Extensive Variation in the Activities of Pseudocerastes and Eristicophis Viper Venoms Suggests Divergent Envenoming Strategies Are Used for Prey Capture

 , , ,
, , ,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results and Discussion
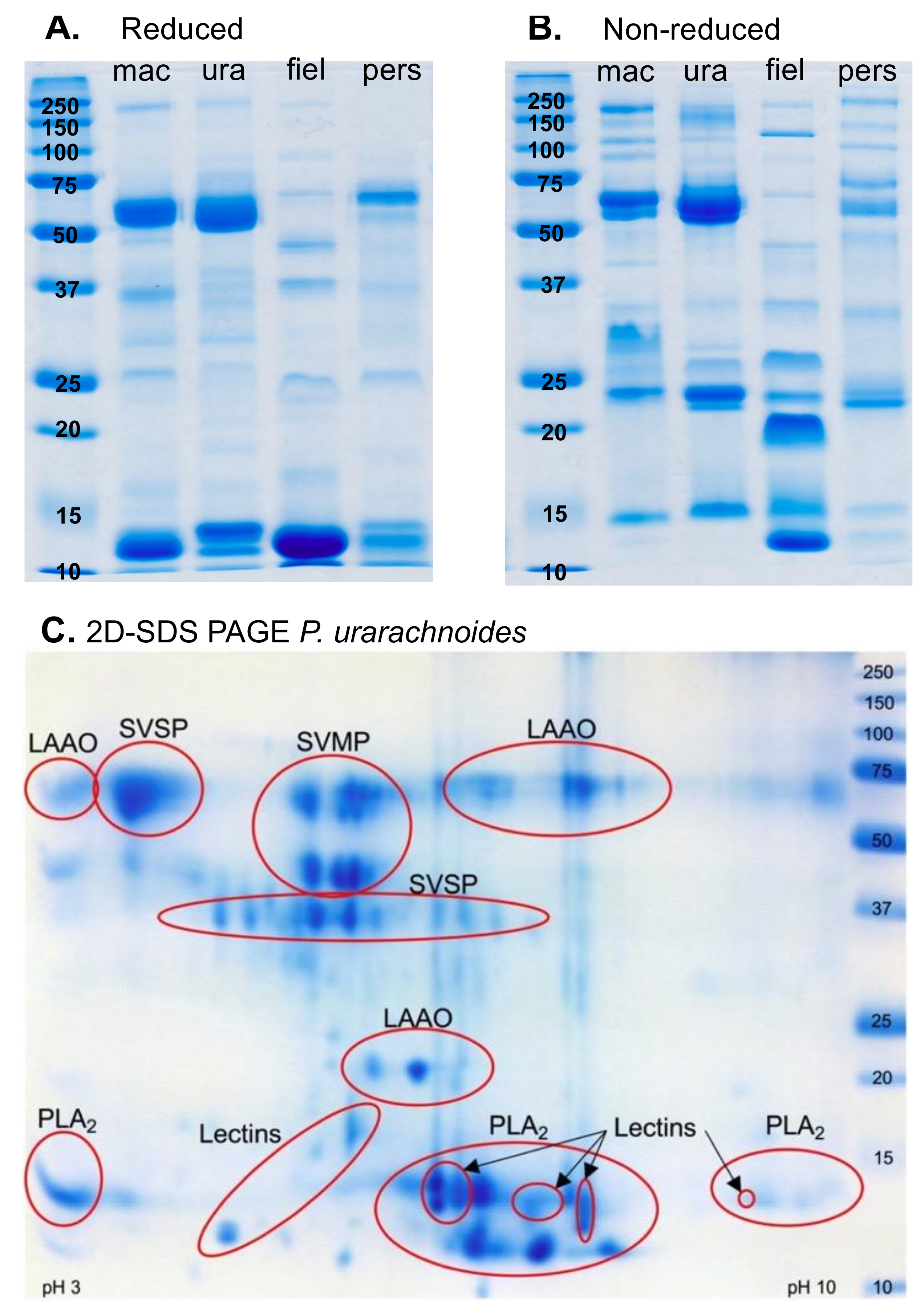
2.1. Venom Proteomics
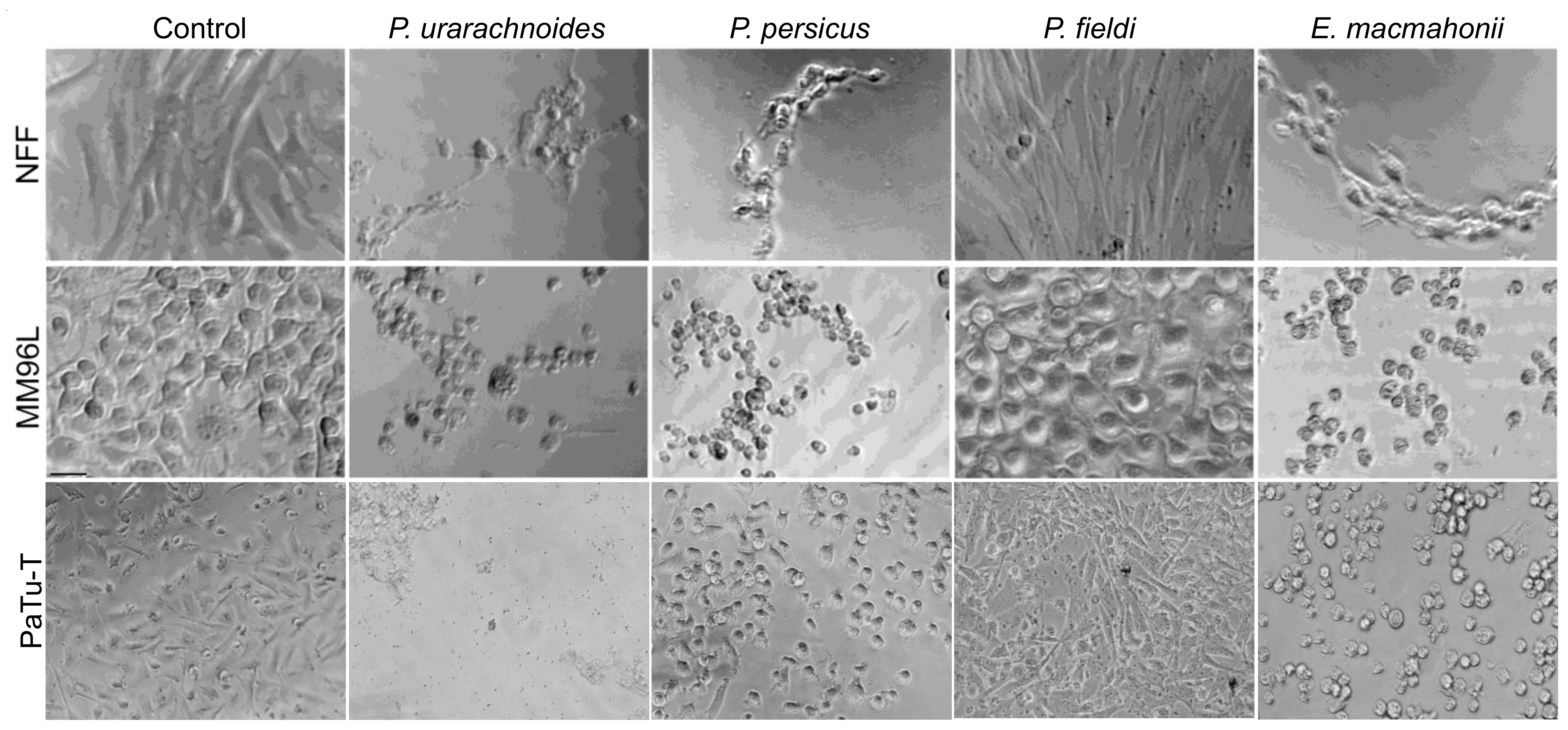
2.2. Venom-Induced Cytotoxicity
2.3. Venom-Induced Neurotoxicity
2.4. Venom-Induced Coagulant Activity
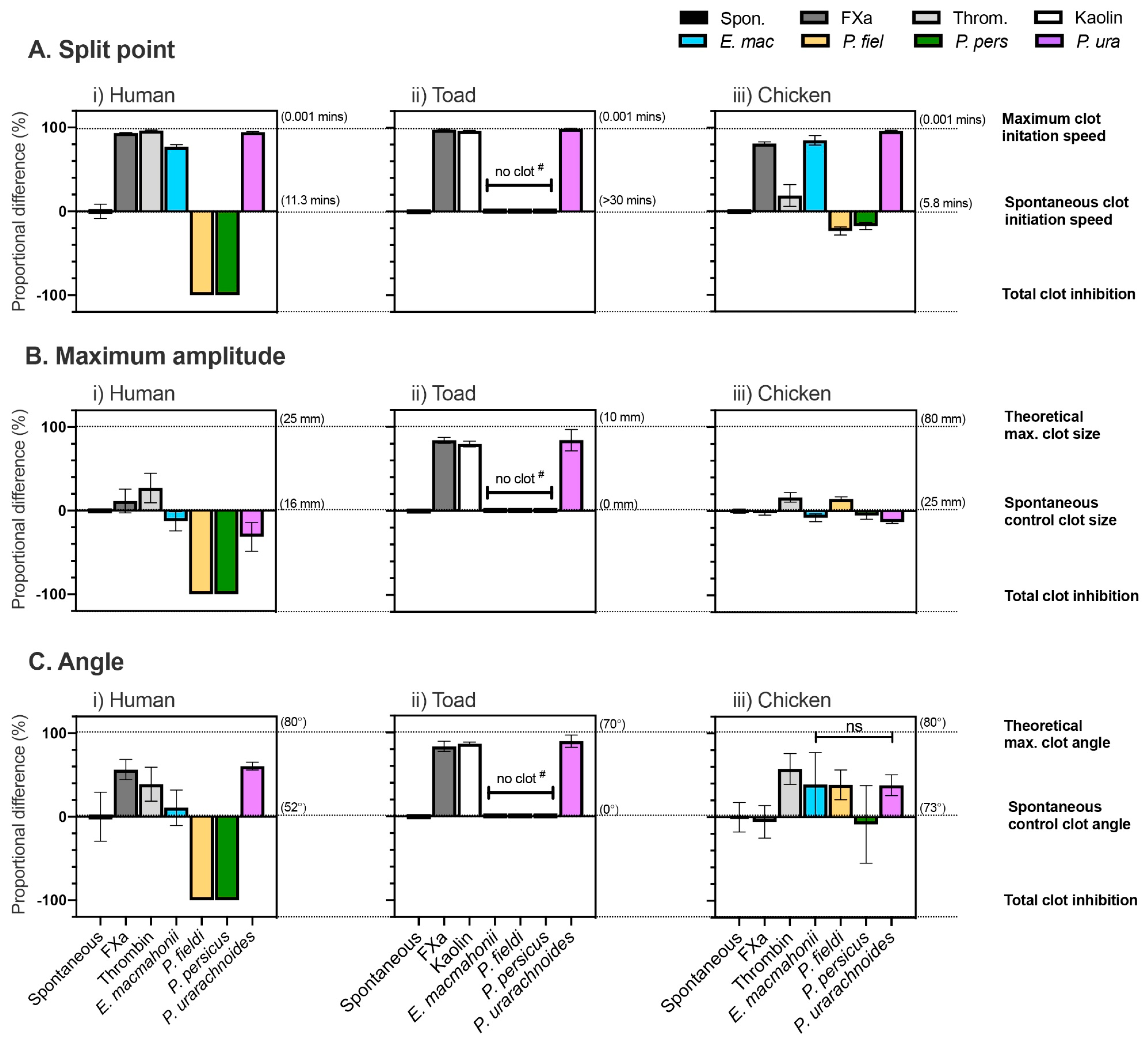
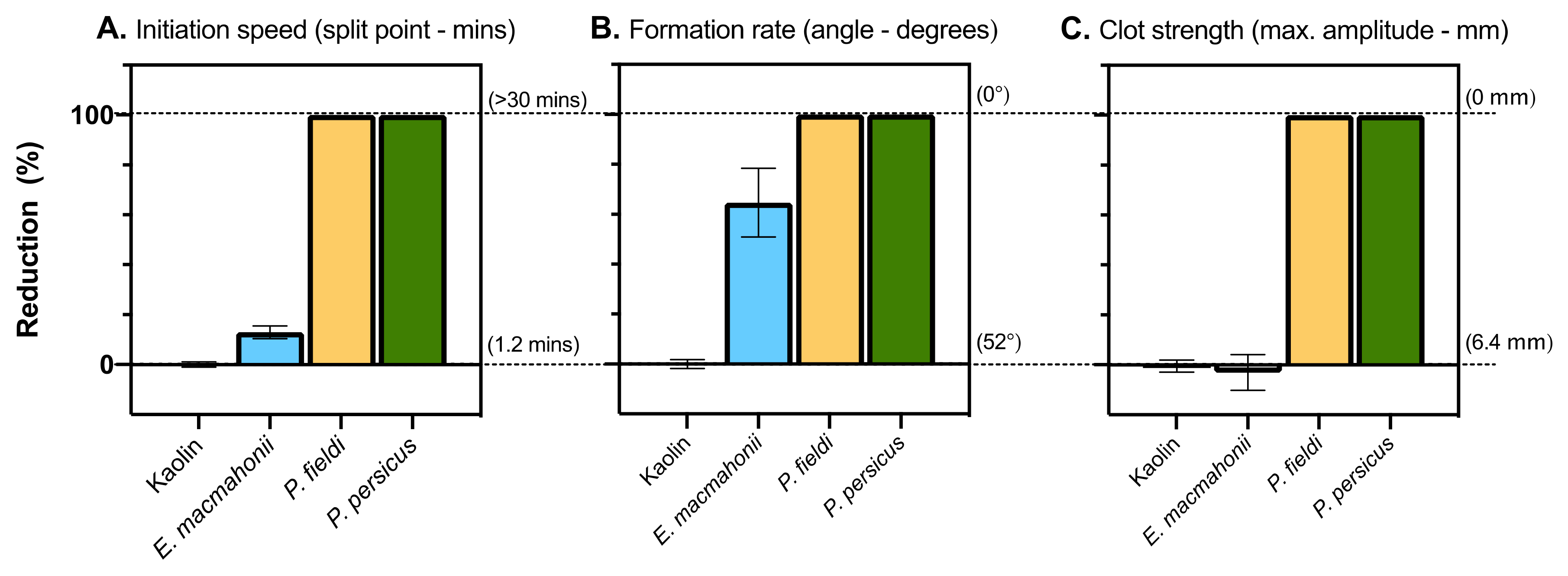
2.4.1. Coagulation Time, Strength, and Potency
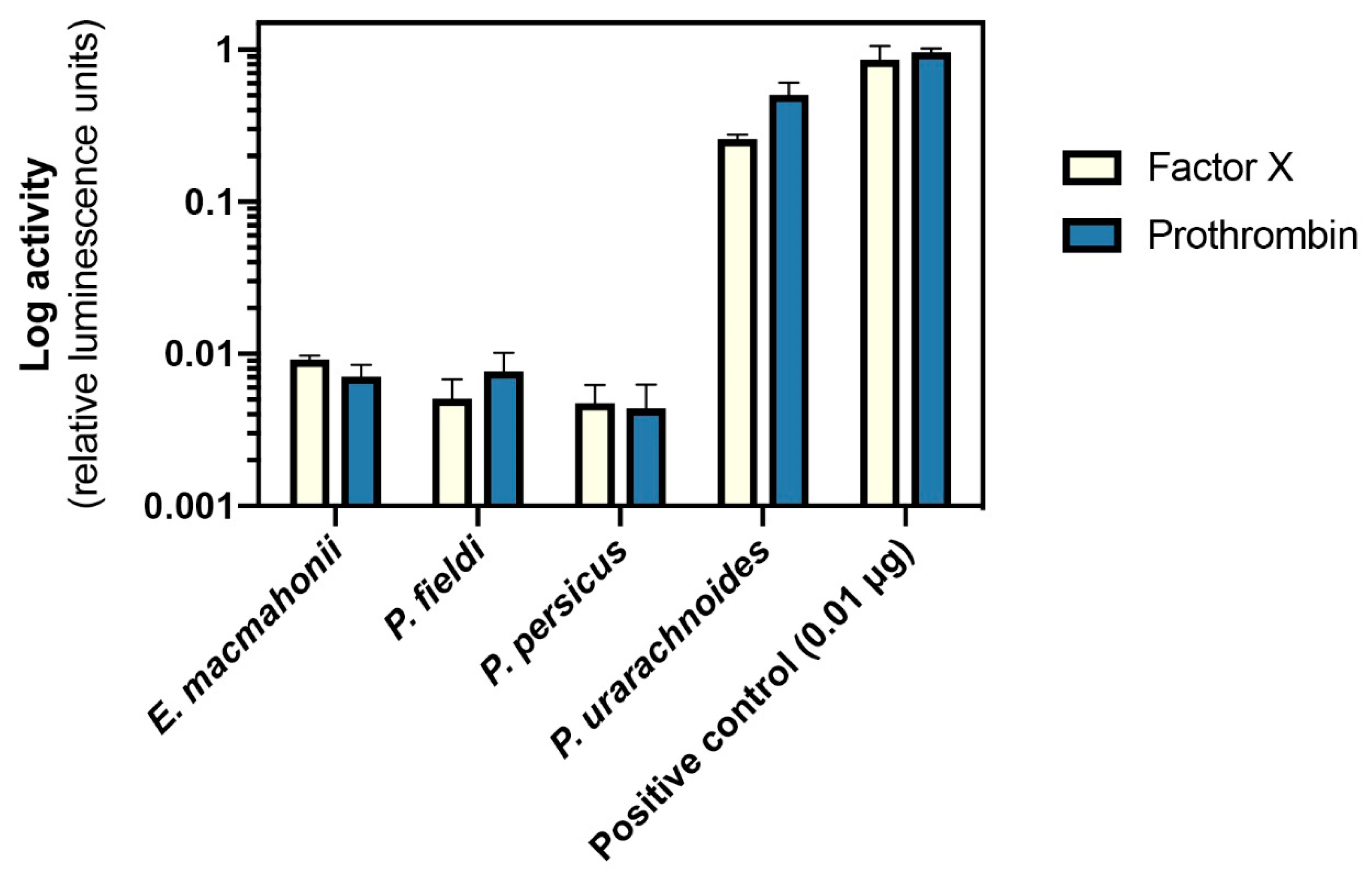
2.4.2. Activation of Factor X and Prothrombin
2.4.3. Thromboelastography
3. Conclusions
4. Materials and Methods
4.1. Venom Samples
4.2. Plasma
4.3. Proteomics
4.3.1. Venom SDS-PAGE Gel Electrophoresis
4.3.2. Liquid Chromatography-Mass Spectrometry
4.4. Cytotoxicity
4.5. Neurotoxicity
4.6. Coagulation Analyses
4.6.1. Clot Formation Time
4.6.2. Factor X and Prothrombin Activation
4.6.3. Thromboelastography
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [Green Version]
- Phelps, T. Old World Vipers, A Natural History of the Azemiopinae and Viperinae; Edition Chimaira: Frankfurt am Main, Germany, 2010. [Google Scholar]
