Antinociceptive and Anti-Inflammatory Effects of Recombinant Crotamine in Mouse Models of Pain

 , , , and
, , , and 
Abstract
:1. Introduction
2. Methods
2.1. Expression and Purification of Recombinant Crotamine from E. coli
2.2. Experimental Animals
2.3. Behavioral Tests for Antinociceptive Activity Assay of Recombinant Crotamine
2.3.1. The Hot-Plate Test
2.3.2. The Acetic-Acid-Induced Writhing Method
2.3.3. The Formalin Test: Systemic (Intraperitoneal) Effect of Recombinant Crotamine
2.3.4. The Formalin Test: Peripheral (Intraplantar) Effect of Recombinant Crotamine
2.4. Behavioral Tests for Anti-Inflammatory Activity Assay: Systemic (Intraperitoneal) and Peripheral (Intraplantar) Effect of Recombinant Crotamine in Formalin Test
2.5. Biochemical Assays
2.6. Behavioral Analysis: The Rotarod Test
2.7. Data and Statistical Analysis
3. Results
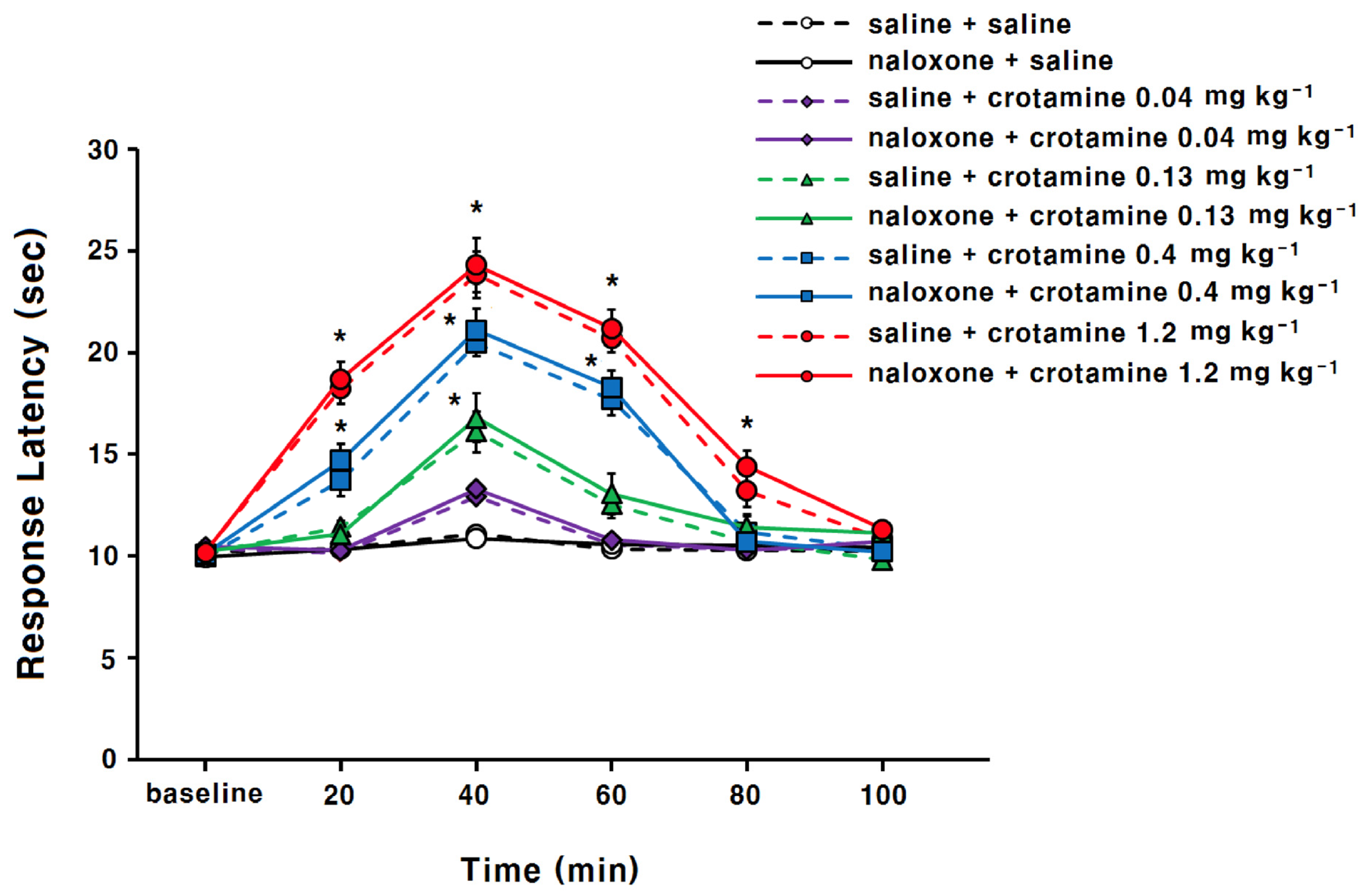
3.1. The Antinociceptive Effects of Systemic (Intraperitoneal) Recombinant Crotamine: The Hot-Plate Test
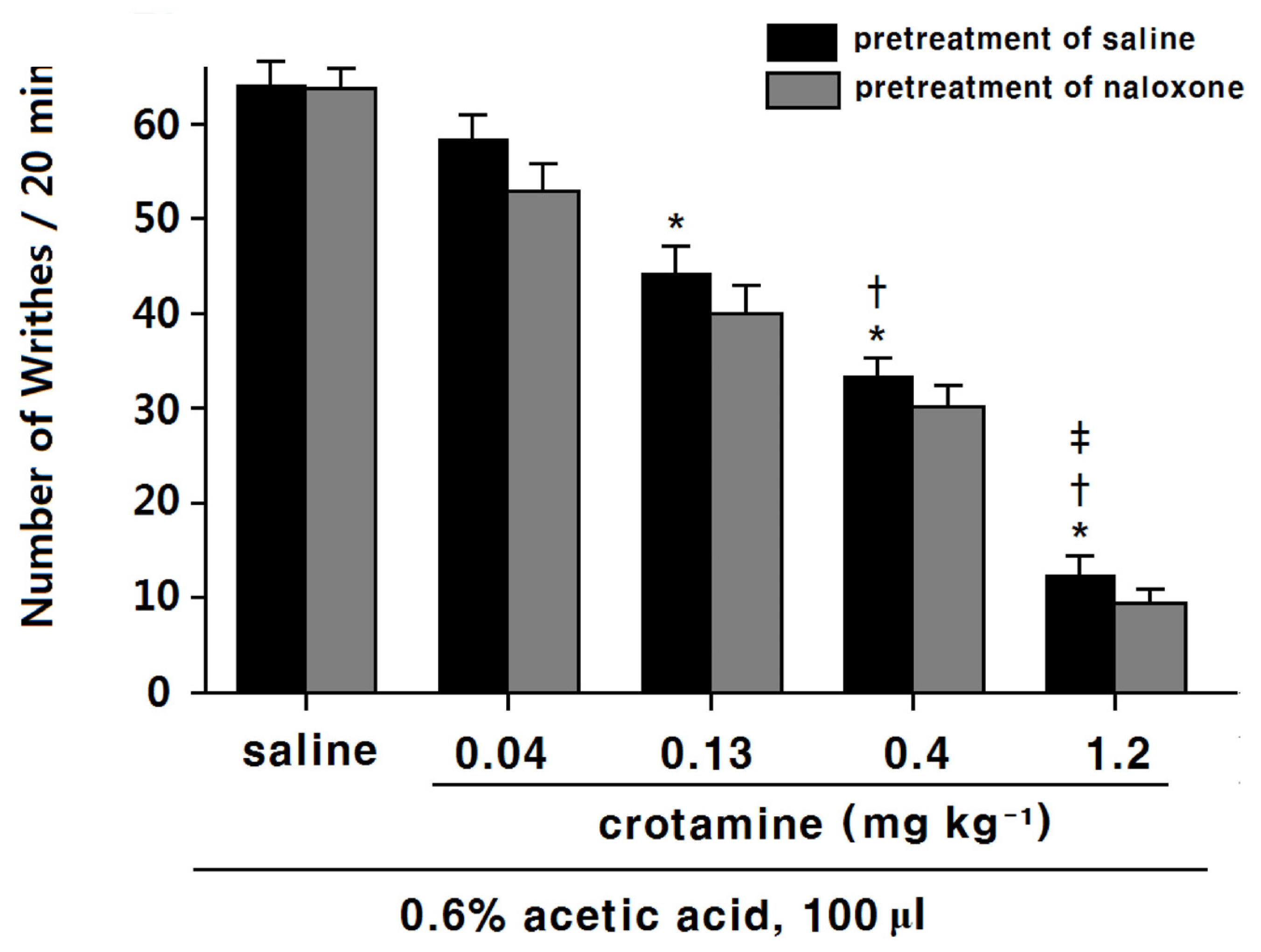
3.2. The Antinociceptive Effects of Systemic (Intraperitoneal) Recombinant Crotamine: Acetic-Acid-Induced Writhing Method
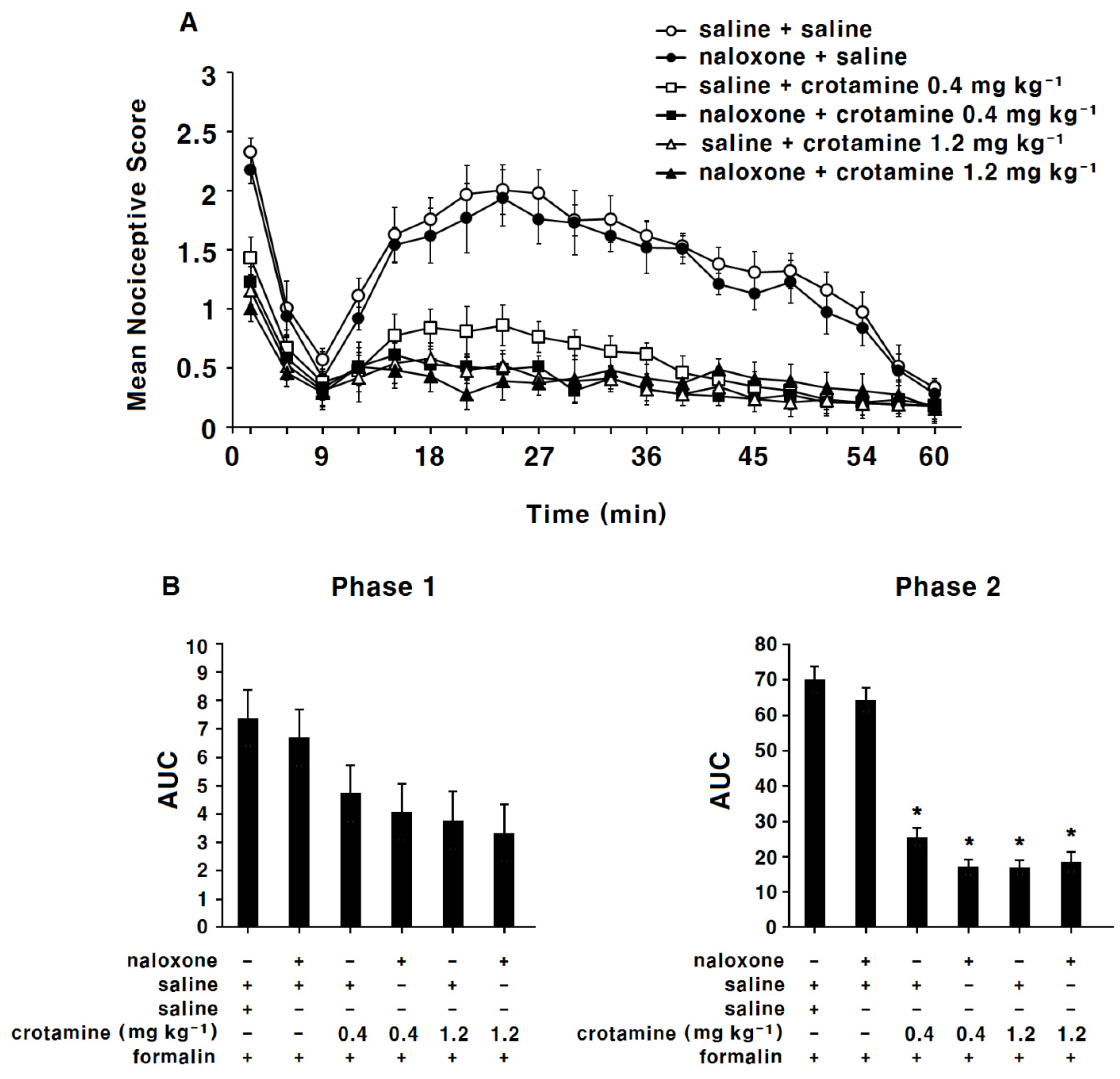
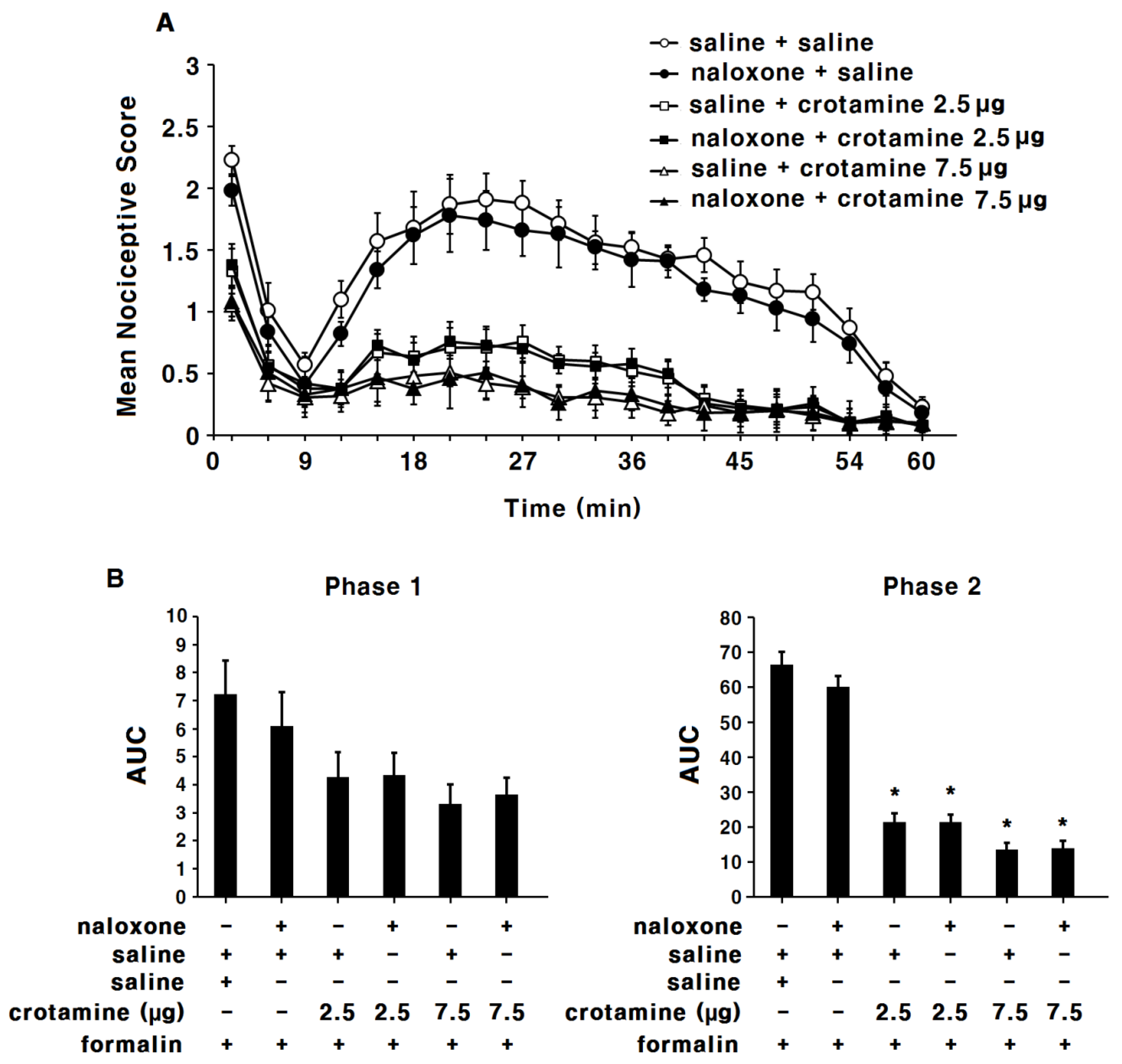
3.3. The Antinociceptive Effects of Systemic (Intraperitoneal) Recombinant Crotamine: The Formalin Test
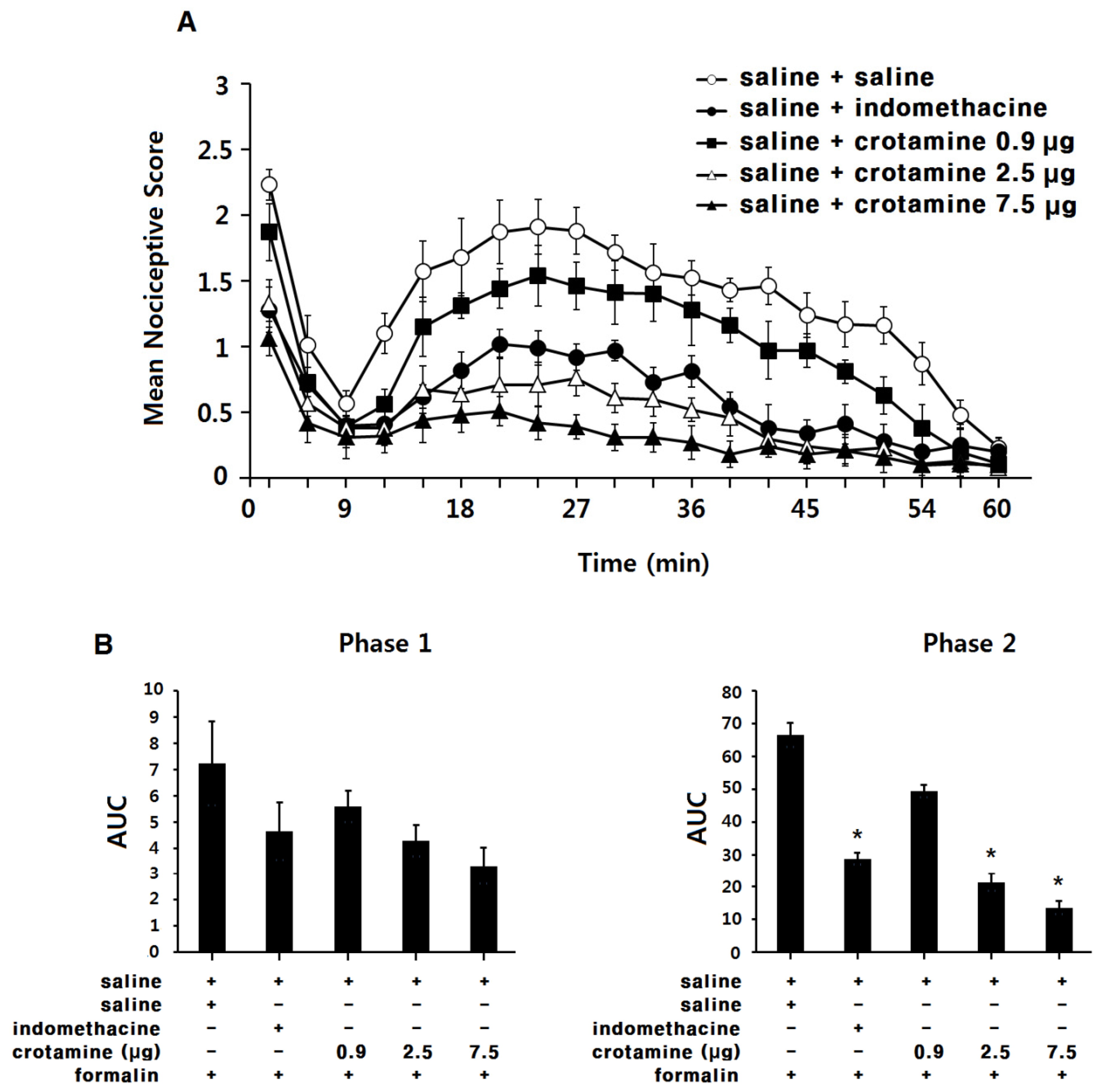
3.4. The Antinociceptive Effects of Peripheral (Intraplantar) Recombinant Crotamine: The Formalin Test
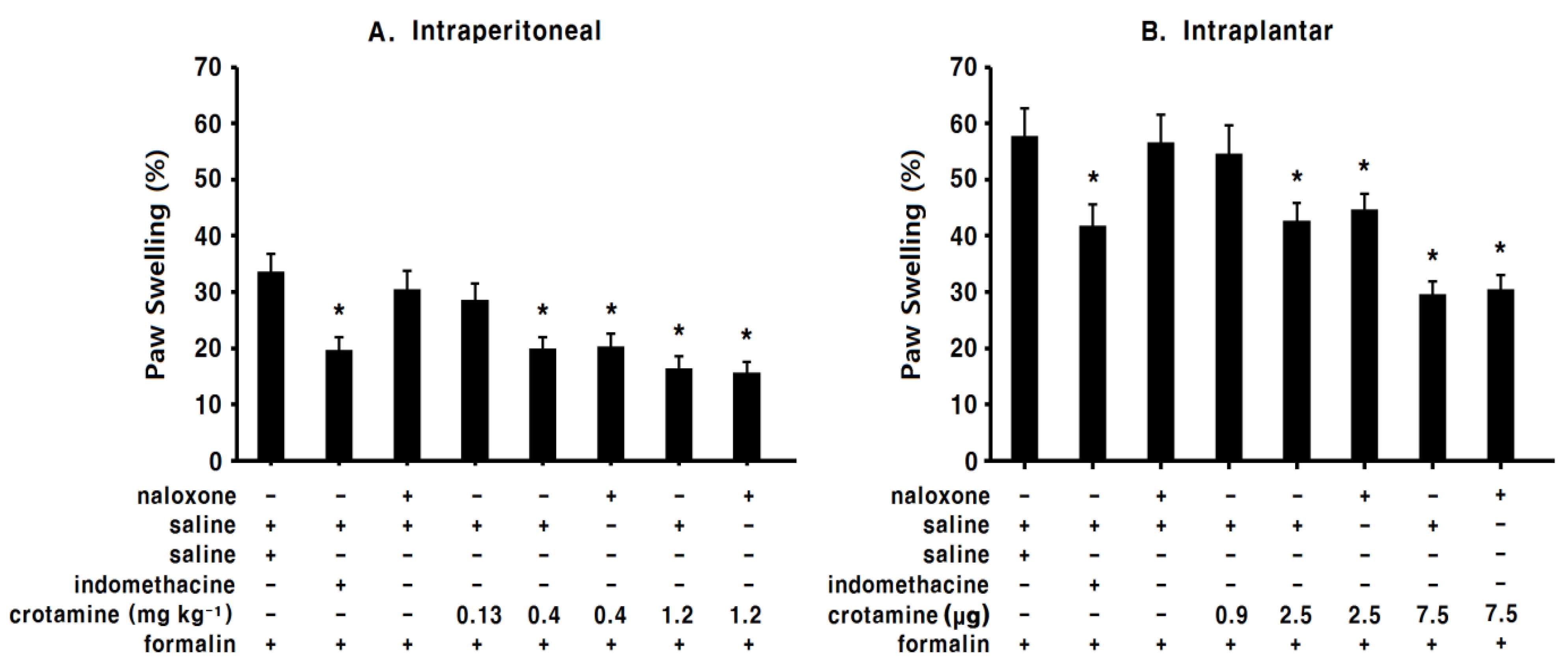
