Functional Role of Aspergillus carbonarius AcOTAbZIP Gene, a bZIP Transcription Factor within the OTA Gene Cluster

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
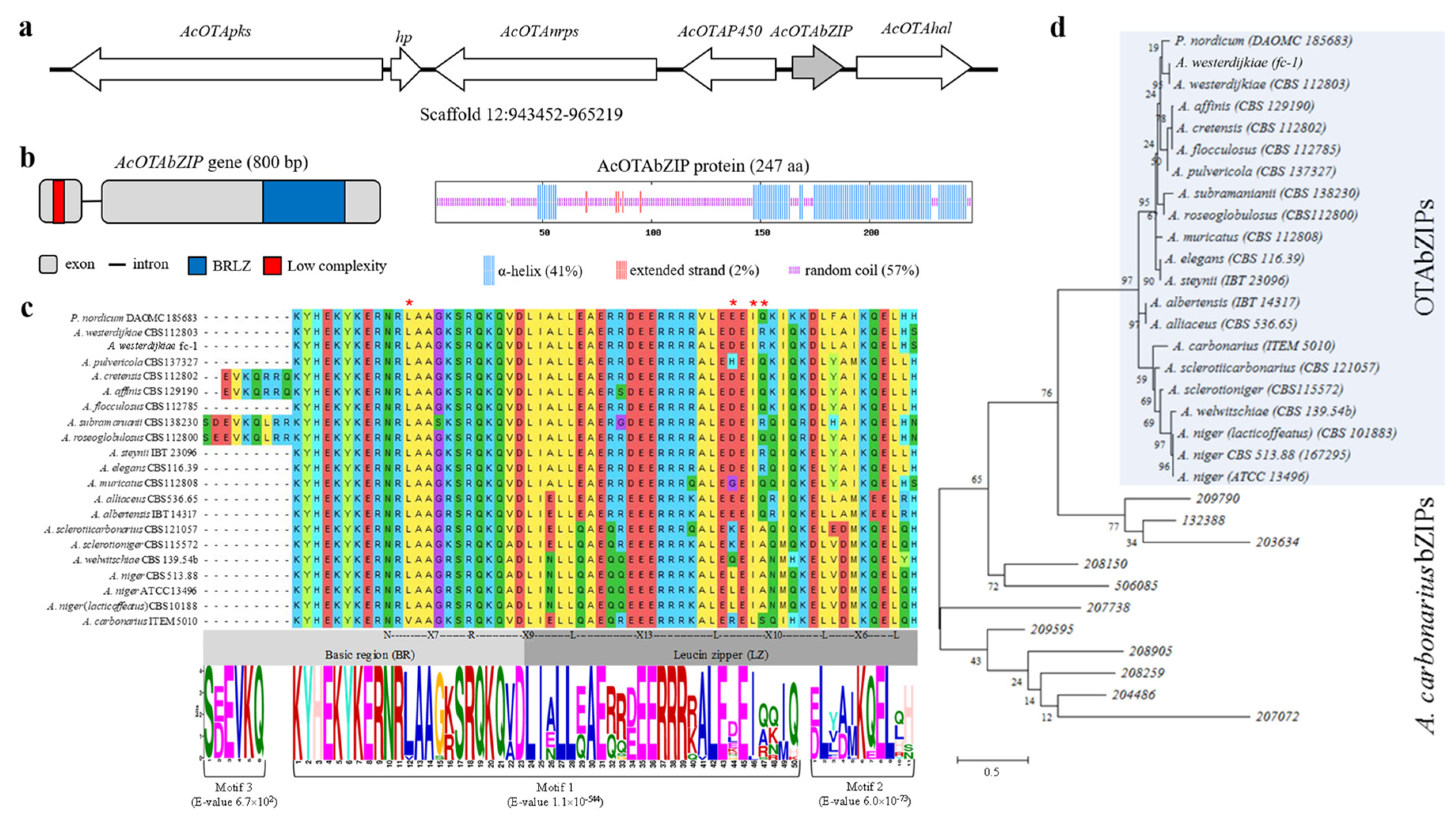
2.1. Characterization of AcOTAbZIP Gene
2.2. Generation of A. carbonarius Deletion Mutants
2.3. Phenotypic Characterization
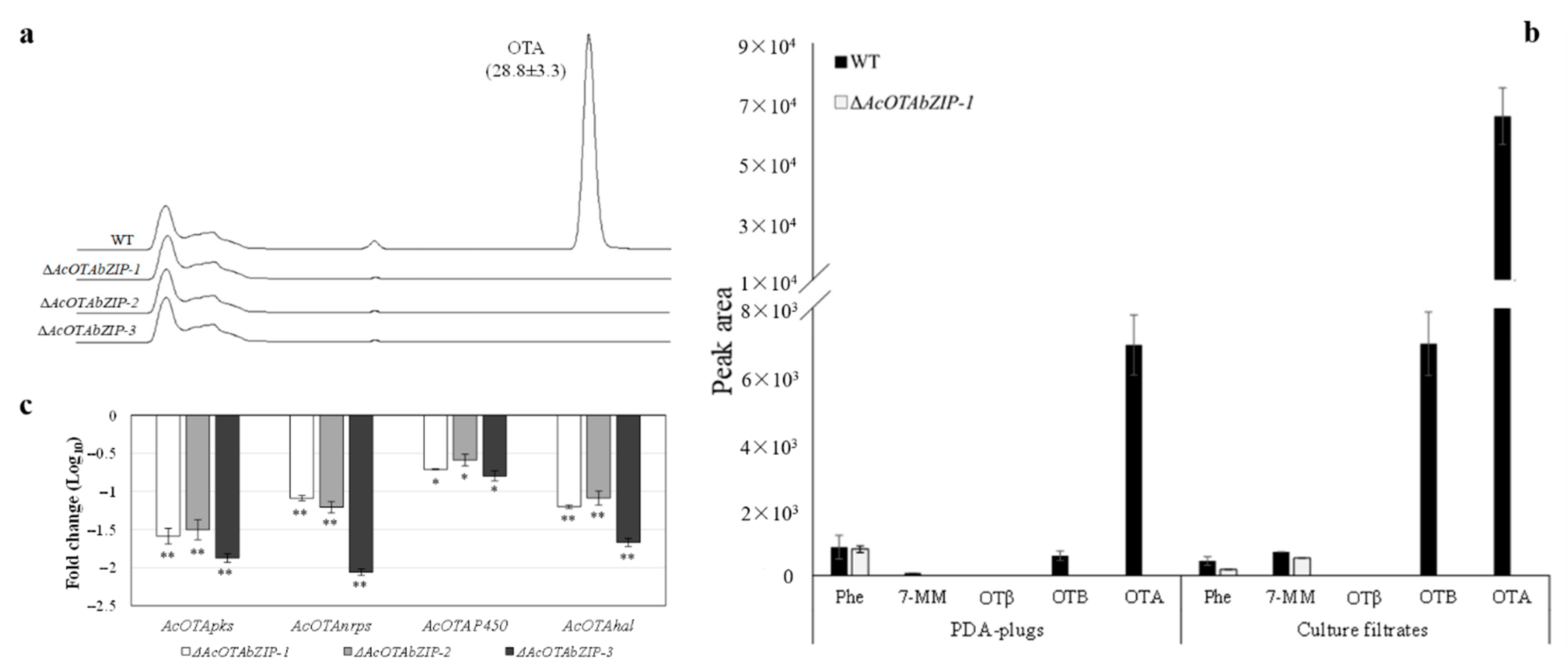
2.4. Chemical Analysis of OTA and Its Intermediates and Gene Expression Studies
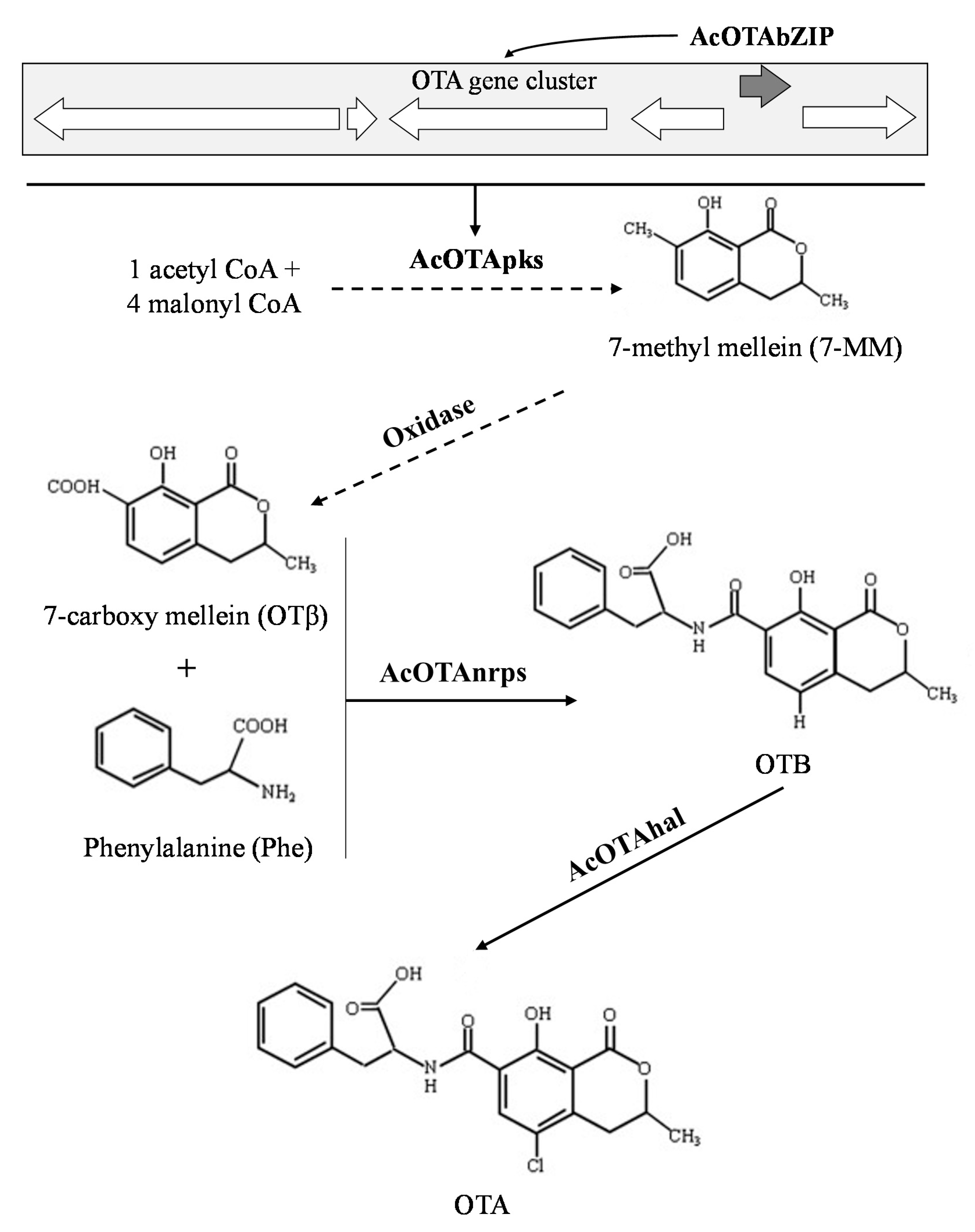
3. Discussion
4. Material and Methods
4.1. Strains and Media
4.2. Identification and Characterization of AcOTAbZIP Gene
4.3. Deletion of AcOTAbZIP Gene in A. carbonarius
4.4. Phenotypic Characterization and In Vivo Assay
4.5. Analysis of OTA and Its Intermediates
4.6. Gene Expression Studies by RT-qPCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Agency for Research on Cancer (IARC). IARC Monographs on the Evaluation of the Carcinogenic Risks to Humans; IARC: Lyon, France, 1993; Volume 56, pp. 489–521. [Google Scholar]
- Abarca, M.L.; Accensi, F.; Bragulat, M.R.; Castella, G.; Cabanes, F.J. Aspergillus carbonarius is the main source of ochratoxin A contamination in dried vine fruits from the Spanish market. J. Food Protect. 2003, 66, 504–506. [Google Scholar] [CrossRef] [PubMed]
- Amezqueta, S.; Schorr-Galindo, S.; Murillo-Arbizu, M.; Gonzalez-Peñas, E.; De Cerain, A.L.; Guiraud, J.P. OTA-producing fungi in foodstuffs: A review. Food Control 2012, 26, 259–268. [Google Scholar] [CrossRef]
- Cabañes, F.J.; Bragulat, M.R.; Castellá, G. Ochratoxin A producing species in the genus Penicillium. Toxins 2010, 2, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Patiño, B.; Cortes, L.; Gonzalez-Jaen, M.T.; Vazquez, C. Aspergillus steynii and Aspergillus westerdijkiae as potential risk of OTA contamination in food products in warm climates. Food Microbiol. 2015, 46, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Gallo, A. Aspergillus Species and Their Associated Mycotoxins; Mycotoxigenic Fungi Humana Press: New York, NY, USA, 2017; pp. 33–49. [Google Scholar]