- Bostanchi, H.; Anderson, S.C.; Kami, H.G.; Papenfuss, T.J. A new species of Pseudocerastes with elaborate tail ornamentation from western Iran (Squamata: Viperidae). Proc. Cal. Acad. Sci. 2006, 57, 443. [Google Scholar]
- Fathinia, B.; Rastegar-Pouyani, N.; Rastegar-Pouyani, E. Molecular phylogeny and historical biogeography of genera Eristicophis and Pseudocerastes (Ophidia, Viperidae). Zool. Scr. 2018, 47, 673–685. [Google Scholar] [CrossRef]
- Wüster, W.; Peppin, L.; Pook, C.E.; Walker, D.E. A nesting of vipers: Phylogeny and historical biogeography of the Viperidae (Squamata: Serpentes). Mol. Phylogenet. Evol. 2008, 49, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Alencar, L.R.; Quental, T.B.; Grazziotin, F.G.; Alfaro, M.L.; Martins, M.; Venzon, M.; Zaher, H. Diversification in vipers: Phylogenetic relationships, time of divergence and shifts in speciation rates. Mol. Phylogenet. Evol. 2016, 105, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Smid, J.; Tolley, K.A. Calibrating the tree of vipers under the fossilized birth-death model. Sci. Rep. 2019, 9, 5510. [Google Scholar] [CrossRef] [PubMed]
- Mallow, D.; Ludwig, D.; Nilson, G. True Vipers: Natural History and Toxinology of Old World Vipers; Kreiger Publishing Company: Malabar, FL, USA, 2003. [Google Scholar]
- Del Marmol, G.M.; Mozaffari, O.; Gállego, J. Pseudocerastes urarachnoides: The ambush specialist. Waterbirds 2016, 10, 117–126. [Google Scholar]
