Varying Intensities of Introgression Obscure Incipient Venom-Associated Speciation in the Timber Rattlesnake (Crotalus horridus)

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Reversed-Phase High-Performance Liquid Chromatography
2.3. Canebrake Toxin PCR Assay
2.4. Morphological Analysis
2.5. Niche Modeling
2.6. mtDNA Sequencing and Phylogenetic Reconstruction
3. Results and Discussion
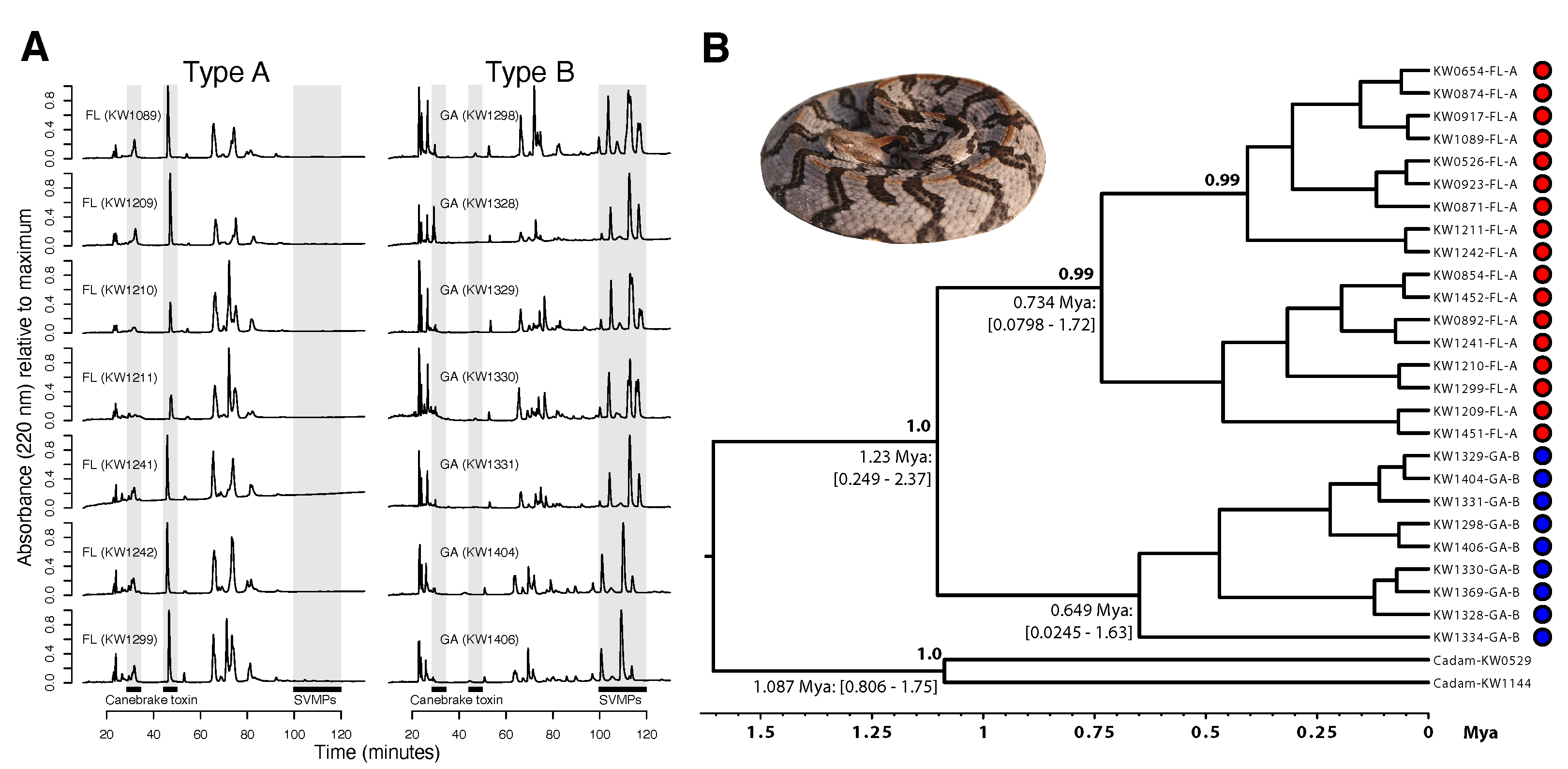
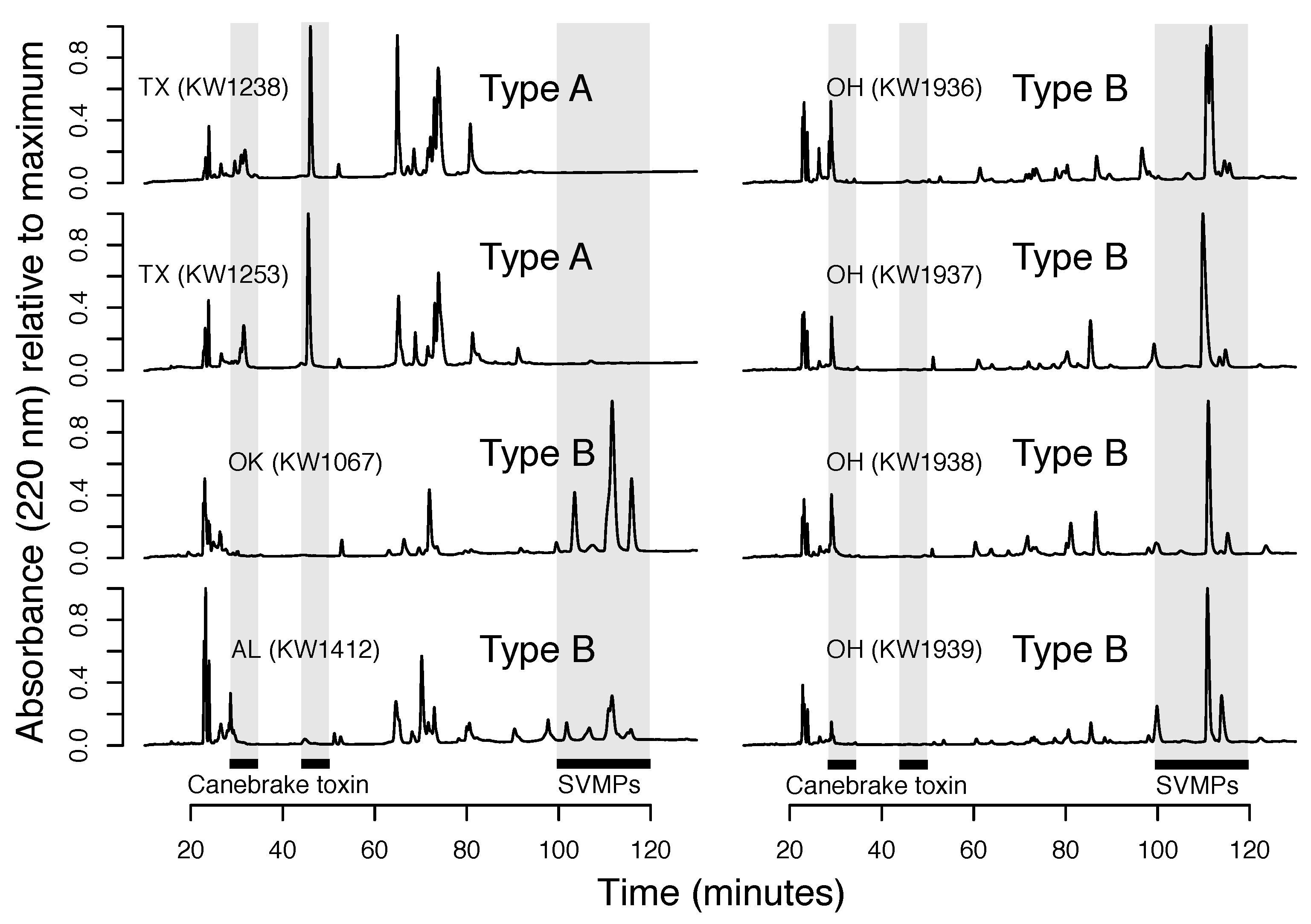
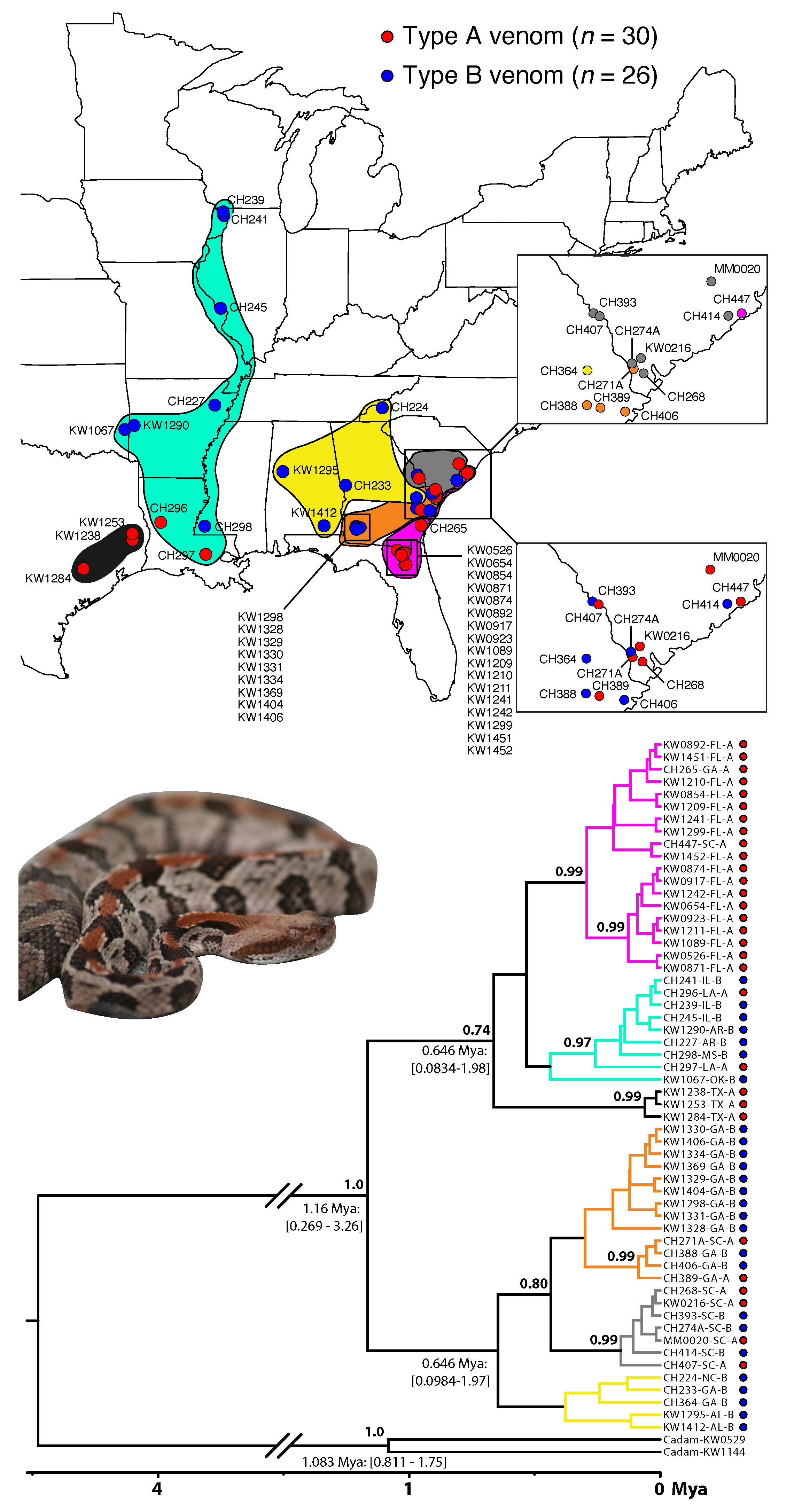
3.1. Fixed Venom Differences, Deep Mitochondrial Genetic Divergence, and Distinct Morphologies over Small Spatial Scales
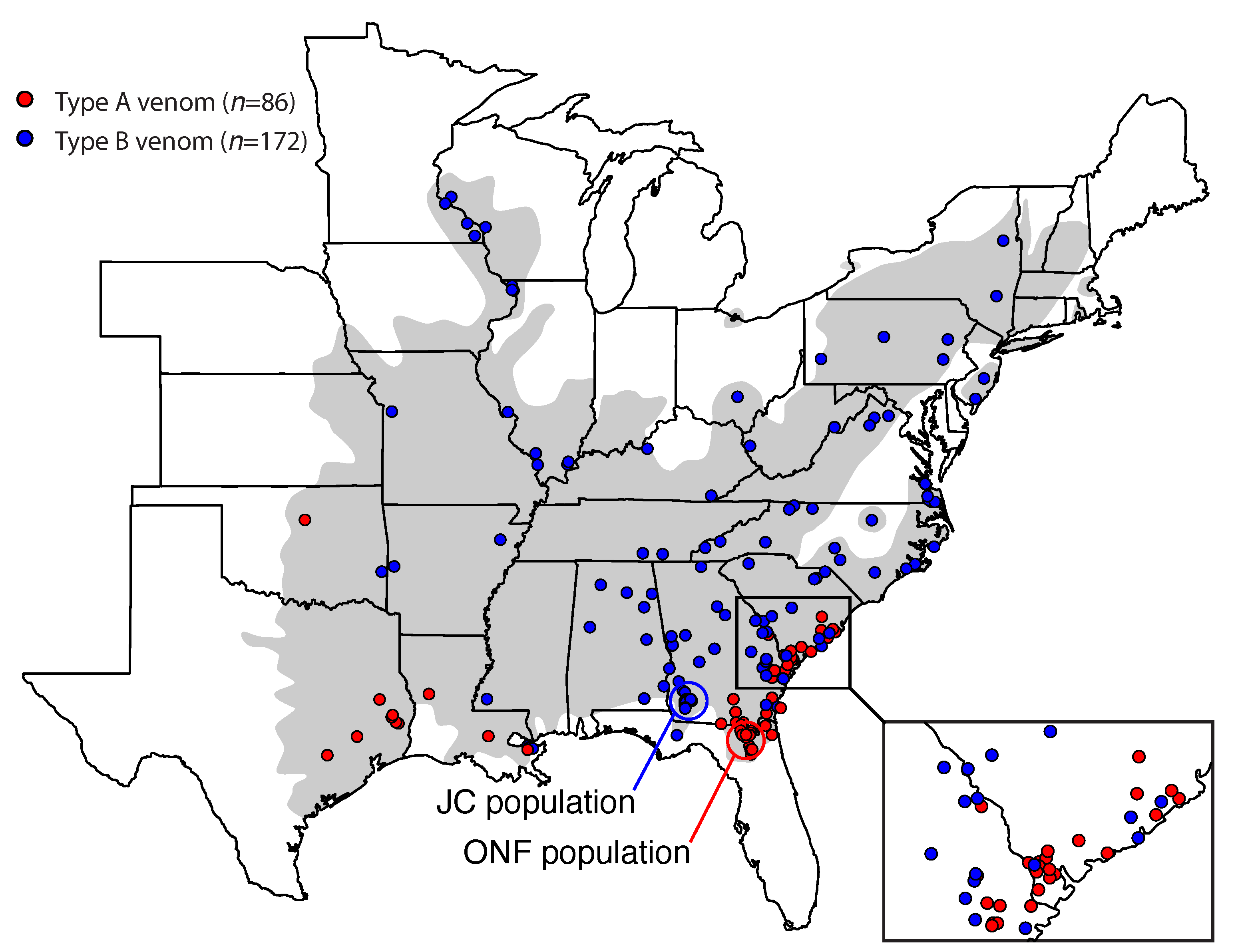
3.2. Type A Venom Dominates along the Southern Periphery of the Range
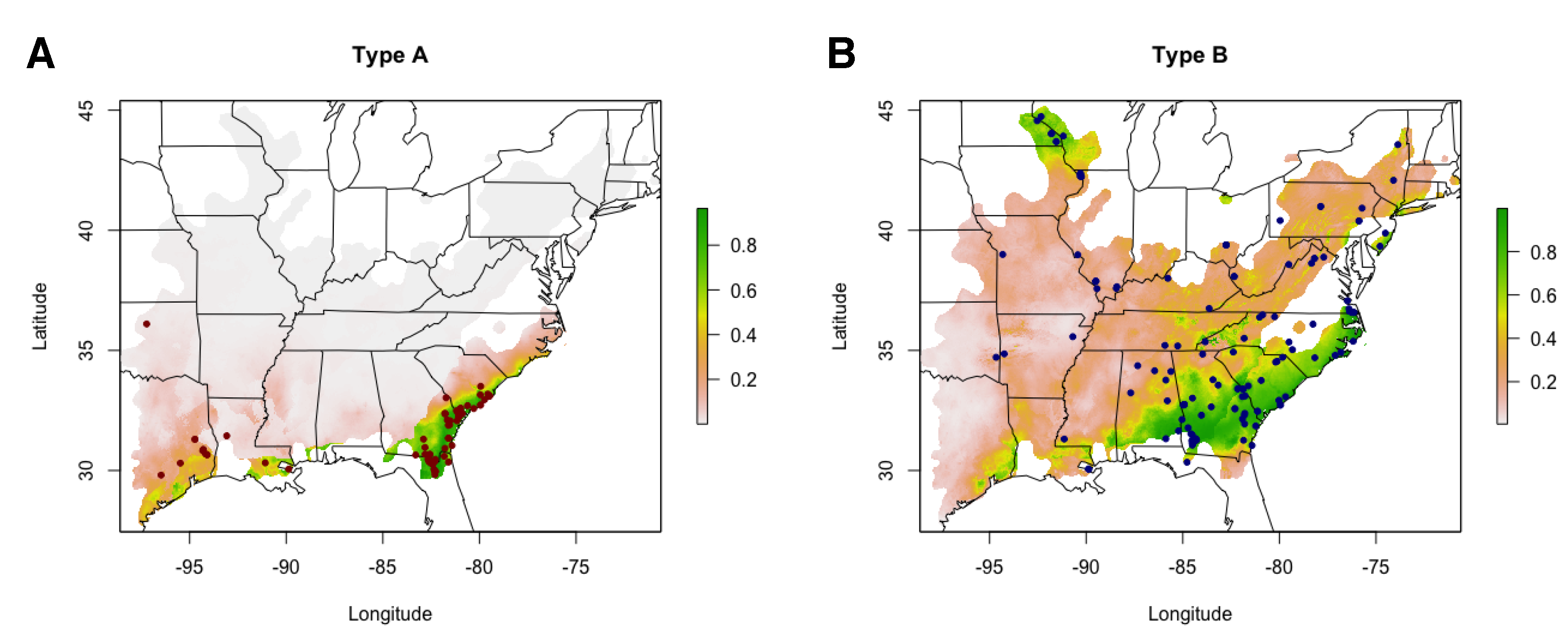
3.3. Type A and B Individuals Occupy Significantly Different Niches
3.4. A Lack of Reciprocal Monophyly Suggests Gene Flow between Venom Types across the Range
3.5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hendry, A.P.; Nosil, P.; Rieseberg, L.H. The speed of ecological speciation. Funct. Ecol. 2007, 21, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Nosil, P.; Funk, D.J.; Ortiz-Barrientos, D. Divergent selection and heterogeneous genomic divergence. Mol. Ecol. 2009, 18, 375–402. [Google Scholar] [CrossRef]
- Schluter, D. Evidence for ecological speciation and its alternative. Science 2009, 323, 737–741. [Google Scholar] [CrossRef] [Green Version]
- Keller, I.; Seehausen, O. Thermal adaptation and ecological speciation. Mol. Ecol. 2012, 21, 782–799. [Google Scholar] [CrossRef]
- Öhlund, G.; Bodin, M.; Nilsson, K.A.; Öhlund, S.O.; Mobley, K.B.; Hudson, A.G.; Peedu, M.; Brännström, Å.; Bartels, P.; Præbel, K.; et al. Ecological speciation in European whitefish is driven by a large-gaped predator. Evol. Lett. 2020, 4, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Garant, D.; Forde, S.E.; Hendry, A.P. The multifarious effects of dispersal and gene flow on contemporary adaptation. Funct. Ecol. 2007, 21, 434–443. [Google Scholar] [CrossRef]
