Tentacle Morphological Variation Coincides with Differential Expression of Toxins in Sea Anemones

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Assembly Statistics
2.2. Functional Differences of Structures
2.3. Venom Repertoire and Toxin Expression
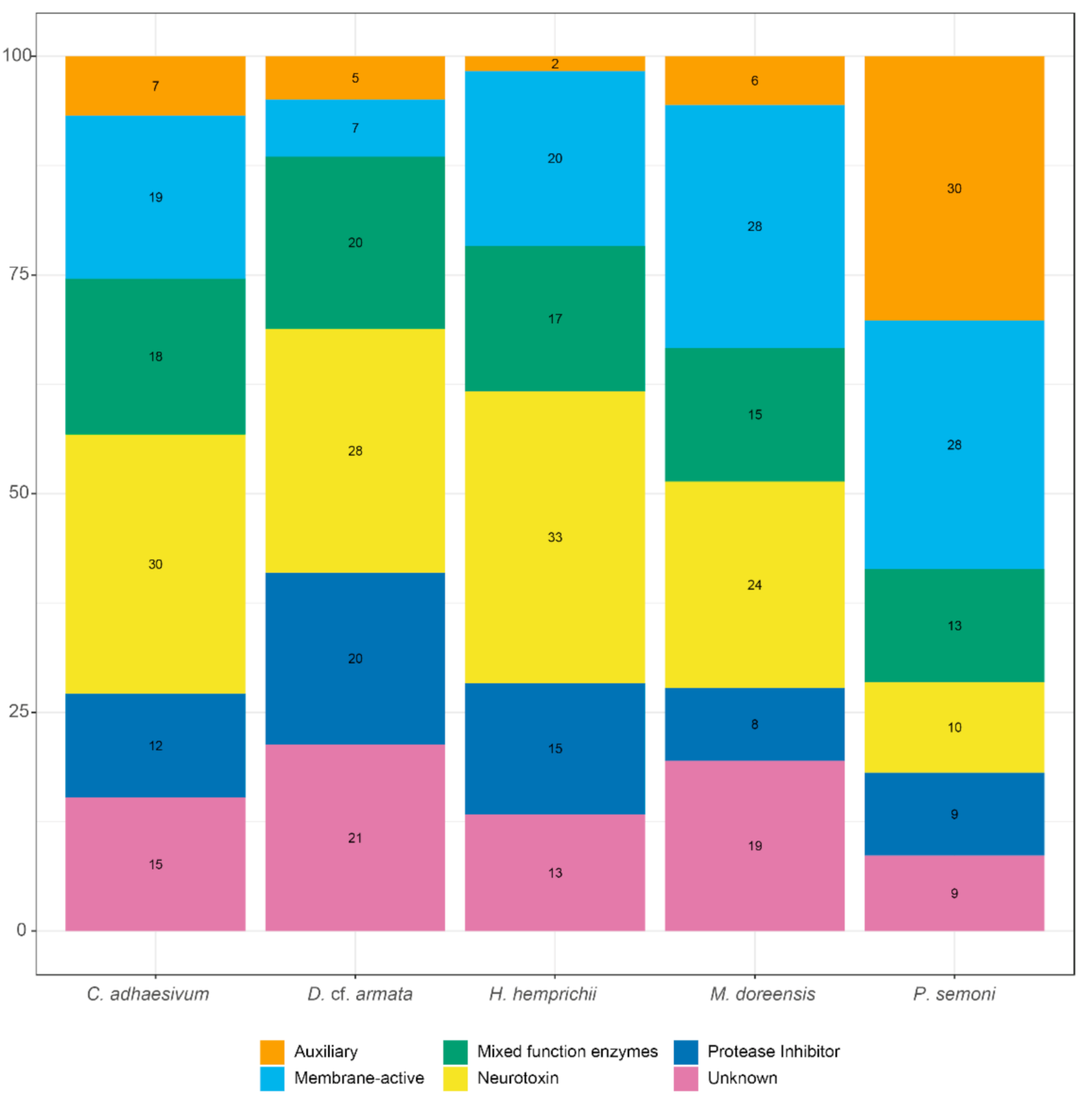
2.3.1. Comparison of Species Venom Arsenal
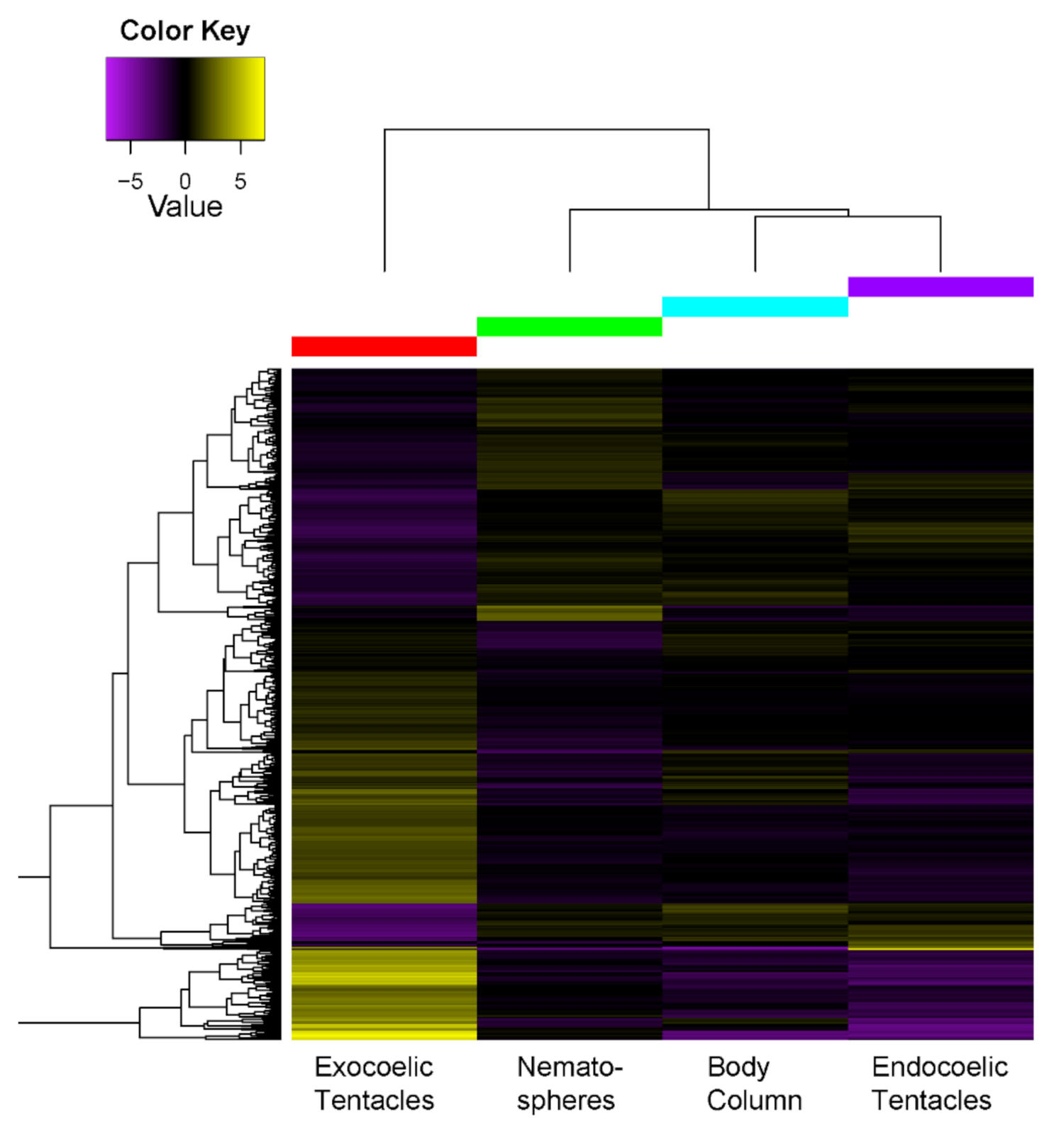
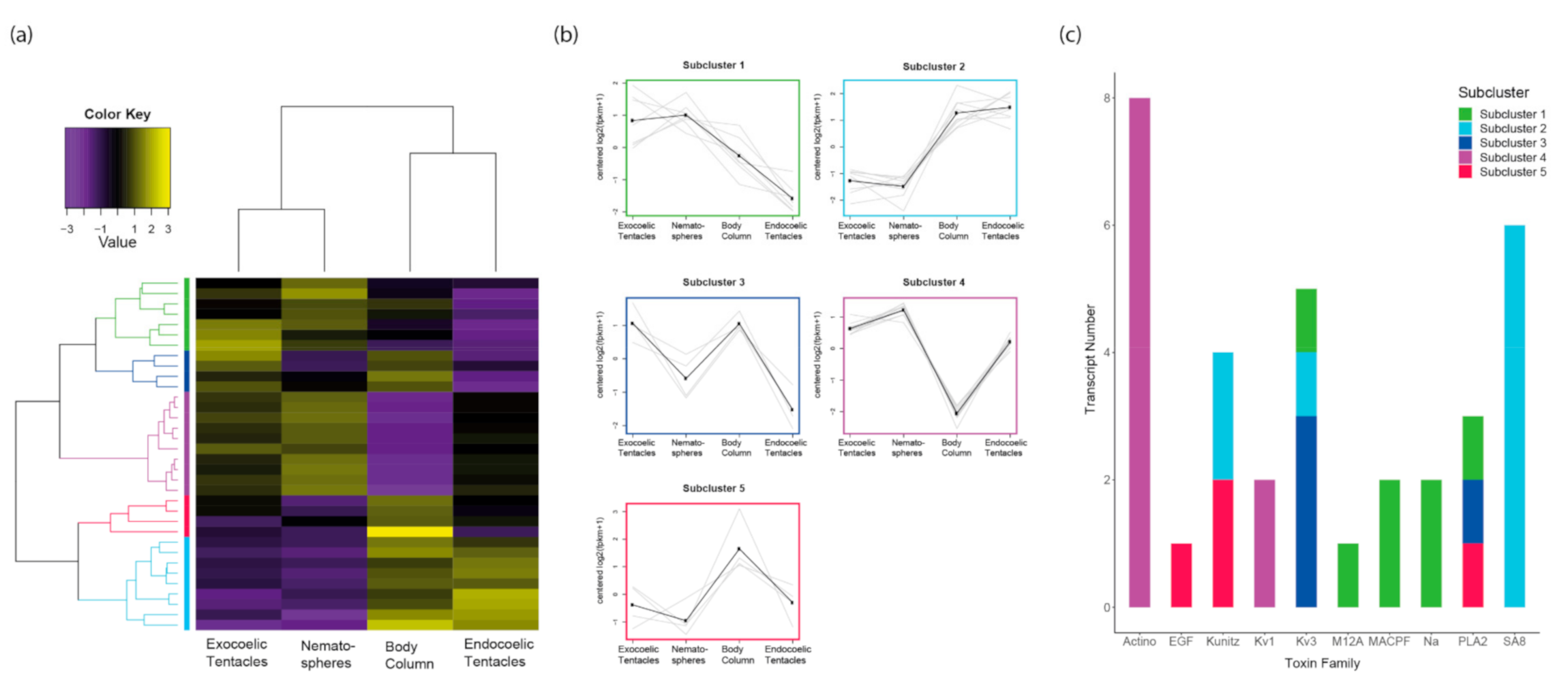
2.3.2. Tissue-Specific Expression of Toxin-Like Transcripts
3. Discussion
3.1. Tissue-Specific Toxin Expression Profiles of Tentacles Show Greater Divergence When Morphological Variation Is Present
3.2. Ecological Significance of Tentacular Structures, and Its Relationship with Toxin Expression Profiles
3.3. Unique Toxin Arsenals Are Required by Structures That Defend Endosymbiont-Hosting Structures
4. Materials and Methods
4.1. Sample Preparation and Sequencing
4.1.1. Heterodactyla hemprichii, Cryptodendrum adhaesivum and Phyllodiscus semoni
4.1.2. Dofleinia cf. armata
4.1.3. Macrodactyla doreensis
4.2. Transcriptome Assembly
4.3. Tissue-Specific Expression Profiles
4.3.1. Functional Annotation of Transcripts
4.3.2. Differential Expression Analysis and Gene Ontology Enrichment Analysis
4.4. Analysis of Toxin-Like Transcripts
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Augusto-de-Oliveira, C.; Stuginski, D.R.; Kitano, E.S.; Andrade-Silva, D.; Liberato, T.; Fukushima, I.; Serrano, S.M.; Zelanis, A. Dynamic rearrangement in snake venom gland proteome: Insights into Bothrops jararaca intraspecific venom variation. J. Proteome Res. 2016, 15, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Dowell, N.L.; Giorgianni, M.W.; Kassner, V.A.; Selegue, J.E.; Sanchez, E.E.; Carroll, S.B. The deep origin and recent loss of venom toxin genes in rattlesnakes. Curr. Biol. 2016, 26, 2434–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves-Machado, L.; Pla, D.; Sanz, L.; Jorge, R.J.B.; Leitao-De-Araujo, M.; Alves, M.L.M.; Alvares, D.J.; De Miranda, J.; Nowatzki, J.; de Morais-Zani, K.; et al. Combined venomics, venom gland transcriptomics, bioactivities, and antivenomics of two Bothrops jararaca populations from geographic isolated regions within the Brazilian Atlantic rainforest. J. Proteom. 