Primary Impacts of the Fungal Toxin Sporidesmin on HepG2 Cells: Altered Cell Adhesion without Oxidative Stress or Cell Death


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
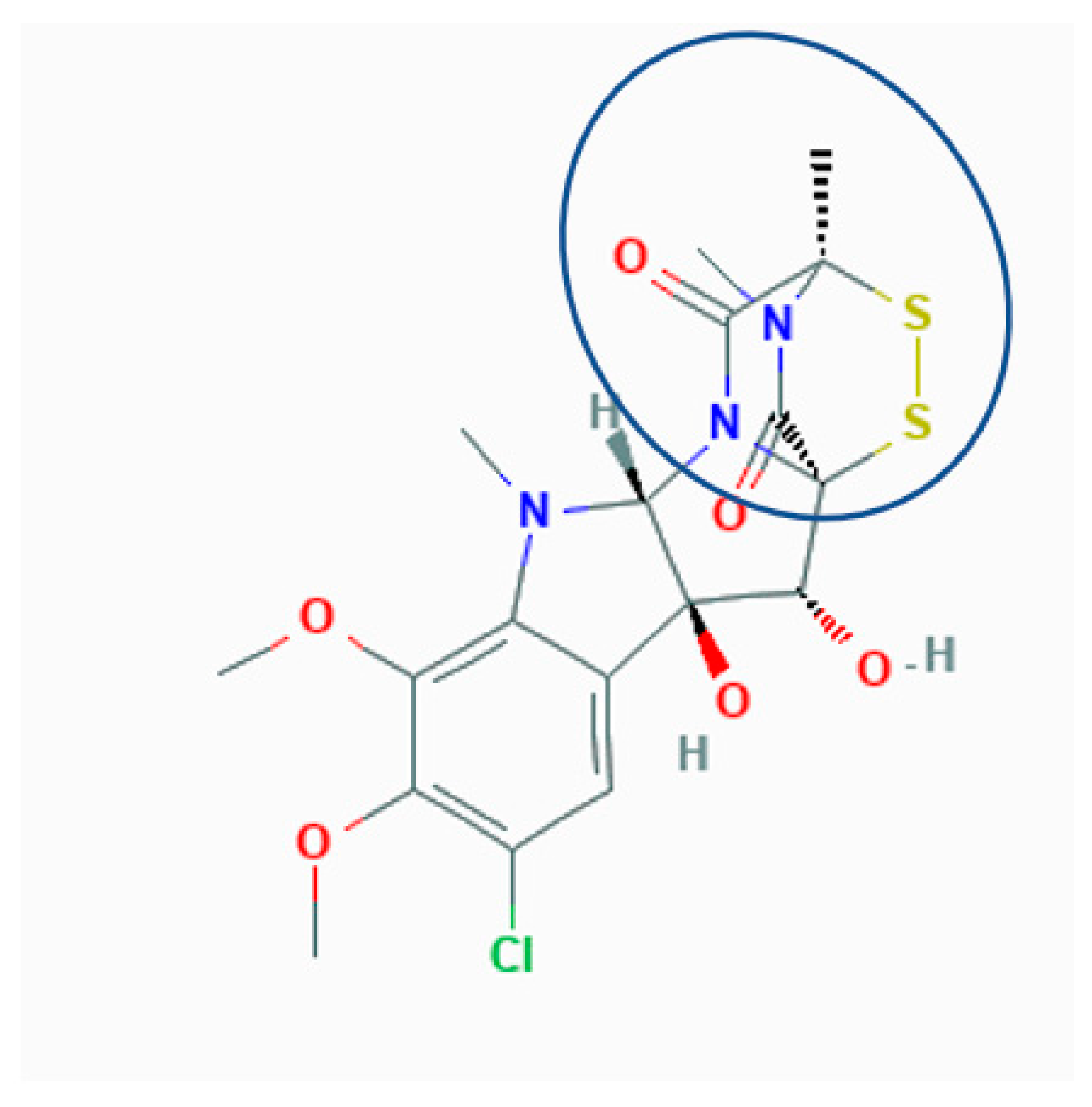
:1. Introduction
2. Results
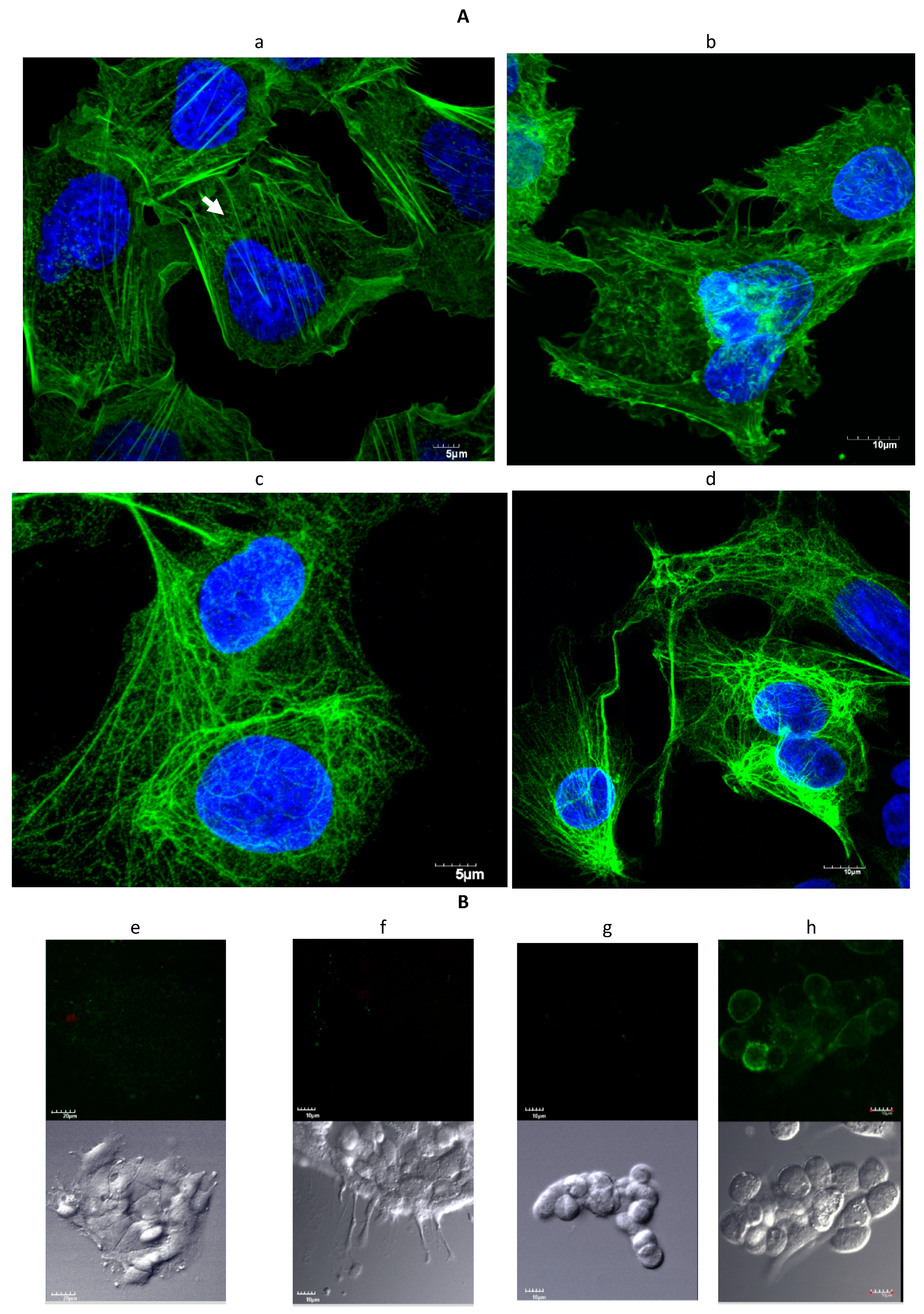
2.1. Microscopy
2.2. Staining with Annexin-V-FLUOS
2.3. Oxidative Stress
3. Discussion
3.1. Oxidative Stress
3.2. Cytotoxicity and Cell Death
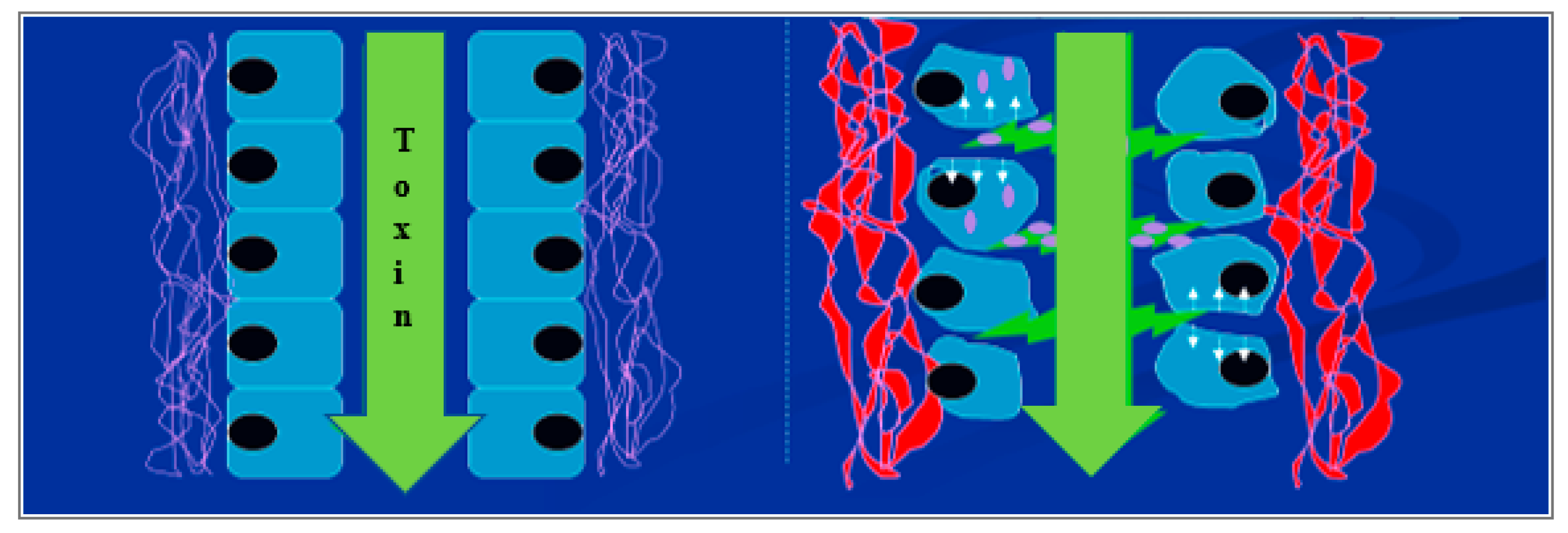
3.3. Cell Adhesion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture
5.2. Microscopy
5.3. Apoptosis
5.4. Oxidative Stress and Cytotoxicity
5.5. Two-Dimensional Gel Electrophoresis of Proteins
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- di Menna, M.E.; Smith, B.L.; Miles, C.O. A history of facial eczema (pithomycotoxicosis) research. N. Z. J. Agric. Res. 2009, 52, 345–376. [Google Scholar] [CrossRef]
- Mortimer, P.H. The experimental intoxication of sheep with sporidesmin, a metabolic product of Pithomyces chartarum. IV—Histological and histochemical examinations of orally-dosed sheep. Res. Vet. Sci. 1963, 4, 166–185. [Google Scholar] [CrossRef]
- Smith, B. Effects of low dose rates of sporidesmin given orally to sheep. N. Z. Vet. J. 2000, 48, 176–181. [Google Scholar] [CrossRef]
- Welch, T.R.; Williams, R.M. Epidithiodioxopiperazines. occurrence, synthesis and biogenesis. Nat. Prod. Rep. 2014, 31, 1376–1404. [Google Scholar] [CrossRef] [Green Version]
- Chai, C.L.; Waring, P. Redox sensitive epidithiodioxopiperazines in biological mechanisms of toxicity. Redox. Rep. 2000, 5, 257–264. [Google Scholar] [CrossRef]
- Jordan, T.W. The cellular and molecular toxicity of sporidesmin. N. Z. Vet. J. 2020, 68, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Munday, R. Studies on the mechanism of toxicity of the mycotoxin, sporidesmin. I. Generation of superoxide radical by sporidesmin. Chem. Biol. Interact. 1982, 41, 361–374. [Google Scholar] [CrossRef]
