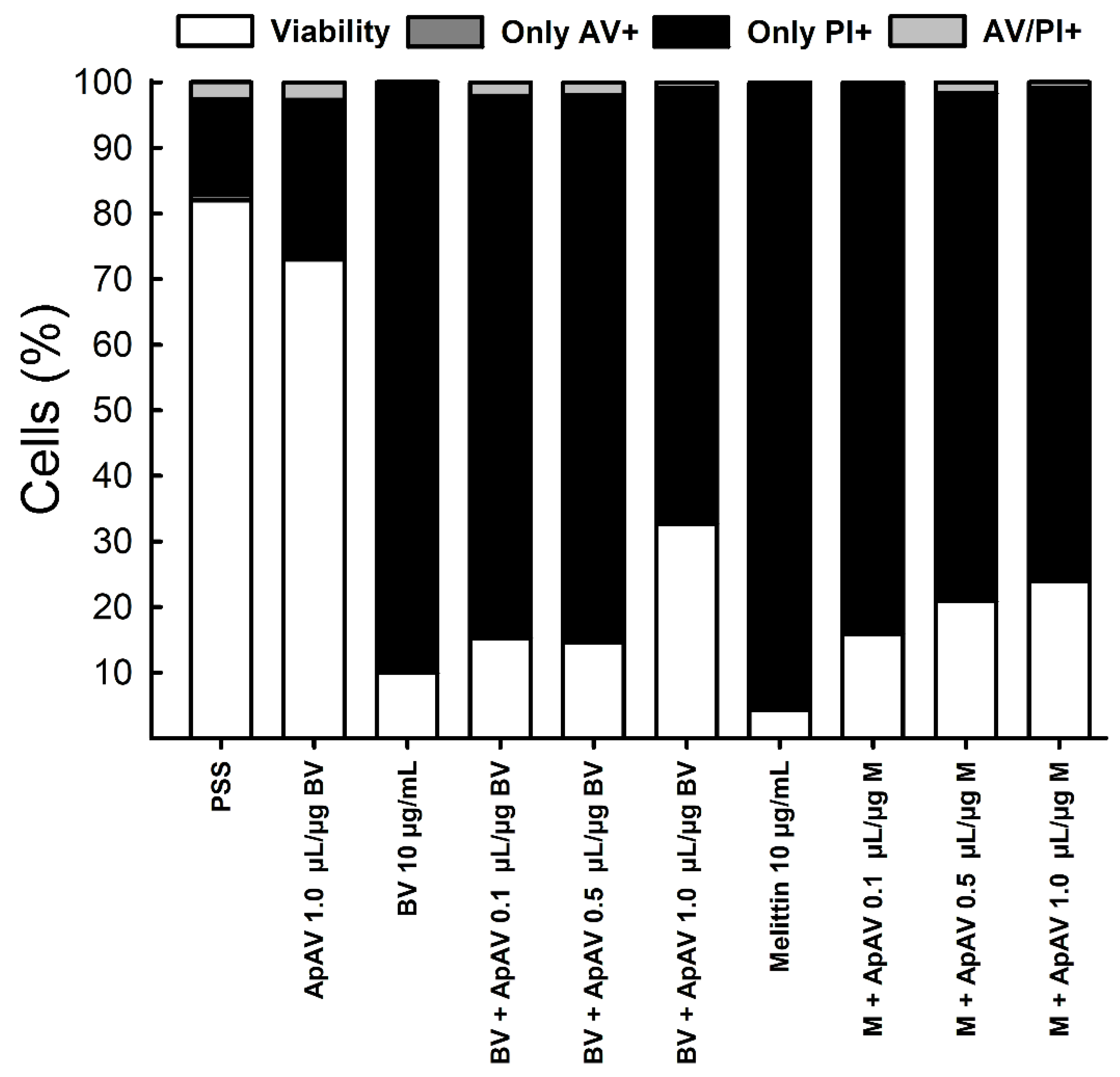
2.7. ApAV Inhibited the Cytotoxic Activity of Africanized A. mellifera Venom and Melittin
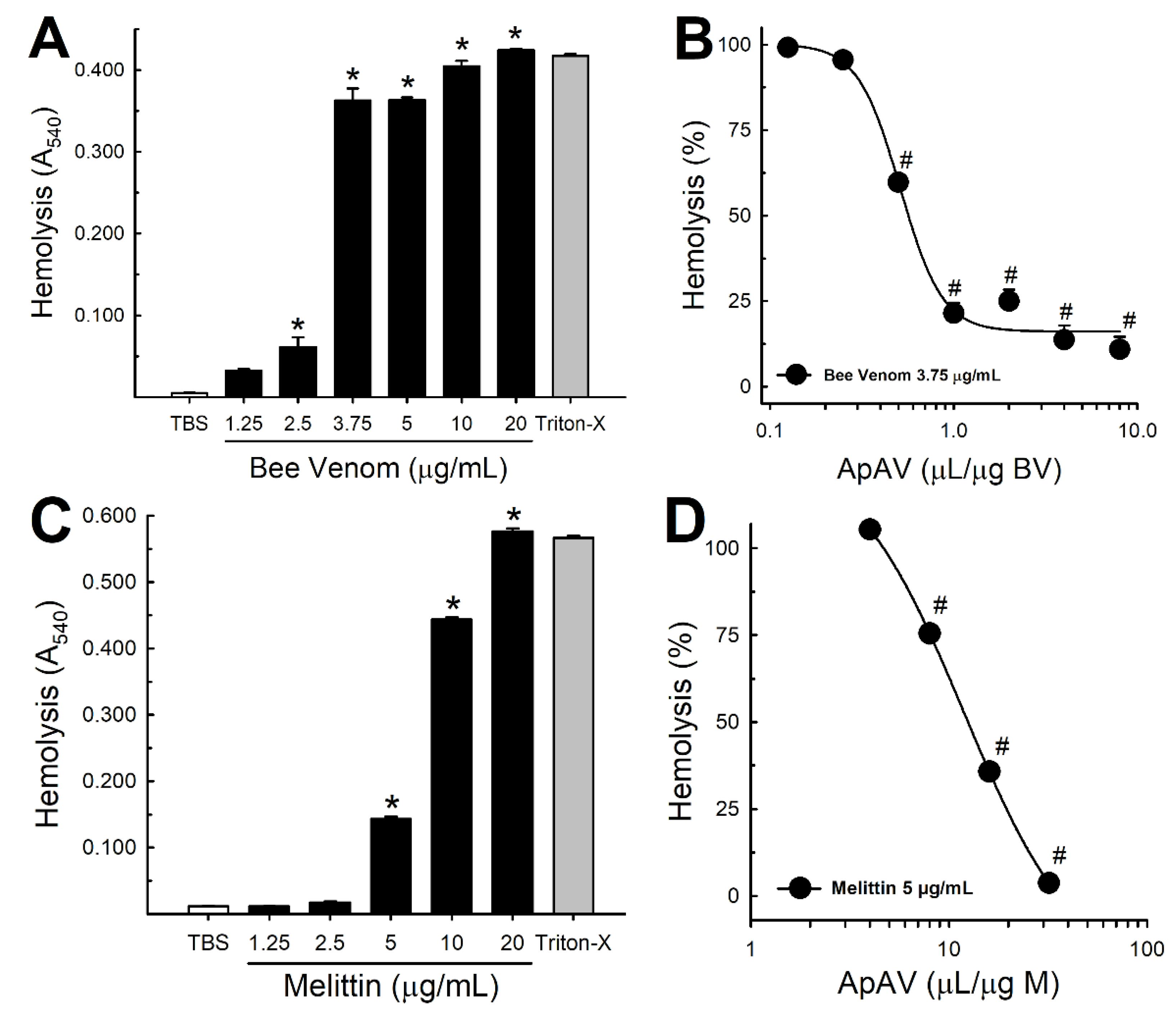
2.8. ApAV Inhibited the Hemolytic Activity of the Africanized A. mellifera Venom and Melittin
3. Discussion
4. Materials and Methods
4.1. Animals and Reagents
4.2. Cell Line
4.3. Lethality
4.4. Myotoxicity In Vivo and Hematocrit
4.5. Vascular Permeability
4.6. Myeloperoxidase Activity
4.7. Paw Edema
4.8. Honeybee Venom Phospholipase Activity
4.9. Honeybee Venom Hyaluronidase Activity
4.10. LDH Assay
4.11. Cell Death Analysis
4.12. Hemolytic Activity
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghazoul, J. Buzziness as usual? Questioning the global pollination crisis. Trends Ecol. Evol. 2005, 20, 367–373. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Boil. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Tarpy, D.R.; Delaney, D.A.; Seeley, T.D. Mating frequencies of honey bee queens (Apis mellifera L.) in a population of feral colonies in the Northeastern United States. PLoS ONE 2015, 10, e0118734. [Google Scholar] [CrossRef]
- Kadri, S.M.; Harpur, B.A.; Orsi, R.O.; Zayed, A. A variant reference data set for the Africanized honeybee, Apis mellifera. Sci. Data 2016, 3, 160097. [Google Scholar] [CrossRef]
- De Oliveira, G.P.; Kadri, S.M.; Benaglia, B.G.E.; Ribolla, P.E.M.; Orsi, R.D.O. [Unesp] Energetic supplementation for maintenance or development of Apis mellifera L. colonies. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200004. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.S., Jr.; Almeida, R.A.M.B.; Barraviera, S.R.C.S.; Barraviera, B. Historical perspective and human consequences of Africanized bee stings in the Americas. J. Toxicol. Environ. Heal. Part. B 2012, 15, 97–108. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture, Agricultural Research Service. Africanized Honey Bees. Available online: https://www.ars.usda.gov/pacific-west-area/tucson-az/honey-bee-research/docs/africanized-honey-bees/ (accessed on 2 March 2020).