2016, 135, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.W.; Liu, B.S.; Chien, K.Y.; Chiang, L.C.; Huang, S.Y.; Sung, W.C.; Wu, W.G. Cobra venom proteome and glycome determined from individual snakes of Naja atra reveal medically important dynamic range and systematic geographic variation. J. Proteom. 2015, 128, 92–104. [Google Scholar] [CrossRef]
- Lyons, K.; Dugon, M.M.; Healy, K. Diet breadth mediates the prey specificity of venom potency in snakes. Toxins 2020, 12, 74. [Google Scholar] [CrossRef] [Green Version]
- Massey, D.J.; Calvete, J.J.; Sanchez, E.E.; Sanz, L.; Richards, K.; Curtis, R.; Boesen, K. Venom variability and envenoming severity outcomes of the Crotalus scutulatus scutulatus (Mojave rattlesnake) from Southern Arizona. J. Proteom. 2012, 75, 2576–2587. [Google Scholar] [CrossRef]
- Mitchell, M.L.; Tonkin-Hill, G.Q.; Morales, R.A.; Purcell, A.W.; Papenfuss, A.T.; Norton, R.S. Tentacle transcriptomes of the speckled anemone (Actiniaria: Actiniidae: Oulactis sp.): Venom-related components and their domain structure. Mar. Biotechnol 2020, 1–13. [Google Scholar] [CrossRef]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient venom systems: A review on cnidaria toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [Green Version]
- Schendel, V.; Rash, D.L.; Jenner, A.R.; Undheim, A.B.E. The diversity of venom: The importance of behavior and venom system morphology in understanding its ecology and evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [Green Version]
- Daly, M. Functional and genetic diversity of toxins in sea anemones. In Evolution of Venomous Animals and Their Toxins; Malhotra, A., Ed.; Springer: Dordrecht, The Netherlands, 2017; pp. 87–104. [Google Scholar] [CrossRef]
