Increased Risk of Thrombocytopenia and Death in Patients with Bacteremia Caused by High Alpha Toxin-Producing Methicillin-Resistant Staphylococcus aureus


Abstract
:1. Introduction
2. Results
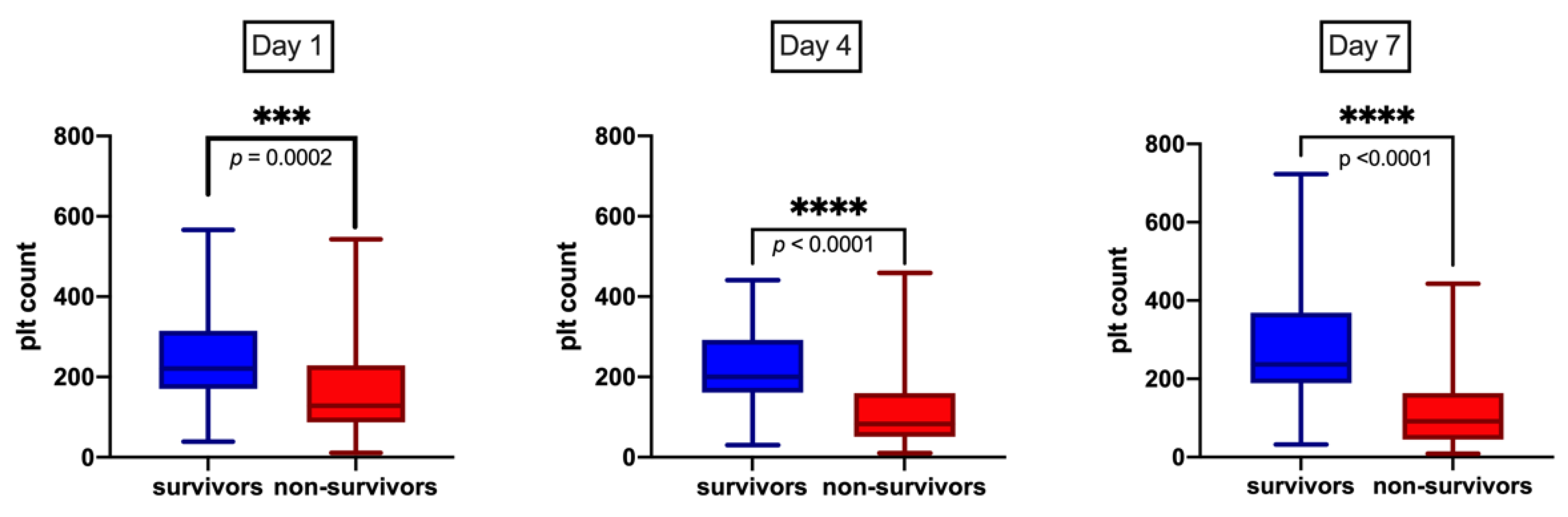
2.1. Patient Characteristics and Platelet Trends
2.2. High Correlation of Hla Protein Level and Hemolytic Activity
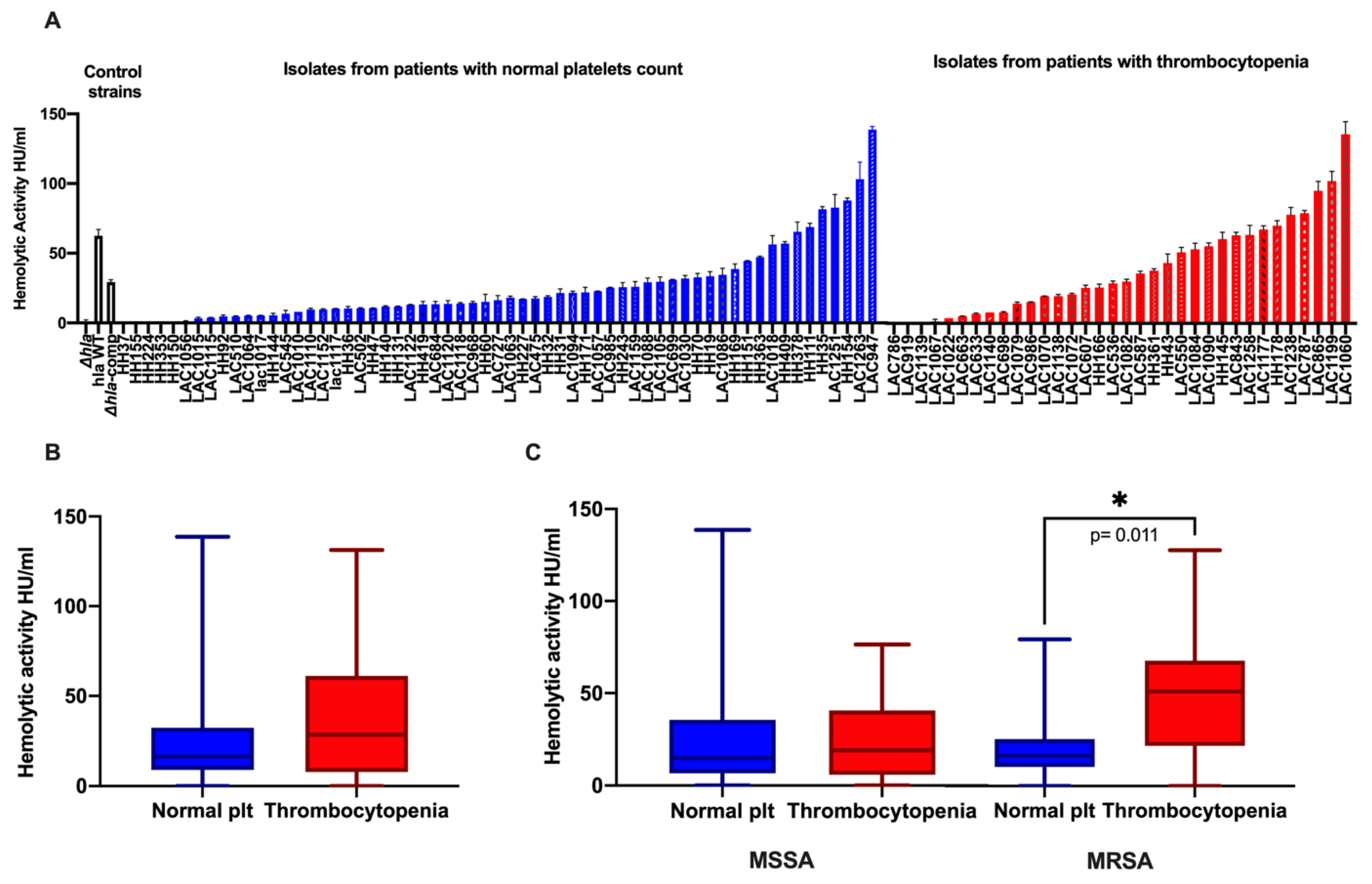
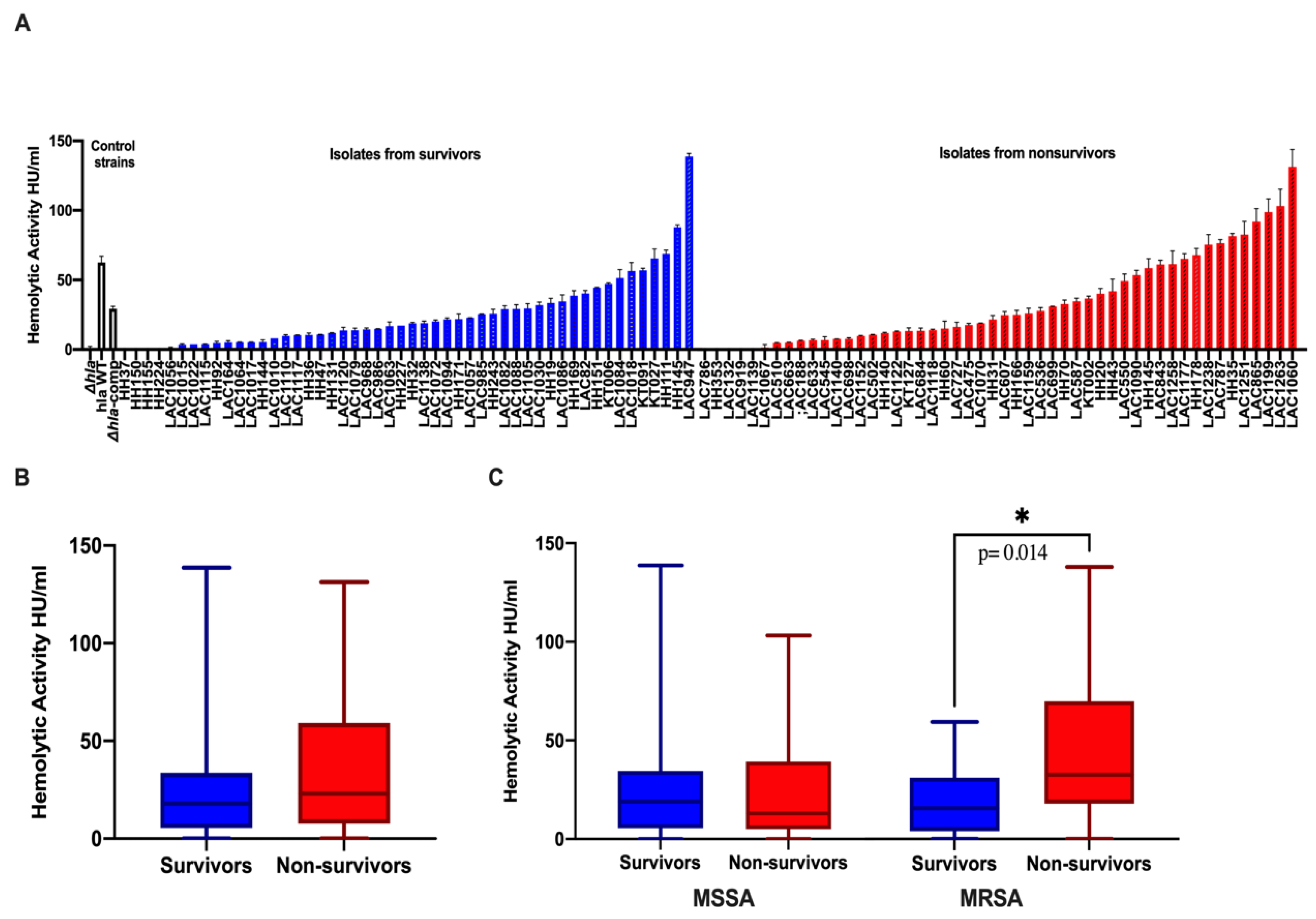
2.3. Association of High Hemolytic Activity with Thrombocytopenia and Mortality
2.4. Association of High Hemolytic Activity with High-Risk Source of Infection
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patient and Bacterial Isolate Selection
5.2. In Vitro Measurement of Hla Expression
5.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of mortality in Staphylococcus aureus Bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef] [Green Version]
- Minejima, E.; Bensman, J.; She, R.C.; Mack, W.J.; Tuan Tran, M.; Ny, P.; Lou, M.; Yamaki, J.; Nieberg, P.; Ho, J.; et al. A Dysregulated Balance of Proinflammatory and Anti-Inflammatory Host Cytokine Response Early During Therapy Predicts Persistence and Mortality in Staphylococcus aureus Bacteremia. Crit. Care Med. 2016, 44, 671–679. [Google Scholar] [CrossRef]
- Berube, B.J.; Bubeck Wardenburg, J. Staphylococcus aureus alpha-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.A.; Wuescher, L.M.; Dona, K.R.; Worth, R.G. Platelets Mediate Host Defense against Staphylococcus aureus through Direct Bactericidal Activity and by Enhancing Macrophage Activities. J. Immunol. 2017, 198, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Wuescher, L.M.; Takashima, A.; Worth, R.G. A novel conditional platelet depletion mouse model reveals the importance of platelets in protection against Staphylococcus aureus bacteremia. J. Thromb. Haemost. 