Aflatoxin in Dairy Cows: Toxicity, Occurrence in Feedstuffs and Milk and Dietary Mitigation Strategies



Abstract
:1. Introduction
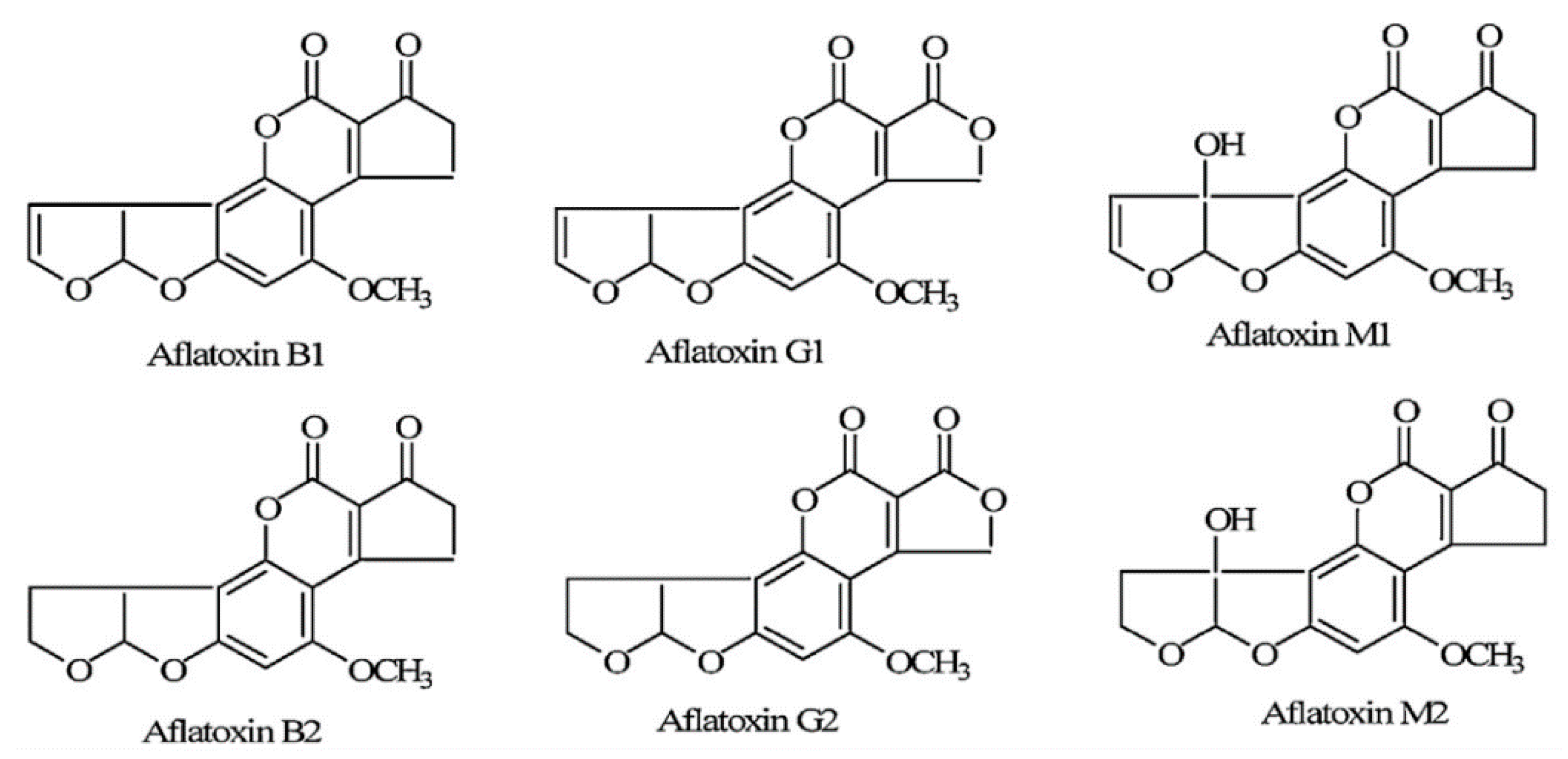
2. Aflatoxin Types
3. Occurrence and Prevalence
4. Risk Factors for Contamination
4.1. Temperature Stress
4.2. Drought Stress
4.3. Diseases, Insects and Other Physical Damage
5. Effects on Ruminant Animals
5.1. Effects on Performance and Health
5.2. Effects on Ruminal Fermentation and Microorganisms
5.3. Effects on Reproduction
6. Strategies for Preventing and Mitigating Aflatoxin Contamination
6.1. Preharvest Prevention
6.2. Post-Harvest Mitigation Treatments
6.2.1. Ammoniation
6.2.2. Ozonation
6.2.3. Enzyme Treatment
6.2.4. Cold Plasma
6.2.5. Clay Sequestering Agents
- Inclusion Level
- Sequestering Agent to Aflatoxin Ratio
- Particle Size
- Mode of Addition
- Saccharomyces cerevisiae Based Sequestering Agents
6.2.6. Lactic Acid Bacteria
6.2.7. Chlorophyll Products and Polyphenol
- Chlorophyll Products
- Polyphenol
6.2.8. Activated Carbon
6.2.9. Antioxidants
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hell, K.; Cardwell, K.; Setamou, M.; Poehling, H.-M. The influence of storage practices on aflatoxin contamination in maize in four agroecological zones of Benin, west Africa. J. Stored Prod. Res. 2000, 36, 365–382. [Google Scholar] [CrossRef]
- Yin, Y.-N.; Yan, L.-Y.; Jiang, J.-H.; Ma, Z.-H. Biological control of aflatoxin contamination of crops. J. Zhejiang Univ. Sci. B 2008, 9, 787–792. [Google Scholar] [CrossRef]
- Kosicki, R.; Błajet-Kosicka, A.; Grajewski, J.; Twarużek, M. Multiannual mycotoxin survey in feed materials and feedingstuffs. Anim. Feed. Sci. Technol. 2016, 215, 165–180. [Google Scholar] [CrossRef]
- Rahimi, E.; Bonyadian, M.; Rafei, M.; Kazemeini, H. Occurrence of aflatoxin M1 in raw milk of five dairy species in Ahvaz, Iran. Food Chem. Toxicol. 2010, 48, 129–131. [Google Scholar] [CrossRef]
- Queiroz, O.; Han, J.; Staples, C.; Adesogan, A. Effect of adding a mycotoxin-sequestering agent on milk aflatoxin M1 concentration and the performance and immune response of dairy cattle fed an aflatoxin B1-contaminated diet. J. Dairy Sci. 2012, 95, 5901–5908. [Google Scholar] [CrossRef]
- Mitchell, N.J.; Bowers, E.; Hurburgh, C.; Wu, F. Potential economic losses to the US corn industry from aflatoxin con-tamination. Food Addit. Contam. Part A 2016, 33, 540–550. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration (FDA). Action Levels for Aflatoxins in Animal Food, Compliance Policy Guide. 2019. Available online: https://www.fda.gov/media/121202/download (accessed on 16 July 2019).