- Fautin, D. Structural diversity, systematics, and evolution of cnidae. Toxicon 2009, 54, 1054–1064. [Google Scholar] [CrossRef]
- Watson, G.M.; Hessinger, D.A. Cnidocyte mechanoreceptors are tuned to the movements of swimming prey by chemoreceptors. Science 1989, 243, 1589–1591. [Google Scholar] [CrossRef]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of venom composition across a complex life cycle. eLife 2018, 7, e35014. [Google Scholar] [CrossRef]
- Surm, J.M.; Smith, H.L.; Madio, B.; Undheim, E.A.; King, G.F.; Hamilton, B.R.; Van der Burg, C.A.; Pavasovic, A.; Prentis, P.J. A process of convergent amplification and tissue-specific expression dominates the evolution of toxin and toxin-like genes in sea anemones. Mol. Ecol. 2019, 28, 2272–2289. [Google Scholar] [CrossRef] [Green Version]
- Macrander, J.; Broe, M.; Daly, M. Tissue-specific venom composition and differential gene expression in sea anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef] [Green Version]
- Purcell, J.E. Aggresive function and induced development of catch tentacles in the sea anemone Metridium Senile (Coelenterata, Actiniaria). Biol. Bull. 1977, 153, 355–368. [Google Scholar] [CrossRef]
- Fukui, Y. Catch tentacles in the sea anemone Haliplanella luciae. Mar. Biol 1986, 91, 245–251. [Google Scholar] [CrossRef]
- Watson, G.M.; Mariscal, R.N. Comparative ultrastructure of catch tentacles and feeding tentacles in the sea anemone Haliplanella. Tissue Cell 1983, 15, 939–953. [Google Scholar] [CrossRef]
- Williams, R.B. Acrorhagi, catch tentacles and sweeper tentacles: A synopsis of aggression of actiniarian and scleractinian cnidaria. Hydrobiologia 1991, 216, 539–545. [Google Scholar] [CrossRef]
- Chornesky, E.A. Induced development of sweeper tentacles on the reef coral Agaricia agaricites: A response to direct competition. Biol. Bull. 1983, 165, 569–581. [Google Scholar] [CrossRef]
