Isolation and Characterization of A2-EPTX-Nsm1a, a Secretory Phospholipase A2 from Malaysian Spitting Cobra (Naja sumatrana) Venom


 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
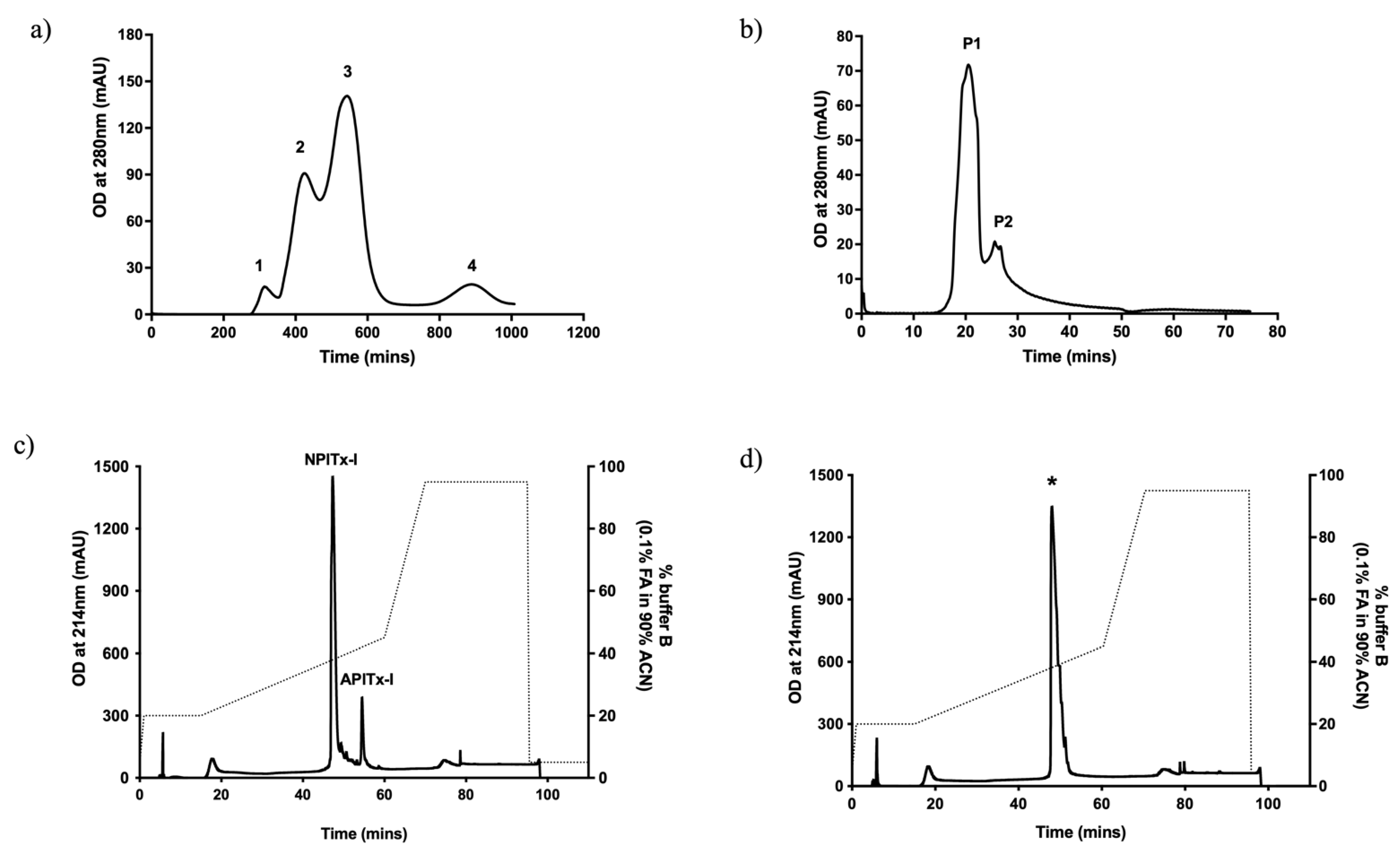
2.1. Purification of A2-EPTX-Nsm1a
2.2. SDS-PAGE
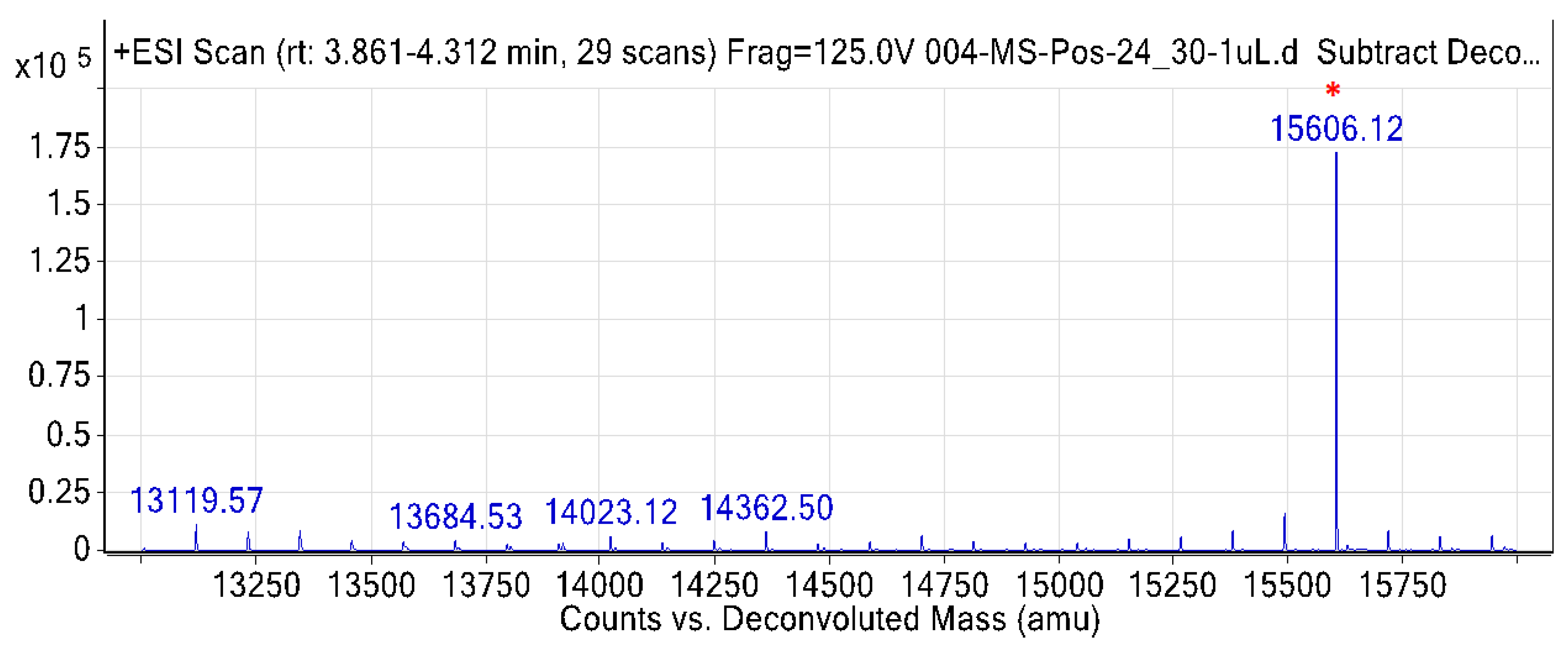
2.3. Intact Protein Using Accurate Mass LC-MS
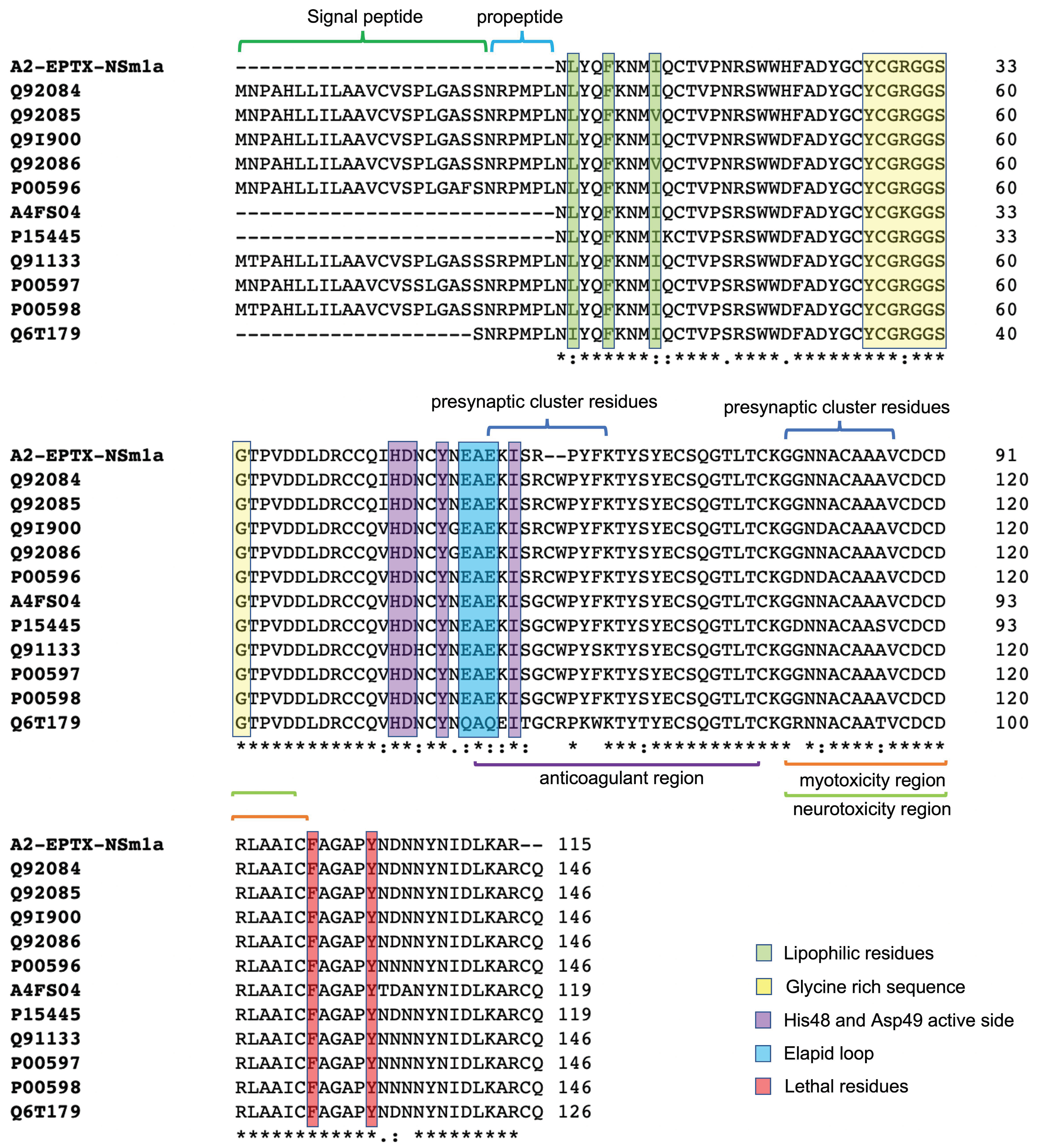
2.4. Protein Identification by ESI-LC-MSMS
2.5. Molecular Modelling
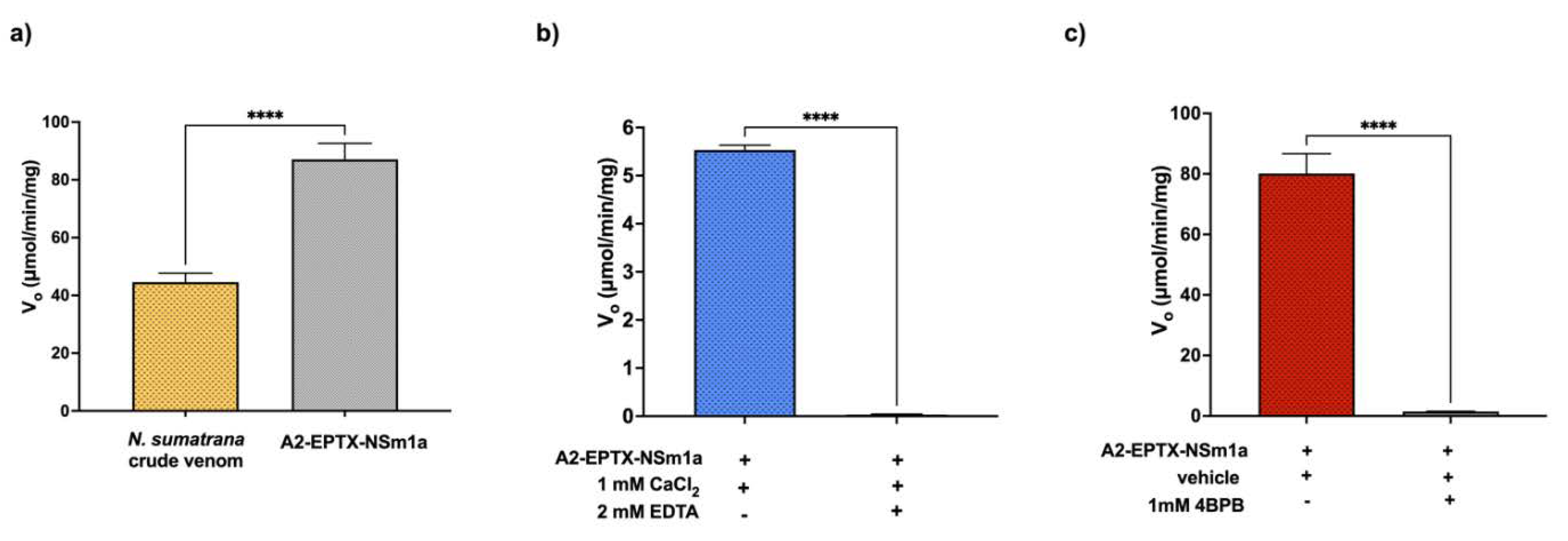
2.6. PLA2 Activity
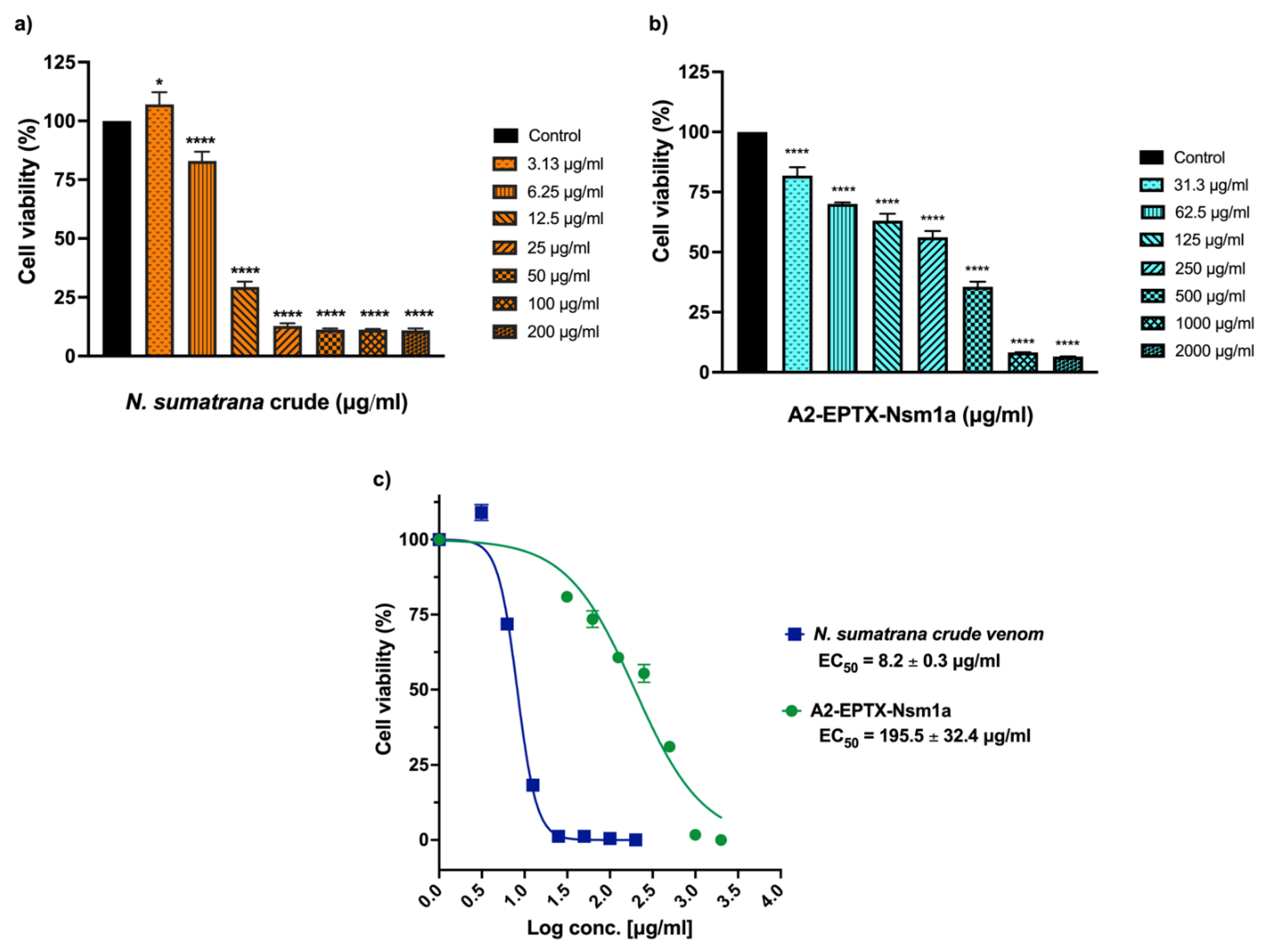
2.7. Cytotoxicity Activity of A2-EPTX-NSm1a on SH-SY5Y
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Crude Venom
5.3. Purification of A2-EPTX-Nsm1a
5.4. Protein Quantification by Bicinchoninic Acid (BCA) Assay
5.5. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
5.6. Intact Protein Analysis with Electrospray-Ionisation Coupled with Mass-Spectrometry
5.7. In-Gel Tryptic Digestion
5.8. Protein Identification with Tandem Mass Spectrometry (ESI-LCMS/MS)
5.9. Automated De Novo Sequencing
5.10. Molecular Modelling
5.11. PLA2 Activity
5.12. PLA2 Inhibition by 4-Bromophenacyl Bromide (4-BPB) and EDTA
5.13. Cell Culture
5.14. Cytotoxicity of A2-EPTX-Nsm1a on SH-SY5Y cells
5.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, I. Field Guide to the Reptiles of South-East Asia; Bloomsbury Publishing: London, UK, 2015; p. 384. [Google Scholar]
- Stuebing, R.B.; Inger, R.F.; Tan, F.L. Field Guide to the Snakes of Borneo; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 1999; p. 254. [Google Scholar]
- WHO. Guidelines for the Management of Snake Bites; Regional office for South-East Asia, World Health Organization: New Delhi, India, 2016. [Google Scholar]
- Ismail, A.K. Snakebite and envenomation management in Malaysia. Clin. Toxicol Asia Pac. Afr. 2015, 2, 71–102. [Google Scholar] [CrossRef]
- Yap, M.K.; Fung, S.Y.; Tan, K.Y.; Tan, N.H. Proteomic characterization of venom of the medically important Southeast Asian Naja sumatrana (Equatorial spitting cobra). Acta Trop. 2014, 133, 15–25. [Google Scholar] [CrossRef]
- Fry, B.G.; Scheib, H.; Van Der Weerd, L.; Young, B.; McNaughtan, J.; Ramjan, S.F.R.; Vidal, N.; Poelmann, R.E.; Norman, J.A. Evolution of an Arsenal: Structural and functional diversification of the venom system in the advanced snakes (Caenophidia). Mol. Cell. Proteom. 2008, 7, 215–246. [Google Scholar] [CrossRef] [Green Version]
- Petan, T.; Krizaj, I.; Pungercar, J. Restoration of enzymatic activity in a Ser-49 phospholipase A2 homologue decreases its Ca2+-independent membrane-damaging activity and increases its toxicity. Biochemistry 2007, 46, 12795–12809. [Google Scholar] [CrossRef]
- Murakami, T.; Kariu, T.; Takazaki, S.; Hattori, S.; Chijiwa, T.; Ohno, M.; Oda-Ueda, N. Island specific expression of a novel [Lys49] phospholipase A2 (BPIII) in Protobothrops flavoviridis venom in Amami–Oshima, Japan. Toxicon 2009, 54, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Resende, L.M.; Almeida, J.R.; Schezaro-Ramos, R.; Collaço, R.C.O.; Simioni, L.R.; Ramírez, D.; González, W.; Soares, A.M.; Calderon, L.A.; Marangoni, S.; et al. Exploring and understanding the functional role, and biochemical and structural characteristics of an acidic phospholipase A2, AplTx-I, purified from Agkistrodon piscivorus leucostoma snake venom. Toxicon 2017, 127, 22–36. [Google Scholar] [CrossRef]