- Srinivasan, U.; Bala, A.; Jao, S.C.; Starke, D.W.; Jordan, T.W.; Mieyal, J.J. Selective inactivation of glutaredoxin by sporidesmin and other epidithiopiperazinediones. Biochemistry 2006, 45, 8978–8987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortimer, P.H.; Collins, B.S. The in vitro toxicity of the sporidesmins and related compounds to tissue-culture cells. Res. Vet. Sci. 1968, 9, 136–142. [Google Scholar] [CrossRef]
- Jordan, T.W.; Pedersen, J.S. Sporidesmin and gliotoxin induce cell detachment and perturb microfilament structure in cultured liver cells. J. Cell Sci. 1986, 85, 33–46. [Google Scholar] [PubMed]
- Waring, P.; Egan, M.; Braithwaite, A.; Mullbacher, A.; Sjaarda, A. Apoptosis induced in macrophages and T blasts by the mycotoxin sporidesmin and protection by Zn2+ salts. Int. J. Immunopharmacol. 1990, 12, 445–457. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Waring, P.; Howlett, B.J. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution, mode of action, functions and biosynthesis. Microbiology 2005, 151, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.H.; Wong, S.W.; Martinez, J. Programmed necrosis and disease: We interrupt your regular programming to bring you necroinflammation. Cell Death Differ. 2019, 26, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Briggs, L.R.; Towers, N.R.; Molan, P.C. Development of an enzyme-linked immunosorbent assay for analysis of sporidesmin A and its metabolites in ovine urine and bile. J. Agric. Food Chem. 1994, 42, 2769–2777. [Google Scholar] [CrossRef]