- Fathinia, B.; Anderson, S.C.; Rastegar-Pouyani, N.; Jahani, H.; Mohamadi, H. Notes on the natural history of Pseudocerastes urarachnoides (Squamata: Viperidae). Rus. J. Herpetol. 2009, 16, 134–138. [Google Scholar]
- Al-Sheikhly, O.F.; Al-Barazengy, A.N.; LAl-Haideri, M. First record of the Iranian Spider Viper Pseudocerastes urarachnoides Bostanchi, Anderson, Kami & Papenfuss, 2006 (Serpentes: Viperidae) in Iraq. SAURIA Berl. 2019, 41, 43–46. [Google Scholar]
- De Pous, P.; Simó-Riudalbas, M.; Els, J.; Jayasinghe, S.; Amat, F.; Carranza, S. Phylogeny and biogeography of Arabian populations of the Persian horned viper Pseudocerastes persicus (Duméril, Bibron & Duméril, 1854). Zool. Mid. East 2016, 62, 231–238. [Google Scholar]
- Bok, B.; Berroneau, M.; Yousefi, M.; Nerz, J.; Deschandol, F.; Berroneau, M.; Tiemann, L. Sympatry of Pseudocerastes persicus and P. urarachnoides in the western Zagros Mountains, Iran. Herpetol. Notes 2017, 10, 323–325. [Google Scholar]
- Gholamifard, A.; Esmaeili, H.R. First record and range extension of Field’s horned viper, Pseudocerastes fieldi Schmidt, 1930 (Squamata: Viperidae), from Fars province, southern Iran. Turk. J. Zool. 2010, 34, 551–552. [Google Scholar]