3.5. The Anti-Inflammatory Effects of Systemic (Intraperitoneal) Recombinant Crotamine: The Formalin Test
3.6. The Anti-Inflammatory Effects of Peripheral (Intraplantar) Recombinant Crotamine: The Formalin Test
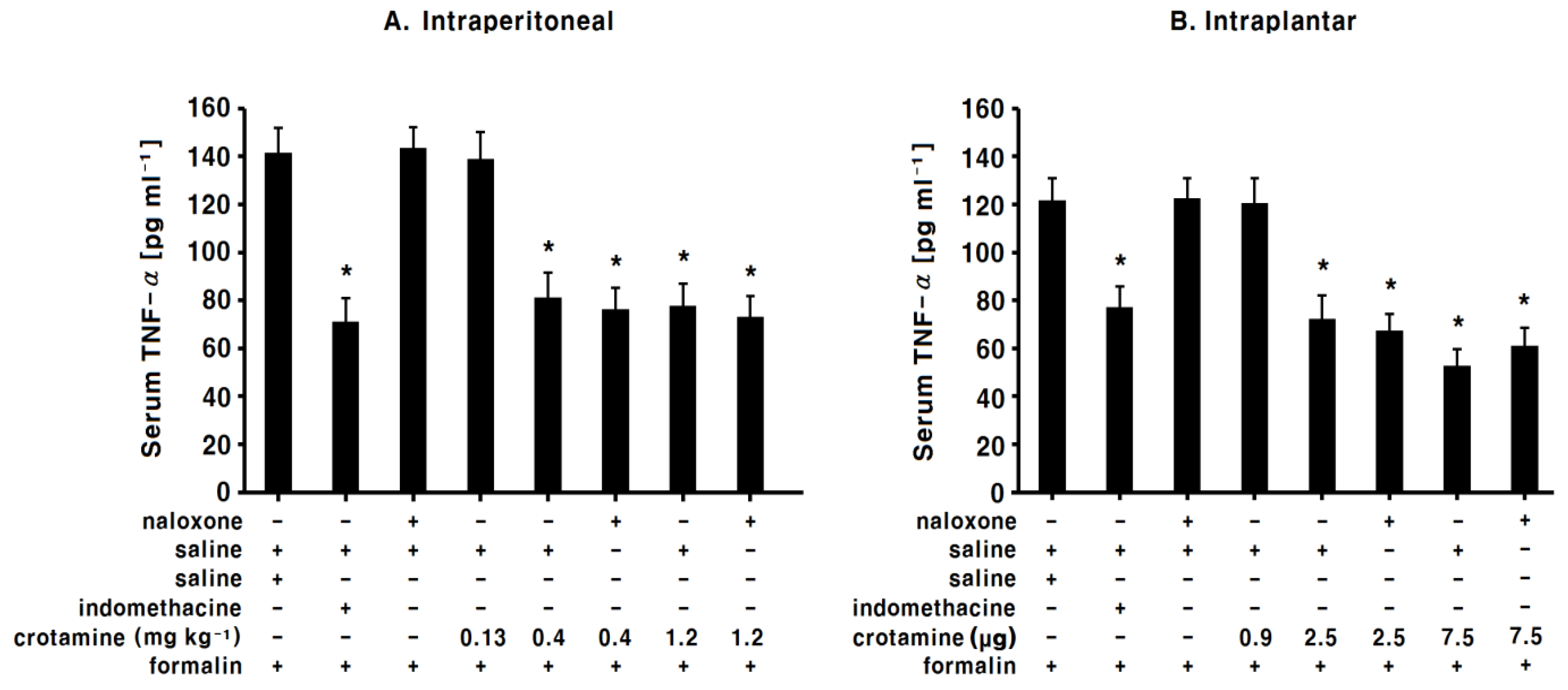
3.7. The Effects of Systemic (Intraperitoneal) Recombinant Crotamine on Serum TNF-α Levels
3.8. The Effects of Peripheral (Intraplantar) Recombinant Crotamine on Serum TNF-α levels
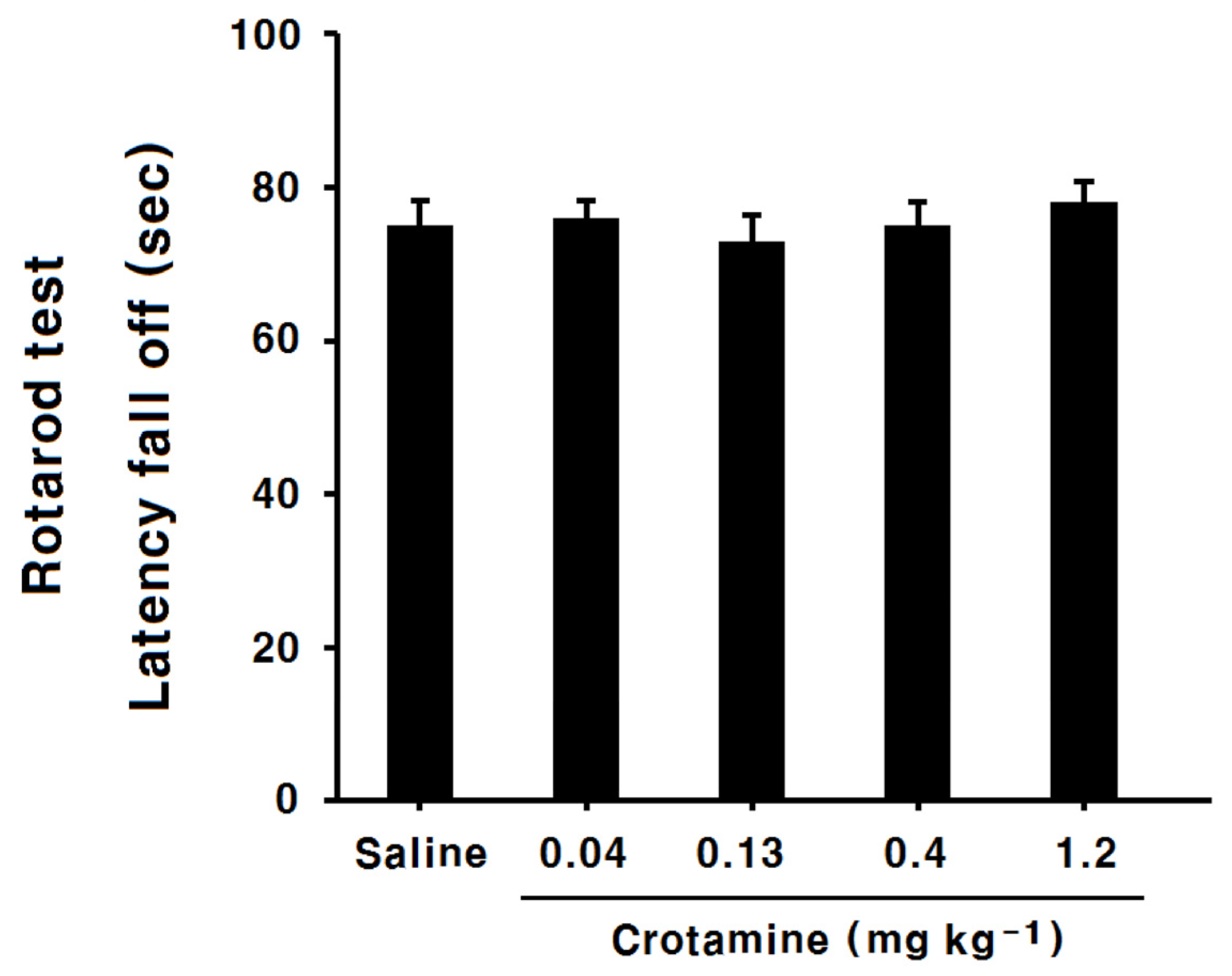
3.9. The Effects of Systemic (Intraperitoneal) Recombinant Crotamine on the Rotarod Test
4. Discussion
4.1. The Antinociceptive Effects of Systemic (Intraperitoneal) Recombinant Crotamine: The Hot-Plate Test and the Acetic-Acid-Induced Writhing Method
4.2. The Antinociceptive Effects of Systemic (Intraperitoneal) or Peripheral (Intraplantar) Recombinant Crotamine: The Formalin Test
4.3. The Anti-Inflammatory Effects of Systemic (Intraperitoneal) or Peripheral (Intraplantar) Recombinant Crotamine: The Formalin Test
4.4. The Effects of Systemic (Intraperitoneal) and Peripheral (Intraplantar) Recombinant Crotamine on Serum TNF-α Levels
4.5. Recombinant Crotamine and Opioid Receptors
4.6. Limitations of This Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, R.Z.; Robinson, S.E. The effect of cholinergic manipulations on the analgesic response to cobrotoxin in mice. Life Sci. 1990, 47, 1949–1954. [Google Scholar] [CrossRef]
- Pu, X.C.; Wong, P.T.; Gopalakrishnakone, P. A novel analgesic toxin (hannalgesin) from the venom of king cobra (Ophiophagus hannah). Toxicon 1995, 33, 1425–1431. [Google Scholar] [CrossRef]
