Bacillus cereus Induces Severe Infections in Preterm Neonates: Implication at the Hospital and Human Milk Bank Level

 ,
,  ,
,
Abstract
:1. Introduction
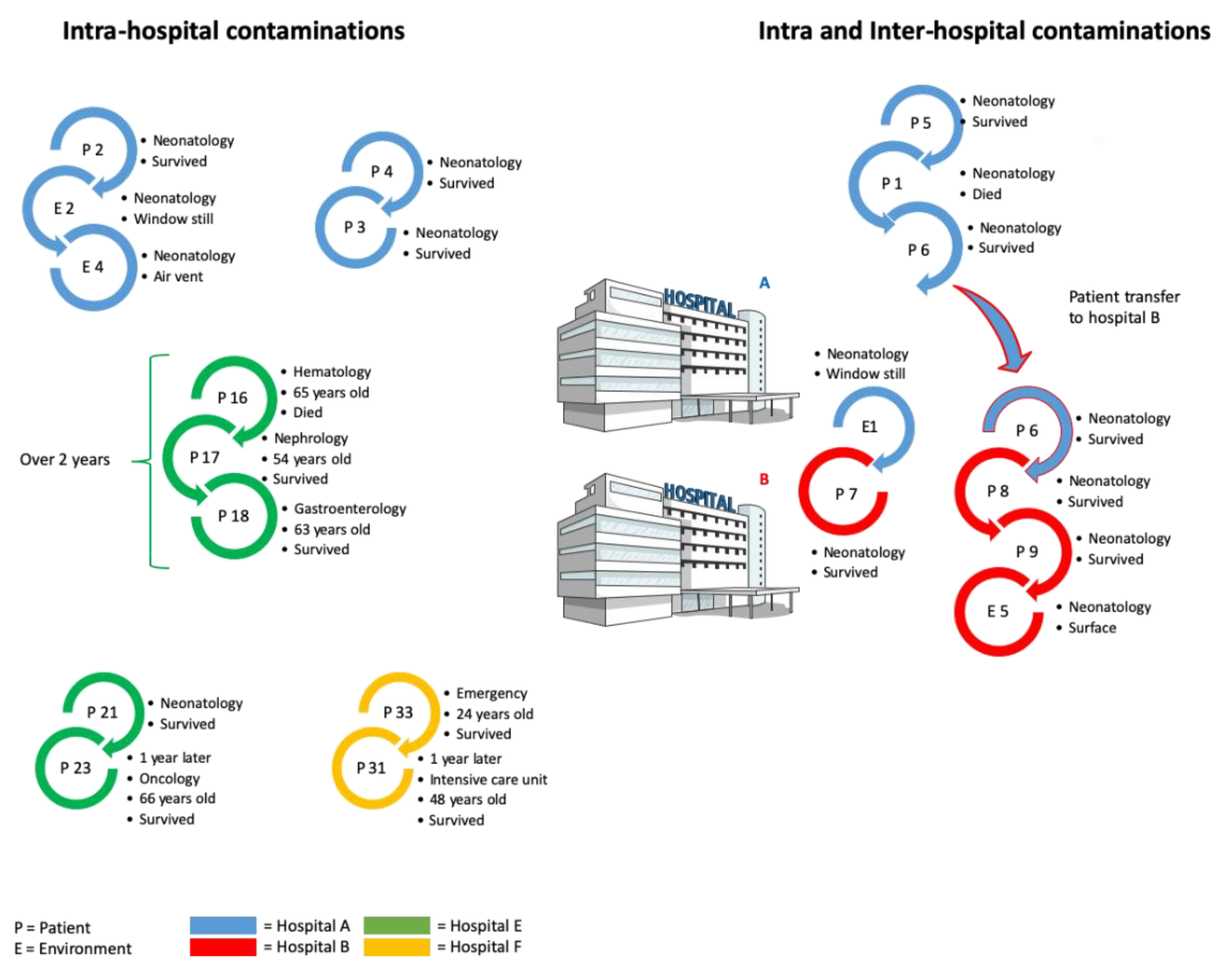
2. B. cereus Induces Severe Pathologies in Preterm Neonates
3. Toxins Potentially Involved in B. cereus Infection
4. Contamination Sources
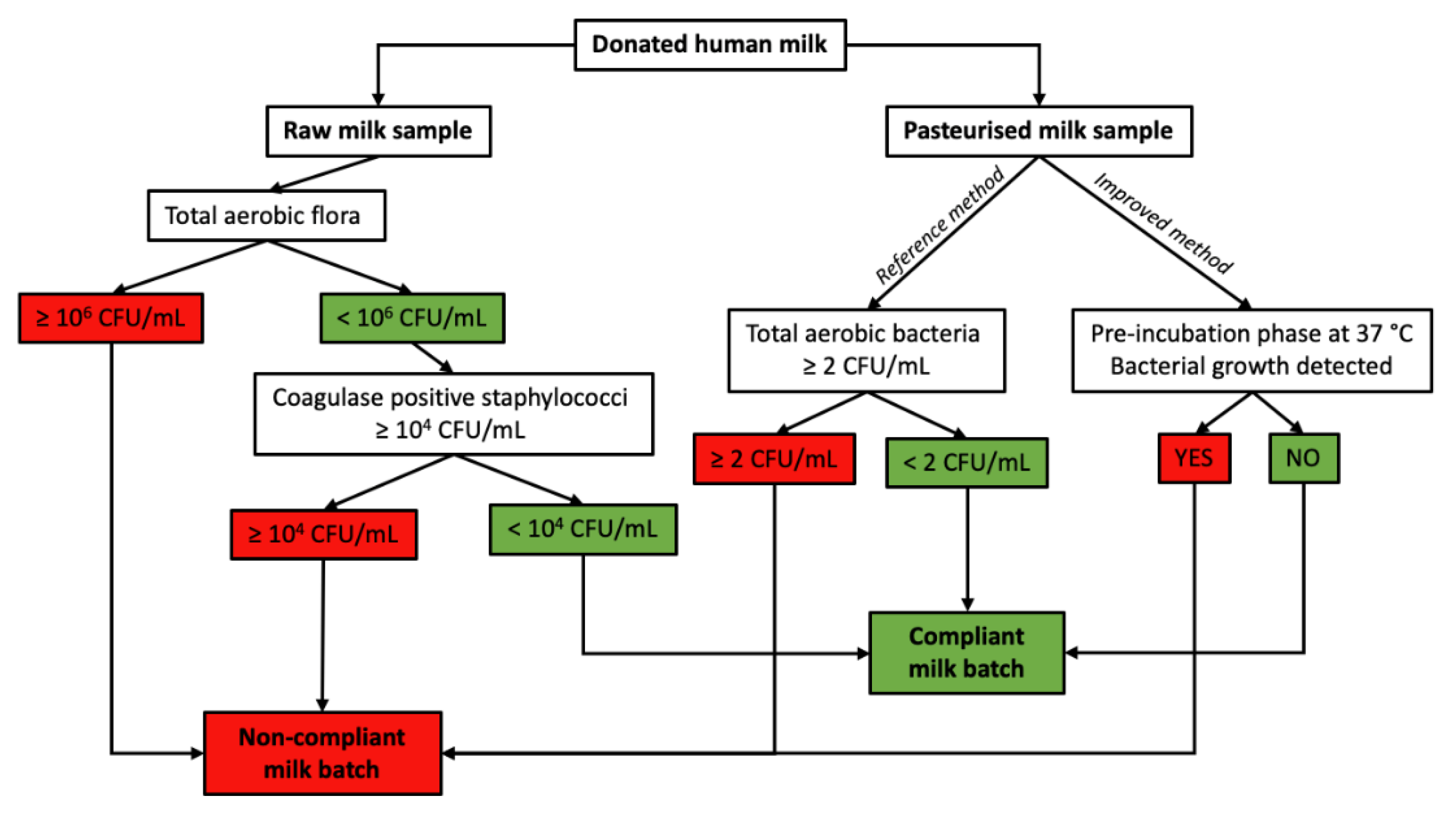
5. Food Safety Regulation
6. Human Milk Banks—Methods, Practices and Issues
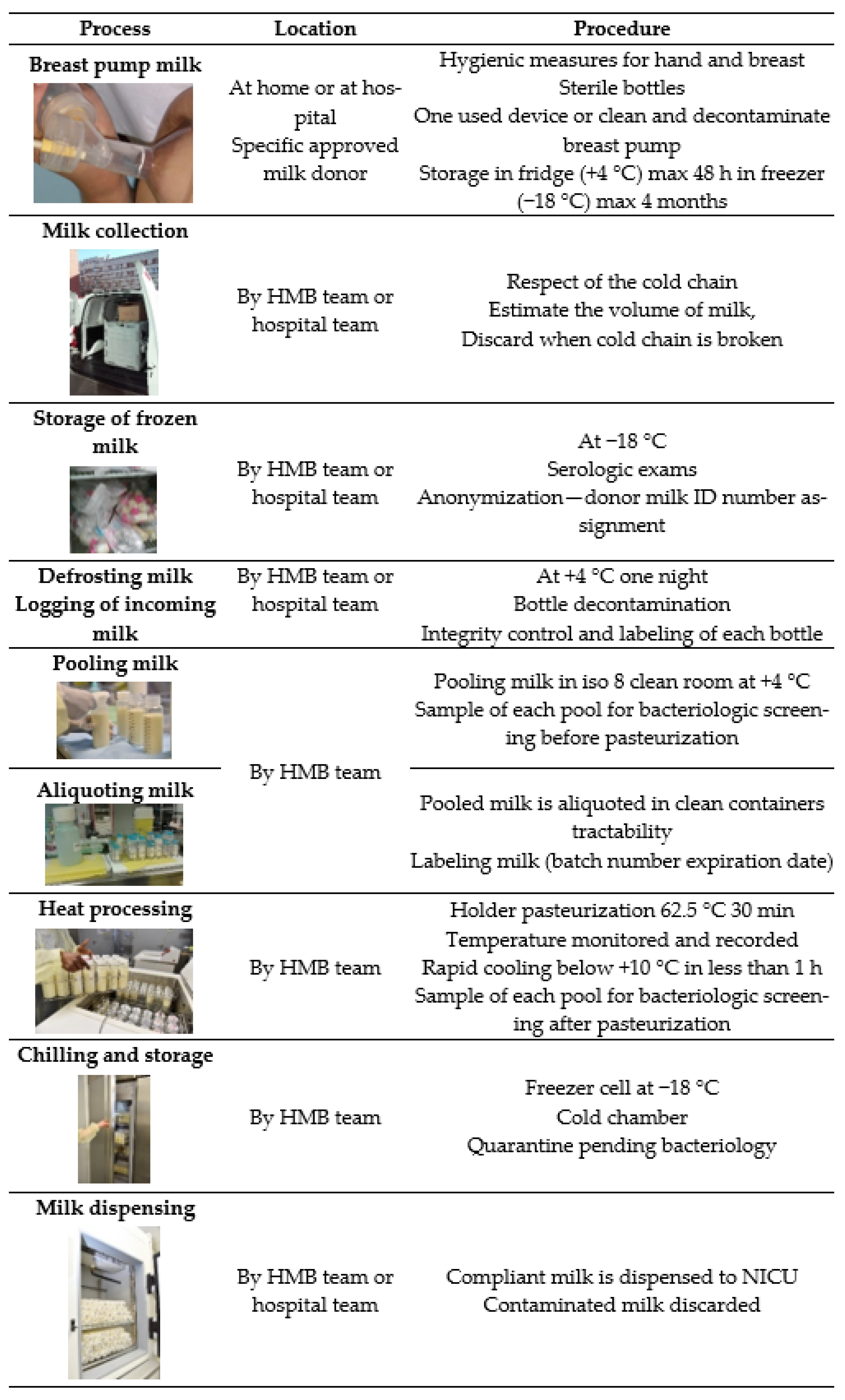
6.1. General Hygiene Procedure and Milk Treatment
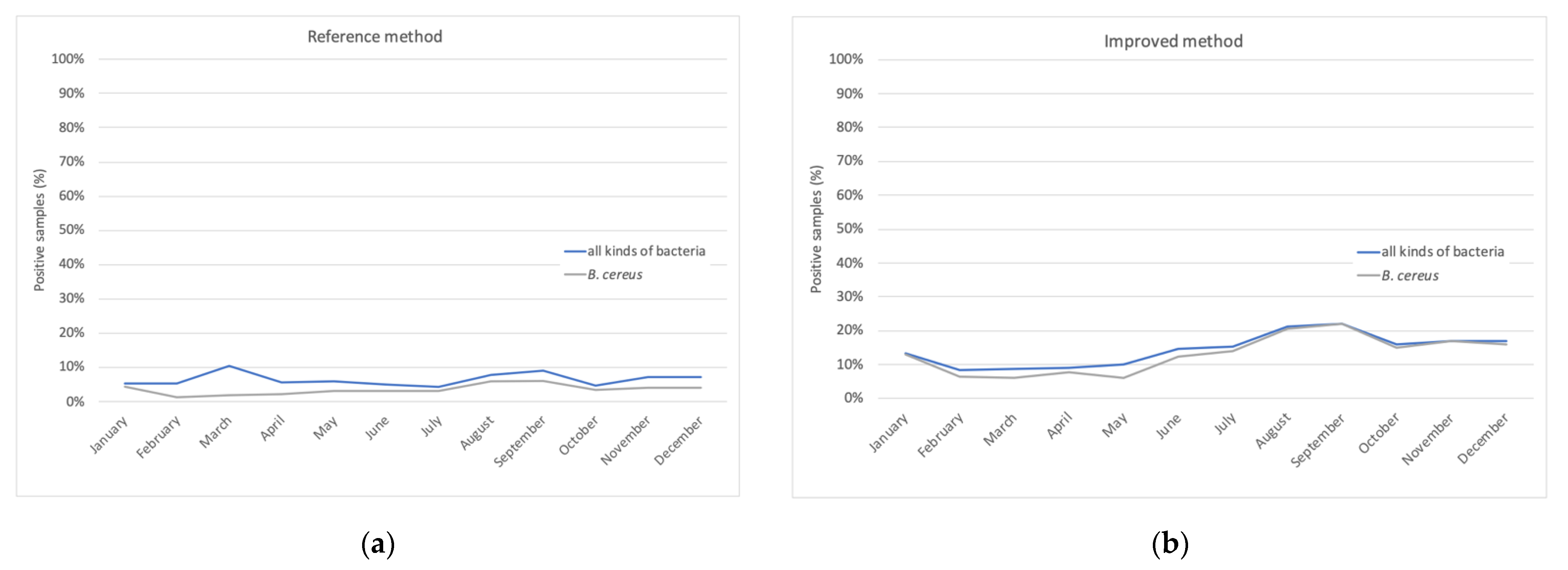
6.2. B. cereus in HBM
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Picaud, J.C.; Buffin, R.; Gremmo-Feger, G.; Rigo, J.; Putet, G.; Casper, C.; Working group of the French Neonatal Society on fresh human milk use in preterm infants. Review concludes that specific recommendations are needed to harmonise the provision of fresh mother’s milk to their preterm infants. Acta Paediatr. 2018, 107, 1145–1155. [Google Scholar] [CrossRef]
- Moro, G.E.; Billeaud, C.; Rachel, B.; Calvo, J.; Cavallarin, L.; Christen, L.; Escuder-Vieco, D.; Gaya, A.; Lembo, D.; Wesolowska, A.; et al. Processing of donor human milk: Update and recommendations from the european milk bank association (EMBA). Front. Pediatr. 2019, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, G.; Bertino, E.; Gebauer, C.; Grovslien, A.; Mileusnic-Milenovic, R.; Arslanoglu, S.; Barnett, D.; Boquien, C.-Y.; Buffin, R.; Gaya, A.; et al. Recommendations for the establishment and operation of human milk banks in Europe: A consensus statement from the european milk bank association (EMBA). Front. Pediatr. 2019, 7, 53. [Google Scholar] [CrossRef]
- Lewin, A.; Quach, C.; Rigourd, V.; Picaud, J.-C.; Perreault, T.; Frange, P.; Domingo, M.-C.; Lalancette, C.; Delage, G.; Germain, M. Bacillus cereus infection in neonates and the absence of evidence for the role of banked human milk: Case reports and literature review. Infect. Control. Hosp. Epidemiol. 2019, 40, 787–793. [Google Scholar] [CrossRef]
- Decousser, J.-W.; RamaRao, N.; Duport, C.; Dorval, M.; Bourgeois-Nicolaos, N.; Guinebretière, M.-H.; Razafimahefa, H.; Doucet-Populaire, F. Bacillus cereus and severe intestinal infections in preterm neonates: Putative role of pooled breast milk. Am. J. Infect. Control 2013, 41, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Schraft, H.; Odumeru, J.A.; Griffiths, M.W. Identification of contamination sources of Bacillus cereus in pasteurized milk. Int. J. Food Microbiol. 1998, 43, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Stenfors Arnesen, L.; Fagerlund, A.; Granum, P. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [Green Version]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.-A.; Ramarao, N.; et al. Large-scale survey of Bacillus cereus-induced food-borne outbreaks: Epidemiologic and genetic characterization. Eurosurveillance 2016, 21, 30413. [Google Scholar]
- European Food Safety Authority ECfDPaC. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSa J. 2018, 13. [Google Scholar]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, V.M. You Can’t B. cereus-A review of Bacillus cereus strains that cause anthrax-like disease. Front. Microbiol. 2020, 11, 1731. [Google Scholar] [CrossRef] [PubMed]
- Lequin, M.H.; Vermeulen, J.R.; Van Elburg, R.M.; Barkhof, F.; Kornelisse, R.F.; Swarte, R.; Govaert, P.P. Bacillus cereus meningoencephalitis in preterm infants: Neuroimaging characteristics. AJNR Am. J. Neuroradiol. 2005, 26, 2137–2143. [Google Scholar] [PubMed]
- Heep, A.; Schaller, C.; Rittmann, N.; Himbert, U.; Bartmann, P. Multiple brain abscesses in an extremely preterm infant: Treatment surveillance with interleukin-6 in the CSF. Eur. J. Nucl. Med. Mol. Imaging 2003, 163, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Jevon, G.P.; Dunne, W.M., Jr.; Hicks, M.J.; Langston, C. Bacillus cereus pneumonia in premature neonates: A report of two cases. Pediatr. Infect. Dis. J. 1993, 12, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Steen, M.K.; Bruno-Murtha, L.A.; Chaux, G.; Lazar, H.; Bernard, S.; Sulis, C. Bacillus cereus endocarditis: Report of a Case and Review. Clin. Infect. Dis. 1992, 14, 945–946. [Google Scholar] [CrossRef]