- Jiménez–Charris, E.; Lopes, D.; Gimenes, S.; Teixeira, S.C.; Montealegre–Sánchez, L.; Solano–Redondo, L.; Fierro–Pérez, L.; Ávila, V.D.M.R. Antitumor potential of Pllans–II, an acidic Asp49–PLA2 from Porthidium lansbergii lansbergii snake venom on human cervical carcinoma HeLa cells. Int. J. Biol. Macromol. 2019, 122, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Salvador, G.H.; Cavalcante, W.L.; dos Santos, J.I.; Gallacci, M.; Soares, A.M.; Fontes, M.R. Structural and functional studies with mytoxin II from Bothrops moojeni reveal remarkable similarities and differences compared to other catalytically inactive phospholipases A2-like. Toxicon 2013, 72, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Clement, H.; De Oliveira, V.C.; Zamudio, F.Z.; Lago, N.R.; Valdez-Cruz, N.A.; Valle, M.B.; Hajos, S.E.; Alagón, A.; Possani, L.D.; De Roodt, A.R. Isolation, amino acid sequence and biological characterization of an “aspartic-49” phospholipase A2 from Bothrops (Rhinocerophis) ammodytoides venom. Toxicon 2012, 60, 1314–1323. [Google Scholar] [CrossRef]
- Bustillo, S.; Gay, C.C.; Denegri, M.E.G.; Ponce-Soto, L.A.; Joffé, E.B.D.K.; Acosta, O.; Leiva, L.C. Synergism between baltergin metalloproteinase and Ba SPII RP4 PLA2 from Bothrops alternatus venom on skeletal muscle (C2C12) cells. Toxicon 2012, 59, 338–343. [Google Scholar] [CrossRef]
- Ullah, A.; Souza, T.; Betzel, C.; Murakami, M.; Arni, R. Crystallographic portrayal of different conformational states of a Lys49 phospholipase A2 homologue: Insights into structural determinants for myotoxicity and dimeric configuration. Int. J. Biol. Macromol. 2012, 51, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huancahuire-Vega, S.; Ponce-Soto, L.A.; Martins-De-Souza, D.; Marangoni, S. Biochemical and pharmacological characterization of PhTX-I a new myotoxic phospholipase A2 isolated from Porthidium hyoprora snake venom. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 154, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Pereañez, J.A.; Núñez, V.; Huancahuire-Vega, S.; Marangoni, S.; Ponce-Soto, L.A. Biochemical and biological characterization of a PLA2 from crotoxin complex of Crotalus durissus cumanensis. Toxicon 2009, 53, 534–542. [Google Scholar] [CrossRef]
- Marques, P.P.; Esteves, A.; Lancellotti, M.; Ponce-Soto, L.A.; Marangoni, S. Novel acidic phospholipase A2 from Porthidium hyoprora causes inflammation with mast cell rich infiltrate. Biochem. Biophys. Rep. 2015, 1, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Nunes, D.C.; Rodrigues, R.S.; Lucena, M.N.; Cologna, C.T.; Oliveira, A.C.S.; Hamaguchi, A.; Homsi-Brandeburgo, M.I.; Arantes, E.C.; Teixeira, D.N.; Ueira-Vieira, C.; et al. Isolation and functional characterization of proinflammatory acidic phospholipase A2 from Bothrops leucurus snake venom. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 154, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Toyama, D.d.O.; Diz Filho, E.B.d.S.; Cavada, B.S.; da Rocha, B.A.M.; de Oliveira, S.C.B.; Cotrim, C.A.; Soares, V.C.G.; Delatorre, P.; Marangoni, S.; Toyama, M.H. Umbelliferone induces changes in the structure and pharmacological activities of Bn IV, a phospholipase A2 isoform isolated from Bothrops neuwiedi. Toxicon 2011, 57, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.-F.; Wei, X.-L.; Mo, Y.-Z.; He, S.-H. Induction of microvascular leakage and histamine release by promutoxin, an Arg49 phospholipase A2. Toxicon 2010, 55, 888–896. [Google Scholar] [CrossRef]