- Brazil, O.V.; Prado-Franceschi, J.; Laure, C.J. Repetitive muscle responses induced by crotamine. Toxicon 1979, 17, 61–67. [Google Scholar] [CrossRef]
- Melani, R.D.; Araujo, G.D.T.; Carvalho, P.C.; Goto, L.; Nogueira, F.C.S.; Junqueira, M.; Domont, G.B. Seeing beyond the tip of the iceberg: A deep analysis of the venome of the Brazilian Rattlesnake, Crotalus durissus terrificus. EuPA Open Proteom. 2015, 8, 144–156. [Google Scholar] [CrossRef] [Green Version]
- Fusco, L.S.; Neto, E.B.; Francisco, A.F.; Alfonso, J.; Soares, A.; Pimenta, D.C.; Leiva, L.C. Fast venomic analysis of Crotalus durissus terrificus from northeastern Argentina. Toxicon X 2020, 7, 100047. [Google Scholar] [CrossRef] [PubMed]
- Giglio, J.R. Analytical studies on crotamine hydrochloride. Anal. Biochem. 1975, 69, 207–221. [Google Scholar] [CrossRef]
- Laure, C.J. The primary structure of crotamine (author’s transl). Hoppe Seylers Z Physiol. Chem. 1975, 356, 213–215. [Google Scholar]
- Mancin, A.C.; Soares, A.M.; Andriao-Escarso, S.H.; Faca, V.M.; Greene, L.J.; Zuccolotto, S.; Pela, I.R.; Giglio, J.R. The analgesic activity of crotamine, a neurotoxin from Crotalus durissus terrificus (South American rattlesnake) venom: A biochemical and pharmacological study. Toxicon 1998, 36, 1927–1937. [Google Scholar] [CrossRef]
- Giorgi, R.; Bernardi, M.M.; Cury, Y. Analgesic effect evoked by low molecular weight substances extracted from Crotalus durissus terrificus venom. Toxicon 1993, 31, 1257–1265. [Google Scholar] [CrossRef]