- Perrone, G.; Susca, A.; Cozzi, G.; Ehrlich, K.; Varga, J.; Frisvad, J.C.; Meijer, M.; Noonim, P.; Mahakarnchanakul, W.; Samson, R.A. Biodiversity of Aspergillus species in some important agricultural products. Stud. Mycol. 2007, 59, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Cabañes, F.J.; Accensi, F.; Bragulat, M.R.; Abarca, M.L.; Castellá, G.; Minguez, S.; Pons, A. What is the source of ochratoxin A in wine? Int. J. Food Microbiol. 2002, 79, 213–215. [Google Scholar] [CrossRef]
- Pollastro, S.; Dongiovanni, C.; Abbatecola, A.; Tauro, G.; Natale, P.; Pascale, M.; Visconti, A.; Faretra, F. Wine contamination by ochratoxin A in South Italy: Causes and preventive actions. J. Plant Pathol. 2003, 85, 281. [Google Scholar]
- Gallo, A.; Bruno, K.S.; Solfrizzo, M.; Perrone, G.; Mulè, G.; Visconti, A.; Baker, S.E. New insight into the ochratoxin A biosynthetic pathway through deletion of a nonribosomal peptide synthetase gene in Aspergillus carbonarius. Appl. Environ. Microb. 2012, 78, 8208–8218. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Knox, B.P.; Bruno, K.S.; Solfrizzo, M.; Baker, S.E.; Perrone, G. Identification and characterization of the polyketide synthase involved in ochratoxin A biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2014, 179, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.; Perrone, G.; Gambacorta, L.; Epifani, F.; Solfrizzo, M.; Gallo, A. Identification of a halogenase involved in the biosynthesis of ochratoxin A in Aspergillus carbonarius. Appl. Environ. Microb. 2016, 82, 5631–5641. [Google Scholar] [CrossRef]
- Ferrara, M.; Gallo, A.; Perrone, G.; Magistà, D.; Baker, S.E. Comparative genomic analysis of ochratoxin A biosynthetic cluster in producing fungi: New evidence of a cyclase gene involvement. Front. Microbiol. 2020, 11, 3289. [Google Scholar]