2015, 13, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, M.E.; Becker, R.E.; Sailer, A.; Turner, J.R.; Bubeck Wardenburg, J. Synergistic Action of Staphylococcus aureus alpha-Toxin on Platelets and Myeloid Lineage Cells Contributes to Lethal Sepsis. Cell Host Microbe 2015, 17, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Uchiyama, S.; Olson, J.; Morodomi, Y.; Cornax, I.; Ando, N.; Kohno, Y.; Kyaw, M.M.T.; Aguilar, B.; Haste, N.M.; et al. Repurposed drugs block toxin-driven platelet clearance by the hepatic Ashwell-Morell receptor to clear Staphylococcus aureus bacteremia. Sci. Transl. Med. 2021, 13, 586. [Google Scholar] [CrossRef]
- Surewaard, B.G.J.; Thanabalasuriar, A.; Zeng, Z.; Tkaczyk, C.; Cohen, T.S.; Bardoel, B.W.; Jorch, S.K.; Deppermann, C.; Bubeck Wardenburg, J.; Davis, R.P.; et al. alpha-Toxin Induces Platelet Aggregation and Liver Injury during Staphylococcus aureus Sepsis. Cell Host Microbe 2018, 24, 271–284.e273. [Google Scholar] [CrossRef]
- Bubeck Wardenburg, J.; Patel, R.J.; Schneewind, O. Surface proteins and exotoxins are required for the pathogenesis of Staphylococcus aureus pneumonia. Infect. Immun. 2007, 75, 1040–1044. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, A.H.; Foster, T.J.; Hayashida, A.; Park, P.W. Alpha-toxin facilitates the generation of CXC chemokine gradients and stimulates neutrophil homing in Staphylococcus aureus pneumonia. J. Infect. Dis. 2008, 198, 1529–1535. [Google Scholar] [CrossRef] [Green Version]
- Rauch, S.; DeDent, A.C.; Kim, H.K.; Bubeck Wardenburg, J.; Missiakas, D.M.; Schneewind, O. Abscess formation and alpha-hemolysin induced toxicity in a mouse model of Staphylococcus aureus peritoneal infection. Infect. Immun. 2012, 80, 3721–3732. [Google Scholar] [CrossRef] [Green Version]
- Kielian, T.; Cheung, A.; Hickey, W.F. Diminished virulence of an alpha-toxin mutant of Staphylococcus aureus in experimental brain abscesses. Infect. Immun. 2001, 69, 6902–6911. [Google Scholar] [CrossRef] [Green Version]
- Hua, L.; Hilliard, J.J.; Shi, Y.; Tkaczyk, C.; Cheng, L.I.; Yu, X.; Datta, V.; Ren, S.; Feng, H.; Zinsou, R.; et al. Assessment of an anti-alpha-toxin monoclonal antibody for prevention and treatment of Staphylococcus aureus-induced pneumonia. Antimicrob. Agents Chemother. 2014, 58, 1108–1117. [Google Scholar] [CrossRef] [Green Version]
