Polydatin Protects Bovine Mammary Epithelial Cells against Zearalenone-Induced Apoptosis by Inhibiting Oxidative Responses and Endoplasmic Reticulum Stress

 , ,
, ,
Abstract
:1. Introduction
2. Results
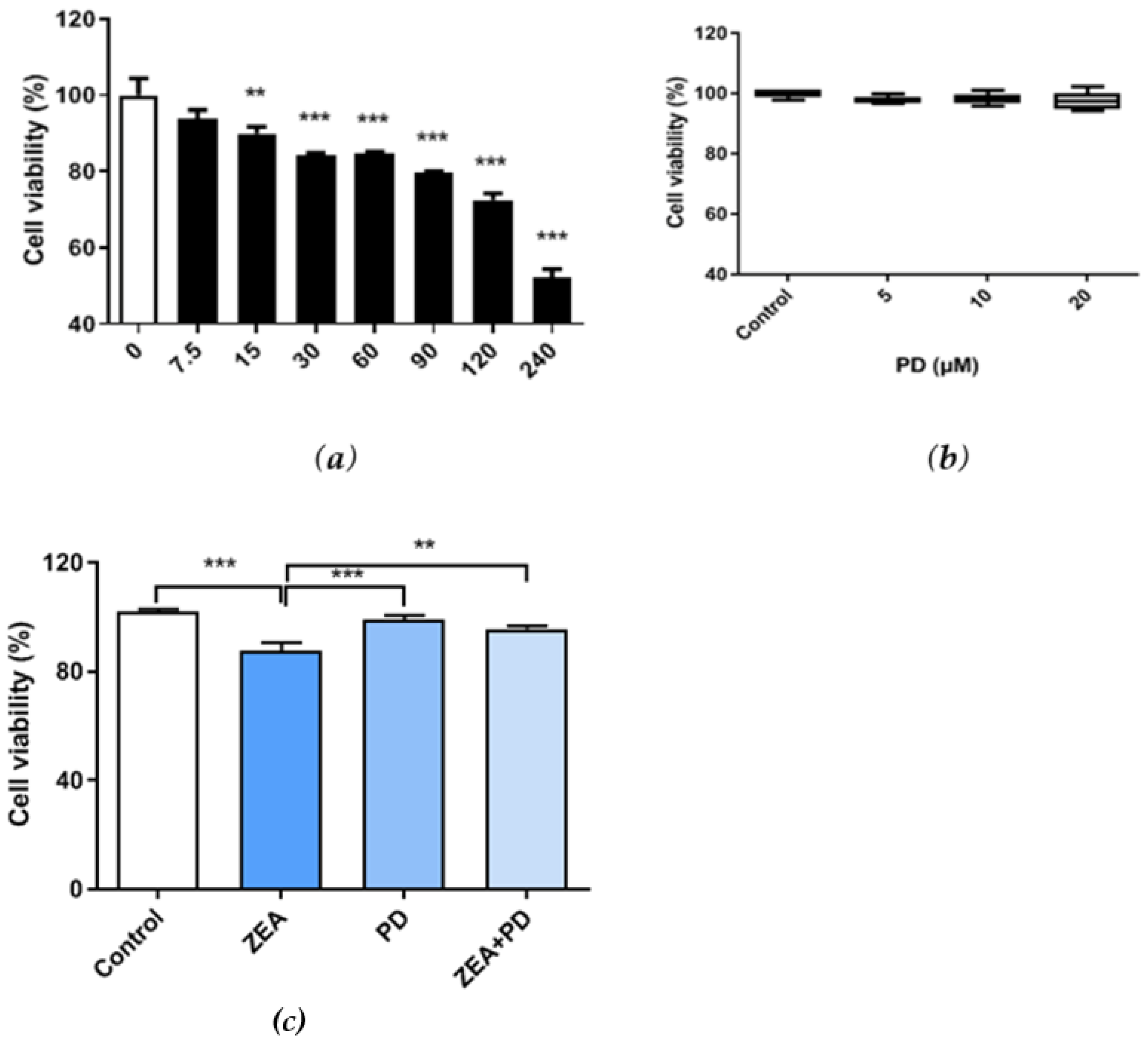
2.1. The Increase in PD Decreased the MAC-T Cell Viability Due to ZEA
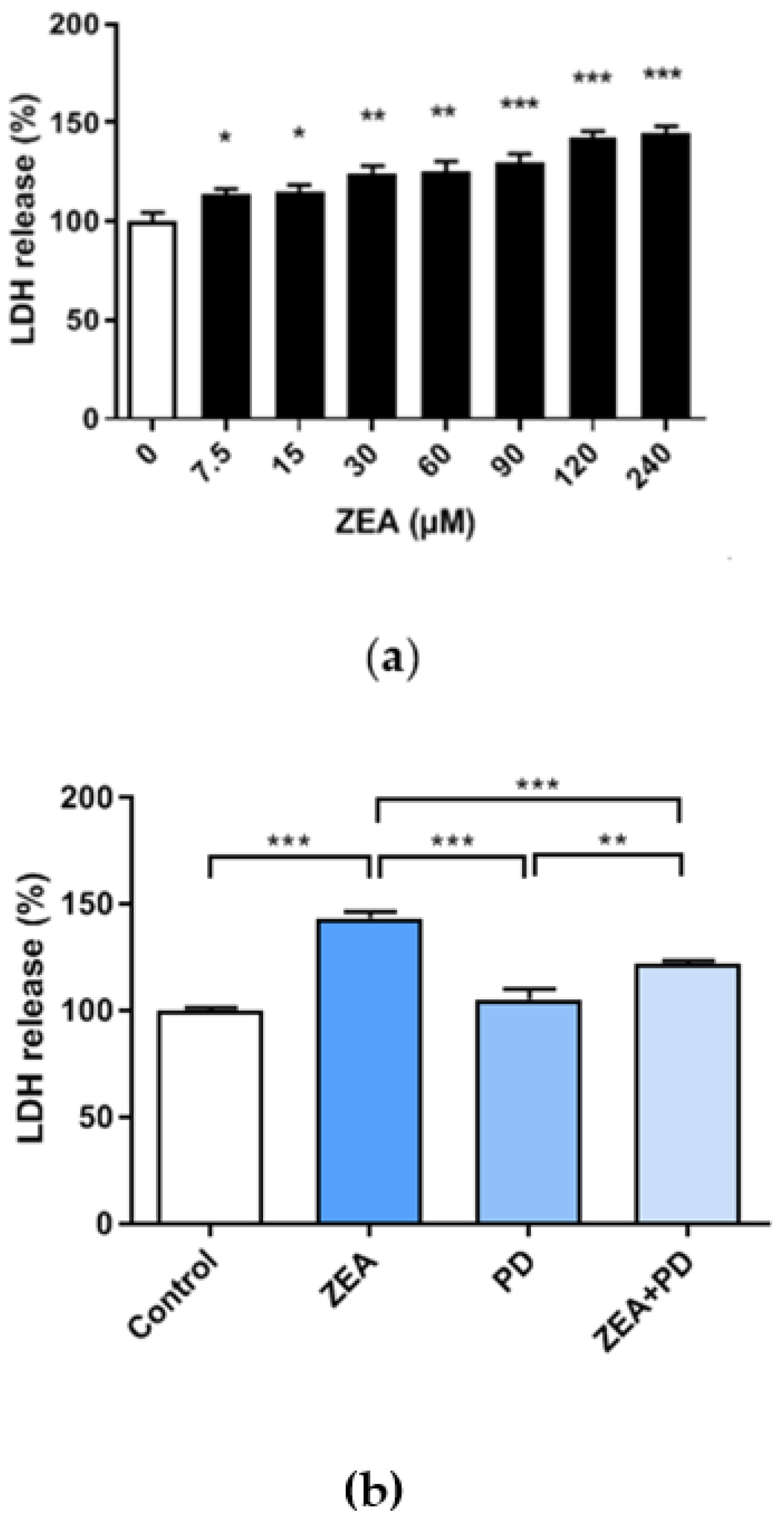
2.2. PD Reduces the ZEA-Induced Cytotoxic Effect in MAC-T Cells
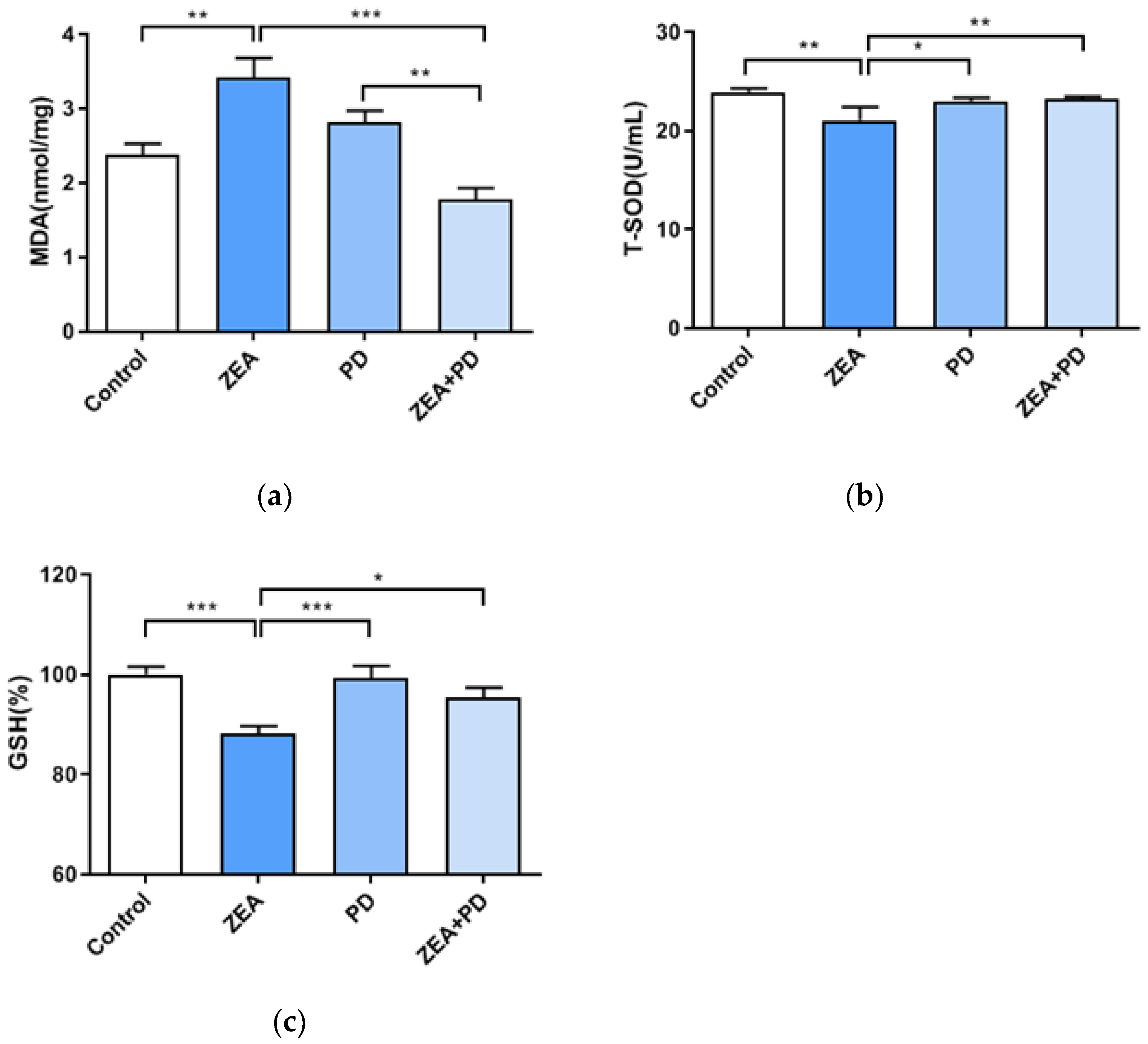
2.3. PD Can Inhibit ZEA-Induced Oxidative Damage
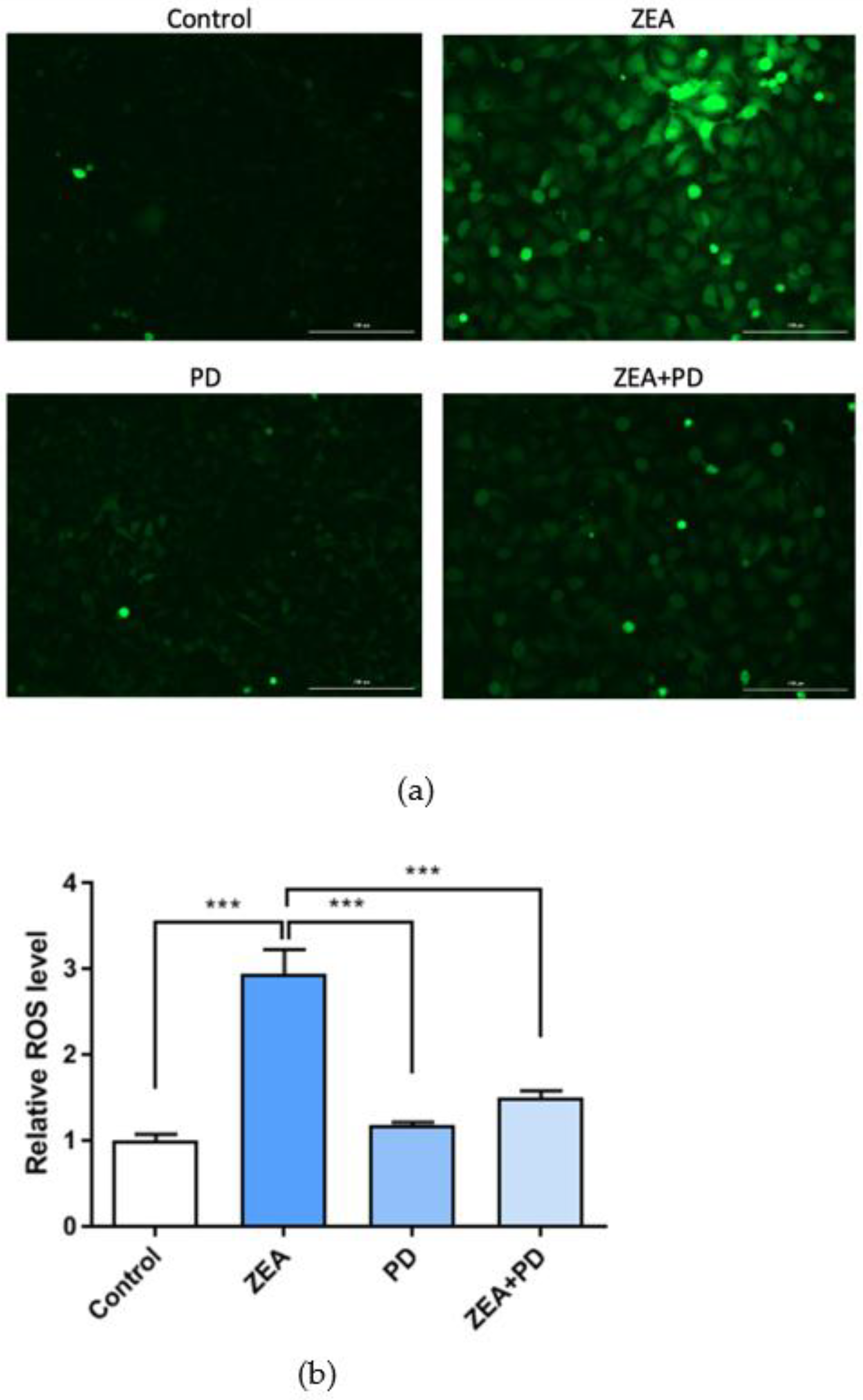
2.4. PD Inhibits ZEA-Induced Increases in ROS Levels
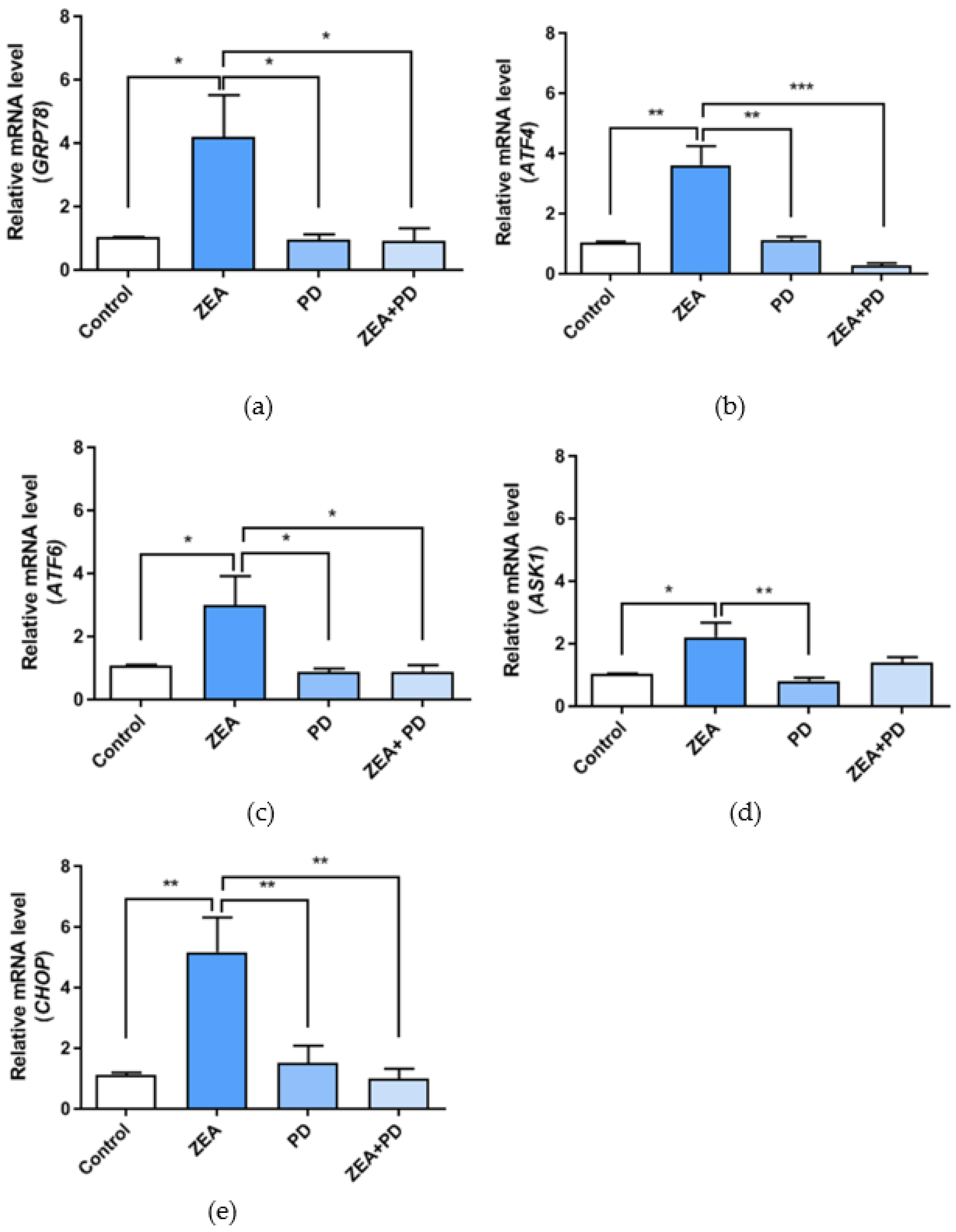
2.5. PD Inhibits ZEA-Induced ER Stress in MAC-T Cells
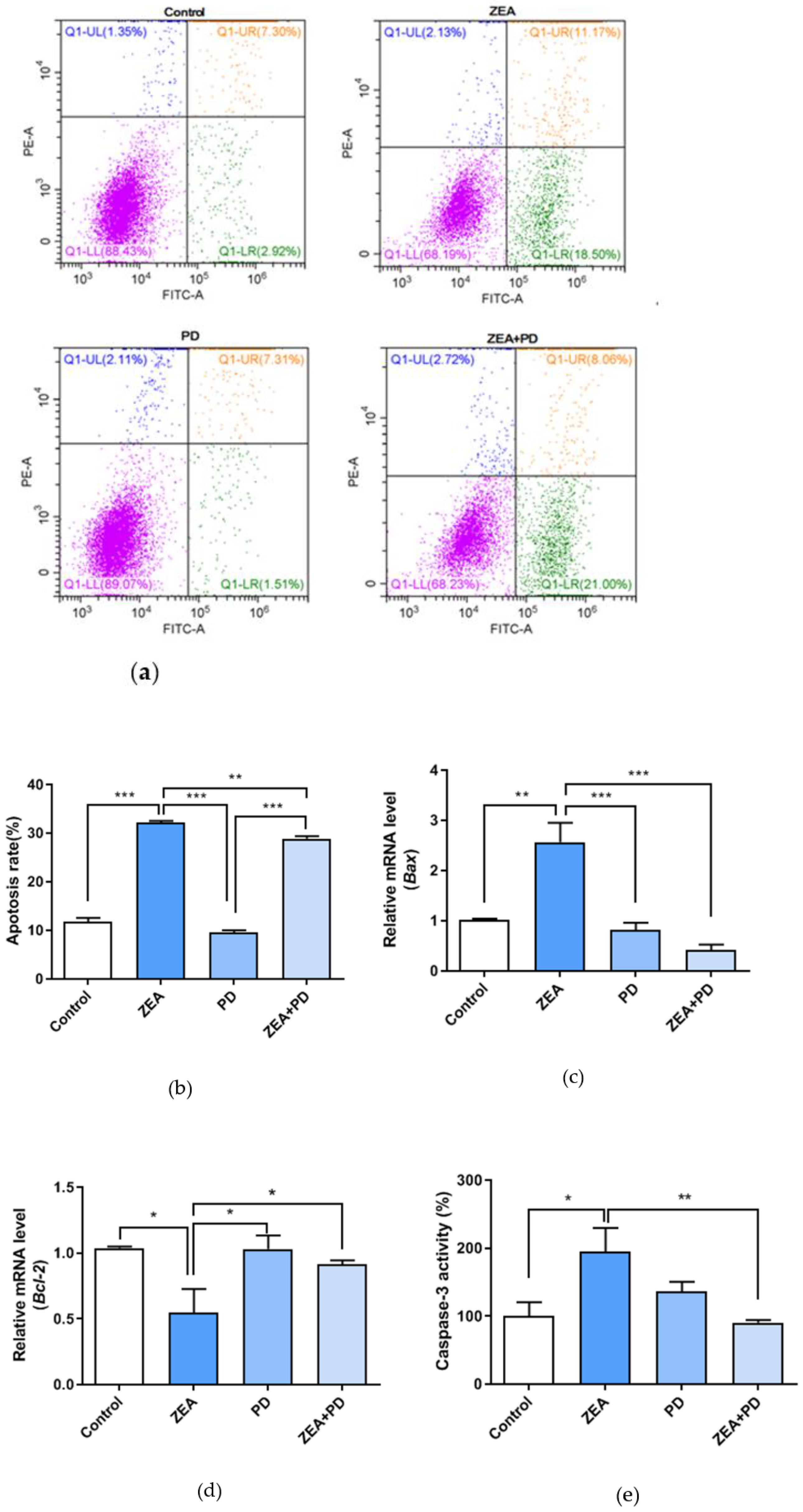
2.6. PD Inhibits ZEA-Induced Apoptosis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cell Culture
5.3. Cell Viability Assay
5.4. Lactate Dehydrogenase Assay
5.5. Detection of GSH, T-SOD, and MDA Levels
5.6. Measurement of ROS Production
5.7. qPCR
5.8. Flow Cytometry Detection of Apoptosis
5.9. Caspase-3 Activity Assay
5.10. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kouadio, J.H.; Mobio, T.A.; Baudrimont, I.; Moukha, S.; Dano, S.D.; Creppy, E.E. Comparative study of cytotoxicity and oxidative stress induced by deoxynivalenol, zearalenone or fumonisin B1 in human intestinal cell line Caco-2. Toxicology 2005, 213, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, U.; Brussow, K.P.; Dannenberger, D.; Jonas, L.; Pohland, R.; Jager, K.; Danicke, S.; Hagemann, E. The effect of feeding a diet naturally contaminated with deoxynivalenol (DON) and zearalenone (ZON) on the spleen and liver of sow and fetus from day 35 to 70 of gestation. Toxicol. Lett. 2008, 179, 113–117. [Google Scholar] [CrossRef]