- David, D.B.; Kirkby, G.R.; Noble, B.A. Bacillus cereus endophthalmitis. Br. J. Ophthalmol. 1994, 78, 577–580. [Google Scholar] [CrossRef] [Green Version]
- Glasset, B.; Herbin, S.; Granier, S.A.; Cavalié, L.; Lafeuille, E.; Guérin, C.; Ruimy, R.; Casagrande-Magne, F.; Levast, M.; Chautemps, N.; et al. Bacillus cereus, a serious cause of nosocomial infections: Epidemiologic and genetic survey. PLoS ONE 2018, 13, e0194346. [Google Scholar] [CrossRef]
- Lotte, R.; Hérissé, A.L.; Berrouane, Y.; Lotte, L.; Casagrande, F.; Landraud, L.; Herbin, S.; Ramarao, N.; Boyer, L.; Ruimy, R.; et al. Virulence analysis of Bacillus cereus isolated after death of preterm neonates, Nice, France. Emerg. Infect. Dis. 2017, 23, 845–848. [Google Scholar] [CrossRef] [Green Version]
- RamaRao, N.; Belotti, L.; Deboscker, S.; Ennahar-Vuillemin, M.; De Launay, J.; Lavigne, T.; Koebel, C.; Escande, B.; Guinebretière, M.H. Two unrelated episodes of Bacillus cereus bacteremia in a neonatal intensive care unit. Am. J. Infect. Control. 2014, 42, 694–695. [Google Scholar] [CrossRef]
- Van Der Zwet, W.C.; Parlevliet, G.A.; Savelkoul, P.H.; Stoof, J.; Kaiser, A.M.; Van Furth, A.M.; Vandenbroucke-Grauls, C.M. Outbreak of Bacillus cereus infections in a neonatal intensive care unit traced to balloons used in manual ventilation. J. Clin. Microbiol. 2000, 38, 4131–4136. [Google Scholar] [CrossRef] [Green Version]
- Fournier, S.; Faraut-Derouin, V.; Casetta, A.; Frange, P.; Doit, C.; Fortineau, N.; Romain, O.; Patkai, J.; de Chillaz, C.; Rigourd, V.; et al. Bactériémies à Bacillus cereus en réanimation néonatale à l’ap-hp en 2016. Bull. Epidémiol. Hebd. BEH 2018, 25, 536–540. [Google Scholar]
- Campbell, J.R.; Hulten, K.; Baker, C.J. Cluster of Bacillus species bacteremia cases in neonates during a hospital construction project. Infect. Control. Hosp. Epidemiol. 2011, 32, 1035–1038. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.P.; Que, T.L.; Lee, W.K.; Wong, S.N. Meningoencephalitis caused by Bacillus cereus in a neonate. Hong Kong Med. J. 2001, 7. [Google Scholar]
- Evreux, F.; Delaporte, B.; Leret, N.; Buffet-Janvresse, C.; Morel, A. A case of fatal neonatal Bacillus cereus meningitis. Arch. Pediatr. 2007, 14, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; George, R.; Durbin, G.; Ewer, A.; Hocking, M.; Morgan, M. An outbreak of Bacillus cereus respiratory tract infections on a neonatal unit due to contaminated ventilator circuits. J. Hosp. Infect. 1999, 41, 19–22. [Google Scholar] [CrossRef]
- Horii, T.; Tamai, K.; Notake, S.; Yanagisawa, H. Bacillus cereus bloodstream infection in a preterm neonate complicated by late meningitis. Case Rep. Infect. Dis. 2012, 2012, 1–3. [Google Scholar]
- Patrick, C.C.; Langston, C.; Baker, C.J. Bacillus Species infections in neonates. Clin. Infect. Dis. 1989, 11, 612–615. [Google Scholar] [CrossRef]
- Sasahara, T.; Hayashi, S.; Morisawa, Y.; Sakihama, T.; Yoshimura, A.; Hirai, Y. Bacillus cereus bacteremia outbreak due to contaminated hospital linens. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 30, 219–226. [Google Scholar] [CrossRef]
- Tokieda, K.; Morikawa, Y.; Maeyama, K.; Mori, K.; Ikeda, K. Clinical manifestations of Bacillus cereus meningitis in newborn infants. J. Paediatr. Child Health 1999, 35, 582–584. [Google Scholar] [CrossRef]
- Hilliard, N.J.; Schelonka, R.L.; Waites, K.B. Bacillus cereus bacteremia in a preterm neonate. J. Clin. Microbiol. 2003, 41, 3441–3444. [Google Scholar] [CrossRef] [Green Version]
- Lewin, A.; Delage, G.; Bernier, F.; Germain, M. Banked human milk and quantitative risk assessment of Bacillus cereus infection in premature infants: A simulation study. Can. J. Infect. Dis. Med Microbiol. 2019, 2019, 6348281. [Google Scholar] [CrossRef] [Green Version]