- Fathinia, B.; Rastegar-Pouyani, N. On the species of Pseudocerastes (Ophidia: Viperidae) in Iran. Russ. J. Herpetol. 2010, 17, 275–279. [Google Scholar]
- Baig, K.J.; Awan, M.R.; Ashraf, N. Ecological studies and zoogeographic affinities of the amphibians and reptiles found in Chagai desert, Balochistan, Pakistan. Pak. J. Zool. 2006, 38, 145. [Google Scholar]
- Khan, M.S. A Guide to the Snakes of Pakistan; Edition Chimaira: Frankfurt am Main, Germany, 2002. [Google Scholar]
- Minton, S.A. A Contribution to the Herpetology of West Pakistan; American Museum of Natural History: New York, NY, USA, 1966. [Google Scholar]
- Mendelssohn, H. On the biology of the venomous snakes of Israel. II. Isr. J. Ecol. Evol. 1965, 14, 185–212. [Google Scholar]
- Amr, Z.S.; Disi, A.M. Systematics, distribution and ecology of the snakes of Jordan. Vertebr. Zool. 2011, 61, 179–266. [Google Scholar]
- Ali, S.A.; Jackson, T.N.; Casewell, N.R.; Low, D.H.; Rossi, S.; Baumann, K.; Fathinia, B.; Visser, J.; Nouwens, A.; Hendrikx, I.; et al. Extreme venom variation in Middle Eastern vipers: A proteomics comparison of Eristicophis macmahonii, Pseudocerastes fieldi and Pseudocerastes persicus. J. Proteom. 2015, 116, 106–113. [Google Scholar] [CrossRef]