- Zhang, L.; Hood, G.R.; Roush, A.M.; Shzu, S.A.; Comerford, M.S.; Ott, J.R.; Egan, S.P. Asymmetric, but opposing reductions in immigrant viability and fecundity promote reproductive isolation among host-associated populations of an insect herbivore. Evolution 2021, 75, 476–489. [Google Scholar] [CrossRef]
- Ryan, P.G.; Bloomer, P.; Moloney, C.L.; Grant, T.J.; Delport, W. Ecological speciation in South Atlantic island finches. Science 2007, 315, 1420–1423. [Google Scholar] [CrossRef] [Green Version]
- Shafer, A.B.; Wolf, J.B. Widespread evidence for incipient ecological speciation: A meta-analysis of isolation-by-ecology. Ecol. Lett. 2013, 16, 940–950. [Google Scholar] [CrossRef]
- Nosil, P.; Harmon, L.J.; Seehausen, O. Ecological explanations for (incomplete) speciation. Trends Ecol. Evol. 2009, 24, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Endepols, S.; Klemann, N.; Richter, D.; Matuschka, F.R.; Shih, C.H.; Nachman, M.W.; Kohn, M.H. Adaptive introgression of anticoagulant rodent poison resistance by hybridization between old world mice. Curr. Biol. 2011, 21, 1296–1301. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, J.S.; Kawji, Y.; Kelly, M.W. An experimental test of adaptive introgression in locally adapted populations of splash pool copepods. Mol. Biol. Evol. 2020, 38, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Margres, M.; Patton, A.; Wray, K.; Hassinger, A.; Ward, M.; Lemmon, E.; Lemmon, A.; Rokyta, D. Tipping the scales: The migration-selection balance leans toward selection in snake venoms. Mol. Biol. Evol. 2019, 36, 271–282. [Google Scholar] [CrossRef]
- Rokyta, D.; Margres, M.; Calvin, K. Post-transcriptional mechanisms contribute little to phenotypic variation in snake venoms. G3 Genes Genomes Genet. 2015, g3, 115. [Google Scholar] [CrossRef] [Green Version]