- Pucca, M.B.; Cerni, F.A.; Oliveira, I.S.; Jenkins, T.P.; Argemí, L.; Sørensen, C.V.; Ahmadi, S.; Barbosa, J.E.; Laustsen, A.H. Bee updated: Current knowledge on bee venom and bee envenoming therapy. Front. Immunol. 2019, 10, 2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gummin, D.D.; Mowry, J.B.; Spyker, D.A.; Brooks, D.E.; Osterthaler, K.M.; Banner, W. Annual report of the American Association of Poison Control Centers’ National Poison Data System (NPDS): 35th anual report. Clin. Toxicol. 2018, 56, 1213–1415. [Google Scholar] [CrossRef] [PubMed]
- QuickStats: Number of deaths from hornet, wasp, and bee stings, among males and females-National Vital Statistics System, United States, 2000–2017. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 649. [CrossRef] [Green Version]
- Pinto, M.A.; Rubink, W.L.; Patton, J.C.; Coulson, R.N.; Johnston, J.S. Africanization in the United States. Genetics 2005, 170, 1653–1665. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, M.J.; Schmidt, J.O.; Egen, N.B. Lethality of ’killer’ bee stings. Nat. Cell Biol. 1989, 337, 413. [Google Scholar] [CrossRef]
- França, F.; Benvenuti, L.A.; Wen, F.H.; Dos Santos, D.R.; Hain, S.H.; Picchi-Martins, F.R.; Cardoso, J.L.; Kamiguti, A.S.; Theakston, R.D.G.; Warrell, D.A. Severe and fatal mass attacks by ‘killer’ bees (Africanized honey bees-Apis mellifera scutellata) in Brazil: Clinicopathological studies with measurement of serum venom concentrations. Qjm: Int. J. Med. 1994, 87, 269–282. [Google Scholar] [CrossRef]
- Barbosa, A.; Leslie, B.; Chippaux, J.-P.; Medolago, N.B.; Caramori, C.A.; Paixão, A.G.; Poli, J.P.V.; Mendes, M.; Santos, L.; Ferreira, R.S., Jr.; et al. A clinical trial protocol to treat massive Africanized honeybee (Apis mellifera) attack with a new Apilic antivenom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochner, R.; De Souza, C.M.V. Divergences between the Brazilian national information systems for recording deaths from venomous animals. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e143018. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, S.K.; Trevisol, D.J.; Parma, G.C.; Júnior, R.S.F.; Barbosa, A.N.; Barraviera, B.; Schuelter-Trevisol, F. Honey bee envenoming in Santa Catarina, Brazil, 2007 through 2017: An observational, retrospective cohort study. Rev. Soc. Bras. Med. Trop. 2019, 52, e20180418. [Google Scholar] [CrossRef] [PubMed]
- Brazilian Ministry of Health. Sistema de Informação de Agravos de Notificação-SINAN/SUS. 2018. Available online: https://portalarquivos2.saude.gov.br/images/pdf/2019/junho/05/04-Dados-Epidemiologicos-SiteSVS--maio-2019---ABELHAS-CASOS.pdf (accessed on 28 January 2018).