- Ramarao, N.; Sanchis, V. The pore-forming haemolysins of Bacillus cereus: A review. Toxins 2013, 5, 1119–1139. [Google Scholar] [CrossRef] [Green Version]
- Guinebretiere, M.; Girardin, H.; Dargaignaratz, C.; Carlin, F.; Nguyen, C. Contamination flows of Bacillus cereus and spore-forming aerobic bacteria in a cooked, pasteurized and chilled zucchini purée processing line. Int. J. Food Microbiol. 2003, 82, 223–232. [Google Scholar] [CrossRef]
- Beecher, D.J.; Schoeni, J.L.; Wong, A.C. Enterotoxic activity of hemolysin BL from Bacillus cereus. Infect. Immun. 1995, 63, 4423–4428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beecher, D.J.; Wong, A.C. Identification of hemolysin BL-producing Bacillus cereus isolates by a discontinuous hemolytic pattern in blood agar. Appl. Environ. Microbiol. 1994, 60, 1646–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beecher, D.J.; Pulido, J.S.; Barney, N.P.; Wong, A.C. Extracellular virulence factors in Bacillus cereus endophthalmitis: Methods and implication of involvement of hemolysin BL. Infect. Immun. 1995, 63, 632–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, T.; Granum, P.E. Comparison of biological effect of the two different enterotoxin complexes isolated from three different strains of Bacillus cereus. Microbiology 1997, 143, 3329–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beecher, D.J.; Macmillan, J.D. A novel bicomponent hemolysin from Bacillus cereus. Infect. Immun. 1990, 58, 2220–2227. [Google Scholar] [CrossRef] [Green Version]
- Beecher, D.J.; Wong, A.C.L. Tripartite haemolysin BL: Isolation and characterization of two distinct homologous sets of components from a single Bacillus cereus isolate. Microbiology 2000, 146, 1371–1380. [Google Scholar] [CrossRef] [Green Version]
- Granum, P.E.; O’Sullivan, K.; Lund, T. The sequence of the non-haemolytic enterotoxin operon from Bacillus cereus. FEMS Microbiol. Lett. 1999, 177, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Granum, P.E.; Tomas, J.M.; Alouf, J.E. A survey of bacterial toxins involved in food poisoning: A suggestion for bacterial food poisoning toxin nomenclature. Int. J. Food Microbiol. 1995, 28, 129–144. [Google Scholar] [CrossRef]
- Fagerlund, A.; Lindback, T.; Storset, A.K.; Granum, P.E.; Hardy, S.P. Bacillus cereus Nhe is a pore-forming toxin with structural and functional properties similar to the ClyA (HlyE, SheA) family of haemolysins, able to induce osmotic lysis in epithelia. Microbiology 2008, 154, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Lund, T.; De Buyser, M.-L.; Granum, P.E. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef]
- Fagerlund, A.; Brillard, J.; Fürst, R.; Guinebretière, M.-H.; Granum, P.E. Toxin production in a rare and genetically remote cluster of strains of the Bacillus cereus group. BMC Microbiol. 2007, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Guinebretiere, M.-H.; Monthã¡n, A.; Berge, O.; Fricker, M.; Svensson, B. Toxin gene profiling of enterotoxic and emetic Bacillus cereus. FEMS Microbiol. Lett. 2006, 260, 232–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehling-Schulz, M.; Svensson, B.; Guinebretiere, M.-H.; Lindbäck, T.; Andersson, M.; Schulz, A.; Fricker, M.; Christiansson, A.; Granum, P.E.; Märtlbauer, E.; et al. Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 2005, 151, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, S.L. Bacillus cereus immune escape: A journey within macrophages. FEMS Microbiol. Lett. 2013, 347, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tran, S.L.; Puhar, A.; Ngo-Camus, M.; Ramarao, N. Trypan blue dye enters viable cells incubated with the pore-forming toxin HlyII of Bacillus cereus. PLoS ONE 2011, 6, e22876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, S.L.; Guillemet, E.; Ngo-Camus, M.; Clybouw, C.; Puhar, A.; Moris, A.; Gohar, M.; Lereclus, D.; Ramarao, N. Hemolysin II is a Bacillus cereus virulence factor that induces apoptosis of macrophages. Cell Microbiol. 2011, 13, 92–108. [Google Scholar] [CrossRef]