- Liu, M.; Gao, R.; Meng, Q.; Zhang, Y.; Bi, C.; Shan, A. Toxic effects of maternal zearalenone exposure on intestinal oxidative stress, barrier function, immunological and morphological changes in rats. PLoS ONE 2014, 9, e106412. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Yang, W.; Feng, Q.; Huang, L.; Zhang, G.; Liu, F.; Jiang, S.; Yang, Z. Effects of purified zearalenone on selected immunological measurements of blood in post-weaning gilts. Anim. Nutr. 2016, 2, 142–148. [Google Scholar] [CrossRef]
- Pietsch, C.; Noser, J.; Wettstein, F.E.; Burkhardt-Holm, P. Unraveling the mechanisms involved in zearalenone-mediated toxicity in permanent fish cell cultures. Toxicon Off. J. Int. Soc. Toxinology 2014, 88, 44–61. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [CrossRef]
- Wang, C.C.; Chiang, Y.M.; Sung, S.C.; Hsu, Y.L.; Chang, J.K.; Kuo, P.L. Plumbagin induces cell cycle arrest and apoptosis through reactive oxygen species/c-Jun N-terminal kinase pathways in human melanoma A375.S2 cells. Cancer Lett. 2008, 259, 82–98. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Kim, M.O.; Kang, S.H.; Choi, Y.H.; Kim, G.Y. Sulforaphane suppresses TNF-alpha-mediated activation of NF-kappaB and induces apoptosis through activation of reactive oxygen species-dependent caspase-3. Cancer Lett. 2009, 274, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.R.; Jin, Y.C.; Zhao, Y.; Shan, A.; Fang, H.T.; Shen, J.L.; Zhou, C.H.; Yu, H.; Zhou, Y.F.; Wang, X.; et al. Zearalenone induces apoptosis inbovine mammary epithelial cells by activating endoplasmic reticulum stress. J. Dairy Sci. 2019, 102, 10543–10553. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.X.; Tanaka, L.Y.; Wosniak, J.; Laurindo, F.R. Mechanisms and implications of reactive oxygen species generation during the unfolded protein response: Roles of endoplasmic reticulum oxidoreductases, mitochondrial electron transport, and NADPH oxidase. Antioxid. Redox Signal. 2009, 11, 2409–2427. [Google Scholar] [CrossRef]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T.; Samini, F. Anti-oxidative effects of curcumin on immobilization-induced oxidative stress in rat brain, liver and kidney. Biomed. Pharmacother. 2017, 87, 223–229. [Google Scholar] [CrossRef]
- Lin, P.; Chen, F.; Sun, J.; Zhou, J.; Wang, X.; Wang, N.; Li, X.; Zhang, Z.; Wang, A.; Jin, Y. Mycotoxin zearalenone induces apoptosis in mouse Leydig cells via an endoplasmic reticulum stress-dependent signalling pathway. Reprod. Toxicol. 2015, 52, 71–77. [Google Scholar] [CrossRef]
- Kadowaki, H.; Nishitoh, H.; Ichijo, H. Survival and apoptosis signals in ER stress: The role of protein kinases. J. Chem. Neuroanat. 2004, 28, 93–100. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a. double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and. human disease. Antioxid. Redox Signal. 2014, 21, 396–413. [Google Scholar] [CrossRef]
- Wang, H.L.; Gao, J.P.; Han, Y.L.; Xu, X.; Wu, R.; Gao, Y. Comparative studies of polydatin and resveratrol on mutual transformation and antioxidative effect in vivo. Phytomedicine 2015, 22, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm. Biol. 2013, 51, 1347–1354. [Google Scholar] [CrossRef]