- Carranza, S.; Xipell, M.; Tarroso, P.; Gardner, A.; Arnold, E.N.; Robinson, M.D.; Simó-Riudalbas, M.; Vasconcelos, R.; de Pous, P.; Amat, F.; et al. Diversity, distribution and conservation of the terrestrial reptiles of Oman (Sauropsida, Squamata). PLoS ONE 2018, 13, e0190389. [Google Scholar] [CrossRef] [Green Version]
- Thomas, O. Persian horned viper (Pseudocerastes persicus): A record at unusually low elevation from the United Arab Emirates. Herpetol. Bull. 2019, 147, 28–29. [Google Scholar] [CrossRef]
- Fathinia, B.; Rastegar-Pouyani, N.; Rastegar-Pouyani, E.; Todehdehghan, F.; Mansouri, M. Annual activity pattern of Pseudocerastes urarachnoides Bostanchi, Anderson, Kami & Papenfuss, 2006, with notes on its natural history (squamata: Serpentes: Viperidae). Herpetozoa 2017, 29, 135–142. [Google Scholar]
- Fathinia, B.; Rastegar-pouyani, N.; Rastegar-pouyani, E. Avian deception using an elaborate caudal lure in Pseudocerastes urarachnoides (Serpentes:Viperidae). Amphib. Reptil. 2015, 36, 223–231. [Google Scholar] [CrossRef]
- Alam, J.M.; Ali, S.A. Age-dependent variability of biochemical and biological properties in venoms of Leaf-Nose Viper snakes, Eristicophis macmahoni. Pak. J. Zool. 2001, 33, 173–177. [Google Scholar]
- Bdolah, A. Comparison of venoms from two subspecies of the false horned viper (Pseudocerastes persicus). Toxicon 1986, 24, 726–729. [Google Scholar] [CrossRef]