- Hider, R.C. Honeybee venom: A rich source of pharmacologically active peptides. Endeavour 1988, 12, 60–65. [Google Scholar] [CrossRef]
- Soman, N.R.; Baldwin, S.L.; Hu, G.; Marsh, J.N.; Lanza, G.M.; Heuser, J.E.; Arbeit, J.M.; Wickline, S.A.; Schlesinger, P.H. Molecularly targeted nanocarriers deliver the cytolytic peptide melittin specifically to tumor cells in mice, reducing tumor growth. J. Clin. Investig. 2009, 119, 2830–2842. [Google Scholar] [CrossRef]
- Ewan, P.W. ABC of allergies: Venom allergy. BMJ 1998, 316, 1365–1368. [Google Scholar] [CrossRef]
- Warrell, D.A. Venomous bites, stings, and poisoning. Infect. Dis. Clin. North. Am. 2019, 33, 17–38. [Google Scholar] [CrossRef]
- Vetter, R.S.; Visscher, P.K.; Camazine, S. Mass envenomations by honey bees and wasps. West. J. Med. 1999, 170, 223–227. [Google Scholar]
- Hughes, R.L. A fatal case of acute renal failure from envenoming syndrome after massive bee attack. Am. J. Forensic Med. Pathol. 2019, 40, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Prado, M.; Solano-Trejos, G.; Lomonte, B. Acute physiopathological effects of honeybee (Apis mellifera) envenoming by subcutaneous route in a mouse model. Toxicon 2010, 56, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Rayamane, A.P.; Kumar, M.P.; Kishor, D.G.; Dayananda, R.; Saraf, A. Honey bee stings and anaphylaxis: Review. J. Forensic Med. Sci. Law 2014, 23, 1–7. [Google Scholar]
- Fundação Nacional de Saúde (FUNASA). Manual de Diagnóstico e Tratamento de Acidentes por Animais Peçonhentos, 2nd ed.; Fundação Nacional de Saúde: Brasilia, Brazil, 2001. [Google Scholar]
- Ferreira, R.S., Jr.; Barraviera, B.; Orsi, R.O.; Cunha, L.E.R.; Pimenta, D.C. Process of Obtaining AntiApilic Equine Serum and Its Uses; Registration number: BR10201502630; INPI-National Institute of Industrial Property: Brasilia, Brazil, 2015; PCT: 10201502630. [Google Scholar]
- El-Kik, C.Z.; Fernandes, F.F.; Tomaz, M.A.; Gaban, G.A.; Fonseca, T.F.; Calil-Eliás, S.; Oliveira, S.D.S.; Silva, C.L.M.; Martinez, A.M.; Melo, P. Neutralization of Apis mellifera bee venom activities by suramin. Toxicon 2013, 67, 55–62. [Google Scholar] [CrossRef]
- WHO. Guidelines for the Production, Control. and Regulation of Snake Antivenom Immunoglobulins; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Pessenda, G.; Silva, L.C.E.; Campos, L.B.; Pacello, E.M.; Pucca, M.B.; Martinez, E.Z.; Barbosa, J.E. Human scFv antibodies (Afribumabs) against Africanized bee venom: Advances in melittin recognition. Toxicon 2016, 112, 59–67. [Google Scholar] [CrossRef]
- Petricevich, V.L. Cytokine and nitric oxide production following severe envenomation. Curr. Drug Target. -Inflamm. Allergy 2004, 3, 325–332. [Google Scholar] [CrossRef]
- Quesada, L.; Sevcik, C.; Lomonte, B.; Rojas, E.; Gutiérrez, J.M. Pharmacokinetics of whole IgG equine antivenom: Comparison between normal and envenomed rabbits. Toxicon 2006, 48, 255–263. [Google Scholar] [CrossRef]
- Smith, C.; Kruger, M.J.; Smith, R.M.; Myburgh, K.H. The inflammatory response to skeletal muscle injury. Sports Med. 2008, 38, 947–969. [Google Scholar] [CrossRef]
- Patrão-Neto, F.C.; Tomaz, M.A.; Strauch, M.A.; Monteiro-Machado, M.; Rocha-Junior, J.R.D.S.; Borges, P.A.; Calil-Elias, S.; Melo, P. Dexamethasone antagonizes the in vivo myotoxic and inflammatory effects of Bothrops venoms. Toxicon 2013, 69, 55–64. [Google Scholar] [CrossRef]