- Cadot, C.; Tran, S.-L.; Vignaud, M.-L.; de Buyser, M.-L.; Kolstø, A.-B.; Brisabois, A.; Nguyen, C.; Lereclus, D.; Guinebretière, M.-H.; Ramarao, N. InhA1, NprA and HlyII as candidates to differentiate pathogenic from non-pathogenic Bacillus cereus strains. J. Clin. Microbiol. 2010, 3, 1358–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RamaRao, N.; Lereclus, D. The InhA1 metalloprotease allows spores of the B. cereus group to escape macrophages. Cell. Microbiol. 2005, 7, 1357–1364. [Google Scholar]
- Haydar, A.; Tran, S.L.; Guillemet, E.; Darrigo, C.; Perchat, S.; Lereclus, D.; Coquet, L.; Jouenne, T.; Ramarao, N. InhA1-mediated cleavage of the metalloprotease NprA allows Bacillus cereus to escape from macrophages. Front Microbiol. 2018, 23, 1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemet, E.; Leréec, A.; Tran, S.-L.; Royer, C.; Barbosa, I.; Sansonetti, P.; Lereclus, D.; RamaRao, N. The bacterial DNA repair protein Mfd confers resistance to the host nitrogen immune response. Sci. Rep. 2016, 6, 29349. [Google Scholar] [CrossRef] [Green Version]
- Darrigo, C.; Guillemet, E.; Dervyn, R.; Ramarao, N. The bacterial Mfd protein prevents DNA damage induced by the host nitrogen immune response in a NER-independent but RecBC-dependent pathway. PLoS ONE 2016, 11, e0163321. [Google Scholar] [CrossRef]
- Tran, S.-L.; Guillemet, E.; Gohar, M.; Lereclus, D.; RamaRao, N. CwpFM (EntFM) is a Bacillus cereus potential cell wall peptidase implicated in adhesion, biofilm formation, and virulence. J. Bacteriol. 2010, 192, 2638–2642. [Google Scholar] [CrossRef] [Green Version]
- Tran, S.-L.; Cormontagne, D.; Vidic, J.; André-Leroux, G.; RamaRao, N. Structural modeling of cell wall peptidase CwpFM (EntFM) reveals distinct intrinsically disordered extensions specific to pathogenic Bacillus cereus strains. Toxins 2020, 12, 593. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, P.-R.; Teng, L.-J.; Yang, P.-C.; Pan, H.-L.; Ho, S.-W.; Luh, K.-T. Nosocomial pseudoepidemic caused by Bacillus cereus traced to contaminated ethyl alcohol from a liquor factory. J. Clin. Microbiol. 1999, 37, 2280–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Hu, Q.; Xu, F.; Ding, S.; Zhu, K. Characterization of Bacillus cereus in dairy products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef]
- Vidic, J.; Chaix, C.; Manzano, M.; Heyndrickx, M. Food sensing: Detection of Bacillus cereus spores in dairy products. Biosensors 2020, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Egopal, N.; Ehill, C.; Ross, P.R.; Beresford, T.P.; Fenelon, M.A.; Cotter, P.D. The prevalence and control of Bacillus and related spore-forming bacteria in the dairy industry. Front. Microbiol. 2015, 6, 1418. [Google Scholar]
- Machado, S.G.; Baglinière, F.; Marchand, S.; Van Coillie, E.; Vanetti, M.C.D.; De Block, J.; Heyndrickx, M. The biodiversity of the microbiota producing heat-resistant enzymes responsible for spoilage in processed bovine milk and dairy products. Front. Microbiol. 2017, 8, 302. [Google Scholar] [CrossRef] [Green Version]
- Sarita Kumari, P.K.S. Bacillus cereus hazard and control in industrial dairy processing environment. Food Control. 2016, 69, 20–29. [Google Scholar] [CrossRef]
- Saleh-Lakha, S.; Leon-Velarde, C.G.; Chen, S.; Lee, S.; Shannon, K.; Fabri, M.; Downing, G.; Keown, B. A study to assess the numbers and prevalence of Bacillus cereus and its toxins in pasteurized fluid milk. J. Food Prot. 2017, 80, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Brandl, H.; Fricker-Feer, C.; Ziegler, M.; Mandal, J.; Stephan, R.; Lehner, A. Distribution and identification of culturable airborne microorganisms in a Swiss milk processing facility. J. Dairy Sci. 2014, 97, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.-D.; Huertas, J.