- Ji, H.; Zhang, X.; Du, Y.; Liu, H.; Li, S.; Li, L. Polydatin modulates inflammation by decreasing NF-kappaB activation and oxidative stress by increasing Gli1, Ptch1, SOD1 expression and ameliorates blood-brain barrier permeability for its neuroprotective effect in pMCAO rat brain. Brain Res. Bull. 2012, 87, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, W.; Deng, J.; Lan, L.; Xue, X.; Zhang, C.; Cai, G.; Luo, X.; Liu, J. Polydatin protects cardiac function against burn injury by inhibiting sarcoplasmic reticulum Ca2+ leak by reducing oxidative modification of ryanodine receptors. Free Radic. Biol. Med. 2013, 60, 292–299. [Google Scholar] [CrossRef]
- Ye, J.; Piao, H.; Jiang, J.; Jin, G.; Zheng, M.; Yang, J.; Jin, X.; Sun, T.; Choi, Y.H.; Li, L.; et al. Polydatin inhibits mast cell-mediated allergic inflammation by targeting PI3K/Akt, MAPK, NF-kappaB and Nrf2/HO-1 pathways. Sci. Rep. 2017, 7, 11895. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.S.; Jin, C.; Huang, X.; Liu, J.; Yan, W.S.; Huang, Q.; Kan, W. The mechanism of Polydatin in shock treatment. Clin. Hemorheol. Microcirc. 2003, 29, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Hou, Y.; Lin, D. Polydatin Protects Bone Marrow Stem Cells against Oxidative Injury: Involvement of Nrf 2/ARE Pathways. Stem Cells Int. 2016, 9394150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Flores-Flores, M.E.; Lizarraga, E.; de Cerain, A.L.; González-Peñas, E. Presence of mycotoxins in animal milk: A review. Food Control 2015, 53, 163–176. [Google Scholar] [CrossRef]