- Monteiro-Machado, M.; Tomaz, M.A.; Fonseca, R.J.; Strauch, M.A.; Cons, B.L.; Borges, P.A.; Patrão-Neto, F.C.; Tavares-Henriques, M.S.; Teixeira-Cruz, J.M.; Calil-Eliás, S.; et al. Occurrence of sulfated fucose branches in fucosylated chondroitin sulfate are essential for the polysaccharide effect preventing muscle damage induced by toxins and crude venom from Bothrops jararacussu snake. Toxicon 2015, 98, 20–33. [Google Scholar] [CrossRef]
- Grisotto, L.S.; Mendes, G.E.; Castro, I.; Baptista, M.A.; Alves, V.A.; Yu, L.; Burdmann, E.A. Mechanisms of bee venom-induced acute renal failure. Toxicon 2006, 48, 44–54. [Google Scholar] [CrossRef]
- Funayama, J.C.; Pucca, M.B.; Roncolato, E.C.; Bertolini, T.B.; Campos, L.B.; Barbosa, J.E. Production of human antibody fragments binding to melittin and phospholipase A2 in Africanised bee venom: Minimising venom toxicity. Basic Clin. Pharmacol. Toxicol. 2011, 110, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Tomaz, M.A.; Fernandes, F.F.; El-Kik, C.Z.; Moraes, R.A.; Calil-Eliás, S.; Oliveira, J.; Martínez, A.M.B.; Ownby, C.L.; Melo, P. Increase of the cytotoxic effect of Bothrops jararacussu venom on mouse extensor digitorum longus and soleus by potassium channel blockers and by Na+/K+-ATPase inhibition. Toxicon 2008, 52, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, M.A.; Costa, R.S.; Coimbra, T.M.; Teixeira, V.P. Acute renal failure in experimental envenomation with Africanized bee venom. Ren. Fail. 1998, 20, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Han, H.J.; Lee, J.H.; Park, S.H.; Choi, H.J.; Yang, I.S.; Mar, W.C.; Kang, S.K.; Lee, H.J. Effect of bee venom and its melittin on apical transporters of renal proximal tubule cells. Kidney Blood Press. Res. 2000, 23, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Mollay, C.; Kreil, G. Enhancement of bee venom phospholipase A2 activity by melittin, direct lytic factor from cobra venom and polymyxin B. FEBS Lett. 1974, 46, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Girish, K.S.; Kemparaju, K. Overlooked issues of snakebite management: Time for strategic approach. Curr. Top. Med. Chem. 2011, 11, 2494–2508. [Google Scholar] [CrossRef]
- Marković-Housley, Z.; Miglierini, G.; Soldatova, L.; Rizkallah, P.J.; Müller, U.; Schirmer, T. Crystal structure of hyaluronidase, a major allergen of bee venom. Structure 2000, 8, 1025–1035. [Google Scholar] [CrossRef]
- Amaral, L.S.; Ferreira, J.M.; Predes, D.; Abreu, J.G.; Noël, F.; Quintas, L.E.M. Telocinobufagin and Marinobufagin produce different effects in LLC-PK1 cells: A case of functional selectivity of bufadienolides. Int. J. Mol. Sci. 2018, 19, 2769. [Google Scholar] [CrossRef] [Green Version]
- Melo, P.A.; Ownby, C.L. Ability of wedelolactone, heparin, and para-bromophenacyl bromide to antagonize the myotoxic effects of two crotaline venoms and their PLA2 myotoxins. Toxicon 1999, 37, 199–215. [Google Scholar] [CrossRef]
- Calil-Elias, S.; Martinez, A.M.B.; Melo, P.A. Effect of heparin and antivenom on skeletal muscle damage produced by Bothrops jararacussu venom. Histol. Histopathol. 2002, 17, 463–470. [Google Scholar] [PubMed]
- Calil-Elias, S.; Thattassery, E.; Martinez, A.; Melo, P. Effect of perimuscular injection of Bothrops jararacussu venom on plasma creatine kinase levels in mice: Influence of dose and volume. Braz. J. Med. Biol. Res. 2002, 35, 1233–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayres, R.; Feijó, P.R.; Cintra, A.C.; Tomaz, M.A.; Melo, P.; Cunha, V.M.; Quintas, L.E.M. Different effects of myotoxins bothropstoxin-I and II from Bothrops snake venom on cation transport ATPases from murine fast twitch skeletal muscle. Toxicon 2015, 103, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Melo, P.A.; Suarez-Kurtz, G. Release of creatine kinase from skeletal muscles by Bothrops venoms: Heparin potentiation of inhibition by antivenin. Braz. J. Med. Biol. Res. 1988, 21, 545–548. [Google Scholar] [PubMed]