- Hayashi, M.A.; Oliveira, E.B.; Kerkis, I.; Karpel, R.L. Crotamine: A novel cell-penetrating polypeptide nanocarrier with potential anti-cancer and biotechnological applications. Methods Mol. Biol. 2012, 906, 337–352. [Google Scholar]
- Vu, T.T.; Jeong, B.; Yu, J.; Koo, B.K.; Jo, S.H.; Robinson, R.C.; Choe, H. Soluble prokaryotic expression and purification of crotamine using an N-terminal maltose-binding protein tag. Toxicon 2014, 92, 157–165. [Google Scholar] [CrossRef]
- Peigneur, S.; Orts, D.J.; Prieto da Silva, A.R.; Oguiura, N.; Boni-Mitake, M.; de Oliveira, E.B.; Zaharenko, A.J.; de Freitas, J.C.; Tytgat, J. Crotamine pharmacology revisited: Novel insights based on the inhibition of KV channels. Mol. Pharmacol. 2012, 82, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Naloxone for Treatement of Opioide Overdose. Available online: https://www.fda.gov/media/100429/download (accessed on 30 September 2021).
- de Oliveira, S.A.; Magalhaes, M.R.; de Oliveira, L.P.; da Cunha, L.C. Identification of antinociceptive fraction of snake venom from Crotalus durissus collilineatus crotamine-negative and its acute toxicity evaluation. Toxicon 2016, 122, 145–151. [Google Scholar] [CrossRef]
- Chung, T.W.; Li, S.; Lin, C.C.; Tsai, S.W. Antinociceptive and anti-inflammatory effects of the citrus flavanone naringenin. Ci Ji Yi Xue Za Zhi 2019, 31, 81–85. [Google Scholar]
- Naloxone DrugFacts. Available online: https://www.drugabuse.gov/publications/drugfacts/naloxone (accessed on 30 September 2021).
- Bannon, A.W.; Malmberg, A.B. Models of nociception: Hot-plate, tail-flick, and formalin tests in rodents. Curr. Protoc. Neurosci. Chapter 8, Unit 8 9. 8. [CrossRef]