- Margres, M.J.; Rautsaw, R.M.; Strickland, J.L.; Mason, A.J.; Schramer, T.D.; Hofmann, E.P.; Stiers, E.; Ellsworth, S.A.; Nystrom, G.S.; Hogan, M.P.; et al. The Tiger Rattlesnake genome reveals a complex genotype underlying a simple venom phenotype. Proc. Natl. Acad. Sci. USA 2021, 118, e2014634118. [Google Scholar] [CrossRef] [PubMed]
- Daltry, J.C.; Wüster, W.; Thorpe, R.S. Diet and snake venom evolution. Nature 1996, 379, 537–540. [Google Scholar] [CrossRef]
- Holding, M.; Biardi, J.; Gibbs, H. Coevolution of venom function and venom resistance in a rattlesnake predator and its squirrel prey. Proc. R. Soc. B 2016, 283, 20152841. [Google Scholar] [CrossRef]
- Margres, M.; Wray, K.; Hassinger, A.; Ward, M.; McGivern, J.; Lemmon, E.; Lemmon, A.; Rokyta, D. Quantity, not quality: Rapid adaptation in a polygenic trait proceeded exclusively through expression differentiation. Mol. Biol. Evol. 2017, 34, 3099–3110. [Google Scholar] [CrossRef] [Green Version]
- Lynch, V.J. Inventing an arsenal: adaptive evolution and neofunctionalization of snake venom phospholipase A2 genes. BMC Evol. Biol. 2007, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Margres, M.; Wray, K.; Seavy, M.; McGivern, J.; Herrera, N.; Rokyta, D. Expression differentiation is constrained to low-expression proteins over ecological timescales. Genetics 2016, 202, 273–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margres, M.; Bigelow, A.; Lemmon, E.; Lemmon, A.; Rokyta, D. Selection to increase expression, not sequence diversity, precedes gene family origin and expansion in rattlesnake venom. Genetics 2017, 206, 1569–1580. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, H.L.; Rossiter, W. Rapid Evolution by Positive Selection and Gene Gain and Loss: PLA2 Venom Genes in Closely Related Sistrurus Rattlesnakes with Divergent Diets. J. Mol. Evol. 2008, 66, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.L.; Strickland, J.L.; Rautsaw, R.M.; Hofmann, E.P.; Mason, A.J.; Hogan, M.P.; Nystrom, G.S.; Ellsworth, S.A.; Colston, T.J.; Borja, M.; et al. Phylogenetically diverse diets favor more complex venoms in North American pitvipers. Proc. Natl. Acad. Sci. USA 2021, 118, e2015579118. [Google Scholar] [CrossRef]
- Pavey, S.; Collin, H.; Nosil, P.; Rogers, S. The role of gene expression in ecological speciation. Ann. N. Y. Acad. Sci. 2010, 1206, 110–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackessy, S.P. Venom composition in rattlesnakes: Trends and biological significance. In The Biology of Rattlesnakes; Hayes, W.K., Beaman, K.R., Cardwell, M.D., Bush, S.P., Eds.; Loma Linda University Press: Loma Linda, CA, USA, 2008; pp. 495–510. [Google Scholar]
- Straight, R.C.; Glenn, J.L.; Wolt, T.B.; Wolfe, M.C. Regional differences in content of small basic peptide toxins in the venoms of Crotalus adamanteus and Crotalus horridus. Comp. Biochem. Physiol. 1991, 100, 51–58. [Google Scholar] [CrossRef]
- Glenn, J.L.; Straight, R.C.; Wolf, T.B. Regional variation in the presence of canebrake toxin in Crotalus horridus venom. Comp. Biochem. Phys. C 1994, 107, 337–346. [Google Scholar] [CrossRef]
- Mackessy, S.P. Evolutionary trends in venom composition in the Western Rattlesnakes (Crotalus viridis sensu lato): Toxicity vs. tenderizers. Toxicon 2010, 55, 1463–1474. [Google Scholar] [CrossRef]
- Dowell, N.; Giorgianni, M.; Kassner, V.; Selegue, J.; Sanchez, E.; Carroll, S. The deep origin and recent loss of venom toxin genes in rattlesnakes. Curr. Biol. 2016, 26, 2434–2445. [Google Scholar] [CrossRef] [Green Version]