- Toyama, D.; Marangoni, S.; Diz-Filho, E.; Oliveira, S.; Toyama, M. Effect of umbelliferone (7-hydroxycoumarin, 7-HOC) on the enzymatic, edematogenic and necrotic activities of secretory phospholipase A2 (sPLA2) isolated from Crotalus durissus collilineatus venom. Toxicon 2009, 53, 417–426. [Google Scholar] [CrossRef]
- de Carvalho, N.D.; Garcia, R.C.; Kleber Ferreira, A.; Rodrigo Batista, D.; Carlos Cassola, A.; Maria, D.; Lebrun, I.; Mendes Carneiro, S.; Castro Afeche, S.; Marcourakis, T.; et al. Neurotoxicity of coral snake phospholipases A2 in cultured rat hippocampal neurons. Brain Res. 2014, 1552, 1–16. [Google Scholar] [CrossRef]
- Vergara, I.; Pedraza-Escalona, M.; Paniagua, D.; Restano-Cassulini, R.; Zamudio, F.; Batista, C.V.F.; Possani, L.D.; Alagón, A. Eastern coral snake Micrurus fulvius venom toxicity in mice is mainly determined by neurotoxic phospholipases A2. J. Proteom. 2014, 105, 295–306. [Google Scholar] [CrossRef]
- Lomeo, R.D.S.; Gonçalves, A.P.D.F.; da Silva, C.N.; de Paula, A.T.; Santos, D.O.C.; Fortes-Dias, C.L.; Gomes, D.; de Lima, M.E. Crotoxin from Crotalus durissus terrificus snake venom induces the release of glutamate from cerebrocortical synaptosomes via N and P/Q calcium channels. Toxicon 2014, 85, 5–16. [Google Scholar] [CrossRef]
- Blacklow, B.; Escoubas, P.; Nicholson, G.M. Characterisation of the heterotrimeric presynaptic phospholipase A2 neurotoxin complex from the venom of the common death adder (Acanthophis antarcticus). Biochem. Pharmacol. 2010, 80, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Menezes, T.N.; Naumann, G.B.; Peixoto, P.; Rouver, W.N.; Gomes, H.L.; Campos, F.V.; Borges, M.H.; dos Santos, R.L.; Bissoli, N.S.; Sanchez, E.F.; et al. Bothrops leucurus venom induces acute hypotension in rats by means of its phospholipase A2 (blD-PLA2). Toxicon 2020, 185, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Zhang, W.; Xue, N.; Teng, M.; Li, X.; Shen, B. Crystal structure of phospholipase PA2-Vb, a protease-activated receptor agonist from the Trimeresurus stejnegeri snake venom. FEBS Lett. 2014, 588, 4604–4612. [Google Scholar] [CrossRef] [Green Version]
- Saikia, D.; Majumdar, S.; Mukherjee, A.K. Mechanism of in vivo anticoagulant and haemolytic activity by a neutral phospholipase A2 purified from Daboia russelii russelii venom: Correlation with clinical manifestations in Russell’s Viper envenomed patients. Toxicon 2013, 76, 291–300. [Google Scholar] [CrossRef]
- Marinho, A.D.; Silveira, J.A.M.; Chaves Filho, A.J.M.; Jorge, A.R.C.; Nogueira Junior, F.A.; Pereira, V.B.M.; de Aquino, P.E.A.; Pereira, C.A.S.; Evangelista, J.; Macedo, D.S.; et al. Bothrops pauloensis snake venom-derived Asp-49 and Lys-49 phospholipases A2 mediates acute kidney injury by oxidative stress and release of inflammatory cytokines. Toxicon 2021, 190, 31–38. [Google Scholar] [CrossRef] [PubMed]
- de Vasconcelos Azevedo, F.V.P.; Zóia, M.A.P.; Lopes, D.S.; Gimenes, S.N.; Vecchi, L.; Alves, P.T.; Rodrigues, R.S.; Silva, A.C.A.; Yoneyama, K.A.G.; Goulart, L.R.; et al. Antitumor and antimetastatic effects of PLA2-BthTX-II from Bothrops jararacussu venom on human breast cancer cells. Int. J. Biol. Macromol. 2019, 135, 261–273. [Google Scholar] [CrossRef]