- Lapid, E.D.; Chadwick, N.E. Long-term effects of competition on coral growth and sweeper tentacle development. Mar. Ecol. Prog. Ser. 2006, 313, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Sebens, K.P.; Miles, J.S. Sweeper tentacles in a Gorgonian octocoral: Morphological modifications for interference competition. Biol. Bull. 1988, 175, 378–387. [Google Scholar] [CrossRef]
- Wellington, G.M. Reversal of digestive interactions between Pacific reef corals: Mediation by sweeper tentacles. Oecologia 1980, 47, 340–343. [Google Scholar] [CrossRef]
- Yosef, O.; Popovits, Y.; Malik, A.; Ofek-Lalzer, M.; Mass, T.; Sher, D. A tentacle for every occasion: Comparing the hunting tentacles and sweeper tentacles, used for territorial competition, in the coral Galaxea fascicularis. BMC Genom. 2020, 21, 548. [Google Scholar] [CrossRef]
- Ashwood, L.M.; Norton, R.S.; Undheim, E.A.; Hurwood, D.A.; Prentis, P.J. Characterising functional venom profiles of anthozoans and medusozoans within their ecological context. Mar. Drugs 2020, 18, 202. [Google Scholar] [CrossRef] [Green Version]
- Crowther, A. Character Evolution in Light OF Phylogenetic Analysis and Taxonomic Revision of the Zooxanthellate Sea Anemone Families Thalassianthidae and Aliciidae. Ph.D. Dissertation, University of Kansas, Lawrence, KS, USA, 2013. [Google Scholar]
- Daly, M. The anatomy, terminology, and homology of acrorhagi and pseudoacrorhagi in sea anemones. Zool. Verh. 2003, 89–102. [Google Scholar]
- Shick, J.M. A Functional Biology of Sea Anemones; Chapman & Hall: London, UK, 1991. [Google Scholar]
- Song, J.-I.; Cha, H.-R. Taxonomy of Actiniidae (Anthozoa, Actiniaria, Thenaria, Endomvaria) from Korea. Anim. Syst. Evol. Divers. 2002, 18, 253–270. [Google Scholar]
- Carlgren, O.H. A Survey of the Ptychodactiaria, Corallimorpharia and Actiniaria. Kungl. Sven. Vetensk. Handl. 1949, 1, 1–122. [Google Scholar]
- Attaran-Fairman, G.; Javid, P.; Shakouri, A. Morphology and phylogeny of the sea anemone Stichodactyla haddoni (Cnidaria: Anthozoa: Actiniaria) from Chabahar Bay, Iran. Turk. Zool. Derg. 2015, 39, 998–1003. [Google Scholar] [CrossRef]