- Gerin, D.; De Miccolis Angelini, R.M.; Pollastro, S.; Faretra, F. RNA-Seq reveals OTA-related gene transcriptional changes in Aspergillus carbonarius. PLoS ONE 2016, 11, e0147089. [Google Scholar] [CrossRef] [PubMed]
- Castellá, G.; Bragulat, M.R.; Puig, L.; Sanseverino, W.; Cabañes, F.J. Genomic diversity in ochratoxigenic and non ochratoxigenic strains of Aspergillus carbonarius. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Castellá, G.; Bragulat, M.R.; Cigliano, R.A.; Cabañes, F.J. Transcriptome analysis of non-ochratoxigenic Aspergillus carbonarius strains and interactions between some black aspergilli species. Int. J. Food Microbiol. 2020, 317, 108498. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Vázquez, C.; Patiño, B. The Genomic Regions That Contain Ochratoxin A Biosynthetic Genes Widely Differ in Aspergillus Section Circumdati Species. Toxins 2020, 12, 754. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.-B.; et al. A consensus ochratoxin A biosynthetic pathway: Insights from the genome sequence of Aspergillus ochraceus and a comparative genomic analysis. Appl. Environ. Microb. 2018, 84, e01009–e01018. [Google Scholar] [CrossRef] [PubMed]
- Marzluf, G.A. Genetic regulation of nitrogen metabolism in the fungi. Microbiol. Mol. Biol. Rev. 1997, 61, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Mihlan, M.; Homann, V.; Liu, T.W.D.; Tudzynski, B. AREA directly mediates nitrogen regulation of gibberellin biosynthesis in Gibberella fujikuroi, but its activity is not affected by NMR. Mol. Microbiol. 2003, 47, 975–991. [Google Scholar] [CrossRef]
- Tilburn, J.; Sarkar, S.; Widdick, D.A.; Espeso, E.A.; Orejas, M.; Mungroo, J.; Peñalva, M.A.; Arst, H.N., Jr. The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid-and alkaline-expressed genes by ambient pH. EMBO J. 1995, 14, 779–790. [Google Scholar] [CrossRef]
- Dowzer, C.E.; Kelly, J.M. Analysis of the creA gene, a regulator of carbon catabolite repression in Aspergillus nidulans. Mol. Cell. Biol. 1991, 11, 5701–5709. [Google Scholar] [CrossRef]