- Batzri-Izraeli, R.; Bdolah, A. Isolation and characterization of the main toxic fraction from the venom of the false horned viper (Pseudocerastes fieldi). Toxicon 1982, 20, 867–875. [Google Scholar] [CrossRef]
- Francis, B.; Bdolah, A.; Kaiser, I.I. Amino acid sequences of a heterodimeric neurotoxin from the venom of the false horned viper (Pseudocerastes fieldi). Toxicon 1995, 33, 863–874. [Google Scholar] [CrossRef]
- Tsai, M.; Lee, C.; Bdolah, A. Mode of neuromuscular blocking action of a toxic phospholipase A2 from Pseudocerastes fieldi (Field’s horned viper) snake venom. Toxicon 1983, 21, 527–534. [Google Scholar] [CrossRef]
- Bdolah, A.; Kinamon, S.; Batzri-Izraeli, R. The neurotoxic complex from the venom of Pseudocerastes fieldi. Contribution of the nontoxic subunit. Biochem. Int. 1985, 11, 627–636. [Google Scholar]
- Latifi, M. Variation in yield and lethality of venoms from Iranian snakes. Toxicon 1984, 22, 373–380. [Google Scholar] [CrossRef]
- Strickland, J.L.; Smith, C.F.; Mason, A.J.; Schield, D.R.; Borja, M.; Castañeda-Gaytán, G.; Spencer, C.L.; Smith, L.L.; Trápaga, A.; Bouzid, N.M.; et al. Evidence for divergent patterns of local selection driving venom variation in Mojave Rattlesnakes (Crotalus scutulatus). Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- David, P.; Vogel, G. Venomous Snakes of Europe Northern, Central and Western Asia; Edition Chimaira: Frankfurt am Main, Germany, 2010. [Google Scholar]