- Lopes, D.M.; Cater, H.L.; Thakur, M.; Wells, S.; McMahon, S.B. A refinement to the formalin test in mice. F1000Res 2019, 8, 891. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, H.; Dubois, D.; Gendron, L. Activation of spinal mu- and delta-opioid receptors potently inhibits substance P release induced by peripheral noxious stimuli. J. Neurosci. 2011, 31, 13068–13077. [Google Scholar] [CrossRef] [Green Version]
- Asahara, M.; Ito, N.; Yokomizo, T.; Nakamura, M.; Shimizu, T.; Yamada, Y. The absence of the leukotriene B4 receptor BLT1 attenuates peripheral inflammation and spinal nociceptive processing following intraplantar formalin injury. Mol. Pain 2015, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.J.; Bak, S.G.; Lim, H.J.; Lee, S.W.; Lee, S.; Ku, S.K.; Park, S.I.; Lee, S.J.; Rho, M.C. Acyclic Triterpenoid Isolated from Alpinia katsumadai Alleviates Formalin-Induced Chronic Mouse Paw Inflammation by Inhibiting the Phosphorylation of ERK and NF-kappaB. Molecules 2020, 25, 15. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Xing, W.; Li, W.; Fan, T.; Hu, H.; Li, Y. Isofraxidin exhibited anti-inflammatory effects in vivo and inhibited TNF-alpha production in LPS-induced mouse peritoneal macrophages in vitro via the MAPK pathway. Int. Immunopharmacol. 2012, 14, 164–171. [Google Scholar] [CrossRef]
- Kayser, V.; Viguier, F.; Melfort, M.; Bourgoin, S.; Hamon, M.; Masson, J. Altered nociception in mice with genetically induced hypoglutamatergic tone. Neuroscience 2015, 293, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Nemirovsky, A.; Chen, L.; Zelman, V.; Jurna, I. The antinociceptive effect of the combination of spinal morphine with systemic morphine or buprenorphine. Anesth. Analg. 2001, 93, 197–203. [Google Scholar] [CrossRef]