- Ardelean, A. Reinterpretation of some tentacular structures in actinodendronid and thalassianthid sea anemones (Cnidaria: Actiniaria). Zool. Verh. 2003, 31–40. [Google Scholar]
- Ardelean, A.; Fautin, D. Variability in nematocysts from a single individual of the sea anemone Actinodendron arboreum (Cnidaria: Anthozoa: Actiniaria). Hydrobiologia 2004, 530, 189–197. [Google Scholar] [CrossRef]
- Fautin, D.G.; Allen, G.R. Field Guide to Anemonefishes and Their Host sea Anemones; Western Australian Museum: Perth, Australia, 1992. [Google Scholar]
- Hoeksema, B.W.; Crowther, A.L. Masquerade, mimicry and crypsis of the polymorphic sea anemone Phyllodiscus semoni and its aggregations in South Sulawesi. Contrib. Zool. 2011, 80, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Gladfelter, W.B. Sea anemone with zooxanthellae: Simultaneous contraction and expansion in response to changing light intensity. Science 1975, 189, 570. [Google Scholar] [CrossRef] [PubMed]
- Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A.; Koshelev, S.G.; Sanamyan, N.P.; Sanamyan, K.E.; Dyachenko, I.A.; Bondarenko, D.A.; Murashev, A.N.; Mineev, K.S.; et al. Sea anemone peptide with uncommon β-hairpin structure inhibits acid-sensing ion channel 3 (ASIC3) and reveals analgesic activity. J. Biol. Chem. 2013, 288, 23116–23127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClounan, S.; Seymour, J. Venom and cnidome ontogeny of the cubomedusae Chironex fleckeri. Toxicon 2012, 60, 1335–1341. [Google Scholar] [CrossRef]
- Edmunds, M.; Potts, G.W.; Swinfen, R.C.; Waters, V.L. Defensive behaviour of sea anemones in response to predation by the opisthobranch mollusc Aeolidia papillosa (L.). J. Mar. Biol. Assoc. UK 1976, 56, 65–83. [Google Scholar] [CrossRef]
- Ho, G. Cryptodendrum Adhaesivum. Available online: https://www.gbri.org.au//SpeciesList//Cryptodendrumadhaesivum%7CGuenHo (accessed on 7 March 2021).