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006. Setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- European Commission. Commission Directive 2003/100/EC of 31 October 2003 amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council on undesirable substances in animal feed (Text with EEA relevance). Off. J. Euro. Union. 2003, 285, 33–37. [Google Scholar]
- Wouters, A.T.B.; Casagrande, R.A.; Wouters, F.; Watanabe, T.T.N.; Boabaid, F.M.; Cruz, C.E.F.; Driemeier, D. An outbreak of aflatoxin poisoning in dogs associated with aflatoxin B1–contaminated maize products. J. Veter. Diagn. Investig. 2013, 25, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin Contamination of Commercial Maize Products during an Outbreak of Acute Aflatoxicosis in Eastern and Central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef]
- Kamala, A.; Shirima, C.; Jani, B.; Bakari, M.; Sillo, H.; Rusibamayila, N.; De Saeger, S.; Kimanya, M.; Gong, Y.; Simba, A.; et al. Outbreak of an acute aflatoxicosis in Tanzania during 2016. World Mycotoxin J. 2018, 11, 311–320. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outbreak of Aflatoxin Poisoning—Eastern and Central Provinces, Kenya, January–July 2004. Morb. Mortal. Wkly. Rep. 2004, 53, 790–793. Available online: http://www.cdc.gov/mmwr/preview/mmwrhtml/mm5334a4.htm (accessed on 20 April 2019).
- Gong, Y.Y.; Egal, S.; Hounsa, A.; Turner, P.C.; Hall, A.J.; Cardwell, K.F.; Wild, C.P. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: The critical role of weaning. Int. J. Epidemiol. 2003, 32, 556–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, P.C.; Moore, S.E.; Hall, A.J.; Prentice, A.M.; Wild, C.P. Modification of immune function through exposure to dietary aflatoxin in Gambian children. Environ. Health Perspect. 2003, 111, 217–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Veter. J. 2008, 176, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Hasheminya, S.M.; Dehghannya, J. Strategies for decreasing aflatoxin in livestock feed and milk. Int. Res. J. Appl. Basic Sci. 2013, 4, 1506–1510. [Google Scholar]
- Komsky-Elbaz, A.; Saktsier, M.; Roth, Z. Aflatoxin B1 impairs sperm quality and fertilization competence. Toxicology 2018, 393, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Hansen, P.J.; Xiao, Y.; Amaral, T.F.; Vyas, D.; Adesogan, A.T. Aflatoxin compromises development of the preim-plantation bovine embryo through mechanisms independent of reactive oxygen production. J. Dairy Sci. 2019, 102, 10506–10513. [Google Scholar] [CrossRef]
- Dors, G.C.; Caldas, S.S.; Feddern, V.; Bemvenuti, R.H.; dos Santos Hackbart, H.C.; de Souza, M.M.; dos Santos, O.M.; Garda-Buffon, J.; Primel, E.G.; Badiale-Furlong, E. Aflatoxins: Contamination, analysis and control. In Aflatoxins-Biochemistry and Molecular Biology; IntechOpen: Shanghai China, 2011. [Google Scholar]
- Zhang, D.; Li, P.; Zhang, Q.; Zhang, W. Ultrasensitive nanogold probe-based immunochromatographic assay for simultaneous detection of total aflatoxins in peanuts. Biosens. Bioelectron. 2011, 26, 2877–2882. [Google Scholar] [CrossRef]
- Horn, B.; Singh, K.; Frisvad, J.C.; Thrane, U.; Mathur, S.B. An Illustrated Manual on Identification of Some Seed-Borne Aspergilli, Fusaria, and Penicillia and Their Mycotoxins. Mycologia 1993, 85, 139. [Google Scholar] [CrossRef]
- Sheng, Y.J.; Eremin, S.; Mi, T.J.; Zhang, S.X.; Shen, J.Z.; Wang, Z.H. The development of a fluorescence polarization im-munoassay for aflatoxin detection. Biomed. Environ. Sci. 2014, 27, 126–129. [Google Scholar] [PubMed]
- International Agency for Research on Cancer (IARC). Some traditional herbal medicines, some mycotoxins, naphthalene and styrene: Summary of data reported and evaluation. In IARC Monographs on the Evaluation of the Carcinogenic Risk to Humans; IARC: Lyon, France, 2002; Volume 82, p. 36. [Google Scholar]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-Mycotoxin Screening Reveals the Occurrence of 139 Different Secondary Metabolites in Feed and Feed Ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [Green Version]
- Cathey, C.; Huang, Z.; Sarr, A.; Clement, B.; Phillips, T. Development and Evaluation of a Minicolumn Assay for the Detection of Aflatoxin M1 in Milk. J. Dairy Sci. 1994, 77, 1223–1231. [Google Scholar] [CrossRef]
- Agag, B.I. Mycotoxins in foods and feeds: 1-aflatoxins. Assiut Univ. Bull. Environ. Res. 2004, 7, 173–205. [Google Scholar]
- Dhanasekaran, D.; Shanmugapriya, S.; Thajuddin, N.; Panneerselvam, A. Aflatoxins and Aflatoxicosis in Human and Animals. In Aflatoxins—Biochemistry and Molecular Biology; InTech Press: Rijeka, Croatia, 2011; pp. 221–254. [Google Scholar]
- Rodrigues, I.; Naehrer, K. Prevalence of mycotoxins in feedstuffs and feed surveyed worldwide in 2009 and 2010. Phytopathol. Mediterr. 2012, 51, 175–192. [Google Scholar]
- Senerwa, D.M.; Sirma, A.J.; Mtimet, N.; Kang’ethe, E.K.; Grace, D.; Lindahl, J.F. Prevalence of aflatoxin in feeds and cow milk from five counties in Kenya. Afr. J. Food Agric. Nutr. Dev. 2016, 16, 11004–11021. [Google Scholar] [CrossRef]
- Nishimwe, K.; Bowers, E.; Ayabagabo, J.D.D.; Habimana, R.; Mutiga, S.; Maier, D. Assessment of Aflatoxin and Fumonisin Contamination and Associated Risk Factors in Feed and Feed Ingredients in Rwanda. Toxins 2019, 11, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang’Ethe, E.K.; Sirma, A.J.; Murithi, G.; Mburugu-Mosoti, C.K.; Ouko, E.O.; Korhonen, H.J.; Nduhiu, G.J.; Mungatu, J.K.; Joutsjoki, V.; Lindfors, E.; et al. Occurrence of mycotoxins in food, feed, and milk in two counties from different agro-ecological zones and with historical outbreak of aflatoxins and fumonisins poisonings in Kenya. Food Qual. Saf. 2017, 1, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, F.M.; Girgin, G.; Baydar, T. Mycotoxin detection in maize, commercial feed, and raw dairy milk samples from Assiut City, Egypt. Vet. Sci. 2019, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Nyangi, C.; Mugula, J.K.; Beed, F.; Boni, S.; Koyano, E.; Sulyok, M. Aflatoxins and fumonisin contamination of marketed maize, maize bran and maize used as animal feed in Northern Tanzania. Afr. J. Food Agric. Nutr. Dev. 2016, 16, 11054–11065. [Google Scholar] [CrossRef]
- Biomin Mycotoxin Survey 2018. Available online: https://engage.biomin.net/hubfs/BIOMIN/Mycotoxin%20Survey%20Report/MAG_MTX-Survey-Report_2018_EN.pdf (accessed on 10 May 2019).
- Biomin Mycotoxin Survey 2019. Available online: https://cdn2.hubspot.net/hubfs/5480243/BIOMIN/Downloads/MAG_MTXSurveyReport_2019_EN.pdf (accessed on 11 October 2019).