- Silva, M.A.; Lopes, D.S.; Teixeira, S.C.; Gimenes, S.N.C.; Azevedo, F.V.P.V.; Polloni, L.; Borges, B.C.; da Silva, M.S.; Barbosa, M.J.; Oliveira Júnior, R.J.d.; et al. Genotoxic effects of BnSP-6, a Lys-49 phospholipase A2 (PLA2) homologue from Bothrops pauloensis snake venom, on MDA-MB-231 breast cancer cells. Int. J. Biol. Macromol. 2018, 118, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, F.V.P.V.; Lopes, D.S.; Cirilo Gimenes, S.N.; Achê, D.C.; Vecchi, L.; Alves, P.T.; Guimarães, D.d.O.; Rodrigues, R.S.; Goulart, L.R.; Rodrigues, V.d.M.; et al. Human breast cancer cell death induced by BnSP-6, a Lys-49 PLA2 homologue from Bothrops pauloensis venom. Int. J. Biol. Macromol. 2016, 82, 671–677. [Google Scholar] [CrossRef]
- Bazaa, A.; Luis, J.; Srairi-Abid, N.; Kallech-Ziri, O.; Kessentini-Zouari, R.; Defilles, C.; Lissitzky, J.-C.; El Ayeb, M.; Marrakchi, N. MVL-PLA2, a phospholipase A2 from Macrovipera lebetina transmediterranea venom, inhibits tumor cells adhesion and migration. Matrix Biol. 2009, 28, 188–193. [Google Scholar] [CrossRef]
- Zouari-Kessentini, R.; Luis, J.; Karray, A.; Kallech-Ziri, O.; Srairi-Abid, N.; Bazaa, A.; Loret, E.; Bezzine, S.; El Ayeb, M.; Marrakchi, N. Two purified and characterized phospholipases A2 from Cerastes cerastes venom, that inhibit cancerous cell adhesion and migration. Toxicon 2009, 53, 444–453. [Google Scholar] [CrossRef]
- Liu, W.-H.; Kao, P.-H.; Chiou, Y.-L.; Lin, S.-r.; Wu, M.-J.; Chang, L.-S. Catalytic activity-independent pathway is involved in phospholipase A2-induced apoptotic death of human leukemia U937 cells via Ca2+-mediated p38 MAPK activation and mitochondrial depolarization. Toxicol. Lett. 2009, 185, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Polloni, L.; Azevedo, F.V.P.V.; Teixeira, S.C.; Moura, E.; Costa, T.R.; Gimenes, S.N.C.; Correia, L.I.V.; Freitas, V.; Yoneyama, K.A.G.; Rodrigues, R.S.; et al. Antiangiogenic effects of phospholipase A2 Lys49 BnSP-7 from Bothrops pauloensis snake venom on endothelial cells: An in vitro and ex vivo approach. Toxicol. Vitr. 2021, 72, 105099. [Google Scholar] [CrossRef]
- Santos-Filho, N.A.; de Freitas, L.M.; dos Santos, C.T.; Piccoli, J.P.; Fontana, C.R.; Fusco-Almeida, A.M.; Cilli, E.M. Understanding the mechanism of action of peptide (p-BthTX-I)2 derived from C-terminal region of phospholipase A2 (PLA2)-like bothropstoxin-I on Gram-positive and Gram-negative bacteria. Toxicon 2021, 196, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Diz Filho, E.B.S.; Marangoni, S.; Toyama, D.O.; Fagundes, F.H.R.; Oliveira, S.C.B.; Fonseca, F.V.; Calgarotto, A.K.; Joazeiro, P.P.; Toyama, M.H. Enzymatic and structural characterization of new PLA2 isoform isolated from white venom of Crotalus durissus ruruima. Toxicon 2009, 53, 104–114. [Google Scholar] [CrossRef]
- de Barros, N.B.; Macedo, S.R.A.; Ferreira, A.S.; Tagliari, M.P.; Zanchi, F.B.; Kayano, A.M.; Soares, A.M.; Nicolete, R. Liposomes containing an ASP49-phospholipase A2 from Bothrops jararacussu snake venom as experimental therapy against cutaneous leishmaniasis. Int. Immunopharmacol. 2016, 36, 225–231. [Google Scholar] [CrossRef]
- Damico, D.C.S.; Vassequi-Silva, T.; Torres-Huaco, F.D.; Nery-Diez, A.C.C.; de Souza, R.C.G.; Da Silva, S.L.; Vicente, C.P.; Mendes, C.B.; Antunes, E.; Werneck, C.C.; et al. LmrTX, a basic PLA2 (D49) purified from Lachesis muta rhombeata snake venom with enzymatic-related antithrombotic and anticoagulant activity. Toxicon 2012, 60, 773–781. [Google Scholar] [CrossRef] [Green Version]