- Dowell, N.; Giorgianni, M.; Griffin, A.; Kassner, V.; Selegue, J.; Sanchez, E.; Carroll, S. Extremely divergent haplotypes in two toxin gene complexes encode alternative venom types within rattlesnake species. Curr. Biol. 2018, 28, 1016–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, J.A.; Glenn, J.L.; Straight, R.C.; Sites, J.W., Jr. Distribution and genetic variation in venom A and B populations of the Mojave rattlesnake Crotalus scutulatus scutulatus in Arizona. Herpetologica 1991, 47, 54–68. [Google Scholar]
- Zancolli, G.; Baker, T.G.; Barlow, A.; Bradley, R.K.; Calvete, J.J.; Carter, K.C.; De Jager, K.; Owens, J.B.; Price, J.F.; Sanz, L.; et al. Is hybridization a source of adaptive venom variation in rattlesnakes? A test, using a Crotalus scutulatus × viridis hybrid zone in southwestern New Mexico. Toxins 2016, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Strickland, J.; Smith, C.; Mason, A.; Schield, D.; Borja, M.; Cataneda-Gaytan, G.; Spencer, C.; Smith, L.; Trapaga, A.; Bouzid, N.; et al. Evidence for divergent patterns of local selection driving venom variation in Mojave Rattlesnakes (Crotalus scutulatus). Sci. Rep. 2018, 8, 17622. [Google Scholar] [CrossRef]
- McCue, M.D. Cost of Producing Venom in Three North American Pitviper Species. Copeia 2006, 2006, 818–825. [Google Scholar] [CrossRef]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. Lond. B 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDiarmid, R.W.; Campbell, J.A.; Touré, T. Snake Species of the World: A Taxonomic and Geographic Reference, Vol. 1; Herpetologists’ League: Washington, DC, USA, 1999. [Google Scholar]
- Rokyta, D.; Wray, K.; McGivern, J.; Margres, M. The transcriptomic and proteomic basis for the evolution of a novel venom phenotype within the Timber Rattlesnake (Crotalus horridus). Toxicon 2015, 98, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Margres, M.; Wray, K.; Seavy, M.; McGivern, J.; Sanader, D.; Rokyta, D. Phenotypic integration in the feeding system of the eastern diamondback rattlesnake (Crotalus adamanteus). Mol. Ecol. 2015, 24, 3405–3420. [Google Scholar] [CrossRef]