- Turna, N.S.; Wu, F. Aflatoxin M1 in milk: A global occurrence, intake, & exposure assessment. Trends Food Sci. Technol. 2021, 110, 183–192. [Google Scholar] [CrossRef]
- Jones, R.; Duncan, H.; Hamilton, P. Planting date, harvest date, and irrigation effects on infection and aflatoxin production by Aspergillus flavus in field corn. Phytopathology 1981, 71, 810–816. [Google Scholar] [CrossRef]
- Diener, U.L.; Cole, R.J.; Sanders, T.H.; Payne, G.A.; Lee, L.S.; Klich, M.A. Epidemiology of Aflatoxin Formation by Aspergillus Flavus*. Annu. Rev. Phytopathol. 1987, 25, 249–270. [Google Scholar] [CrossRef]
- Klich, M.A. Aspergillus flavus: The major producer of aflatoxin. Mol. Plant Pathol. 2007, 8, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Cotty, P.J.; Jaime-Garcia, R. Influences of climate on aflatoxin producing fungi and aflatoxin contamination. Int. J. Food Microbiol. 2007, 119, 109–115. [Google Scholar] [CrossRef]
- Kos, J.; Mastilović, J.; Hajnal, E.J.; Šarić, B. Natural occurrence of aflatoxins in maize harvested in Serbia during 2009–2012. Food Control 2013, 34, 31–34. [Google Scholar] [CrossRef]
- Schindler, A.F.; Palmer, J.G.; Eisenberg, W.V. Aflatoxin production by Aspergillus flavus as related to various tempera-tures. Appl. Microbiol. 1967, 15, 1006–1009. [Google Scholar] [CrossRef]
- OBrian, G.R.; Georgianna, D.R.; Wilkinson, J.R.; Yu, J.; Abbas, H.K.; Bhatnagar, D.; Cleveland, T.E.; Nierman, W.; Payne, G.A. The effect of elevated temperature on gene transcription and aflatoxin biosynthesis. Mycologia 2007, 99, 232–239. [Google Scholar] [CrossRef]
- Liu, X.; Guan, X.; Xing, F.; Lv, C.; Dai, X.; Liu, Y. Effect of water activity and temperature on the growth of Aspergillus flavus, the expression of aflatoxin biosynthetic genes and aflatoxin production in shelled peanuts. Food Control 2017, 82, 325–332. [Google Scholar] [CrossRef]
- Payne, G.A.; Cassel, D.K.; Adkins, C.R. Reduction of aflatoxin contamination in corn by irrigation and till-age. Phytopathology 1986, 76, 679–684. [Google Scholar] [CrossRef]
- Hell, K.; Cardwell, K.F.; Setamou, M.; Schulthess, F. Influence of insect infestation on aflatoxin contamination of stored maize in four agroecological regions in Benin. Afr. Entomol. 2000, 8, 169–177. [Google Scholar]
- Ogunade, I.; Arriola, K.; Jiang, Y.; Driver, J.; Staples, C.; Adesogan, A. Effects of 3 sequestering agents on milk aflatoxin M1 concentration and the performance and immune status of dairy cows fed diets artificially contaminated with aflatoxin B1. J. Dairy Sci. 2016, 99, 6263–6273. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.B.; Kubena, L.F.; Phillips, T.D.; Corrier, D.E.; Elissalde, M.H.; Huff, W.E. Diminution of aflatoxin toxicity to growing lambs by dietary supplementation with hydrated sodium calcium aluminosilicate. Am. J. Veter. Res. 1991, 52, 152–156. [Google Scholar]
- Edrington, T.S.; Harvey, R.B.; Kubena, L.F. Effect of aflatoxin in growing lambs fed ruminally degradable or escape protein sources. J. Anim. Sci. 1994, 72, 1274–1281. [Google Scholar] [CrossRef]
- Sulzberger, S.; Melnichenko, S.; Cardoso, F. Effects of clay after an aflatoxin challenge on aflatoxin clearance, milk production, and metabolism of Holstein cows. J. Dairy Sci. 2017, 100, 1856–1869. [Google Scholar] [CrossRef]
- Rodrigues, R.; Rodrigues, R.O.; Ledoux, D.; Rottinghaus, G.; Borutova, R.; Averkieva, O.; McFadden, T. Feed additives containing sequestrant clay minerals and inactivated yeast reduce aflatoxin excretion in milk of dairy cows. J. Dairy Sci. 2019, 102, 6614–6623. [Google Scholar] [CrossRef] [Green Version]
- Applebaum, R.S.; Brackett, R.E.; Wiseman, D.W.; Marth, E.H. Responses of Dairy Cows to Dietary Aflatoxin: Feed Intake and Yield, Toxin Content, and Quality of Milk of Cows Treated with Pure and Impure Aflatoxin. J. Dairy Sci. 1982, 65, 1503–1508. [Google Scholar] [CrossRef]
- Gallo, A.; Moschini, M.; Masoero, F. Aflatoxins absorption in the gastro-intestinal tract and in the vaginal mucosa in lactating dairy cows. Ital. J. Anim. Sci. 2008, 7, 53–63. [Google Scholar] [CrossRef]
- Fernández, A.; Hernández, M.; Verde, M.T.; Sanz, M. Effect of aflatoxin on performance, hematology, and clinical immunology in lambs. Can. J. Veter. Res. 2000, 64, 53–58. [Google Scholar]
- Butler, W.H.; Neal, G.E. Mode of action and human health aspects of aflatoxin carcinogenesis. Pure Appl. Chem. 1977, 49, 1747–1751. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Park, M.A.; Ha, J.K. Mycotoxins and Their Biotransformation in the Rumen: A Review. Asian Austr. J. Anim. Sci. 2010, 23, 1250–1260. [Google Scholar] [CrossRef]
- Mertens, D.R. Biological effects of mycotoxins upon rumen fermentation and lactating dairy cows. In Interactions of Mycotoxins in Animal Production; National Academy Press: Washington, DC, USA, 1979; pp. 118–136. [Google Scholar]
- Sinha, R.R.P.; Arora, S.P. Influence of different levels of aflatoxins on rumen fermentation (in vitro). J. Nucl. Agric. Biol. 1982, 11, 87–89. [Google Scholar]
- Jiang, Y.; Yang, H.; Lund, P. Effect of aflatoxin B1 on in vitro ruminal fermentation of rations high in alfalfa hay or ryegrass hay. Anim. Feed Sci. Technol. 2012, 175, 85–89. [Google Scholar] [CrossRef]
- Westlake, K.; Mackie, R.; Dutton, M. In vitro metabolism of mycotoxins by bacterial, protozoal and ovine ruminal fluid preparations. Anim. Feed Sci. Technol. 1989, 25, 169–178. [Google Scholar] [CrossRef]
- Jiang, Y.; Ogunade, I.; Arriola, K.; Pech-Cervantes, A.; Kim, D.; Li, X.; Xue, Y.; Vyas, D.; Adesogan, A. Short communication: Effects of a physiologically relevant concentration of aflatoxin B1 with or without sequestering agents on in vitro rumen fermentation of a dairy cow diet. J. Dairy Sci. 2020, 103, 1559–1565. [Google Scholar] [CrossRef]