- Diep, B.A.; Hilliard, J.J.; Le, V.T.; Tkaczyk, C.; Le, H.N.; Tran, V.G.; Rao, R.L.; Dip, E.C.; Pereira-Franchi, E.P.; Cha, P.; et al. Targeting Alpha Toxin To Mitigate Its Lethal Toxicity in Ferret and Rabbit Models of Staphylococcus aureus Necrotizing Pneumonia. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Sampedro, G.R.; DeDent, A.C.; Becker, R.E.; Berube, B.J.; Gebhardt, M.J.; Cao, H.; Bubeck Wardenburg, J. Targeting Staphylococcus aureus alpha-toxin as a novel approach to reduce severity of recurrent skin and soft-tissue infections. J. Infect. Dis. 2014, 210, 1012–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, V.T.; Tkaczyk, C.; Chau, S.; Rao, R.L.; Dip, E.C.; Pereira-Franchi, E.P.; Cheng, L.; Lee, S.; Koelkebeck, H.; Hilliard, J.J.; et al. Critical Role of Alpha-Toxin and Protective Effects of Its Neutralization by a Human Antibody in Acute Bacterial Skin and Skin Structure Infections. Antimicrob. Agents Chemother. 2016, 60, 5640–5648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, R.P.; Ajao, A.O.; Aman, M.J.; Karauzum, H.; Sarwar, J.; Lydecker, A.D.; Johnson, J.K.; Nguyen, C.; Chen, W.H.; Roghmann, M.C. Lower antibody levels to Staphylococcus aureus exotoxins are associated with sepsis in hospitalized adults with invasive S. aureus infections. J. Infect. Dis. 2012, 206, 915–923. [Google Scholar] [CrossRef]
- Li, S.; Arvidson, S.; Mollby, R. Variation in the agr-dependent expression of alpha-toxin and protein A among clinical isolates of Staphylococcus aureus from patients with septicaemia. FEMS Microbiol. Lett. 1997, 152, 155–161. [Google Scholar] [CrossRef]
- Stulik, L.; Malafa, S.; Hudcova, J.; Rouha, H.; Henics, B.Z.; Craven, D.E.; Sonnevend, A.M.; Nagy, E. alpha-Hemolysin activity of methicillin-susceptible Staphylococcus aureus predicts ventilator-associated pneumonia. Am. J. Respir Crit. Care Med. 2014, 190, 1139–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, A.; Nielsen, J.B.; Boye, K.; Rohde, S.; Paulo, A.C.; Westh, H.; Schonning, K.; de