- Lindsay, G.C.; Morris, C.A.; Boucher, M.; Capundan, K.; Jordan, T.W. Effects of sporidesmin on cultured biliary tract cells from Romney lambs that differed in their sensitivity to sporidesmin. N. Z. Vet. J. 2018, 66, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Truman, P.; Boucher, M.; Keyzers, R.A.; Northcote, P.; Jordan, T.W. The algal metabolite yessotoxin affects heterogeneous nuclear ribonucleoproteins in HepG2 cells. Proteomics 2009, 9, 2529–2542. [Google Scholar] [CrossRef]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munday, R. Studies on the mechanism of toxicity of the mycotoxin, sporidesmin. V. Generation of hydroxyl radical by sporidesmin. J. App. Toxicol. 1987, 7, 17–22. [Google Scholar] [CrossRef]
- Rabilloud, T.; Heller, M.; Gasnier, F.; Luche, S.; Rey, C.; Aebersold, R.; Benahmed, M.; Louisot, P.; Lunardi, J. Proteomics analysis of cellular response to oxidative stress. Evidence for in vivo overoxidation of peroxiredoxins at their active site. J. Biol. Chem. 2002, 277, 19396–19401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Reest, J.; Lilla, S.; Zheng, L.; Zanivan, S.; Gottlieb, E. Proteome-wide analysis of cysteine oxidation reveals metabolic sensitivity to oxidative stress. Nat. Commun. 2018, 9, 1581. [Google Scholar] [CrossRef]
- Cesaratto, L.; Vascotto, C.; D’Ambrosio, C.; Scaloni, A.; Baccarani, U.; Paron, I.; Damante, G.; Calligaris, S.; Quadrifoglio, F.; Tiribelli, C.; et al. Overoxidation of peroxiredoxins as an immediate and sensitive marker of oxidative stress in HepG2 cells and its application to the redox effects induced by ischemia/reperfusion in human liver. Free Radic. Res. 2005, 39, 255–268. [Google Scholar] [CrossRef]
- Rabilloud, T.; Lescuyer, P. Proteomics in mechanistic toxicology: History, concepts, achievements, caveats, and potential. Proteomics 2015, 15, 1051–1074. [Google Scholar] [CrossRef] [Green Version]