-P.; Fernández, P.; Palop, A. Effect of the medium characteristics and the heating and cooling rates on the nonisothermal heat resistance of Bacillus sporothermodurans IC4 spores. Food Microbiol. 2013, 34, 158–163. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, Y.; Liu, X.; Xia, X.; Ding, S.; Zhu, K. Evaluation of the toxicity and toxicokinetics of Cereulide from an Emetic Bacillus cereus strain of milk origin. Toxins 2016, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Märtlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Ducrest, P.J.; Pfammatter, S.; Stephan, D.; Vogel, G.; Thibault, P.; Schnyder, B. Rapid detection of Bacillus ionophore cereulide in food products. Sci. Rep. 2019, 9, 5814. [Google Scholar] [CrossRef] [Green Version]
- Carlin, F.; Fricker, M.; Pielaat, A.; Heisterkamp, S.; Shaheen, R.; Salonen, M.S.; Svensson, B.; Nguyen, C.; Ehling-Schulz, M. Emetic toxin-producing strains of Bacillus cereus show distinct characteristics within the Bacillus cereus group. Int. J. Food Microbiol. 2006, 109, 132–138. [Google Scholar] [CrossRef] [Green Version]
- RamaRao, N.; Tran, S.-L.; Marin, M.; Vidic, J. Advanced methods for detection of Bacillus cereus and its pathogenic factors. Sensors 2020, 20, 2667. [Google Scholar] [CrossRef] [PubMed]
- Price, E.; Weaver, G.; Hoffman, P.; Jones, M.; Gilks, J.; O’Brien, V.; Ridgway, G. Decontamination of breast pump milk collection kits and related items at home and in hospital: Guidance from a joint working group of the healthcare infection society and infection prevention society. J. Hosp. Infect. 2016, 92, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigourd, V.; Barnier, J.P.; Ferroni, A.; Nicloux, M.; Hachem, T.; Magny, J.F.; Lapillonne, A.; Frange, P.; Nassif, X.; Bille, E. Recent actuality about Bacillus cereus and human milk bank: A new sensitive method for microbiological analysis of pasteurized milk. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1297–1303. [Google Scholar] [CrossRef]
- Peila, C.; Emmerik, N.E.; Giribaldi, M.; Stahl, B.; Ruitenberg, J.E.; Van Elburg, R.M.; Cavallarin, L. Human milk processing: A systematic review of innovative techniques to ensure the safety and quality of donor milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 353–361. [Google Scholar] [CrossRef]
- Wesolowska, A.; Sinkiewicz-Darol, E.; Barbaska, O.; Bernatowicz-Lojko, U.; Borszewska-Kornacka, M.; van Goudoever, J. Innovative techniques of processing human milk to preserve key components. Nutrients 2019, 11, 1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramarao, N. Bacillus cereus: Caractéristiques et pathogénicité. EMC Biol. Medic. 2020, 7, 1–11. [Google Scholar] [CrossRef]
- Arslanoglu, S.; Moro, G.E.; Ziegler, E.E.; the WAPM Working Group on Nutrition. Optimization of human milk fortification for preterm infants: New concepts and recommendations. J. Périnat. Med. 2010, 38, 233–238. [Google Scholar]
- Moro, G.E.; Arslanoglu, S. Heat treatment of human milk. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 165–166. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref | Sex | Birth Weight (g) | Gestational Age, Weeks + Days | Age at First Positive Culture, Days | Predisposing Factors | Treatment | Outcome | Suspected or Proven Source of Infection | First B. cereus Identification | Toxin Identified |
|---|---|---|---|---|---|---|---|---|---|---|
| [21] | NS | 750 | 30 +2 | 3 | Premature, low weight | NS | Died | Pasteurized milk, packs of diapers, linen, baby bath, benches (suspected) | Blood culture | ND |
| NS | 3000 | 40 | 1 | NS | Survived | Cavum | ND | |||
| NS | 1075 | 29 + 2 | 5 | Premature, low weight | NS | Died | Blood culture | ND | ||
| NS | 2815 | 37 + 2 | 2 | NS | Survived | Blood culture | ND | |||
| NS | 3515 | 38 + 6 | 6 | NS | Survived | Blood culture | ND | |||
| NS | 3240 | 39 | 9 | NS | Survived | Blood culture | ND | |||
| NS | 1380 | 31 | 9 | Premature, low weight | NS | Survived | Blood culture | ND | ||