Lencastre, H.; Miragaia, M. Insights into alpha-hemolysin (Hla) evolution and expression among Staphylococcus aureus clones with hospital and community origin. PLoS ONE 2014, 9, e98634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, A.; Marco, F.; Martinez, J.A.; Pisos, E.; Almela, M.; Dimova, V.P.; Alamo, D.; Ortega, M.; Lopez, J.; Mensa, J. Influence of vancomycin minimum inhibitory concentration on the treatment of methicillin-resistant Staphylococcus aureus bacteremia. Clin. Infect. Dis. 2008, 46, 193–200. [Google Scholar] [CrossRef]
- Gafter-Gvili, A.; Mansur, N.; Bivas, A.; Zemer-Wassercug, N.; Bishara, J.; Leibovici, L.; Paul, M. Thrombocytopenia in Staphylococcus aureus bacteremia: Risk factors and prognostic importance. Mayo Clin. Proc. 2011, 86, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Akca, S.; Haji-Michael, P.; de Mendonca, A.; Suter, P.; Levi, M.; Vincent, J.L. Time course of platelet counts in critically ill patients. Crit. Care Med. 2002, 30, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Vandijck, D.M.; Blot, S.I.; De Waele, J.J.; Hoste, E.A.; Vandewoude, K.H.; Decruyenaere, J.M. Thrombocytopenia and outcome in critically ill patients with bloodstream infection. Heart Lung 2010, 39, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Coia, J.E.; Browning, L.; Haines, L.; Birkbeck, T.H.; Platt, D.J. Comparison of enterotoxins and haemolysins produced by methicillin-resistant (MRSA) and sensitive (MSSA) Staphylococcus aureus. J. Med. Microbiol. 1992, 36, 164–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsson, G.; Colque-Navarro, P.; Gustafsson, E.; Andersson, R.; Mollby, R. Antibody responses in patients with invasive Staphylococcus aureus infections. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 715–725. [Google Scholar] [CrossRef]