- Espeso, E.A.; Peñalva, M.A. Carbon catabolite repression can account for the temporal pattern of expression of a penicillin biosynthetic gene in Aspergillus nidulans. Mol. Microbiol. 1992, 6, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Sempere, A.; Marin, S.; Sanchis, V.; Ramos, A.J. VeA and LaeA transcriptional factors regulate ochratoxin A biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2013, 166, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Montalbano, B.G.; Cary, J.W. Binding of the C6-zinc cluster protein, AFLR, to the promoters of aflatoxin pathway biosynthesis genes in Aspergillus parasiticus. Gene 1999, 230, 249–257. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, C.; Wu, C.; Sun, P.; Hou, R.; Liu, H.; Wang, C.; Xu, J.R. TRI6 and TRI10 play different roles in the regulation of deoxynivalenol (DON) production by cAMP signalling in Fusarium graminearum. Environ. Microbiol. 2016, 18, 3689–3701. [Google Scholar] [CrossRef] [PubMed]
- Landschulz, W.H.; Johnson, P.F.; McKnight, S.L. The leucine zipper: A hypothetical structure common to a new class of DNA binding proteins. Science 1988, 240, 1759–1764. [Google Scholar] [CrossRef]
- Vinson, C.; Acharya, A.; Taparowsky, E.J. Deciphering B-ZIP transcription factor interactions in vitro and in vivo. BBA-Gene Struct. Expr. 2006, 1759, 4–12. [Google Scholar] [CrossRef]
- Gil-Serna, J.; García-Díaz, M.; González-Jaén, M.T.; Vázquez, C.; Patiño, B. Description of an orthologous cluster of ochratoxin A biosynthetic genes in Aspergillus and Penicillium species. A comparative analysis. Int. J. Food Microbiol. 2018, 268, 35–43. [Google Scholar] [CrossRef]
- Huffman, J.; Gerber, R.; Du, L. Recent advancements in the biosynthetic mechanisms for polyketide derived mycotoxins. Biopolymers 2010, 93, 764–776. [Google Scholar] [CrossRef]
- Huff, W.E.; Hamilton, P.B. Mycotoxins–their biosynthesis in fungi: Ochratoxins–metabolites of combined pathways. J. Food Protect. 1979, 42, 815–820. [Google Scholar] [CrossRef]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef]