- Amico-Roxas, M.; Caruso, A.; Trombadore, S.; Scifo, R.; Scapagnini, U. Gangliosides antinociceptive effects in rodents. Arch. Int. Pharmacodyn. Ther. 1984, 272, 103–117. [Google Scholar]
- Dubuisson, D.; Dennis, S.G. The formalin test: A quantitative study of the analgesic effects of morphine, meperidine, and brain stem stimulation in rats and cats. Pain 1977, 4, 161–174. [Google Scholar] [CrossRef]
- Hunskaar, S.; Hole, K. The formalin test in mice: Dissociation between inflammatory and non-inflammatory pain. Pain 1987, 30, 103–114. [Google Scholar] [CrossRef]
- Bingham, S.; Beswick, P.J.; Blum, D.E.; Gray, N.M.; Chessell, I.P. The role of the cylooxygenase pathway in nociception and pain. Semin. Cell Dev. Biol. 2006, 17, 544–554. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [Green Version]
- Viggiano, E.; Monda, M.; Viggiano, A.; Viggiano, A.; Aurilio, C.; De Luca, B. Persistent facial pain increases superoxide anion production in the spinal trigeminal nucleus. Mol. Cell Biochem. 2010, 339, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Porreca, F.; Cuzzocrea, S.; Galen, K.; Lightfoot, R.; Masini, E.; Muscoli, C.; Mollace, V.; Ndengele, M.; Ischiropoulos, H.; et al. A newly identified role for superoxide in inflammatory pain. J. Pharmacol. Exp. Ther. 2004, 309, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Cunha, F.Q.; Poole, S.; Lorenzetti, B.B.; Ferreira, S.H. The pivotal role of tumor necrosis factor alpha in the development of inflammatory hyperalgesia. Br. J. Pharmacol. 1992, 107, 660–664. [Google Scholar] [CrossRef]