- Jiang, Y.; Ogunade, I.M.; Kim, D.H.; Li, X.; Pech-Cervantes, A.A.; Arriola, K.G.; Oliveira, A.S.; Driver, J.P.; Ferraretto, L.F.; Staples, C.R.; et al. Effect of adding clay with or without a Saccharomyces cerevisiae fermentation product on the health and performance of lactating dairy cows challenged with dietary aflatoxin B1. J. Dairy Sci. 2018, 101, 3008–3020. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ogunade, I.; Pech-Cervantes, A.; Fan, P.; Li, X.; Kim, D.; Arriola, K.; Poindexter, M.; Jeong, K.; Vyas, D.; et al. Effect of sequestering agents based on a Saccharomyces cerevisiae fermentation product and clay on the ruminal bacterial community of lactating dairy cows challenged with dietary aflatoxin B1. J. Dairy Sci. 2020, 103, 1431–1447. [Google Scholar] [CrossRef]
- Komsky-Elbaz, A.; Kalo, D.; Roth, Z. Effect of aflatoxin B1 on bovine spermatozoa’s proteome and embryo’s transcriptome. Reproduction 2020, 160, 709–723. [Google Scholar] [CrossRef]
- Agarwal, A.; Allamaneni, S.S.R. Role of free radicals in female reproductive diseases and assisted reproduction. Reprod. Biomed. Online 2004, 9, 338–347. [Google Scholar] [CrossRef]
- Huang, S.; Zheng, N.; Fan, C.; Cheng, M.; Wang, S.; Jabar, A.; Wang, J.; Cheng, J. Effects of aflatoxin B1 combined with ochratoxin A and/or zearalenone on metabolism, immune function, and antioxidant status in lactating dairy goats. Asian Austr. J. Anim. Sci. 2018, 31, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.-M.; Shi, C.-Y.; Shen, Y.; Ong, C.-N. Detection of elevated reactive oxygen species level in cultured rat hepatocytes treated with aflatoxin B1. Free Radic. Biol. Med. 1996, 21, 139–146. [Google Scholar] [CrossRef]
- Dorner, J.W.; Cole, R.J.; Blankenship, P.D. Effect of inoculum rate of biological control agents on preharvest aflatoxin con-tamination of peanuts. Biol. Control 1998, 12, 171–176. [Google Scholar] [CrossRef]
- Ehrlich, K.C.; Cotty, P.J. An isolate of Aspergillus flavus used to reduce aflatoxin contamination in cottonseed has a defective polyketide synthase gene. Appl. Microbiol. Biotechnol. 2004, 65, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.F.; Ameye, M.; De Saeger, S.; Audenaert, K.; Haesaert, G. Biological Control of Mycotoxigenic Fungi and Their Toxins: An Update for the Pre-Harvest Approach. In Mycotoxins—Impact and Management Strategies; IntechOpen: London, UK, 2019. [Google Scholar]
- Marechera, G.; Ndwiga, J. Estimation of the potential adoption of Aflasafe among smallholder maize farmers in lower eastern Kenya. Afr. J. Agri. Res. Econ. 2015, 10, 311-2016-5622. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Atehnkeng, J.; Ortega-Beltran, A.; Akande, A.; Falade, T.; Cotty, P.J. Ground-truthing” efficacy of bio-logical control for aflatoxin mitigation in farmers’ fields in Nigeria: From field trials to commercial usage, a 10-year study. Front. Microbiol. 2019, 10, 2528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agbetiameh, D.; Ortega-Beltran, A.; Awuah, R.T.; Atehnkeng, J.; Elzein, A.; Cotty, P.J.; Bandyopadhyay, R. Field efficacy of two atoxigenic biocontrol products for mitigation of aflatoxin contamination in maize and groundnut in Ghana. Biol. Control. 2020, 150, 104351. [Google Scholar] [CrossRef]
- Dövényi-Nagy, T.; Rácz, C.; Molnár, K.; Bakó, K.; Szláma, Z.; Jóźwiak, Á.; Farkas, Z.; Pócsi, I.; Dobos, A.C. Pre-Harvest Modelling and Mitigation of Aflatoxins in Maize in a Changing Climatic Environment—A Review. Toxins 2020, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.; Barros, G.; Palacios, S.; Chulze, S.; Battilani, P. Review on pre- and post-harvest management of peanuts to minimize aflatoxin contamination. Food Res. Int. 2014, 62, 11–19. [Google Scholar] [CrossRef]
- Piva, G.; Galvano, F.; Pietri, A.; Piva, A. Detoxification methods of aflatoxins. A review. Nutr. Res. 1995, 15, 767–776. [Google Scholar] [CrossRef]
- Gardner Jr, H.K.; Koltun, S.P.; Dollear, F.G.; Rayner, E.T. Inactivation of aflatoxins in peanut and cottonseed meals by ammoniation. J. Am. Oil Chem. Soc. 1971, 48, 70–73. [Google Scholar] [CrossRef]
- Samarajeewa, U.; Sen, A.C.; Cohen, M.D.; Wei, C.I. Detoxification of Aflatoxins in Foods and Feeds by Physical and Chemical Methods1. J. Food Prot. 1990, 53, 489–501. [Google Scholar] [CrossRef]
- Fremy, J.M.; Gautier, J.P.; Herry, M.P.; Terrier, C.; and Calett, C. Effects of ammoniation on the “carry-over” of aflatoxins into bovine milk. Food Addit. Contam. 1988, 5, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Cullen, P.; Tiwari, B.; O’Donnell, C.; Muthukumarappan, K. Modelling approaches to ozone processing of liquid foods. Trends Food Sci. Technol. 2009, 20, 125–136. [Google Scholar] [CrossRef]
- De Alencar, E.R.; Faroni, L.R.D.A.; Soares, N.D.F.F.; da Silva, W.A.; da Silva Carvalho, M.C. Efficacy of ozone as a fun-gicidal and detoxifying agent of aflatoxins in peanuts. J. Sci. Food Agric. 2012, 92, 899–905. [Google Scholar] [CrossRef]
- Freitas-Silva, O.; Venâncio, A. Ozone applications to prevent and degrade mycotoxins: A review. Drug Metab. Rev. 2010, 42, 612–620. [Google Scholar] [CrossRef]
- Chen, R.; Ma, F.; Li, P.-W.; Zhang, W.; Ding, X.-X.; Zhang, Q.; Li, M.; Wang, Y.-R.; Xu, B.-C. Effect of ozone on aflatoxins detoxification and nutritional quality of peanuts. Food Chem. 2014, 146, 284–288. [Google Scholar] [CrossRef]
- Luo, X.; Wang, R.; Wang, L.; Li, Y.; Bian, Y.; Chen, Z. Effect of ozone treatment on aflatoxin B1 and safety evaluation of ozo-nized corn. Food Control. 2014, 37, 171–176. [Google Scholar] [CrossRef]
- Motomura, M.; Toyomasu, T.; Mizuno, K.; Shinozawa, T. Purification and characterization of an aflatoxin degradation enzyme fromPleurotus ostreatus. Microbiol. Res. 2003, 158, 237–242. [Google Scholar] [CrossRef]
- Das, A.; Bhattacharya, S.; Palaniswamy, M.; Angayarkanni, J. Biodegradation of aflatoxin B1 in contaminated rice straw by Pleurotus ostreatus MTCC 142 and Pleurotus ostreatus GHBBF10 in the presence of metal salts and surfactants. World J. Microbiol. Biotechnol. 2014, 30, 2315–2324. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mulè, G. Mycotoxin Biotransformation by Native and Commercial Enzymes: Present and Future Perspectives. Toxins 2017, 9, 111. [Google Scholar] [CrossRef]
- Alberts, J.; Gelderblom, W.; Botha, A.; Van Zyl, W. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef]
- Novozymes A/S. Detoxification of Aflatoxin in Feed Products. U.S. Patent Application No. 12,398,284, 5 March 2009.