| NS | 1025 | 29 + 4 | 11 | Premature, low weight | NS | Survived | Blood culture | ND | ||
| NS | 750 | 27 + 5 | 76 | Premature, low weight | NS | Died | Blood culture | ND | ||
| NS | 1720 | 31 | 10 | Premature, low weight | NS | Survived | Blood culture | ND | ||
| [18] | F | 880 | 27 + 2 | 4 | Premature, low weight | Cefotaxime, gentamicin, vancomycin, fluoroquinolone | Died | Incubator, ultrasonographic probe, bench used for bottle feeding (suspected) | Tracheobronchial aspiration | CytK2, Nhe |
| M | 1480 | 29 + 4 | 4 | Premature, low weight | Cefotaxime, gentamicin, vancomycin | Died | Blood culture | |||
| [22] | NS | 1650 | 31 | 70 | Premature, low weight | Vancomycin | Survived | Bioaerosol and surface contamination. Work stations, storage room, linens (suspected) | Blood culture | ND |
| NS | 1148 | 29 | 58 | Premature, low weight | Vancomycin | Survived | Blood culture | ND | ||
| NS | 1515 | 28 | 23 | Premature, low weight | Vancomycin | Survived | Blood culture | ND | ||
| NS | 710 | 24 | 14 | Premature, low weight | Vancomycin | Survived | Blood culture | ND | ||
| NS | 945 | 25 | 59 | Premature, low weight | Vancomycin | Survived | Blood culture | ND | ||
| [23] | M | 1580 | 26 | 30 | Premature, low weight | Ampicillin, cefotaxime then amikacin and vancomycin | Died | ND | Blood culture | ND |
| [5] | NS | 960 | 29 | 3 | Premature, low weight | Vancomycin, cefotaxime and metronidazole | Survived | Food-related origin: milk (suspected) | Gastric fluid culture | ND |
| NS | 1500 | 30 | 3 | Premature, low weight | Vancomycin, cefotaxime and metronidazole | Survived | Gastric fluid culture | ND | ||
| [24] | F | 1670 | 31 | 28 | Premature, low weight | Cefotaxime, amoxicillin, metronidazole, amikacin | Died | ND | Blood culture and central catheter | ND |
| [25] | NS | 880 | 27 | 1 | Premature, low weight | Vancomycin | Survived | Water from washing machine chamber (proven) | ET aspirates | ND |
| NS | 880 | 28 | 1 | Premature, low weight | Vancomycin | Died | ET aspirates | ND | ||
| NS | 720 | 27 | 3 | Premature, low weight | ND | Survived | ET aspirates | ND | ||
| NS | 880 | 29 | 3 | Premature, low weight | ND | Survived | ET aspirates | ND | ||
| NS | 640 | 28 | 2 | Premature, low weight | Vancomycin | Died | Vial of pulmonary surfactant used for both of them (but no growth) | ET aspirates | ND | |
| NS | 530 | 26 | 3 | Premature, low weight | Vancomycin | Died | ET aspirates | ND | ||
| [26] | F | 800 | NS | 1 | Premature, low weight | Vancomycin, meropenem and for meningitis: linezolid, meropenem and clindamycin | Survived | Ventilator equipment, intravascular catheters and linen (suspected) | Blood culture | ND |
| [27] | F | 830 | 26 | 8 | Premature, low weight | Vancomycin, amikacin | Died | ND | Blood culture | ND |
| [19] | NS | 650 | 24 + 5 | 32 | Premature, low weight | Cefotaxime, vancomycin and amikacin | Survived | Arterial catheter (suspected) | Blood culture | Nhe |
| NS | 615 | 26 + 5 | 5 | Premature, low weight | NS | Died | Catheter (suspected) | Peripheral catheter and central catheter | Nhe | |
| [28] | M | NS | NS | 1 | NS | Cefozopran | Died | Hospital linens (proven) | ND | |
| F | NS | NS | 19 | NS | Ampicillin, meropenem, vancomycin, panipenem | Died | ND | |||
| [29] | F | 3764 | 37 | 6 | Premature | Ampicillin, gentamicin | Died | Peripheral vein catheter (suspected) | CSF and blood culture | ND |
| F | 1506 | 36 | 9 | Premature, low weight | Ampicillin, cefotaxime | Died | Nasal feeding tube (suspected) | CSF culture | ND | |
| [20] | M | 895 | 28 + 5 | 5 | Premature, low weight | Amoxicillin and cefotaxime | Died | Balloons of manual ventilation, person to person transmission (health care workers hands) (suspected) | Blood culture, CSF, trachea aspirate, necrotic Skin lesion at insertion site of arterial catheter | ND |
| F | 1000 | 26 + 4 | 5 | Premature, low weight | Meropenem, vancomycin | Survived | Blood, aspirate from left knee | ND | ||