- Seo, Y.J.; Kwon, M.S.; Shim, E.J.; Park, S.H.; Choi, O.S.; Suh, H.W. Changes in pain behavior induced by formalin, substance P, glutamate and pro-inflammatory cytokines in immobilization-induced stress mouse model. Brain Res. Bull. 2006, 71, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.J.; Allchorne, A.; Safieh-Garabedian, B.; Poole, S. Cytokines, nerve growth factor and inflammatory hyperalgesia: The contribution of tumor necrosis factor alpha. Br. J. Pharmacol. 1997, 121, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.S.; Billings, J.A. Opioid antagonists: A review of their role in palliative care, focusing on use in opioid-related constipation. J. Pain Symptom Manage. 2002, 24, 71–90. [Google Scholar] [CrossRef]
- Vanky, E.; Hellmundt, L.; Bondesson, U.; Eksborg, S.; Lundeberg, S. Pharmacokinetics after a single dose of naloxone administered as a nasal spray in healthy volunteers. Acta Anaesthesiol. Scand. 2017, 61, 636–640. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saline | Crotamine (mg/kg) | ||||
|---|---|---|---|---|---|
| 0.04 | 0.13 | 0.4 | 1.2 | ||
| Saline | Group 1 | Group 3 | Group 5 | Group 7 | Group 9 |
| Naloxone | Group 2 | Group 4 | Group 6 | Group 8 | Group 10 |
| Saline | Crotamine (mg/kg) | ||||
|---|---|---|---|---|---|
| 0.04 | 0.13 | 0.4 | 1.2 | ||
| Saline | Group 1 | Group 3 | Group 5 | Group 7 | Group 9 |
| Naloxone | Group 2 | Group 4 | Group 6 | Group 8 | Group 10 |