- Melo, P.A.; Homsi-Brandeburgo, M.I.; Giglio, J.R.; Suarez-Kurtz, G. Antagonism of the myotoxic effects of Bothrops jararacussu venom and bothropstoxin by polyanions. Toxicon 1993, 31, 285–291. [Google Scholar] [CrossRef]
- Melo, P.A.; Nascimento, M.C.D.; Mors, W.B.; Suarez-Kurtz, G. Inhibition of the myotoxic and hemorrhagic activities of crotalid venoms by Eclipta prostrata (Asteraceae) extracts and constituents. Toxicon 1994, 32, 595–603. [Google Scholar] [CrossRef]
- Posadas, I.; Bucci, M.; Roviezzo, F.; Rossi, A.; Parente, L.; Sautebin, L.; Cirino, G. Carrageenan-induced mouse paw oedema is biphasic, age-weight dependent and displays differential nitric oxide cyclooxygenase-2 expression. Br. J. Pharmacol. 2004, 142, 331–338. [Google Scholar] [CrossRef]
- Marinetti, G. The action of phospholipase A on lipoproteins. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 2003, 98, 554–565. [Google Scholar] [CrossRef]
- Di Ferrante, N. Turbidimetric measurement of acid mucopolysaccharides and hyaluronidase activity. J. Biol. Chem. 1956, 220, 303–306. [Google Scholar] [PubMed]
- Moraes, J.; Frony, A.C.; Barcellos-De-Souza, P.; Da Cunha, M.M.; Calcia, T.B.B.; Benjamim, C.F.; Boisson-Vidal, C.; Barja-Fidalgo, C. Downregulation of microparticle release and pro-inflammatory properties of activated human polymorphonuclear neutrophils by LMW fucoidan. J. Innate Immun. 2018, 11, 330–346. [Google Scholar] [CrossRef]
- Fischer, D.; Li, Y.; Ahlemeyer, B.; Krieglstein, J.; Kissel, T. In vitro cytotoxicity testing of polycations: Influence of polymer structure on cell viability and hemolysis. Biomaterials 2003, 24, 1121–1131. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira-Cruz, J.M.; Strauch, M.A.; Monteiro-Machado, M.; Tavares-Henriques, M.S.; de Moraes, J.A.; Ribeiro da Cunha, L.E.; Ferreira, R.S., Jr.; Barraviera, B.; Quintas, L.E.M.; Melo, P.A. A Novel Apilic Antivenom to Treat Massive, Africanized Honeybee Attacks: A Preclinical Study from the Lethality to Some Biochemical and Pharmacological Activities Neutralization. Toxins 2021, 13, 30. https://doi.org/10.3390/toxins13010030
Teixeira-Cruz JM, Strauch MA, Monteiro-Machado M, Tavares-Henriques MS, de Moraes JA, Ribeiro da Cunha LE, Ferreira RS Jr., Barraviera B, Quintas LEM, Melo PA. A Novel Apilic Antivenom to Treat Massive, Africanized Honeybee Attacks: A Preclinical Study from the Lethality to Some Biochemical and Pharmacological Activities Neutralization. Toxins. 2021; 13(1):30. https://doi.org/10.3390/toxins13010030
Chicago/Turabian StyleTeixeira-Cruz, Jhonatha Mota, Marcelo Abrahão Strauch, Marcos Monteiro-Machado, Matheus Silva Tavares-Henriques, João Alfredo de Moraes, Luís Eduardo Ribeiro da Cunha, Rui Seabra Ferreira, Jr., Benedito Barraviera, Luis Eduardo M. Quintas, and Paulo A. Melo. 2021. "A Novel Apilic Antivenom to Treat Massive, Africanized Honeybee Attacks: A Preclinical Study from the Lethality to Some Biochemical and Pharmacological Activities Neutralization" Toxins 13, no. 1: 30. https://doi.org/10.3390/toxins13010030
APA StyleTeixeira-Cruz, J. M., Strauch, M. A., Monteiro-Machado, M., Tavares-Henriques, M. S., de Moraes, J. A., Ribeiro da Cunha, L. E., Ferreira, R. S., Jr., Barraviera, B., Quintas, L. E. M., & Melo, P. A. (2021). A Novel Apilic Antivenom to Treat Massive, Africanized Honeybee Attacks: A Preclinical Study from the Lethality to Some Biochemical and Pharmacological Activities Neutralization. Toxins, 13(1), 30. https://doi.org/10.3390/toxins13010030