- Gerin, D.; González-Candelas, L.; Ballester, A.R.; Pollastro, S.; De Miccolis Angelini, R.M.; Faretra, F. Functional Characterization of the alb1 Orthologue Gene in the Ochratoxigenic Fungus Aspergillus carbonarius (AC49 strain). Toxins 2018, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Barda, O.; Maor, U.; Sadhasivam, S.; Bi, Y.; Zakin, V.; Prusky, D.; Sionov, E. The pH-Responsive Transcription Factor PacC Governs Pathogenicity and Ochratoxin A Biosynthesis in Aspergillus carbonarius. Front. Microbiol. 2020, 11, 210. [Google Scholar] [CrossRef] [PubMed]
- Hof, H.; Kupfahl, C. Gliotoxin in Aspergillus fumigatus: An example that mycotoxins are potential virulence factors. Mycotoxin Res. 2009, 25, 123. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.J.; Desjardins, A.E.; Plattner, R.D.; Nicholson, P.; Butler, G.; Young, J.C.; Weston, G.; Proctor, R.H.; Hohn, T.M. Possible role of trichothecene mycotoxins in virulence of Fusarium graminearum on maize. Plant Dis. 1999, 83, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef]
- Sanderson, K.E.; Srb, A.M. Heterokaryosis and parasexuality in the fungus Ascochyta imperfecta. Am. J. Bot. 1965, 52, 72–81. [Google Scholar] [CrossRef]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deleage, G. NPS@: Network protein sequence analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef]
- Ponting, C.P.; Schultz, J.; Milpetz, F.; Bork, P. SMART: Identification and annotation of domains from signalling and extracellular protein sequences. Nucleic Acids Res. 1999, 27, 229–232. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Lu, Y.; Zinta, G.; Lang, Z.; Zhu, J.K. UTR-dependent control of gene expression in plants. Trends Plant Sci. 2018, 23, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, R.J.; Andersson, J.A.; Kristensen, M.B.; Giese, H. Efficient four fragment cloning for the construction of vectors for targeted gene replacement in filamentous fungi. BMC Mol. Biol. 2008, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | In Vitro Assay | Assay on Grape Berries | ||||||