- Hassen, W.; Ayed-Boussema, I.; Oscoz, A.A.; Ade, C.L.; Bacha, H. The role of oxidative stress in zearalenone-mediated toxicity in Hep G2 cells: Oxidative DNA damage, gluthatione depletion and stress proteins induction. Toxicology 2007, 232, 294–302. [Google Scholar] [CrossRef]
- Wu, M.; Liu, M.; Guo, G.; Zhang, W.; Liu, L. Polydatin Inhibits Formation of Macrophage-Derived Foam Cells. Evid.-Based Complementary Altern. Med. 2015, 729017. [Google Scholar] [CrossRef] [Green Version]
- Koneru, M.; Sahu, B.D.; Gudem, S.; Kuncha, M.; Ravuri, H.G.; Kumar, J.M.; Kilari, E.K.; Sistla, R. Polydatin alleviates alcohol-induced acute liver injury in mice: Relevance of matrix metalloproteinases (MMPs) and hepatic anti-oxidants. Phytomedicine Int. J. Phytother. Phytopharm. 2017, 27, 23–32. [Google Scholar] [CrossRef]
- Ince, S.; Avdatek, F.; Demirel, H.H.; Arslan-Acaroz, D.; Goksel, E.; Kucukkurt, I. Ameliorative effect of polydatin on oxidative stress-mediated testicular damage by chronic arsenic exposure in rats. Andrologia 2016, 48, 518–524. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, S.; Zhang, Y.; Wu, J.; Peng, H.; Fan, J.; Liao, J. Reactive oxygen species-mediated endoplasmic reticulum stress and mitochondrial dysfunction contribute to polydatin-induced apoptosis in human nasopharyngeal carcinoma CNE cells. J. Cell. Biochem. 2011, 112, 3695–3703. [Google Scholar] [CrossRef] [PubMed]
- Ben Salah-Abbes, J.; Abbes, S.; Abdel-Wahhab, M.A.; Oueslati, R. Raphanus sativus extract protects against Zearalenone induced reproductive toxicity, oxidative stress and mutagenic alterations in male Balb/c mice. Toxicon Off. J. Int. Soc. Toxinology 2009, 53, 525–533. [Google Scholar] [CrossRef]