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.C.; Quintieri, L.; Cimmarusti, M.T.; Monaci, L.; Haidukowski, M.; Logrieco, A.F.; Sanjust, E.; et al. Aflatoxin B1 and M1 Degradation by Lac2 from Pleurotus pulmonarius and Redox Mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef] [Green Version]
- Das, C.; Mishra, H.N. Mishra. In vitro degradation of aflatoxin B1 by horseradish peroxidase. Food Chem. 2000, 68, 309–313. [Google Scholar] [CrossRef]
- Liu, D.-L.; Yao, D.-S.; Liang, Y.-Q.; Zhou, T.-H.; Song, Y.-P.; Zhao, L.; Ma, L. Production, purification, and characterization of an intracellular aflatoxin-detoxifizyme from Armillariella tabescens (E-20). Food Chem. Toxicol. 2001, 39, 461–466. [Google Scholar] [CrossRef]
- Adamović, M.; Grubić, G.; Milenković, I.; Jovanović, R.; Protić, R.; Sretenović, L.; Stoićević, L. The biodegradation of wheat straw by Pleurotus ostreatus mushrooms and its use in cattle feeding. Anim. Feed Sci. Technol. 1998, 71, 357–362. [Google Scholar] [CrossRef]
- Beg, S.; Zafar, S.I.; Shah, F. Rice husk biodegradation by Pleurotus ostreatus to produce a ruminant feed. Agric. Wastes 1986, 17, 15–21. [Google Scholar] [CrossRef]
- Siciliano, I.; Spadaro, D.; Prelle, A.; Vallauri, D.; Cavallero, M.C.; Garibaldi, A.; Gullino, M.L. Use of Cold Atmospheric Plasma to Detoxify Hazelnuts from Aflatoxins. Toxins 2016, 8, 125. [Google Scholar] [CrossRef]
- Shi, H.; Ileleji, K.; Stroshine, R.L.; Keener, K.; Jensen, J.L. Reduction of Aflatoxin in Corn by High Voltage Atmospheric Cold Plasma. Food Bioprocess Technol. 2017, 10, 1042–1052. [Google Scholar] [CrossRef]
- Shi, H.; Cooper, B.; Stroshine, R.L.; Ileleji, K.E.; Keener, K.M. Structures of Degradation Products and Degradation Pathways of Aflatoxin B1 by High-Voltage Atmospheric Cold Plasma (HVACP) Treatment. J. Agric. Food Chem. 2017, 65, 6222–6230. [Google Scholar] [CrossRef]
- Hojnik, N.; Cvelbar, U.; Tavčar-Kalcher, G.; Walsh, J.; Križaj, I. Mycotoxin decontamination of food, Cold atmospheric pressure plasma versus “classic” decontamination. Toxins 2017, 9, 151. [Google Scholar] [CrossRef]
- Pankaj, S.; Shi, H.; Keener, K.M. A review of novel physical and chemical decontamination technologies for aflatoxin in food. Trends Food Sci. Technol. 2018, 71, 73–83. [Google Scholar] [CrossRef]
- Kutz, R.E.; Sampson, J.D.; Pompeu, L.B.; Ledoux, D.R.; Spain, J.N.; Vazquez-Anon, M.; Rottinghaus, G.E. Efficacy of Solis, NovasilPlus, and MTB-100 to reduce aflatoxin M1 levels in milk of early to midlactation dairy cows fed aflatoxin B1. J. Dairy Sci. 2009, 92, 3959–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, C.; Thomas, A.; Elmore, S.; Romoser, A.; Harvey, R.; Ramirez-Ramirez, H.; Phillips, T. Effects of calcium montmorillonite clay and aflatoxin exposure on dry matter intake, milk production, and milk composition. J. Dairy Sci. 2016, 99, 1039–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, D.E.; Hagler, W.M., Jr.; Blackwelder, J.T.; Eve, J.A.; Hopkins, B.A.; Anderson, K.L.; Jones, F.T.; Whitlow, L.W. Aflatoxin Binders II: Reduction of aflatoxin M1 in milk by sequestering agents of cows consuming aflatoxin in feed. Mycopathologia 2004, 157, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Wang, Y.; Nennich, T.; Li, Y.; Liu, J. Transfer of dietary aflatoxin B1 to milk aflatoxin M1 and effect of inclusion of adsorbent in the diet of dairy cows. J. Dairy Sci. 2015, 98, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- Pietri, A.; Bertuzzi, T.; Piva, G.; Binder, E.M.; Schatzmayr, D.; Rodrigues, I. Aflatoxin transfer from naturally contaminated feed to milk of dairy cows and the efficacy of a mycotoxin deactivating product. Int. J. Dairy Sci. 2009, 4, 34–42. [Google Scholar] [CrossRef]
- Pate, R.; Compart, D.P.; Cardoso, F. Aluminosilicate clay improves production responses and reduces inflammation during an aflatoxin challenge in lactating Holstein cows. J. Dairy Sci. 2018, 101, 11421–11434. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.B.; Phillips, T.D.; Ellis, J.A.; Kubena, L.F.; Huff, W.E.; Petersen, H.D. Effects on aflatoxin M1 residues in milk by addition of hydrated sodium calcium aluminosilicate to aflatoxin-contaminated diets of dairy cows. Am. J. Veter. Res. 1991, 52, 1556–1559. [Google Scholar]
- Masoero, F.; Gallo, A.; Diaz, D.; Piva, G.; Moschini, M. Effects of the procedure of inclusion of a sequestering agent in the total mixed ration on proportional aflatoxin M1 excretion into milk of lactating dairy cows. Anim. Feed. Sci. Technol. 2009, 150, 34–45. [Google Scholar] [CrossRef]
- Sumantri, I.; Murti, T.; Van Der Poel, A.; Boehm, J.; Agus, A. Carry-over of aflatoxin B1-feed into aflatoxin M1-milk in dairy cows treated with natural sources of aflatoxin and bentonite. J. Indones. Trop. Anim. Agric. 2012, 37, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Kissell, L.; Davidson, S.; Hopkins, B.A.; Smith, G.W.; Whitlow, L.W. Effect of experimental feed additives on aflatoxin in milk of dairy cows fed aflatoxin-contaminated diets. J. Anim. Physiol. Anim. Nutr. 2012, 97, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Katsoulos, P.D.; Karatzia, M.A.; Boscos, C.; Wolf, P.; Karatzias, H. In-field evaluation of clinoptilolite feeding efficacy on the reduction of milk aflatoxin M1 concentration in dairy cattle. J. Anim. Sci. Technol. 2016, 58, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, B.; Rosim, R.; Cappato, L.; Cruz, A.; Oliveira, C.; Corassin, C. Effects of different sources of Saccharomyces cerevisiae biomass on milk production, composition, and aflatoxin M1 excretion in milk from dairy cows fed aflatoxin B1. J. Dairy Sci. 2017, 100, 5701–5708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, C.R.; Haney, S.; Wang, M.; Ward, S.H.; Rude, B.J.; Bailey, R.H.; Harvey, R.B.; Phillips, T.D. Calcium montmorillonite clay for the reduction of aflatoxin residues in milk and dairy products. J. Dairy Vet. Sci. 2017, 2, 1–8. [Google Scholar]