- Wilmes, A.; Chan, A.; Rawson, P.; Jordan, T.W.; Miller, J.H. Paclitaxel effects on the proteome of HL-60 promyelocytic leukemic cells: Comparison to peloruside A. Investig. New Drugs 2012, 30, 121–129. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim Biophys Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative stress in cell culture: An under-appreciated problem? FEBS Lett. 2003, 540, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lou, Q.; He, L.; Wen, C.; Lin, M.; Zhu, Z.; Wang, F.; Huang, L.; Lan, W.; Iwamoto, A.; et al. Reduced gliotoxin induces ROS-mediated anoikis in human colorectal cancer cells. Int. J. Oncol. 2018, 52, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Haun, F.; Neumann, S.; Peintner, L.; Wieland, K.; Habicht, J.; Schwan, C.; Østevold, K.; Koczorowska, M.M.; Biniossek, M.; Kist, M.; et al. Identification of a novel anoikis signalling pathway using the fungal virulence factor gliotoxin. Nat. Commun. 2018, 9, 3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachir, A.I.; Horwitz, A.R.; Nelson, W.J.; Bianchini, J.M. Actin-based adhesion modules mediate cell interactions with the extracellular matrix and neighboring cells. Cold Spring Harb. Perspect. Biol. 2017, 9, a023234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Liu, X.; Chen, F.; Hu, Y.; Li, Z.; Liu, Y.; Han, X.; Sun, Y.; Han, L. Gliotoxin destructs the pulmonary epithelium barrier function by reducing cofilin oligomer formation to promote the dissolution of actin stress fibers. Microb. Pathog. 2018, 123, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, F.; Liu, X.; Han, X.; Hu, Y.; Su, X.; Chen, Y.; Sun, Y.; Han, L. Gliotoxin induces cofilin phosphorylation to promote actin cytoskeletal dynamics and internalization of Aspergillus fumigatus into Type II human pneumocyte cells. Front. Microbiol. 2019, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Bhathal, P.S.; Jordan, T.W.; Mackay, I.R. Mouse strain differences in susceptibility to sporidesmin-induced biliary tract injury. Liver 1990, 10, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.K.; Karlsen, T.H.; Folseraas, T. Cholangiocytes in the pathogenesis of primary sclerosing cholangitis and development of cholangiocarcinoma. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1390–1400. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.M.; Pinto, C.; Maroni, L.; Benedetti, A.; Marzioni, M. Inflammation and the gut-liver axis in the pathophysiology of cholangiopathies. Int. J. Mol. Sci. 2018, 19, 3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lechuga, S.; Ivanov, A.I. Actin cytoskeleton dynamics during mucosal inflammation: A view from broken epithelial barriers. Curr. Opin. Physiol. 2021, 19, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Roca Suarez, A.A.; El Saghire, H.; Saviano, A.; Schuster, C.; Lupberger, J.; Baumert, T.F. Tight junction proteins and the biology of hepatobiliary disease. Int. J. Mol. Sci. 2020, 21, 825. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boucher, M.; Jordan, T.W. Primary Impacts of the Fungal Toxin Sporidesmin on HepG2 Cells: Altered Cell Adhesion without Oxidative Stress or Cell Death. Toxins 2021, 13, 179. https://doi.org/10.3390/toxins13030179
Boucher M, Jordan TW. Primary Impacts of the Fungal Toxin Sporidesmin on HepG2 Cells: Altered Cell Adhesion without Oxidative Stress or Cell Death. Toxins. 2021; 13(3):179. https://doi.org/10.3390/toxins13030179
Chicago/Turabian StyleBoucher, Magalie, and T. William Jordan. 2021. "Primary Impacts of the Fungal Toxin Sporidesmin on HepG2 Cells: Altered Cell Adhesion without Oxidative Stress or Cell Death" Toxins 13, no. 3: 179. https://doi.org/10.3390/toxins13030179
APA StyleBoucher, M., & Jordan, T. W. (2021). Primary Impacts of the Fungal Toxin Sporidesmin on HepG2 Cells: Altered Cell Adhesion without Oxidative Stress or Cell Death. Toxins, 13(3), 179. https://doi.org/10.3390/toxins13030179