- Li, R.P.; Wang, Z.Z.; Sun, M.X.; Hou, X.L.; Sun, Y.; Deng, Z.F.; Xiao, K. Polydatin protects learning and memory impairments in a rat model of vascular dementia. Phytomedicine Int. J. Phytother. Phytopharm. 2012, 19, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Quan, Z.; Gu, J.; Dong, P.; Lu, J.; Wu, X.; Wu, W.; Fei, X.; Li, S.; Wang, Y.; Wang, J.; et al. Reactive oxygen species-mediated endoplasmic reticulum stress and mitochondrial dysfunction contribute to cirsimaritin-induced apoptosis in human gallbladder carcinoma GBC-SD cells. Cancer Lett. 2010, 295, 252–259. [Google Scholar] [CrossRef]
- Ben Salem, I.; Prola, A.; Boussabbeh, M.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. Crocin and Quercetin protect HCT116 and HEK293 cells from Zearalenone-induced apoptosis by reducing endoplasmic reticulum stress. Cell Stress Chaperones 2015, 20, 927–938. [Google Scholar] [CrossRef] [Green Version]
- Banjerdpongchai, R.; Kongtawelert, P.; Khantamat, O.; Srisomsap, C.; Chokchaichamnankit, D.; Subhasitanont, P. Mitochondrial and endoplasmic reticulum stress pathways cooperate in zearalenone-induced apoptosis of human leukemic cells. J. Hematol. Oncol. 2010, 3, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Jiang, K.; Wu, H.; Qiu, C.; Deng, G.; Peng, X. Polydatin reduces Staphylococcus aureus lipoteichoic acid-induced injury by attenuating reactive oxygen species generation and TLR2-NFkappaB signalling. J. Cell. Mol. Med. 2017, 21, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ni, M.; Lee, B.; Barron, E.; Hinton, D.R.; Lee, A.S. The unfolded protein response regulator GRP78/BiP is required for endoplasmic reticulum integrity and stress-induced autophagy in mammalian cells. Cell Death Differ. 2008, 15, 1460–1471. [Google Scholar] [CrossRef] [PubMed]