- Weatherly, M.; Pate, R.; Rottinghaus, G.; Filho, F.R.; Cardoso, F. Physiological responses to a yeast and clay-based adsorbent during an aflatoxin challenge in Holstein cows. Anim. Feed. Sci. Technol. 2018, 235, 147–157. [Google Scholar] [CrossRef]
- Intanoo, M.; Kongkeitkajorn, M.B.; Pattarajinda, V.; Bernard, J.K.; Callaway, T.R.; Suriyasathaporn, W.; Phasuk, Y. Isolation and screening of aflatoxin-detoxifying yeast and bacteria from ruminal fluids to reduce aflatoxin B1 contamination in dairy cattle feed. J. Appl. Microbiol. 2018, 125, 1603–1613. [Google Scholar] [CrossRef]
- Pfliegler, W.P.; Pusztahelyi, T.; Pócsi, I. Mycotoxins—Prevention and decontamination by yeasts. J. Basic Microbiol. 2015, 55, 805–818. [Google Scholar] [CrossRef]
- Jensen, G.S.; Carter, S.G.; Reeves, S.G.; Robinson, L.E.; Benson, K.F. Anti-inflammatory properties of a dried fermentate in vitro and in vivo. J. Med. Food. 2015, 18, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Alugongo, G.; Chung, R.; Dong, S.; Li, S.; Yoon, I.; Wu, Z.; Cao, Z. Effects of Saccharomyces cerevisiae fermentation products on dairy calves: Ruminal fermentation, gastrointestinal morphology, and microbial community. J. Dairy Sci. 2016, 99, 5401–5412. [Google Scholar] [CrossRef] [Green Version]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Firmin, S.; Morgavi, D.; Yiannikouris, A.; Boudra, H. Effectiveness of modified yeast cell wall extracts to reduce aflatoxin B1 absorption in dairy ewes. J. Dairy Sci. 2011, 94, 5611–5619. [Google Scholar] [CrossRef] [PubMed]
- Pierides, M.; El-Nezami, H.; Peltonen, K.; Salminen, S.; Ahokas, J. Ability of Dairy Strains of Lactic Acid Bacteria to Bind Aflatoxin M1 in a Food Model. J. Food Prot. 2000, 63, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Peltonen, K.; El-Nezami, H.; Haskard, C.; Ahokas, J.; Salminen, S. Aflatoxin B1 Binding by Dairy Strains of Lactic Acid Bacteria and Bifidobacteria. J. Dairy Sci. 2001, 84, 2152–2156. [Google Scholar] [CrossRef]
- Ma, Z.X.; Amaro, F.; Romero, J.; Pereira, O.; Jeong, K.; Adesogan, A. The capacity of silage inoculant bacteria to bind aflatoxin B1 in vitro and in artificially contaminated corn silage. J. Dairy Sci. 2017, 100, 7198–7210. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.G.; Muck, R.E.; Weimer, P.J.; Chen, Y.; Gamburg, M. Lactic Acid Bacteria Used in Inoculants for Silage as Probiotics for Ruminants. Appl. Biochem. Biotechnol. 2004, 118, 001–010. [Google Scholar] [CrossRef]
- Oliveira, A.S.; Weinberg, Z.G.; Ogunade, I.M.; Cervantes, A.A.; Arriola, K.G.; Jiang, Y.; Kim, D.; Li, X.; Gonçalves, M.C.; Vyas, D.; et al. Meta-analysis of effects of inoculation with homofermentative and facultative heterofermentative lactic acid bacteria on silage fermentation, aerobic stability, and the performance of dairy cows. J. Dairy Sci. 2017, 100, 4587–4603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonich, M.T.; Egner, P.A.; Roebuck, B.D.; Orner, G.A.; Jubert, C.; Pereira, C.; Groopman, J.D.; Kensler, T.W.; Dashwood, R.H.; Williams, D.E.; et al. Natural chlorophyll inhibits aflatoxin B1-induced multi-organ carcinogenesis in the rat. Carcinogenesis 2007, 28, 1294–1302. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-Y.; Chen, Y.-H.; Chao, P.-Y.; Chen, C.-M.; Hsieh, L.-L.; Hu, S.-P. Naturally occurring chlorophyll derivatives inhibit aflatoxin B1-DNA adduct formation in hepatoma cells. Mutat. Res. Toxicol. Environ. Mutagen. 2008, 657, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Egner, P.A.; Wang, J.-B.; Zhu, Y.-R.; Zhang, B.-C.; Wu, Y.; Zhang, Q.-N.; Qian, G.-S.; Kuang, S.-Y.; Gange, S.J.; Jacobson, L.P.; et al. Chlorophyllin intervention reduces aflatoxin-DNA adducts in individuals at high risk for liver cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 14601–14606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Liu, F.; Zhu, Q.; Zhang, M.; Li, T.; Chen, J.; Huang, Y.; Wang, X.; Sheng, J. Aflatoxin B1 can be complexed with oxidised tea polyphenols and the absorption of the complexed aflatoxin B1 is inhibited in rats. J. Sci. Food Agric. 2016, 97, 1910–1915. [Google Scholar] [CrossRef] [PubMed]
- Marsh, H.; Reinoso, F.R. Activated Carbon; Elsevier: Amsterdam, The Netherlands, 2006; pp. 1–9. [Google Scholar]
- Galvano, F.; Pietri, A.; Fallico, B.; Bertuzzi, T.; Scirè, S.; Galvano, M.; Maggiore, R. Activated Carbons: In Vitro Affinity for Aflatoxin B1 and Relation of Adsorption Ability to Physicochemical Parameters. J. Food Prot. 1996, 59, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Galvano, F.; Pietri, A.; Bertuzzi, T.; Fusconi, G.; Galvano, M.; Piva, A.; Piva, G. Reduction of carry-over of aflatoxin from cow feed to milk by addition of activated carbons. J. Food Prot. 1996, 59, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Chopra, R. Influence of sodium bentonite and activated charcoal on aflatoxin M1 excretion in milk of goats. Small Rumin. Res. 2001, 41, 203–213. [Google Scholar] [CrossRef]