- Cleuren, S.G.; Hocking, D.P.; Evans, A.R. Fang evolution in venomous snakes: Adaptation of 3D tooth shape to the biomechanical properties of their prey. Evolution 2021, 75, 1377–1394. [Google Scholar] [CrossRef]
- Marshall, T.L.; Chambers, E.A.; Matz, M.V.; Hillis, D.M. How mitonuclear discordance and geographic variation have confounded species boundaries in a widely studied snake. Mol. Phylogenet. Evol. 2021, 162, 107194. [Google Scholar] [CrossRef]
- Margres, M.; McGivern, J.; Wray, K.; Seavy, M.; Calvin, K.; Rokyta, D. Linking the transcriptome and proteome to characterize the venom of the eastern diamondback rattlesnake (Crotalus adamanteus). J. Proteom. 2014, 96, 145–158. [Google Scholar] [CrossRef]
- Margres, M.; McGivern, J.; Seavy, M.; Wray, K.; Facente, J.; Rokyta, D. Contrasting modes and tempos of venom expression evolution in two snake species. Genetics 2015, 199, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Campos, P.; Gilbert, T. DNA Extraction from Keratin and Chitin. Methods Mol. Biol. (Clifton N.J.) 2012, 840, 43–49. [Google Scholar] [CrossRef]
- Wooldridge, B.J.; Pineda, G.; Banuelas-Ornelas, J.J.; Dagda, R.K.; Gasanov, S.E.; Rael, E.D.; Lieb, C.S. Mojave rattlesnakes (Crotalus scutulatus scutulatus) lacking the acidic subunit DNA sequence lack Mojave toxin in their venom. Comp. Biochem. Phys. B 2001, 130, 169–179. [Google Scholar] [CrossRef]
- Lombarte, A.; Lleonart, J. Otolith size changes related with body growth, habitat depth and temperature. Environ. Biol. Fish. 1993, 37, 297–306. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R.J. Package ‘dismo’. Circles 2017, 9, 1–68. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hammerson, G. Crotalus horridus. The IUCN Red List of Threatened Species 2007: e.T64318A12765920. 2007. Available online: http://dx.doi.org/10.2305/IUCN.UK.2007.RLTS.T64318A12765920.en (accessed on 22 September 2021).