- Dvash, E.; Har-Tal, M.; Barak, S.; Meir, O.; Rubinstein, M. Leukotriene C4 is the major trigger of stress-induced oxidative DNA damage. Nat. Commun. 2015, 6, 10112. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-Q.; Yuan, A.-H.; Yang, J.; Bi-xiang, Z.H.A.; Zhang, M. Effect of acupuncture on the endoplasmic reticulum stress IRE1-CHOP pathway and the expression levels of Bax and Bcl-2 protein as well as genes in pancreatic tissue of rats with diabetes mellitus. World J. Acupunct. Moxibustion 2017, 27, 41–46. [Google Scholar] [CrossRef]
- Liu, Y.H.; Huang, Q.H.; Wu, X.; Wu, J.Z.; Liang, J.L.; Lin, G.S.; Xu, L.Q.; Lai, X.P.; Su, Z.R.; Chen, J.N. Polydatin protects against acetaminophen-induced hepatotoxicity in mice via anti-oxidative and anti-apoptotic activities. Food Funct. 2018, 9, 5891–5902. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | GenBank Accession No. | Product Size (bp) |
|---|---|---|---|---|
| β-actin | 5′-CCCTGGAGAAGAGCTACGAG-3′ | 5′-GTAGTTTCGTGAATGCCGCAG-3′ | NM_173979.3 | 130 |
| GRP78 | 5′-CGACCCCTGACGAAAGACAA-3′ | 5′-AGGTGTCAGGCGATTTTGGT-3′ | NM_001075148.1 | 198 |
| ATF4 | 5′-AGATGACCTGGAAACCATGC-3′ | 5′-AGGGGGAAGAGGTTGAAAGA-3′ | NM_001034342.2 | 190 |
| ATF6 | 5′-ATATTCCTCCGCCTCCCTGT-3′ | 5′-GTCCTTTCCACTTCGTGCCT-3′ | XM_024989876.1 | 103 |
| ASK1 | 5′-GCTATGGAAAGGCAGCCAGA-3′ | 5′-TCTGCTGACATGGACTCTGG-3′ | NM_001144081.2 | 160 |