- Verma, R.J.; Nair, A. Ameliorative effect of vitamin E on aflatoxin-induced lipid peroxidation in the testis of mice. Asian J. Androl. 2001, 3, 217–321. [Google Scholar] [PubMed]
- Alpsoy, L.; Yildirim, A.; Agar, G. The antioxidant effects of vitamin A, C, and E on aflatoxin B1-induced oxidative stress in human lymphocytes. Toxicol. Ind. Health 2009, 25, 121–127. [Google Scholar] [CrossRef]


{kind=link}
| Europe | Middle East | Africa | Asia | North America | South and Central America | ||
|---|---|---|---|---|---|---|---|
| Finished Feed | No. of samples | 1146 | 87 | 161 | 1458 | 537 | 1033 |
| % of samples positive for aflatoxin | 12% | 11% | 11% | 44% | 12% | 25% | |
| Average of positive samples, µg/kg | 5 | 3 | 9 | 26 | 9 | 9 | |
| Median of positive, µg/kg | 2 | 2 | 4 | 8 | 4 | 4 | |
| Maximum, µg/kg | 136 | 15 | 26 | 697 | 57 | 216 | |
| Corn | No. of samples | 371 | 14 | 191 | 685 | 478 | 3656 |
| % of samples positive for aflatoxin | 18% | 29% | 3% | 37% | 9% | 19% | |
| Average of positive samples, µg/kg | 9 | 2 | 3 | 42 | 36 | 11 | |
| Median of positive, µg/kg | 2 | 1 | 2 | 8 | 15 | 4 | |
| Maximum, µg/kg | 76 | 6 | 8 | 636 | 280 | 402 | |
| Cereals 1 | No. of samples | 743 | 8 | 28 | 267 | 48 | 586 |
| % of samples positive for aflatoxin | 11% | 13% | 21% | 13% | 6% | 53% | |
| Average of positive samples, µg/kg | 2 | 2 | 8 | 13 | 6 | 5 | |
| Median of positive, µg/kg | 1 | 2 | 4 | 1 | 7 | 4 | |
| Maximum, µg/kg | 19 | 2 | 27 | 88 | 8 | 40 |
| Europe | Middle East and North Africa | Africa | Asia | North America | South and Central America | ||
|---|---|---|---|---|---|---|---|
| Finished Feed | No. of samples | 1042 | 84 | 326 | 1589 | 690 | 1530 |
| % of samples positive for aflatoxin | 8% | 2% | 25% | 30% | 6% | 28% | |
| Average of positive samples, µg/kg | 10 | 42 | 33 | 19 | 11 | 5 | |
| Median of positive, µg/kg | 4 | 12 | 13 | 8 | 5 | 4 | |
| Maximum, µg/kg | 237 | 615 | 370 | 430 | 94 | 134 | |
| Corn | No. of samples | 427 | 30 | 376 | 717 | 524 | 4091 |
| % of samples positive for aflatoxin | 9% | 37% | 7% | 31% | 4% | 21% | |
| Average of positive samples, µg/kg | 8 | 2 | 16 | 43 | 132 | 10 | |
| Median of positive, µg/kg | 4 | 1 | 6 | 10 | 5 | 4 | |
| Maximum, µg/kg | 54 | 5 | 64 | 773 | 1327 | 1264 | |
| Cereals 1 | No. of samples | 766 | 3 | 20 | 90 | 64 | 375 |
| % of samples positive for aflatoxin | 21% | 0% | 5% | 12% | 3% | 54% | |
| Average of positive samples, µg/kg | 2 | - | 1 | 13 | 5 | 4 | |
| Median of positive, µg/kg | 2 | - | 1 | 4 | 5 | 2 | |
| Maximum, µg/kg | 6 | 0 | 1 | 68 | 7 | 30 |
| Name | Company | Compounds | Reference |
|---|---|---|---|
| Astra-Ben 20 | Prince AgriProducts, Quincy, IL | Sodium bentonite | Diaz et al., 2004 [103] |
| Flow Guard | Laporte Biochem, Inc., Milwaukee, WI | Sodium bentonite | Diaz et al., 2004 [103] |
| Mycosorb | American Colloid Co., Arlington Heights, IL | Sodium bentonite | Diaz et al., 2004 [103] |
| Red Crown bentonite | Prince AgriProducts, Quincy, IL | Bentonite | Diaz et al., 2004 [103] |
| SA-20 | Westvaco, Covington, VA | Activated Carbon | Diaz et al., 2004 [103] |
| Calibrin A | Amlan International, Chicago, IL | Calcium montmorillonite bentonite | Queiroz et al., 2012 [5] |
| MTB-100 | Alltech Inc., Nicholasville, KY | Esterified glucomannan with HSCAS | Kutz et al., 2009 [101] |
| NovasilPlus | BSAF, Ludwigshafen, Germany | Smectite clay | Kutz et al., 2009 [101] |
| Solis | Novus International, Saint Charles, MO | A blend of layered aluminosilicate mineral clays | Kutz et al., 2009 [101] |
| Solis Mos | Novus International, Saint Charles, MO | Sodium montmorillonite with live yeast, yeast culture, mannan oligosaccharide, and vitamin E | Xiong et al., 2015 [104] |
| Toxy-Ni | Nutriad Animal Feed Additives, Dendermonde, Belgium | Adsorbent clay minerals and inactivated yeast (Saccharomyces cerevisiae) | Rodrigues et al., 2019 [52] |
| Unike Plus | Nutriad Animal Feed Additives, Dendermonde, Belgium | Adsorbent clay minerals, inactivated yeast (S. cerevisiae), botanical components, and a mixture of antioxidants and preservatives | Rodrigues et al., 2019 [52] |
| Mycofix Plus | Biomin GmbH, Herzogenburg, Australia | Bentonites, enzymes, Eubacterium strain (BBSH 797), and yeast strain T. mycotoxinivorans | Pietri et al., 2009 [105] |
| FloMatrix | PMI nutritional Additives, Arden Hills, MN | Aluminosilicate clay matrices and yeast components | Pate et al., 2018 [106] |
| Study | Aflatoxin Dose in Diet | Sequestering Agents | % of Sequestering Agents as Diet DM | % Reduction of Milk AFM1 |
|---|---|---|---|---|
| Harvey et al., 1991 [107] | 200 µg/kg AF | HSCAS | 0.5% | 24% |
| 100 µg/kg AF | HSCAS | 1% | 44% | |
| Diaz et al., 2004 [103], Exp 1 | 100 µg/kg total AF, 55% AFB1; 40% AFG1; 2% AFB2 and 3% AFG2 | Astra-Ben 20 1 | 1.2% | 61% |
| FlowGuard | 1.2% | 65% | ||
| Mycrosorb | 1.2% | 50% | ||
| Diaz et al., 2004 [103], Exp 2 | 100 µg/kg total AF. 55% AFB1; 40% AFG1; 2% AFB2 and 3% AFG2 | Astra-Ben 20 1 | 1.2% | 64.4% |
| Red Crown bentonite | 1.2% | 31.4% | ||
| MTB-100 1 | 0.05% | 58.5% | ||
| Activated Carbons | 0.25% | 5.4% | ||
| Masoero et al., 2008 [108] | 7.4 µg/kg AFB1 Exp 1 | Magnesium smectite clay (Atox) | 0.82% | 47.4% |
| 7.5 µg/kg AFB1 Exp 2 | AFB1-contaminated complete concentrate with magnesium smectite clay as a pellet | 0.83% | 76 ng/kg AFM1 in milk | |
| AFB1-contaminated complete concentrate with magnesium smectite clay as a meal | 0.83% | 111 ng/kg AFM1 in milk | ||