- Sharma-Kuinkel, B.K.; Wu, Y.; Tabor, D.E.; Mok, H.; Sellman, B.R.; Jenkins, A.; Yu, L.; Jafri, H.S.; Rude, T.H.; Ruffin, F.; et al. Characterization of alpha-toxin hla gene variants, alpha-toxin expression levels, and levels of antibody to alpha-toxin in hemodialysis and postsurgical patients with Staphylococcus aureus bacteremia. J. Clin. Microbiol. 2015, 53, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, K.; Lucar, J.; Blackshear, C.; Hobbs, C.V. Methicillin-susceptible and Methicillin-resistant Staphylococcus aureus Bacteremia: Nationwide Estimates of 30-Day Readmission, In-hospital Mortality, Length of Stay, and Cost in the United States. Clin. Infect. Dis. 2019, 69, 2112–2118. [Google Scholar] [CrossRef] [PubMed]
- Whitby, M.; McLaws, M.L.; Berry, G. Risk of death from methicillin-resistant Staphylococcus aureus bacteraemia: A meta-analysis. Med. J. Aust. 2001, 175, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, S.E.; Sakoulas, G.; Perencevich, E.N.; Schwaber, M.J.; Karchmer, A.W.; Carmeli, Y. Comparison of mortality associated with methicillin-resistant and methicillin-susceptible Staphylococcus aureus bacteremia: A meta-analysis. Clin. Infect. Dis. 2003, 36, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Cheung, G.Y.; Wang, R.; Khan, B.A.; Sturdevant, D.E.; Otto, M. Role of the accessory gene regulator agr in community-associated methicillin-resistant Staphylococcus aureus pathogenesis. Infect. Immun. 2011, 79, 1927–1935. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. MRSA virulence and spread. Cell Microbiol. 2012, 14, 1513–1521. [Google Scholar] [CrossRef] [Green Version]
- Chua, K.Y.; Monk, I.R.; Lin, Y.H.; Seemann, T.; Tuck, K.L.; Porter, J.L.; Stepnell, J.; Coombs, G.W.; Davies, J.K.; Stinear, T.P.; et al. Hyperexpression of alpha-hemolysin explains enhanced virulence of sequence type 93 community-associated methicillin-resistant Staphylococcus aureus. BMC Microbiol. 2014, 14, 31. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Dai, Y.; Zhu, Y.; Fu, C.L.; Tan, V.Y.; Wang, Y.; Wang, X.; Hong, X.; Liu, Q.; Li, T.; et al. Virulence determinants associated with the Asian community-associated methicillin-resistant Staphylococcus aureus lineage ST59. Sci. Rep. 2016, 6, 27899. [Google Scholar] [CrossRef] [Green Version]