| CHOP | 5′-GAGCTGGAAGCCTGGTATGA-3′ | 5′-CTCCTTGTTTCCAGGGGGTG-3′ | NM_001078163.1 | 90 |
| Bax | 5′-GCTCTGAGCAGATCATGAAGAC-3′ | 5′-CAATTCATCTCCGATGCGCT-3′ | NM_173894.1 | 167 |
| Bcl-2 | 5′-GATGACCGAGTACCTGAACC-3′ | 5′-AGAGACAGCCAGGAGAAATCA-3′ | NM_001166486.1 | 123 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Jin, Y.; Shan, A.; Zhang, J.; Tang, H.; Shen, J.; Zhou, C.; Yu, H.; Fang, H.; Zhao, Y.; et al. Polydatin Protects Bovine Mammary Epithelial Cells against Zearalenone-Induced Apoptosis by Inhibiting Oxidative Responses and Endoplasmic Reticulum Stress. Toxins 2021, 13, 121. https://doi.org/10.3390/toxins13020121
Fu Y, Jin Y, Shan A, Zhang J, Tang H, Shen J, Zhou C, Yu H, Fang H, Zhao Y, et al. Polydatin Protects Bovine Mammary Epithelial Cells against Zearalenone-Induced Apoptosis by Inhibiting Oxidative Responses and Endoplasmic Reticulum Stress. Toxins. 2021; 13(2):121. https://doi.org/10.3390/toxins13020121
Chicago/Turabian StyleFu, Yurong, Yongcheng Jin, Anshan Shan, Jing Zhang, Hongyu Tang, Jinglin Shen, Changhai Zhou, Hao Yu, Hengtong Fang, Yun Zhao, and et al. 2021. "Polydatin Protects Bovine Mammary Epithelial Cells against Zearalenone-Induced Apoptosis by Inhibiting Oxidative Responses and Endoplasmic Reticulum Stress" Toxins 13, no. 2: 121. https://doi.org/10.3390/toxins13020121
APA StyleFu, Y., Jin, Y., Shan, A., Zhang, J., Tang, H., Shen, J., Zhou, C., Yu, H., Fang, H., Zhao, Y., Wang, J., & Tian, Y. (2021). Polydatin Protects Bovine Mammary Epithelial Cells against Zearalenone-Induced Apoptosis by Inhibiting Oxidative Responses and Endoplasmic Reticulum Stress. Toxins, 13(2), 121. https://doi.org/10.3390/toxins13020121