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Glor, R.E.; Huron, N.A.; Simões, M.; Iglesias, T.L.; et al. ENMTools 1.0: An R package for comparative ecological biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Lawson, R.; Slowinski, J. Mitochondrial DNA phylogeography of the polytypic North American rat snake (Elaphe obsoleta): A critique of the subspecies concept. Evolution 2000, 54, 2107–2118. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; pp. 1–8. [Google Scholar]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, A.M.; Lo, N.; Ho, S.Y. The impact of the tree prior on molecular dating of data sets containing a mixture of inter-and intraspecies sampling. Syst. Biol. 2017, 66, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.; Ho, S.; Phillips, M.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef]
- Ho, S. Calibrating molecular estimates of substitution rates and divergence times in birds. J. Avian Biol. 2007, 38, 409–414. [Google Scholar] [CrossRef]
- Holman, J. Fossil Snakes of North America: Origin, Evolution, Distribution, Paleoecology; Indiana University Press: Bloomington, IN, USA, 2000. [Google Scholar]
- Rambaut, A.; Drummond, A. Tracer v1.4. 2007. Available online: http://beast.bio.ed.ac.uk (accessed on 22 September 2021).
- Bert, T. Speciation in western Atlantic stone crabs (genus Menippe): The role of geological processes and climatic events in the formation and distribution of species. Mar. Biol. 1986, 93, 157–170. [Google Scholar] [CrossRef]
- Randazzo, A.; Jones, D. (Eds.) The Geology of Florida; University Press of Florida: Gainesville, FL, USA, 1997. [Google Scholar]
- Wang, I.J.; Bradburd, G.S. Isolation by environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef]
- Harrison, C.M.; Colbert, J.; Richter, C.J.; McDonald, P.J.; Trumbull, L.M.; Ellsworth, S.A.; Hogan, M.P.; Rokyta, D.R.; Margres, M.J. Using morphological, genetic, and venomic analyses to present current and historic evidence of Crotalus horridus × Crotalus adamanteus hybridization on Jekyll Island GA. Southeast. Nat. 2021, in press. [Google Scholar]
- Holding, M.L.; Margres, M.J.; Rokyta, D.R.; Gibbs, H.L. Local prey community composition and genetic distance predict venom divergence among populations of the Northern Pacific rattlesnake (Crotalus oreganus). J. Evol. Biol. 2018, 31, 1513–1528. [Google Scholar] [CrossRef]
- Babcock, H.L. Food habits of the timber rattlesnake. Bull. Boston Soc. Nat. Hist. 1929, 51, 12–14. [Google Scholar]
- King, W. A Survey of the Herpetology of Great Smoky Mountains National Park. Am. Midl. Nat. 1939, 21, 531–582. [Google Scholar] [CrossRef]
- Uhler, F.M.; Cottam, C.; Clarke, T.E. Food of snakes of the George Washington National Forest, Virginia. In Proceedings of the Fourth National American Wildlife Conference, Detroit, MI, USA, 13–15 February 1939; pp. 605–662. [Google Scholar]
- Smyth, T. Notes on the Timber Rattlesnake at Mountain Lake, Virginia. Copeia 1949, 1949, 78. [Google Scholar] [CrossRef]
- Barbour, R.W. The Reptiles of Big Black Mountain, Harlan County, Kentucky. Copeia 1950, 1950, 100–107. [Google Scholar] [CrossRef]
- Martin, J.R.; Wood, J.T. Notes on the Poisonous Snakes of the Dismal Swamp Area. Herpetologica 1955, 11, 237–238. [Google Scholar]
- Hamilton, W.J.; Pollack, J.A. The Food of Some Colubrid Snakes from Fort-Benning, Georgia. Ecology 1956, 37, 519–526. [Google Scholar] [CrossRef]
- Bush, F.M. Foods of Some Kentucky Herptiles. Herpetologica 1959, 15, 73–77. [Google Scholar]
- Keenlyne, K.D. Sexual Differences in Feeding Habits of Crotalus horridus horridus. J. Herpetol. 1972, 6, 234–237. [Google Scholar] [CrossRef]
- Brown, E.E. Some Snake Food Records From The Carolinas, USA. Brimleyana 1979, 1, 113–124. [Google Scholar]