- Yamaki, J.; Synold, T.; Wong-Beringer, A. Tigecycline induction of phenol-soluble modulins by invasive methicillin-resistant Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2013, 57, 4562–4565. [Google Scholar] [CrossRef] [Green Version]
- Berube, B.J.; Sampedro, G.R.; Otto, M.; Bubeck Wardenburg, J. The psmalpha locus regulates production of Staphylococcus aureus alpha-toxin during infection. Infect. Immun. 2014, 82, 3350–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34, table of contents. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.; Borges, A.; Simoes, M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, P.A.; Taylor, R.; Thielke, R.; Payne, J.; Gonzalez, N.; Conde, J.G. Research electronic data capture (REDCap)--a metadata-driven methodology and workflow process for providing translational research informatics support. J. Biomed. Inform. 2009, 42, 377–381. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.D.; Bubeck Wardenburg, J.; Gardner, D.J.; Long, D.; Whitney, A.R.; Braughton, K.R.; Schneewind, O.; DeLeo, F.R. Targeting of alpha-hemolysin by active or passive immunization decreases severity of USA300 skin infection in a mouse model. J. Infect. Dis. 2010, 202, 1050–1058. [Google Scholar] [CrossRef] [Green Version]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Ross, H.F.; Projan, S.J.; Kornblum, J.; Kreiswirth, B.; Moghazeh, S. Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J. 1993, 12, 3967–3975. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Thrombocytopenia (n = 34) | Non-Thrombocytopenia (n = 61) | p Value | Non-Survivors (n = 50) | Survivors (n = 50) | p Value |
|---|---|---|---|---|---|---|
| Sex (male) | 21 (61%) | 42 (68.8%) | 0.504 | 31 (62%) | 35 (70%) | 0.53 |
| Age, y, mean (SD) | 58.91 (15.24) | 56.84 (18.28) | 0.575 | 62.66 (14.74) | 52.60 (17.86) | 0.002 * |
| IV drug use | 3 (8.8%) | 7 (11.4%) | >0.99 | 6 (12%) | 5 (10%) | >0.99 |
| Alcohol use | 5 (14.7%) | 12 (19.6%) | 0.591 | 3 (6%) | 14 (28%) | 0.006 * |
| Active malignancy | 5 (14.7%) | 6 (9.8%) | 0.498 | 10 (20%) | 3 (6%) | 0.071 |