| M | 2780 | 37 + 3 | 14 | Premature | Meropenem, vancomycin | Survived | Blood, CSF, tip of peripheral catheter | ND | ||
| [30] | M | 585 | 24 | 19 | Premature, low weight | Vancomycin, tobramycin | Survived | ND | Blood culture | ND |
| [31] | NS | 590 | 25 | 10 | Premature, low weight | Linezolid, meropenem, vancomycin | Died | Batches of HBM but the strains from the baby are different (suspected) | Blood culture | ND |
| NS | 560 | 24 | 6 | Premature, low weight | Ampicillin, tobramycin, vancomycin, meropenem, piperacillin-tazobactam, fluconazole, amphotericin and trimethoprim-sulfamethoxazole | Died | HBM (suspected) | Blood culture | ND | |
| [32] | NS | NS | NS | NS | Premature | Died | ND | Blood culture and cerebrospinal fluid | Nhe | |
| NS | NS | NS | NS | Premature | Vancomycin, cefotaxime | Survived | ND | Blood culture | Nhe, Hbl | |
| NS | NS | NS | NS | Premature | Vancomycin | Survived | ND | Blood culture | Nhe | |
| NS | NS | NS | NS | Vancomycin | Survived | ND | Neonatal gastric liquid | Nhe | ||
| NS | NS | NS | NS | Cefotaxime, amoxicillin, amikacin | Survived | ND | Umbilical | Nhe | ||
| NS | NS | NS | NS | Ceftriaxone | Survived | ND | Axilla later feces | Nhe | ||
| NS | NS | NS | NS | Premature | Cefotaxime, amoxicillin, amikacin | Survived | ND | Stomach tube feeding | Nhe | |
| NS | NS | NS | NS | Premature | Vancomycin | Survived | ND | Gastric acid | Nhe | |
| NS | NS | NS | NS | Premature | Amoxicillin, amikacin, vancomycin | Survived | ND | Central venous catheter | Nhe | |
| NS | NS | NS | NS | Premature | Vancomycin, cefotaxime, amikacin | Survived | ND | Blood culture | Nhe | |
| NS | NS | NS | NS | Premature | Died | ND | Blood culture | Nhe | ||
| NS | NS | NS | NS | Premature | Vancomycin, cefotaxime, metronidazole | Survived | ND | Stomach tube feeding | ND | |
| NS | NS | NS | NS | Premature | Vancomycin, cefotaxime, metronidazole | Survived | ND | Stomach tube feeding | ND | |
| NS | NS | NS | NS | Ceftriaxone, gentamicin | Survived | ND | Blood culture | ND | ||
| NS | NS | NS | NS | Premature | Vancomycin | Died | ND | Blood culture from umbilical venous catheter and peripheral veins | Nhe, Hbl | |
| NS | NS | NS | NS | Premature | Vancomycin | Died | ND | Bronchial aspiration (lung) | Nhe |
| Term and Birth Weight | Hours or Days After Birth | Bacteriological Analyses | Symptoms | Evolution |
|---|---|---|---|---|
| 25 WA 675 g | D14 | Blood (+) UVC(+) LS (+) MB (-) | Diffuse multiple abscess 2 hemispheres | Death |
| 39 WA + 1 2500 g | D22 | Blood (+) OMM (-) HL | Respiratory breaks | Favorable |
| 25 WA + 1 665 g | D134 | Blood (+) OMM(-) | Respiratory breaks | Favorable |
| 25 WA 600 g | D15 | Blood (+) OMM (-) | No symptom | Favorable |
| 38 WA 2700 g | D32 | Blood (+) HL | No symptom | Favorable |
| 26 WA 675 g | D26 | Blood (+) MB (-) | No symptom | Favorable |
| ND | Blood (+) Parenteral | No symptom | Favorable | |
| 28 WA + 5 585 g | D13 | Blood (+) Incubator (+) | No symptom | Favorable |
| 32 WA 1530 g | D3 | Blood (+) MB (-) | No symptom | Favorable |
| ND | Blood (+) MB (-) | Favorable | ||
| 29 WA + 5 1240 g | D20 | Blood (+) | No symptom | Favorable |
| 31 WA 1410 g | D4 | Blood (+) MB (-) | No symptom | Favorable |
| 25 WA 635 g | D6 | Blood (+) MB (-) | Respiratory breaks | Favorable |
| 29 WA 1260 g | D18 | Blood (+) | Respiratory breaks | Favorable |
| 31 WA + 2 1940 g | D16 | Blood (+) MB(-) | No symptom | Favorable |
| 25 WA + 6 855 g | D6 | Blood (+) Peritonitis MB (-) | Increase needs in 02 | Favorable |
| 37 WA + 2 2035 g | D16 | Blood (+) | No symptom | Favorable |
| 28 WA + 2 865 g | D16 | Blood (+) MB (-) | Favorable | |
| 4 premature babies | Blood (+) MB (-) | One with a brain abscess | 1 Death/4 | |
| 1 premature baby | MB (-) | Favorable | ||
| 1 premature baby | Blood (+) LS (+) MB (-) | Death | ||
| 29 WA 1210 g | D21 | Electric ramp (+) Blood (+ Bacillus pumilus) MB (-) | Bone localization | Favorable |
| 29 WA + 5 1285 g | D6 | Blood (+) MB (-) | Extensive periventricular hemorrhage and cytotoxic Involvement of the cortex and central gray nuclei | Death |