- Saikia, D.; Thakur, R.; Mukherjee, A.K. An acidic phospholipase A2 (RVVA-PLA2-I) purified from Daboia russelli venom exerts its anticoagulant activity by enzymatic hydrolysis of plasma phospholipids and by non-enzymatic inhibition of factor Xa in a phospholipids/Ca2+ independent manner. Toxicon 2011, 57, 841–850. [Google Scholar] [CrossRef]
- Muller, V.D.M.; Russo, R.R.; Oliveira Cintra, A.C.; Sartim, M.A.; De Melo Alves-Paiva, R.; Figueiredo, L.T.M.; Sampaio, S.V.; Aquino, V.H. Crotoxin and phospholipases A2 from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon 2012, 59, 507–515. [Google Scholar] [CrossRef]
- da Silva Cunha, K.C.; Fuly, A.L.; Giestal de Araujo, E. A phospholipase A2 isolated from Lachesis muta snake venom increases the survival of retinal ganglion cells in vitro. Toxicon 2011, 57, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Vija, H.; Samel, M.; Siigur, E.; Aaspõllu, A.; Trummal, K.; Tõnismägi, K.; Subbi, J.; Siigur, J. Purification, characterization, and cDNA cloning of acidic platelet aggregation inhibiting phospholipases A2 from the snake venom of Vipera lebetina (Levantine viper). Toxicon 2009, 54, 429–439. [Google Scholar] [CrossRef]
- Hiu, J.J.; Yap, M.K.K. Cytotoxicity of snake venom enzymatic toxins: Phospholipase A2 and l-amino acid oxidase. Biochem. Soc. Trans. 2020, 48, 719–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armugam, A.; Earnest, L.; Chung, M.C.; Gopalakrishnakone, P.; Tan, C.H.; Tan, N.H.; Jeyaseelan, K. Cloning and characterization of cDNAs encoding three isoforms of phospholipase A2 in Malayan spitting cobra (Naja naja sputatrix) venom. Toxicon 1997, 35, 27–37. [Google Scholar] [CrossRef]
- Jeyaseelan, K.; Armugam, A.; Donghui, M.; Tan, N.H. Structure and phylogeny of the venom group I phospholipase A2 gene. Mol. Biol. Evol. 2000, 17, 1010–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, S.-I.; Tu, A.T. Phospholipase A2 from Naja naja sputatrix venom is a muscarinic acetylcholine receptor inhibitor. Arch. Biochem. Biophys. 1996, 328, 17–25. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.S.; Fuentes-Silva, D.; King, G.F. Development of a rational nomenclature for naming peptide and protein toxins from sea anemones. Toxicon 2012, 60, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.Y.; Tan, C.H.; Chanhome, L.; Tan, N.H. Comparative venom gland transcriptomics of Naja kaouthia (monocled cobra) from Malaysia and Thailand: Elucidating geographical venom variation and insights into sequence novelty. PeerJ 2017, 5, e3142. [Google Scholar] [CrossRef] [Green Version]
- Chong, H.P.; Tan, K.Y.; Tan, N.H.; Tan, C.H. Exploring the Diversity and Novelty of Toxin Genes in Naja sumatrana, the Equatorial Spitting Cobra from Malaysia through De Novo Venom-Gland Transcriptomics. Toxins 2019, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Kazandjian, T.D.; Petras, D.; Robinson, S.D.; van Thiel, J.; Greene, H.W.; Arbuckle, K.; Barlow, A.; Carter, D.A.; Wouters, R.M.; Whiteley, G.; et al. Convergent evolution of pain-inducing defensive venom components in spitting cobras. Science 2021, 371, 386–390. [Google Scholar] [CrossRef]
- Serino-Silva, C.; Morais-Zani, K.; Hikari Toyama, M.; Toyama, D.O.; Gaeta, H.H.; Rodrigues, C.F.B.; Aguiar, W.D.S.; Tashima, A.K.; Grego, K.F.; Tanaka-Azevedo, A.M. Purification and characterization of the first γ-phospholipase inhibitor (γPLI) from Bothrops jararaca snake serum. PLoS ONE 2018, 13, e0193105. [Google Scholar] [CrossRef] [Green Version]