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Rodríguez, E.; Barbeitos, M.S.; Brugler, M.R.; Crowley, L.M.; Grajales, A.; Gusmão, L.; Häussermann, V.; Reft, A.; Daly, M. Hidden among sea anemones: The first comprehensive phylogenetic reconstruction of the order Actiniaria (Cnidaria, Anthozoa, Hexacorallia) reveals a novel group of hexacorals. PLoS ONE 2014, 9, e96998. [Google Scholar] [CrossRef]
- Dunn, D.F. The clownfish sea anemones: Stichodactylidae (Coelenterata: Actiniaria) and other sea anemones symbiotic with Pomacentrid fishes. Trans. Am. Philos. Soc. 1981, 71, 3–115. [Google Scholar] [CrossRef]
- Carlgren, O.H. Actiniaria and Corallimorpharia. In Scientific Reports of the Great Barrier Reef Expedition 1928–1929; BM(NH): London, UK, 1950; Volume 5, pp. 427–457. [Google Scholar]
- Carlgren, O.H. A contribution to the knowledge of the structure and distribution of the cnidae in the Anthozoa. K. Fysiogr. Sällskapets Handl. 1940, 51, 1–62. [Google Scholar]
- Dutertre, S.; Jin, A.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J. Evolution of separate predation-and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.A.; Mayhew, M.L.; Jin, J.; Herzig, V.; Undheim, E.A.B.; Sombke, A.; Fry, B.G.; Meritt, D.J.; King, G.F. The assassin bug Pristhesancus plagipennis produces two distinct venoms in separate gland lumens. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sachkova, M.Y.; Macrander, J.; Surm, J.M.; Aharoni, R.; Menard-Harvey, S.S.; Klock, A.; Leach, W.B.; Reitzel, A.M.; Moran, Y. Some like it hot: Population-specific adaptations in venom production to abiotic stressors in a widely distributed cnidarian. BMC Biol. 2020, 18, 121. [Google Scholar] [CrossRef]
- O’Hara, E.P.; Caldwell, G.S.; Bythell, J. Equistatin and equinatoxin gene expression is influenced by environmental temperature in the sea anemone Actinia equina. Toxicon 2018, 153, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Izumi, T.; Yanagi, K.; Fujita, T. Comprehensive revision of Anemonactis (Cnidaria: Anthozoa: Actiniaria: Haloclavidae) in Japan: Reestablishment of Anemonactis minuta (Wassilieff, 1908) comb. nov. and description of Anemonactis tohrui sp. nov. Mar. Biodivers. 2020, 50, 73. [Google Scholar] [CrossRef]
- Rodríguez, E.; López-González, P.J. Stephanthus antarcticus, a new genus and species of sea anemone (Actiniaria, Haloclavidae) from the South Shetland Islands, Antarctica. Helgol. Mar. Res. 2003, 57, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, M.; Nishikawa, K.; Yuzawa, Y.; Kanie, T.; Mori, H.; Araki, Y.; Hotta, N.; Matsuo, S. Acute renal failure after a sea anemone sting. Am. J. Kidney Dis. 2000, 36, E10. [Google Scholar] [CrossRef]
- Nakamoto, M.; Uezato, H. Stings of box-jellyfish and sea anemones. Clin. Derm. 1998, 52, 29–33. [Google Scholar]
- Brush, D.E. Marine envenomations. In Goldfrank’s Toxicologic Emergencies, 8th ed.; Flomenbaum, N.E., Goldfrank, L.R., Hoffman, R.S., Howland, M.A., Lewin, N.A., Nelson, L.S., Eds.; McGraw-Hill Medical: New York, NY, USA, 2006; pp. 1629–1642. [Google Scholar]
- Haddad, V., Jr.; Lupi, O.; Lonza, J.P.; Tyring, S.K. Tropical dermatology: Marine and aquatic dermatology. J. Am. Acad. Derm. 2009, 61, 733–750. [Google Scholar] [CrossRef]
- Ulrich, H.; Landthaler, M.; Vogt, T. Aquatic dermatoses. J. Dtsch. Dermatol. Ges. 2008, 6, 133–146. [Google Scholar] [CrossRef]