- Maduwage, K.P.; Scorgie, F.E.; Lincz, L.F.; O’Leary, M.A.; Isbister, G.K. Procoagulant snake venoms have differential effects in animal plasmas: Implications for antivenom testing in animal models. Throm. Res. 2016, 137, 174–177. [Google Scholar] [CrossRef]
- Doolittle, R.F. Coagulation in vertebrates with a focus on evolution and inflammation. J. Innate Immun. 2010, 3, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, R.F. The evolution of the vertebrate plasma proteins. Biol. Bull. 1987, 172, 269–283. [Google Scholar] [CrossRef]
- Youngman, N.J.; Zdenek, C.N.; Dobson, J.S.; Bittenbinder, M.A.; Gillett, A.; Hamilton, B.; Dunstan, N.; Allen, L.; Veary, A.; Veary, E.; et al. Mud in the blood: Novel potent anticoagulant coagulotoxicity in the venoms of the Australian elapid snake genus Denisonia (mud adders) and relative antivenom efficacy. Toxicol. Lett. 2019, 302, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, D.V.; Abe, A.S. Relationship of venom ontogeny and diet in Bothrops. Herpetologica 1999, 55, 200–204. [Google Scholar]
- National Center for Biotechnology Information (NCBI). Bethesda (MD): National Library of Medicine (US). Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 12 August 2020).