- Torres-Huaco, F.D.; Marunak, S.; Teibler, P.; Bustillo, S.; Acosta de Perez, O.; Leiva, L.C.; Ponce-Soto, L.A.; Marangoni, S. Local and systemic effects of BtaMP-1, a new weakly hemorrhagic snake venom metalloproteinase purified from Bothriopsis taeniata Snake Venom. Int. J. Biol. Macromol. 2019, 141, 1044–1054. [Google Scholar] [CrossRef]
- Vivas-Ruiz, D.E.; Gonzalez-Kozlova, E.E.; Delgadillo, J.; Palermo, P.M.; Sandoval, G.A.; Lazo, F.; Rodriguez, E.; Chavez-Olortegui, C.; Yarleque, A.; Sanchez, E.F. Biochemical and molecular characterization of the hyaluronidase from Bothrops atrox Peruvian snake venom. Biochimie 2019, 162, 33–45. [Google Scholar] [CrossRef]
- Rivas Mercado, E.; Neri Castro, E.; Benard Valle, M.; Rucavado-Romero, A.; Olvera Rodriguez, A.; Zamudio Zuniga, F.; Alagon Cano, A.; Garza Ocanas, L. Disintegrins extracted from totonacan rattlesnake (Crotalus totonacus) venom and their anti-adhesive and anti-migration effects on MDA-MB-231 and HMEC-1 cells. Toxicol. Vitr. 2020, 65, 104809. [Google Scholar] [CrossRef]
- Dennis, E.A. Diversity of group types, regulation, and function of phospholipase A2. J. Biol. Chem. 1994, 269, 13057–13060. [Google Scholar] [CrossRef]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasquez, A.M.; Mouchlis, V.D.; Dennis, E.A. Review of four major distinct types of human phospholipase A2. Adv. Biol. Regul. 2018, 67, 212–218. [Google Scholar] [CrossRef]
- Filkin, S.Y.; Lipkin, A.V.; Fedorov, A.N. Phospholipase Superfamily: Structure, Functions, and Biotechnological Applications. Biochemistry 2020, 85, S177–S195. [Google Scholar] [CrossRef]
- Soares, A.M.; Giglio, J.R. Chemical modifications of phospholipases A2 from snake venoms: Effects on catalytic and pharmacological properties. Toxicon 2003, 42, 855–868. [Google Scholar] [CrossRef]
- Kudo, I.; Murakami, M. Phospholipase A2 enzymes. Prostaglandins Other Lipid Mediat. 2002, 68–69, 3–58. [Google Scholar] [CrossRef]
- Corasolla Carregari, V.; Stuani Floriano, R.; Rodrigues-Simioni, L.; Winck, F.V.; Baldasso, P.A.; Ponce-Soto, L.A.; Marangoni, S. Biochemical, pharmacological, and structural characterization of new basic PLA2 Bbil-TX from Bothriopsis bilineata snake venom. BioMed Res. Int. 2013, 2013, 612649. [Google Scholar] [CrossRef] [Green Version]
- van den Bergh, C.J.; Slotboom, A.J.; Verheij, H.M.; de Haas, G.H. The role of Asp-49 and other conserved amino acids in phospholipases A2 and their importance for enzymatic activity. J. Cell. Biochem. 1989, 39, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Kramer, R.M.; Hession, C.; Johansen, B.; Hayes, G.; McGray, P.; Chow, E.P.; Tizard, R.; Pepinsky, R.B. Structure and properties of a human non-pancreatic phospholipase A2. J. Biol. Chem. 1989, 264, 5768–5775. [Google Scholar] [CrossRef]
- Tomoo, K.; Ohishi, H.; Doi, M.; Ishida, T.; Inoue, M.; Ikeda, K.; Mizuno, H. Interaction mode of n-dodecylphosphorylcholine, a substrate analogue, with bovine pancreas phospholipase A2 as determined by X-ray crystal analysis. Biochem. Biophys. Res. Commun. 1992, 187, 821–827. [Google Scholar] [CrossRef]
- Yap, M.K.; Tan, N.H.; Sim, S.M.; Fung, S.Y.; Tan, C.H. Pharmacokinetics of Naja sumatrana (equatorial spitting cobra) venom and its major toxins in experimentally envenomed rabbits. PLoS Negl. Trop. Dis. 2014, 8, e2890. [Google Scholar] [CrossRef] [PubMed]