- Randall, J.; Fautin, D. Fishes other than anemonefishes that associate with sea anemones. Coral Reefs 2002, 21, 188–190. [Google Scholar] [CrossRef]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea anemones: Quiet achievers in the field of peptide toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Erhardt, H.; Knop, D. Corals: Indo-Pacific Field Guide; IKAN Unterwasserarchiv: Frankfurt, Germany, 2005. [Google Scholar]
- Acuña, F.H.; Garese, A. The cnidae of the acrospheres of the corallimorpharian Corynactis carnea (Studer, 1878) (Cnidaria, Corallimorpharia, Corallimorphidae): Composition, abundance and biometry. Belg. J. Zool. 2009, 139, 50–57. [Google Scholar]
- Riemann-Zürneck, K.; Iken, K. Corallimorphus profundus in shallow Antarctic habitats: Bionomics, histology, and systematics (Cnidaria: Hexacorallia). Zool. Verh. 2003, 345, 367–386. [Google Scholar]
- den Hartog, J.C. The genus Telmatactis Gravier, 1916 (Actiniaria: Acontiaria: Isophelliidae) in Greece and the eastern Mediterranean. Zool. Meded. 1995, 69, 153–176. [Google Scholar]
- van der Burg, C.A.; Prentis, P.J.; Surm, J.M.; Pavasovic, A. Insights into the innate immunome of actiniarians using a comparative genomic approach. BMC Genom. 2016, 17, 850. [Google Scholar] [CrossRef]
- Prentis, P.J.; Pavasovic, A. The Anadara trapezia transcriptome: A resource for molluscan physiological genomics. Mar. Genom. 2014, 18, 113–115. [Google Scholar] [CrossRef]
- Mitchell, M.L.; Shafee, T.; Papenfuss, A.T.; Norton, R.S. Evolution of cnidarian trans-defensins: Sequence, structure and exploration of chemical space. Proteins 2019, 87, 551–560. [Google Scholar] [CrossRef]
- Madio, B.; Undheim, E.A.; King, G.F. Revisiting venom of the sea anemone Stichodactyla haddoni: Omics techniques reveal the complete toxin arsenal of a well-studied sea anemone genus. J. Proteom. 2017, 166, 83–92. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Garber, M.; Grabherr, M.G.; Guttman, M.; Trapnell, C. Computational methods for transcriptome annotation and quantification using RNA-seq. Nat. Methods 2011, 8, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, J.K.; Wallace, C.C. Australian Anemones Final Report Accompanied by Attribution Database of Australian Anemones (on CD rom): Prepared for the Department of Environment and Heritage, Heritage Division; Museum of Tropical Queensland: Townsville, Australia, 2004. [Google Scholar]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranda, M.; Li, Y.; Liew, Y.J.; Baumgarten, S.; Simakov, O.; Wilson, M.C.; Piel, J.; Ashoor, H.; Bougouffa, S.; Bajic, V.B.; et al. Genomes of coral dinoflagellate symbionts highlight evolutionary adaptations conducive to a symbiotic lifestyle. Sci. Rep. 2016, 6, 39734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Cheng, S.; Song, B.; Zhong, X.; Lin, X.; Li, W.; Li, L.; Zhang, Y.; Zhang, H.; Ji, Z.; et al. The Symbiodinium kawagutii genome illuminates dinoflagellate gene expression and coral symbiosis. Science 2015, 350, 691–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoguchi, E.; Shinzato, C.; Kawashima, T.; Gyoja, F.; Mungpakdee, S.; Koyanagi, R.; Takeuchi, T.; Hisata, K.; Tanaka, M.; Fujiwara, M.; et al. Draft assembly of the Symbiodinium minutum nuclear genome reveals dinoflagellate gene structure. Curr. Biol. 2013, 23, 1399–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Young, M.; Wakefield, M.; Smyth, G.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Liao, Q.; Gong, G.; Poon, T.C.; Ang, I.L.; Lei, K.M.; Siu, S.W.I.; Wong, C.T.T.; Rádis-Baptista, G.; Lee, S.M.-Y. Combined transcriptomic and proteomic analysis reveals a diversity of venom-related and toxin-like peptides expressed in the mat anemone Zoanthus natalensis (Cnidaria, Hexacorallia). Arch. Toxicol. 2019, 93, 1745–1767. [Google Scholar] [CrossRef]