| Liver cirrhosis | 8 (23.5%) | 5 (8.2%) | 0.059 | 7 (14%) | 6 (12%) | >0.99 |
| Renal disease | 12 (35.3%) | 21 (34.4%) | >0.99 | 17 (34%) | 17 (34%) | >0.99 |
| High risk mortality a | 17 (50%) | 15 (24.5%) | 0.022 * | 22 (44%) | 11(22%) | 0.032 * |
| Intermediate risk mortality b | 10 (29.4%) | 20 (32.7%) | 0.649 | 17 (34%) | 17 (34%) | >0.99 |
| Low risk mortality c | 7 (20.5%) | 26 (42.6%) | 0.042 * | 11(22%) | 22 (44%) | 0.032 * |
| MRSA as causative Pathogen | 18(53%) | 19 (31%) | 0.048 * | 25 (50%) | 15 (30%) | 0.065 |
| Platelet count at onset, 109/L (median, IQR) | 96.5 (56, 123) | 241 (201, 319) | <0.0001 * | 128 (87, 229) | 221 (170, 315) | 0.0002 * |
| Death | 27 (79%) | 20 (32.8%) | <0.0001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhurayri, F.; Porter, E.; Douglas-Louis, R.; Minejima, E.; Wardenburg, J.B.; Wong-Beringer, A. Increased Risk of Thrombocytopenia and Death in Patients with Bacteremia Caused by High Alpha Toxin-Producing Methicillin-Resistant Staphylococcus aureus. Toxins 2021, 13, 726. https://doi.org/10.3390/toxins13100726
Alhurayri F, Porter E, Douglas-Louis R, Minejima E, Wardenburg JB, Wong-Beringer A. Increased Risk of Thrombocytopenia and Death in Patients with Bacteremia Caused by High Alpha Toxin-Producing Methicillin-Resistant Staphylococcus aureus. Toxins. 2021; 13(10):726. https://doi.org/10.3390/toxins13100726
Chicago/Turabian StyleAlhurayri, Fatimah, Edith Porter, Rachid Douglas-Louis, Emi Minejima, Juliane Bubeck Wardenburg, and Annie Wong-Beringer. 2021. "Increased Risk of Thrombocytopenia and Death in Patients with Bacteremia Caused by High Alpha Toxin-Producing Methicillin-Resistant Staphylococcus aureus" Toxins 13, no. 10: 726. https://doi.org/10.3390/toxins13100726
APA StyleAlhurayri, F., Porter, E., Douglas-Louis, R., Minejima, E., Wardenburg, J. B., & Wong-Beringer, A. (2021). Increased Risk of Thrombocytopenia and Death in Patients with Bacteremia Caused by High Alpha Toxin-Producing Methicillin-Resistant Staphylococcus aureus. Toxins, 13(10), 726. https://doi.org/10.3390/toxins13100726