| Kutz et al., 2009 [101] | 100 µg/kg AFB1, a mixture of AFs contains 61% AFB1, 2% AFB2, 26% AFG1, 1% AFG2 | Solis 1 | 0.56% | 44.8% |
| NovasilPlus 1 | 0.56% | 47.9% | ||
| MTB-100 1 | 0.56% | 4.2% (NS) | ||
| Pietri et al., 2009 [105] | 97.3 µg/kg AFB1 | Mycofix Plus 1 | 0.08% | 31% |
| Mycofix Plus 1 | 0.2% | 41% | ||
| Queiroz et al., 2012 [5] | 75 µg/kg AF, 64% AFB1, 2% AFB2, 33% AFG1, and 0.003% AFG2 | Calibrin A 1 | 0.05% | −22% (NS) |
| Calibrin A 1 | 2% | 16% | ||
| Sumantri et al., 2012 [109] | 30.8 µg/kg AFB1 | Bentonite (type not described) | 0.005% | 1.7% (NS) |
| Bentonite (type not described) | 0.045% | 9.6% (NS) | ||
| Kissell et al., 2013 [110] Exp1 | 91 µg/kg AFB1 | Experimental product (yeast cell wall extract. glucomannan) and aluminosilicate (Lallemand) | 0.04% | −5.2% (NS) |
| Exp 2 | 94 µg/kg AFB1 | MTB-100-2004, formulation of 2004 | 004% | −8.0% (NS) |
| MTB-100-2006, formulation of 2006 | 0.04% | −6.2% (NS) | ||
| Experimental product (Alltech) | 0.04% | −9.5% (NS) | ||
| Exp 3 | 86 µg/kg AFB1 | MTB-100-2006 1 | 0.2% | −9.5% (NS) |
| Astra-Ben 20 1 | 0.9% | 60.4% | ||
| Xiong et al., 2015 [104] Exp1 | 20 µg/kg of AFB1 | Solis Mos 1 | 0.25% | 16% |
| Exp2 | 40 µg/kg of AFB1 | Solis Mos 1 | 0.25% | 2% (NS) |
| Maki et al., 2016 [102] | 100 µg/kg AF, 79% AFB1, 16% AFG1, 4% AFB2, and 1% AFG2. | NovasilPlus 1 | 0.58% | 47.3% |
| NovasilPlus 1 | 1.17% | 70.9% | ||
| Katsoulos et al., 2016 [111] | Not a feeding trial, data is from 15 commercial herds with milk AFM1> 0.05 µg/kg | Clinoptilolite, a natural zeolite, particle size <0.15mm | 1% | 58.1% |
| Clinoptilolite, a natural zeolite, particle size <0.8 mm | 1% | 53.2% | ||
| Ogunade et al., 2016 [48] | 75 µg/kg AFB1 | SCFP with low dose chlorophyll-based additive (Diamond V) | 0.09% | −0.01% (NS) |
| SCFP with high dose of chlorophyll-based additive (Diamond V) | 0.09% | 0% (NS) | ||
| Low dose of the additive and sodium bentonite clay (Diamond V) | 0.05% | −0.01% (NS) | ||
| Gonçalves et al., 2017 [112] | 480 µg/d AFB1, intake not reported | Yeast cell wall (ICC Brazil) | 20 g/cow/d | 69.4% |
| Autolyzed yeast from sugarcane industry (ICC Brazil) | 20 g/cow/d | 45.6% | ||
| Dried yeast from sugarcane industry (ICC Brazil) | 20 g/cow/d | 47.5% | ||
| Partially dehydrated yeast from brewery industry (ICC Brazil) | 20 g/cow/d | 62.8% | ||
| Sulzberger et al., 2017 [51] | 100 µg/kg AFB1, 71% AFB1, 2.5% AFB2, 24.8% AFG1, 0.006% AFG2 | Clay (composition not described, UMG Minerals Group) | 0.5% | 18.6% |
| Clay, (UMG Minerals Group) | 1% | 30.2% | ||
| Clay, (UMG Minerals Group) | 2% | 41.9% | ||
| Maki et al., 2017 [113] | 50 µg/kg AFB1 | Novasil Plus 1 | 0.125% | 17.3% |
| Novasil Plus 1 | 0.25% | 22.7% | ||
| Novasil Plus 1 | 0.5% | 71% | ||
| Pate et al., 2018 [106] | 100 µg/kg AFB1 | FloMatrix 1 | 0.4% | NS |
| FloMatrix 1 | 0.8% | NS | ||
| Weatherly et al., 2018 [114] | 100 µg/kg AFB1 | Yeast cell wall and bentonite clay (Biorigin) | 0.13% | −3.9% (NS) |
| Yeast cell wall and bentonite clay (Biorigin) | 0.26% | −4.6% (NS) | ||
| A prototype adsorbent (Biorigin) | 0.26% | −8.8% (NS) | ||
| Xiong et al., 2018 [104] | 20 µg/kg AFB1 | Solis Mos 1 | 0.25% | 31.6% |
| Rodrigues et al., 2019 [52] | 105.5 µg/kg AF, 72.2% AFB1, 2.4% AFB2, 24.8% AFG1, 0.6% AFG1 | Toxy-Ni 1 | 0.4% | 66.7% |
| Unike Plus 1 | 0.4% | 50% | ||
| Gallo et al., 2020 [54] | 17.53 µg/kg AFB1 | Smectite clay | 0.5% | 64.8% |
| Intanoo et al., 2020 [115] | 22.28 µg/kg AFB1 | K. marxianus CPY1 | Total 2 g, 1 × 109 CFU/g | 76.6% |
| K. marxianus RSY5 | Total 2 g, 1 × 109 CFU/g | 72.1% | ||
| P. kudriavzevii YSY2 | Total 2 g, 1 × 109 CFU/g | 66.9% |
| Approach | Most Effective Strains Identified | Maximum Binding Efficiency | Factors Affecting Binding Efficiency | Reference |
|---|---|---|---|---|
| In vitro | S. cerevisiae strain A18 | 69.1% | Strain, temperature, acidity environment, incubation time, aflatoxin concentration, growing phase of bacteria | Shetty and Jesperson (2007) [119] |
| In vitro | Dead L. buchneri R1102 Dead L. plantarum R2014 P. acidilactici EQ01 | 66.5% 60.5%, 56.9% | Dose, viability, bacteria species and pH | Ma et al. (2017) [123] |
| In vitro | L. rhamnosus strain GG L. rhamnosus strain LC-705 | 77% 75% | Strain Viability | Pierides et al. (2000) [121] |
| In vitro | L. amylovorus strain CSCC5160 | 73.2% | Stain and incubation time | Peltonen et al. (2001) [122] |
| L. amylovorus CSCC 5197 | 72.4 | |||
| Lactobacillus rhamnosus strain LC1/3 | 76.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Ogunade, I.M.; Vyas, D.; Adesogan, A.T. Aflatoxin in Dairy Cows: Toxicity, Occurrence in Feedstuffs and Milk and Dietary Mitigation Strategies. Toxins 2021, 13, 283. https://doi.org/10.3390/toxins13040283
Jiang Y, Ogunade IM, Vyas D, Adesogan AT. Aflatoxin in Dairy Cows: Toxicity, Occurrence in Feedstuffs and Milk and Dietary Mitigation Strategies. Toxins. 2021; 13(4):283. https://doi.org/10.3390/toxins13040283
Chicago/Turabian StyleJiang, Yun, Ibukun M. Ogunade, Diwakar Vyas, and Adegbola T. Adesogan. 2021. "Aflatoxin in Dairy Cows: Toxicity, Occurrence in Feedstuffs and Milk and Dietary Mitigation Strategies" Toxins 13, no. 4: 283. https://doi.org/10.3390/toxins13040283
APA StyleJiang, Y., Ogunade, I. M., Vyas, D., & Adesogan, A. T. (2021). Aflatoxin in Dairy Cows: Toxicity, Occurrence in Feedstuffs and Milk and Dietary Mitigation Strategies. Toxins, 13(4), 283. https://doi.org/10.3390/toxins13040283