| 1 premature baby | Blood (+) MB (-) | Favorable | ||
| 31 WA + 2 1430 g | D29 | Blood (+) MB (+ 103 Bc) | Favorable | |
| Blood (+) | Favorable | |||
| Near born | Blood (+) Incubator (+) | |||
| 25 WA | D19 | 2 Blood (+) Bacillus subtilis MB (-) | Favorable | |
| ND | Tracheal suction (+) MB (-) | |||
| ND | MB (-) | |||
| 1 premature baby | Blood (+) MB(-) | |||
| 1 premature baby | Blood (+) MB (-) | |||
| 29 WA 550 g | D3 | Blood (+) Parenteral (+) | Brain abscess | Death |
| 30 WA 740 g | H50 | Blood (+) | Brain abscess | Death |
| 36 WA 2000 g | D1 | Nasopharynx (+) | Favorable |
| Year | Bacteriologic Contamination Liters (%) |
|---|---|
| 2000 | 192 (3.3%) |
| 2001 | 174 (3.4%) |
| 2002 | 172 (3.2%) |
| 2003 | 155 (2.5%) |
| 2004 | 256 (4%) |
| 2005 | 215 (3.3%) |
| 2006 | 249 (3.4%) |
| 2007 | 293 (4.2%) |
| 2008 * | 524 (8.1%) |
| 2009 | 401 (6.4%) |
| 2010 | 465 (7.2%) |
| 2011 | 541 (8.1%) |
| 2012 ** | 628 (11.3%) |
| 2013 | 515 (8.1%) |
| 2014 *** | 1221 (14.5%) |
| 2015 | 815 (9.6%) |
| 2016 **** | 1097 (13.9%) |
| 2017 ■ | 1851(19.45%) |
| 2018 | 2018 (19%) |
| 2019 | 2463 (21.2%) |
| Date | Culture Post Pasteurization | Culture Post Pasteurization/Fast-Freezing |
|---|---|---|
| 21-Feb | Bacillus cereus | Negative |
| 16-Apr | Bacillus cereus | Bacillus cereus |
| 28-May | Bacillus cereus | Negative |
| 11-Apr | Bacillus cereus | Streptococcus thermophilus |
| 29-May | Bacillus cereus | Bacillus cereus |
| 05-Apr | Bacillus cereus | Bacillus cereus |
| 13-May | Bacillus cereus | Negative |
| 29-Mar | Bacillus cereus | Negative |
| 10-Apr | Bacillus cereus | Negative |
| 05-Apr | Bacillus cereus | Negative |
| 07-Mar | Bacillus cereus | Bacillus cereus |
| 04-Mar | Bacillus cereus | Negative |
| 12-Mar | Bacillus cereus | Bacillus cereus |
| 21-Mar | Bacillus cereus | Bacillus cereus |
| 18-Mar | Bacillus cereus | Bacillus cereus |
| 16-Apr | Bacillus cereus | Negative |
| 03-Jun | Bacillus cereus | Bacillus cereus |
| 28-Feb | Bacillus cereus | Enterococcus faecalis |
| 20-Feb | Bacillus cereus | Negative |
| 14-May | Bacillus cereus | Bacillus cereus |
| 22-May | Bacillus cereus | Negative |
| 27-Feb | Bacillus cereus | Bacillus cereus |
| 27-Feb | Bacillus cereus | Bacillus cereus |
| 04-Feb | Bacillus cereus | Negative |
| 08-Feb | Bacillus cereus | Bacillus cereus |
| 05-Feb | Bacillus cereus | Bacillus cereus |
| 15-Feb | Bacillus cereus | Bacillus cereus |
| 27-Feb | Bacillus cereus | Bacillus cereus |
| 06-May | Bacillus cereus | Bacillus cereus |
| 12-Feb | Bacillus cereus | Bacillus cereus |
| 27-Feb | Bacillus cereus | Bacillus spp. |
| 07-Feb | Bacillus cereus | Bacillus cereus |
| 12-Feb | Bacillus cereus | Bacillus cereus |
| 01-Mar | Bacillus cereus | Bacillus cereus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cormontagne, D.; Rigourd, V.; Vidic, J.; Rizzotto, F.; Bille, E.; Ramarao, N. Bacillus cereus Induces Severe Infections in Preterm Neonates: Implication at the Hospital and Human Milk Bank Level. Toxins 2021, 13, 123. https://doi.org/10.3390/toxins13020123
Cormontagne D, Rigourd V, Vidic J, Rizzotto F, Bille E, Ramarao N. Bacillus cereus Induces Severe Infections in Preterm Neonates: Implication at the Hospital and Human Milk Bank Level. Toxins. 2021; 13(2):123. https://doi.org/10.3390/toxins13020123
Chicago/Turabian StyleCormontagne, Delphine, Virginie Rigourd, Jasmina Vidic, Francesco Rizzotto, Emmanuelle Bille, and Nalini Ramarao. 2021. "Bacillus cereus Induces Severe Infections in Preterm Neonates: Implication at the Hospital and Human Milk Bank Level" Toxins 13, no. 2: 123. https://doi.org/10.3390/toxins13020123
APA StyleCormontagne, D., Rigourd, V., Vidic, J., Rizzotto, F., Bille, E., & Ramarao, N. (2021). Bacillus cereus Induces Severe Infections in Preterm Neonates: Implication at the Hospital and Human Milk Bank Level. Toxins, 13(2), 123. https://doi.org/10.3390/toxins13020123