- Shina, R.; Yates, S.L.; Ghassemi, A.; Rosenberg, P.; Condrea, E. Inhibitory effect of EDTA · Ca2+ on the hydrolysis of synaptosomal phospholipids by phospholipase A2 toxins and enzymes. Biochem. Pharmacol. 1990, 40, 2233–2239. [Google Scholar] [CrossRef]
- Huancahuire-Vega, S.; Ponce-Soto, L.A.; Marangoni, S. PhTX-II a basic myotoxic phospholipase A2 from Porthidium hyoprora snake venom, pharmacological characterization and amino acid sequence by mass spectrometry. Toxins 2014, 6, 3077–3097. [Google Scholar] [CrossRef] [Green Version]
- Rusmili, M.R.; Yee, T.T.; Mustafa, M.R.; Hodgson, W.C.; Othman, I. Isolation and characterization of a presynaptic neurotoxin, P-elapitoxin-Bf1a from Malaysian Bungarus fasciatus venom. Biochem. Pharmacol. 2014, 91, 409–416. [Google Scholar] [CrossRef]
- Fuly, A.L.; Calil-Elias, S.; Zingali, R.B.; Guimarães, J.A.; Melo, P.A. Myotoxic activity of an acidic phospholipase A2 isolated from Lachesis muta (Bushmaster) snake venom. Toxicon 2000, 38, 961–972. [Google Scholar] [CrossRef]
- Xicoy, H.; Wieringa, B.; Martens, G.J. The SH-SY5Y cell line in Parkinson’s disease research: A systematic review. Mol. Neurodegener. 2017, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Kovalevich, J.; Langford, D. Considerations for the use of SH-SY5Y neuroblastoma cells in neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, R.A.; Spengler, B.A.; Biedler, J.L. Coordinate morphological and biochemical interconversion of human neuroblastoma cells. J. Natl. Cancer Inst. 1983, 71, 741–747. [Google Scholar] [CrossRef]
- Emanuelsson, I.; Norlin, M. Protective effects of 27- and 24-hydroxycholesterol against staurosporine-induced cell death in undifferentiated neuroblastoma SH-SY5Y cells. Neurosci. Lett. 2012, 525, 44–48. [Google Scholar] [CrossRef]
- Gill, I.; Kaur, S.; Kaur, N.; Dhiman, M.; Mantha, A.K. Phytochemical Ginkgolide B attenuates amyloid-β1-42 induced oxidative damage and altered cellular responses in human neuroblastoma SH-SY5Y cells. J. Alzheimers Dis. 2017, 60, S25–S40. [Google Scholar] [CrossRef] [PubMed]
- Jantas, D.; Greda, A.; Golda, S.; Korostynski, M.; Grygier, B.; Roman, A.; Pilc, A.; Lason, W. Neuroprotective effects of metabotropic glutamate receptor group II and III activators against MPP+-induced cell death in human neuroblastoma SH-SY5Y cells: The impact of cell differentiation state. Neuropharmacology 2014, 83, 36–53. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Rasà, D.M.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. PACAP and VIP regulate hypoxia-inducible factors in neuroblastoma cells exposed to hypoxia. Neuropeptides 2018, 69, 84–91. [Google Scholar] [CrossRef]
- Shipley, M.M.; Mangold, C.A.; Szpara, M.L. Differentiation of the SH-SY5Y human neuroblastoma cell line. J. Vis. Exp. 2016, 108, 53193. [Google Scholar] [CrossRef]
- Zhang, Y.; Anoopkumar-Dukie, S.; Mallik, S.B.; Davey, A.K. SIRT1 and SIRT2 modulators reduce LPS-induced inflammation in HAPI microglial cells and protect SH-SY5Y neuronal cells in vitro. J. Neural Transm. 2021, 128, 631–644. [Google Scholar] [CrossRef]
- Moreira, V.; Lomonte, B.; Vinolo, M.A.R.; Curi, R.; Gutiérrez, J.M.; Teixeira, C. An Asp49 phospholipase A2 snake venom induces cyclooxygenase-2 expression and prostaglandin E2 production via activation of NF-κB, p38MAPK, and PKC in macrophages. Mediat. Inflamm. 2014, 2014, 105879. [Google Scholar] [CrossRef] [Green Version]
