Lonomia obliqua Venom Induces NF-κB Activation and a Pro-Inflammatory Profile in THP-1-Derived Macrophage

 , ,
, ,
Abstract
:1. Introduction
2. Results
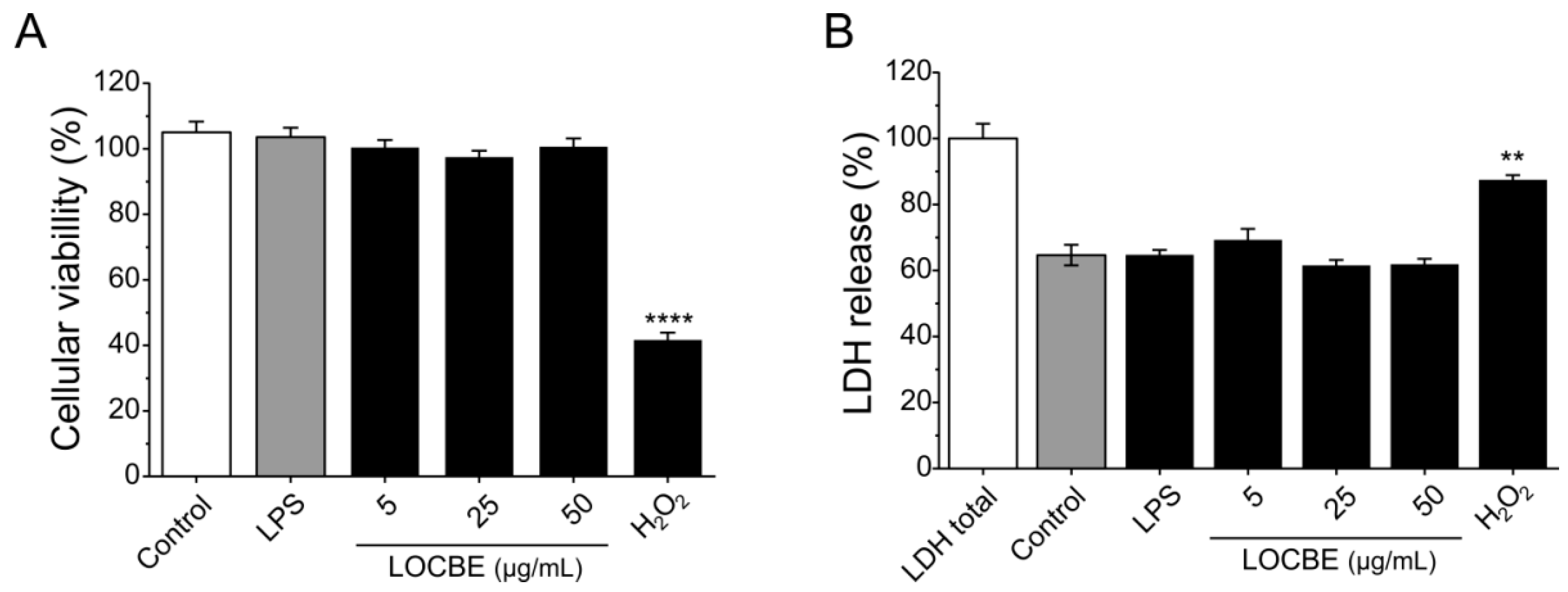