|---|---|---|---|---|---|---|---|---|
| Growth Rate (mm day−1) | Conidia [(No. × 104)/mm2] | Growth Rate (mm/day) | ||||||
| MM | PDA | MEA | MM | PDA | MEA | Italia | Red Globe | |
| WT | 4.2 ± 0.3 a | 7.6 ± 0.7 a | 6.4 ± 0.3 a | 8.2 ± 0.2 a | 0.4 ± 0.4 a | 0.5 ± 0.1 a | 3.0 ± 0.1 a | 2.3 ± 0.1 a |
| ΔAcOTAbZIP-1 | 4.3 ± 0.3 a | 7.5 ± 0.7 a | 6.4 ± 0.3 a | 10.6 ± 1.0 a | 0.4 ± 0.3 a | 0.5 ± 0.1 a | 2.9 ± 0.1 a | 2.2 ± 0.1 a |
| ΔAcOTAbZIP-2 | 4.3 ± 0.3 a | 7.5 ± 0.7 a | 6.4 ± 0.4 a | 7.5 ± 1.1 a | 0.4 ± 0.1 a | 0.5 ± 0.2 a | 3.0 ± 0.1 a | 2.3 ± 0.1 a |
| ΔAcOTAbZIP-3 | 4.3 ± 0.3 a | 7.5 ± 0.7 a | 6.4 ± 0.4 a | 7.5 ± 1.1 a | 0.4 ± 0.2 a | 0.5 ± 0.1 a | 3.0 ± 0.2 a | 2.2 ± 0.1 a |
| Target Region | Primer Name | Primer Sequence (5′-3′) |
|---|---|---|
| Promoter and terminator amplification in A. carbonarius (AC49) | ||
| AcOTAbZIP promoter | AcOTAbZIP_O1 | GGTCTTAAUTGTTGAAGGTGCGGTTCTTG |
| AcOTAbZIP_O2 | GGCATTAAUCATGAGCATTGACACGAGCC | |
| AcOTAbZIP terminator | AcOTAbZIP_A3 | GGACTTAAUTGAGCGCATGTCTAGCAAAC |
| AcOTAbZIP_A4 | GGGTTTAAUTCGGCCGTGAAGCAGTTATA | |
| Screening in E. coli (DH5α) | ||
| pRFHU2-AcOTAbZIP plasmid | RF-2 | TCTCCTTGCATGCACCATTCCTTG |
| RF-5 | GTTTGCAGGGCCATAGAC | |
| RF-1 | AAATTTTGTGCTCACCGCCTGGAC | |
| RF-6 | ACGCCAGGGTTTTCCCAGTC | |
| Screening in A. carbonarius (AC49 and ΔAcOTAbZIP strains) | ||
| AcOTAbZIP promoter | AcOTAbZIP_1F | AGGCGTTATAGGACCAGTCG |
| HPH1F | ACGAGGTCGCCAACATCTTCTTCT | |
| AcOTAbZIP terminator | AcOTAbZIP_2R | CACTCGCTCCTCCGTGATAT |
| HPHPRO4 | GCACCAAGCAGCAGATGATA | |
| Hygromycin B | HMBF1 | CTGTCGAGAAGTTTCTGATCG |
| HMBR1 | CTGATAGAGTTGGTCAAGACC | |
| AcOTAbZIP | AcOTAbZIP_3F | CATCCATGCCCCAATTCGAG |
| AcOTAbZIP_4R | TGCTTGAGGTCTAAGAGTTCCT | |
| T-DNA copy numbers integrated into A. carbonarius ΔAcOTAbZIP strains genome | ||
| AcOTAbZIP | AcOTAbZIP_CN_F | AATTGACAGCGAGGCGAATC |