- Fitch, H.S. Resources of a Snake Community in Prairie-Woodland Habitat of Northeastern Kansas. In Herpetological Communities: A Symposium of the Society for the Study of Amphibians and Reptiles and the Herpetologists’ League, August 1977; Scott, N.J., Jr., Ed.; U.S. Fish and Wildlife Service: Washington, DC, USA, 1982; pp. 83–97. [Google Scholar]
- Reinert, H.K.; Cundall, D.; Bushar, L.M. Foraging Behavior of the Timber Rattlesnake, Crotalus horridus. Copeia 1984, 1984, 976–981. [Google Scholar] [CrossRef]
- Palmer, W.M.; Braswell, A.L. Reptiles of North Carolina; The University of North Carolina Press: Chapel Hill, NC, USA, 1995. [Google Scholar]
- Trauth, S.E.; McAllister, C.T.; Worth, F. Vertebrate Prey of Selected Arkansas Snakes. J. Ark. Acad. Sci. 1995, 49, 188–192. [Google Scholar]
- Parmley, D.; Parmley, A.M. Food habits of the Canebrake Rattlesnake (Crotalus horridus atricaudatus) in Central Georgia. Ga. J. Sci. 2001, 59, 172. [Google Scholar]
- Sajdak, R.A.; Bartz, A.W. Crotalus horridus (timber rattlesnake). Arboreality, diet. Herpetol. Rev. 2004, 35, 60–61. [Google Scholar]
- Wittenberg, R.D. Foraging Ecology of the Timber Rattlesnake (Crotalus horridus) in a Fragmented Agricultural Landscape. Herpetol. Conserv. Biol. 2012, 7, 449–461. [Google Scholar]
- Hammerstein, G.; Steen, D.A.; Stevenson, D.J. Crotalus horridus (timber rattlesnake). Diet. Herpetol. Rev. 2015, 46, 640–641. [Google Scholar]
- Goetz, S.M.; Petersen, C.E.; Rose, R.K.; Kleopfer, J.D.; Savitzky, A.H. Diet and Foraging Behaviors of Timber Rattlesnakes, Crotalus horridus, in Eastern Virginia. J. Herpetol. 2016, 50, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Schalk, C.M.; Trees, T.; Pierce, J.B.; Rudolph, D.C. Food habits of sympatric pitvipers from the West Gulf Coastal Plain, USA. Herpetol. Rev. 2018, 49, 1–5. [Google Scholar]
- Hewlett, J. Crotalus horridus (timber rattlesnake). Diet. Herpetol. Rev. 2019, 50, 589–590. [Google Scholar]
- Pynne, J.T.; Castleberry, S.B.; Conner, L.M.; Parsons, E.I.; Gitzen, R.A.; Austin, J.D.; Duncan, S.I.; McCleery, R.A. Timber Rattlesnake (Crotalus horridus) Predation on a Southeastern Pocket Gopher (Geomys pinetis). Southeast. Nat. 2019, 18, N34–N36. [Google Scholar] [CrossRef]
- Rautsaw, R.M.; Schramer, T.D.; Acuña, R.; Arick, L.N.; DiMeo, M.; Mercier, K.P.; Schrum, M.; Mason, A.J.; Margres, M.J.; Strickland, J.L.; et al. Genomic Adaptations to Salinity Resist Gene Flow in the Evolution of Floridian Watersnakes. Mol. Biol. Evol. 2021, 38, 745–760. [Google Scholar] [CrossRef]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Nosil, P. Speciation with gene flow could be common. Mol. Ecol. 2008, 17, 2103–2106. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margres, M.J.; Wray, K.P.; Sanader, D.; McDonald, P.J.; Trumbull, L.M.; Patton, A.H.; Rokyta, D.R. Varying Intensities of Introgression Obscure Incipient Venom-Associated Speciation in the Timber Rattlesnake (Crotalus horridus). Toxins 2021, 13, 782. https://doi.org/10.3390/toxins13110782
Margres MJ, Wray KP, Sanader D, McDonald PJ, Trumbull LM, Patton AH, Rokyta DR. Varying Intensities of Introgression Obscure Incipient Venom-Associated Speciation in the Timber Rattlesnake (Crotalus horridus). Toxins. 2021; 13(11):782. https://doi.org/10.3390/toxins13110782
Chicago/Turabian StyleMargres, Mark J., Kenneth P. Wray, Dragana Sanader, Preston J. McDonald, Lauren M. Trumbull, Austin H. Patton, and Darin R. Rokyta. 2021. "Varying Intensities of Introgression Obscure Incipient Venom-Associated Speciation in the Timber Rattlesnake (Crotalus horridus)" Toxins 13, no. 11: 782. https://doi.org/10.3390/toxins13110782
APA StyleMargres, M. J., Wray, K. P., Sanader, D., McDonald, P. J., Trumbull, L. M., Patton, A. H., & Rokyta, D. R. (2021). Varying Intensities of Introgression Obscure Incipient Venom-Associated Speciation in the Timber Rattlesnake (Crotalus horridus). Toxins, 13(11), 782. https://doi.org/10.3390/toxins13110782