| Saline | Indomethacin | Crotamine (mg/kg) | |||
|---|---|---|---|---|---|
| 0.13 | 0.4 | 1.2 | |||
| Saline | Group 1 | Group 2 | Group 4 | Group 5 | Group 7 |
| Naloxone | Group 3 | Group 6 | Group 8 | ||
| Saline | Indomethacin | Crotamine (μg 10 μL−1) | |||
|---|---|---|---|---|---|
| 0.9 | 2.5 | 7.5 | |||
| Saline | Group 1 | Group 2 | Group 4 | Group 5 | Group 7 |
| Naloxone | Group 3 | Group 6 | Group 8 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.Y.; Do, B.H.; Lee, J.-S.; Yang, H.C.; Nguyen, A.N.; Krupa, M.; Kim, C.J.; Jang, Y.J.; Choe, H. Antinociceptive and Anti-Inflammatory Effects of Recombinant Crotamine in Mouse Models of Pain. Toxins 2021, 13, 707. https://doi.org/10.3390/toxins13100707
Park JY, Do BH, Lee J-S, Yang HC, Nguyen AN, Krupa M, Kim CJ, Jang YJ, Choe H. Antinociceptive and Anti-Inflammatory Effects of Recombinant Crotamine in Mouse Models of Pain. Toxins. 2021; 13(10):707. https://doi.org/10.3390/toxins13100707
Chicago/Turabian StylePark, Jong Yeon, Bich Hang Do, Ju-Seung Lee, Hyun Cheol Yang, Anh Ngoc Nguyen, Martin Krupa, Chong Jai Kim, Yeon Jin Jang, and Han Choe. 2021. "Antinociceptive and Anti-Inflammatory Effects of Recombinant Crotamine in Mouse Models of Pain" Toxins 13, no. 10: 707. https://doi.org/10.3390/toxins13100707
APA StylePark, J. Y., Do, B. H., Lee, J. -S., Yang, H. C., Nguyen, A. N., Krupa, M., Kim, C. J., Jang, Y. J., & Choe, H. (2021). Antinociceptive and Anti-Inflammatory Effects of Recombinant Crotamine in Mouse Models of Pain. Toxins, 13(10), 707. https://doi.org/10.3390/toxins13100707