| AcOTAbZIP_CN_R | CCTGCAGCAACTCGATCAAA | |
| Calmodulin | Cal_CN_F | CCTTACCATGATGGCTCG |
| Cal_CN_R | TTCTCACCGATGGAGGTCAT | |
| RT-PCR and RTqPCR (AC49 and ΔAcOTAbZIP strains) | ||
| AcOTAbZIP | bZIPFor | TTTCCCTAGGATCTCTCCTA |
| bZIPRev | TATTGGGGTCGGACAGGAAT | |
| AcOTApks | pks4For | TCTGTATGAGCGCATCGCC |
| pks4Rev | GCAGAAGGCCACTTTCCAG | |
| AcOTAnrps | nrps6For | GATTCCGATGGAACTGCAAT |
| nrps6Rev | CTGCCCCAGCATATCAATCT | |
| AcOTAP450 | P450For | GCCATACCTGACCGGGATCA |
| P450Rev | GGGAAAATGGTCTCGTCGTG | |
| AcOTAhal | halFor | AAAGAAGCCTACACCGACTT |
| halRev | GAATTCGATGGATCCCGTGC | |
| Ubiquitin | ubFor | CCGAAGGTCAACTTCACCAC |
| ubRev | GGCATATTTGCGAGTCCATT | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerin, D.; Garrapa, F.; Ballester, A.-R.; González-Candelas, L.; De Miccolis Angelini, R.M.; Faretra, F.; Pollastro, S. Functional Role of Aspergillus carbonarius AcOTAbZIP Gene, a bZIP Transcription Factor within the OTA Gene Cluster. Toxins 2021, 13, 111. https://doi.org/10.3390/toxins13020111
Gerin D, Garrapa F, Ballester A-R, González-Candelas L, De Miccolis Angelini RM, Faretra F, Pollastro S. Functional Role of Aspergillus carbonarius AcOTAbZIP Gene, a bZIP Transcription Factor within the OTA Gene Cluster. Toxins. 2021; 13(2):111. https://doi.org/10.3390/toxins13020111
Chicago/Turabian StyleGerin, Donato, Federica Garrapa, Ana-Rosa Ballester, Luis González-Candelas, Rita Milvia De Miccolis Angelini, Francesco Faretra, and Stefania Pollastro. 2021. "Functional Role of Aspergillus carbonarius AcOTAbZIP Gene, a bZIP Transcription Factor within the OTA Gene Cluster" Toxins 13, no. 2: 111. https://doi.org/10.3390/toxins13020111
APA StyleGerin, D., Garrapa, F., Ballester, A. -R., González-Candelas, L., De Miccolis Angelini, R. M., Faretra, F., & Pollastro, S. (2021). Functional Role of Aspergillus carbonarius AcOTAbZIP Gene, a bZIP Transcription Factor within the OTA Gene Cluster. Toxins, 13(2), 111. https://doi.org/10.3390/toxins13020111