- Bazaa, A.; Pasquier, E.; Defilles, C.; Limam, I.; Kessentini-Zouari, R.; Kallech-Ziri, O.; Battari, A.E.; Braguer, D.; Ayeb, M.E.; Marrakchi, N.; et al. MVL-PLA2, a snake venom phospholipase A2, inhibits angiogenesis through an increase in microtubule dynamics and disorganization of focal adhesions. PLoS ONE 2010, 5, e10124. [Google Scholar] [CrossRef] [Green Version]
- Manns, J.M. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE) of proteins. Curr. Protoc. Microbiol. 2011, 22, A.3M.1–A.3M.13. [Google Scholar] [CrossRef]
- Craig, D.B.; Dombkowski, A.A. Disulfide by Design 2.0: A web-based tool for disulfide engineering in proteins. BMC Bioinform. 2013, 14, 1471–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrè, F.; Clote, P. DiANNA: A web server for disulfide connectivity prediction. Nucleic Acids Res. 2005, 33, W230–W232. [Google Scholar] [CrossRef] [Green Version]
- Fariselli, P.; Riccobelli, P.; Casadio, R. Role of evolutionary information in predicting the disulfide-bonding state of cysteine in proteins. Proteins 1999, 36, 340–346. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | −10lgP | Coverage (%) | No. of Peptides | Average Mass | Description | Origin | Identity (%) |
|---|---|---|---|---|---|---|---|
| Q92084 | 233.5 | 76 | 17 | 16,189 | Neutral phospholipase A2 muscarinic inhibitor | Naja sputatrix | 97.8 |
| Q92085 | 224.51 | 72 | 15 | 16,175 | Neutral phospholipase A2 B | Naja sputatrix | 97.4 |
| Q9I900 | 200.54 | 74 | 12 | 16,097 | Acidic phospholipase A2 D | Naja sputatrix | 94.9 |
| Q92086 | 198.43 | 70 | 11 | 16,082 | Acidic phospholipase A2 C | Naja sputatrix | 94.9 |
| P00596 | 184.86 | 64 | 10 | 16,271 | Acidic phospholipase A2 1 | Naja kaouthia | 94 |
| P00598 | 179.51 | 63 | 8 | 16,013 | Acidic phospholipase A2 1 | Naja atra | 94 |
| P00597 | 179.51 | 63 | 8 | 16,016 | Acidic phospholipase A2 2 | Naja kaouthia | 94 |
| Q91133 | 179.00 | 54 | 7 | 15,949 | Acidic phospholipase A2 2 | Naja atra | 92.3 |
| P15445 | 172.39 | 65 | 7 | 13,346 | Acidic phospholipase A2 2 | Naja naja | 92.3 |
| A4FS04 | 161.13 | 77 | 7 | 13,188 | Acidic phospholipase A2 natratoxin | Naja atra | 92.3 |
| Q6T179 | 149.66 | 51 | 6 | 14,198 | Acidic phospholipase A2 4 (fragment) | Naja sagittifera | 87.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdullah, N.A.H.; Rusmili, M.R.A.; Zainal Abidin, S.A.; Shaikh, M.F.; Hodgson, W.C.; Othman, I. Isolation and Characterization of A2-EPTX-Nsm1a, a Secretory Phospholipase A2 from Malaysian Spitting Cobra (Naja sumatrana) Venom. Toxins 2021, 13, 859. https://doi.org/10.3390/toxins13120859
Abdullah NAH, Rusmili MRA, Zainal Abidin SA, Shaikh MF, Hodgson WC, Othman I. Isolation and Characterization of A2-EPTX-Nsm1a, a Secretory Phospholipase A2 from Malaysian Spitting Cobra (Naja sumatrana) Venom. Toxins. 2021; 13(12):859. https://doi.org/10.3390/toxins13120859
Chicago/Turabian StyleAbdullah, Nur Atiqah Haizum, Muhamad Rusdi Ahmad Rusmili, Syafiq Asnawi Zainal Abidin, Mohd Farooq Shaikh, Wayne C. Hodgson, and Iekhsan Othman. 2021. "Isolation and Characterization of A2-EPTX-Nsm1a, a Secretory Phospholipase A2 from Malaysian Spitting Cobra (Naja sumatrana) Venom" Toxins 13, no. 12: 859. https://doi.org/10.3390/toxins13120859
APA StyleAbdullah, N. A. H., Rusmili, M. R. A., Zainal Abidin, S. A., Shaikh, M. F., Hodgson, W. C., & Othman, I. (2021). Isolation and Characterization of A2-EPTX-Nsm1a, a Secretory Phospholipase A2 from Malaysian Spitting Cobra (Naja sumatrana) Venom. Toxins, 13(12), 859. https://doi.org/10.3390/toxins13120859