- Coimbra, F.C.; Dobson, J.; Zdenek, C.N.; Op den Brouw, B.; Hamilton, B.; Debono, J.; Masci, P.; Frank, N.; Ge, L.; Kwok, H.F.; et al. Does size matter? Venom proteomic and functional comparison between night adder species (Viperidae: Causus) with short and long venom glands. Comp. Biochem. Phys. Part C Toxicol. Pharm. 2018, 211, 7–14. [Google Scholar] [CrossRef]
- Minton, S.A.; Minton, M.R. Toxicity of some Australian snake venoms for potential prey species of reptiles and amphibians. Toxicon 1981, 19, 749–755. [Google Scholar] [CrossRef]
- Davies, E.-L.; Arbuckle, K. Coevolution of snake venom toxic activities and diet: Evidence that ecological generalism favours toxicological diversity. Toxins 2019, 11, 711. [Google Scholar] [CrossRef] [Green Version]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. B 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [Green Version]
- Zancolli, G.; Calvete, J.J.; Cardwell, M.D.; Greene, H.W.; Hayes, W.K.; Hegarty, M.J.; Herrmann, H.W.; Holycross, A.T.; Lannutti, D.I.; Mulley, J.F.; et al. When one phenotype is not enough: Divergent evolutionary trajectories govern venom variation in a widespread rattlesnake species. Proc. R. Soc. B 2019, 286, 20182735. [Google Scholar] [CrossRef] [Green Version]
- Dessauer, H.C. Blood chemistry of reptiles: Physiological and evolutionary aspects. Biol. Reptil. 1970, 3, 1–72. [Google Scholar]
- Johnson, G.S.; Turrentine, M.A.; Swayne, D.E. Coagulation of plasma from the chicken (Gallus domesticus): Phospholipids influence clotting rates induced by components from Russell’s viper venom. Comp. Biochem. Phys. Part B Biochem. Mol. Biol. 1985, 82, 647–653. [Google Scholar] [CrossRef]
- Stopforth, A. A study of coagulation mechanisms in domestic chickens. J. Comp. Path. 1970, 80, 525–533. [Google Scholar] [CrossRef]
- Hackett, E.; Lepage, R. The clotting of the blood of an amphibian, Bufo marinus. Aust. J. Exp. Biol. Med. Sci. 1961, 39, 67–78. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. urarachnoides Venom vs. Spontaneous Control Difference Between Means ± SD (%); Unpaired t-Test | ||
|---|---|---|
| Plasma (n = 4) | Clot Initiation Speed | Clot Formation Rate |
| Human | 95 ± 4.3%; t (6) = 22.08, p < 0.000001 | 61 ± 14.8%; t (6) = 4.08, p = 0.00651 |
| Toad | 99 ± 4.3%; t (6) = 23.15, p < 0.000001 | 63 ± 2.6%; t (6) = 24.37, p < 0.000001 |
| Chicken | 96 ± 4.3%; t (6) = 22.46, p < 0.000001 | 38 ± 10.9%; t (6) = 3.519, p = 0.01254 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
op den Brouw, B.; Coimbra, F.C.P.; Bourke, L.A.; Huynh, T.M.; Vlecken, D.H.W.; Ghezellou, P.; Visser, J.C.; Dobson, J.S.; Fernandez-Rojo, M.A.; Ikonomopoulou, M.P.; et al. Extensive Variation in the Activities of Pseudocerastes and Eristicophis Viper Venoms Suggests Divergent Envenoming Strategies Are Used for Prey Capture. Toxins 2021, 13, 112. https://doi.org/10.3390/toxins13020112
op den Brouw B, Coimbra FCP, Bourke LA, Huynh TM, Vlecken DHW, Ghezellou P, Visser JC, Dobson JS, Fernandez-Rojo MA, Ikonomopoulou MP, et al. Extensive Variation in the Activities of Pseudocerastes and Eristicophis Viper Venoms Suggests Divergent Envenoming Strategies Are Used for Prey Capture. Toxins. 2021; 13(2):112. https://doi.org/10.3390/toxins13020112
Chicago/Turabian Styleop den Brouw, Bianca, Francisco C. P. Coimbra, Lachlan A. Bourke, Tam Minh Huynh, Danielle H. W. Vlecken, Parviz Ghezellou, Jeroen C. Visser, James S. Dobson, Manuel A. Fernandez-Rojo, Maria P. Ikonomopoulou, and et al. 2021. "Extensive Variation in the Activities of Pseudocerastes and Eristicophis Viper Venoms Suggests Divergent Envenoming Strategies Are Used for Prey Capture" Toxins 13, no. 2: 112. https://doi.org/10.3390/toxins13020112
APA Styleop den Brouw, B., Coimbra, F. C. P., Bourke, L. A., Huynh, T. M., Vlecken, D. H. W., Ghezellou, P., Visser, J. C., Dobson, J. S., Fernandez-Rojo, M. A., Ikonomopoulou, M. P., Casewell, N. R., Ali, S. A., Fathinia, B., Hodgson, W. C., & Fry, B. G. (2021). Extensive Variation in the Activities of Pseudocerastes and Eristicophis Viper Venoms Suggests Divergent Envenoming Strategies Are Used for Prey Capture. Toxins, 13(2), 112. https://doi.org/10.3390/toxins13020112