- Klompen, A.M.; Macrander, J.; Reitzel, A.M.; Stampar, S.N. Transcriptomic analysis of four cerianthid (Cnidaria, Ceriantharia) venoms. Mar. Drugs 2020, 18, 413. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. adhaesivum | D. cf. armata | H. hemprichii | M. doreensis | P. semoni | |
|---|---|---|---|---|---|
| Raw Reads | 200.5 million | 160.2 million | 240.3 million | 142.2 million | 305.0 million |
| Transcripts | 628,468 | 164,583 | 101,150 | 108,541 | 208,537 |
| Genes | 451,132 | 97,982 | 74,496 | 64,558 | 107,984 |
| DE 1 Transcripts | 3672 | 156 | 3118 | 271 | 1411 |
| N50 | 609 | 1514 | 1370 | 1493 | 2823 |
| E90 N50 | 1346 | 2068 | 1991 | 1549 | 3241 |
| E90 transcripts | 105,056 | 35,114 | 19,888 | 18,285 | 19,460 |
| BUSCO 2 | 93.9% | 97.2% | 92.0% | 88.9% | 95.0% |
| Category | ToxProt Family | C. adhaesivum | D. cf. armata | H. hemprichii | M. doreensis | P. semoni |
|---|---|---|---|---|---|---|
| Auxiliary | Peptidase M12A | 8 | 3 | 1 | 4 | 35 |
| Membrane-active | Actinoporin family | 15 | 0 | 9 | 9 | 4 |
| Jellyfish toxin family | 0 | 1 | 0 | 0 | 0 | |
| MACPF toxin family | 7 | 3 | 3 | 11 | 29 | |
| Mixed function enzymes | Phospholipase A2 family | 21 | 12 | 10 | 11 | 15 |
| Neurotoxin | Sea anemone type 3 (BDS) potassium channel toxin family | 24 | 11 | 13 | 7 | 2 |
| Sea anemone type 1 potassium channel toxin family | 6 | 2 | 3 | 6 | 2 | |
| Sea anemone type 5 potassium channel toxin family | 1 | 0 | 0 | 0 | 1 | |
| Sea anemone sodium channel inhibitory toxin family | 2 | 1 | 3 | 1 | 5 | |
| Cnidaria small cysteine-rich protein (SCRiP) family | 1 | 1 | 0 | 0 | 2 | |
| Sea anemone short toxin (type III) family | 1 | 0 | 1 | 0 | 0 | |
| Sea anemone structural class 9a family | 0 | 2 | 0 | 3 | 0 | |
| Protease Inhibitor | Venom Kunitz-type family | 14 | 12 | 9 | 6 | 11 |
| Unknown | EGF domain peptide family | 2 | 0 | 1 | 2 | 1 |
| Sea anemone 8 toxin family | 14 | 5 | 6 | 8 | 9 | |
| Acrorhagin | 2 | 3 | 1 | 1 | 0 | |
| Unknown | 0 | 5 | 0 | 3 | 0 | |
| 118 | 61 | 60 | 72 | 116 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashwood, L.M.; Mitchell, M.L.; Madio, B.; Hurwood, D.A.; King, G.F.; Undheim, E.A.B.; Norton, R.S.; Prentis, P.J. Tentacle Morphological Variation Coincides with Differential Expression of Toxins in Sea Anemones. Toxins 2021, 13, 452. https://doi.org/10.3390/toxins13070452
Ashwood LM, Mitchell ML, Madio B, Hurwood DA, King GF, Undheim EAB, Norton RS, Prentis PJ. Tentacle Morphological Variation Coincides with Differential Expression of Toxins in Sea Anemones. Toxins. 2021; 13(7):452. https://doi.org/10.3390/toxins13070452
Chicago/Turabian StyleAshwood, Lauren M., Michela L. Mitchell, Bruno Madio, David A. Hurwood, Glenn F. King, Eivind A. B. Undheim, Raymond S. Norton, and Peter J. Prentis. 2021. "Tentacle Morphological Variation Coincides with Differential Expression of Toxins in Sea Anemones" Toxins 13, no. 7: 452. https://doi.org/10.3390/toxins13070452
APA StyleAshwood, L. M., Mitchell, M. L., Madio, B., Hurwood, D. A., King, G. F., Undheim, E. A. B., Norton, R. S., & Prentis, P. J. (2021). Tentacle Morphological Variation Coincides with Differential Expression of Toxins in Sea Anemones. Toxins, 13(7), 452. https://doi.org/10.3390/toxins13070452