2.1. LOCBE Cytotoxic Effect on Macrophages
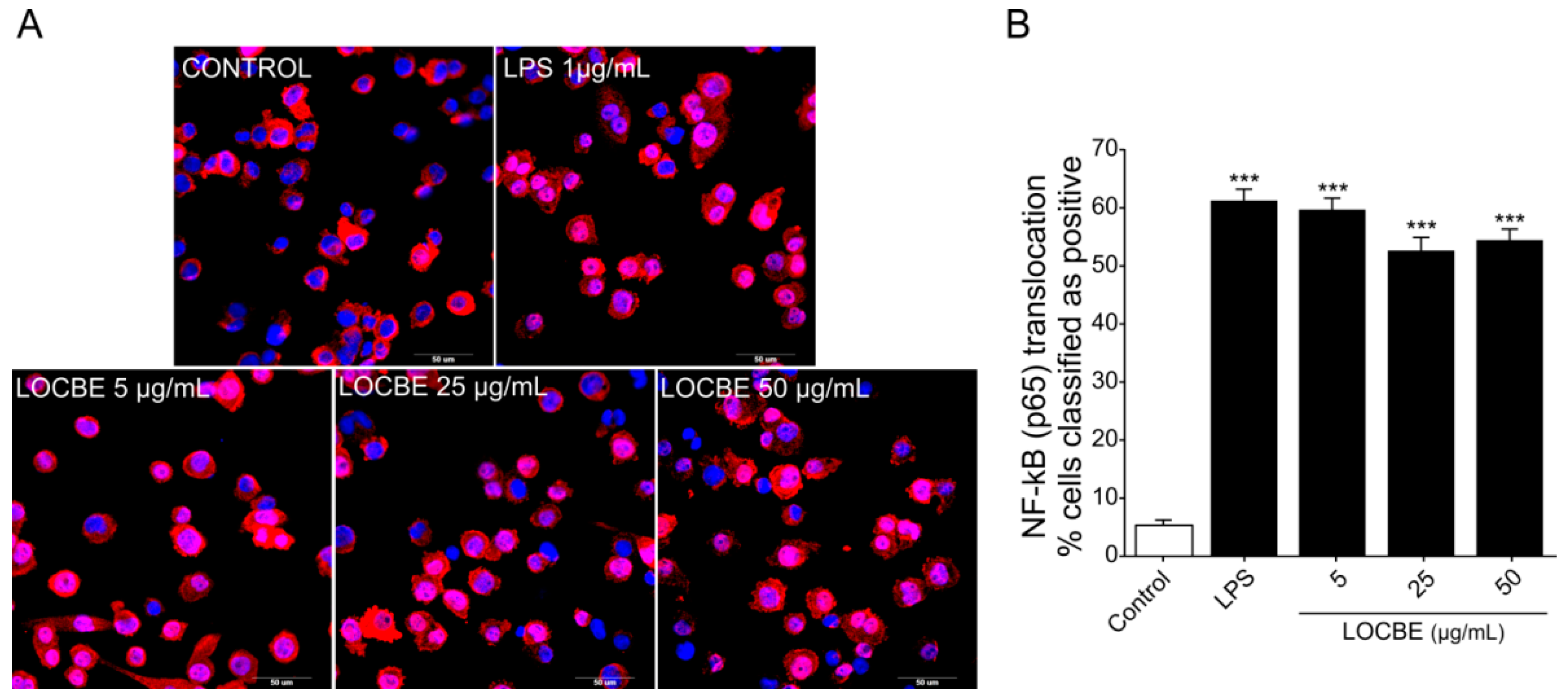
2.2. LOCBE Capacity to Activate NF-κB Pathway
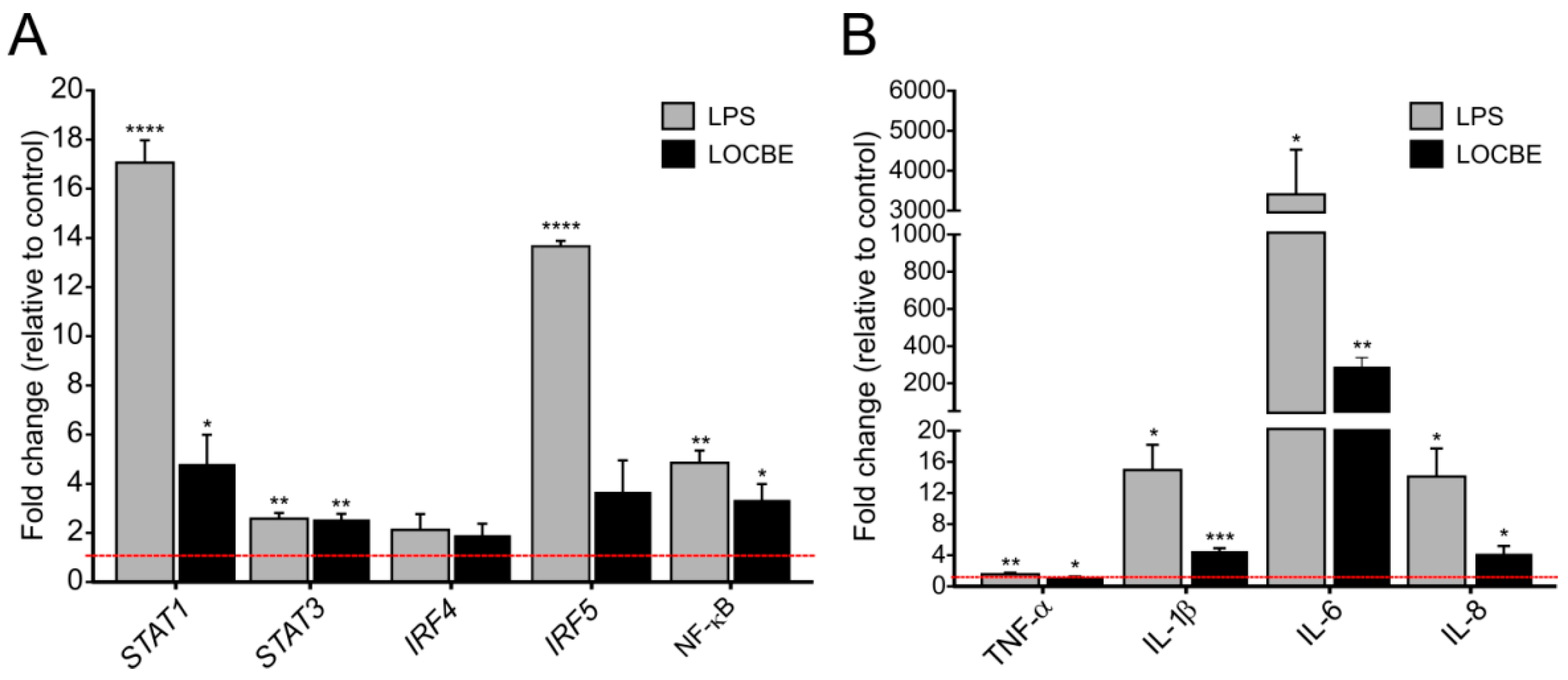
2.3. LOCBE Upregulates Inflammatory Genes Expression
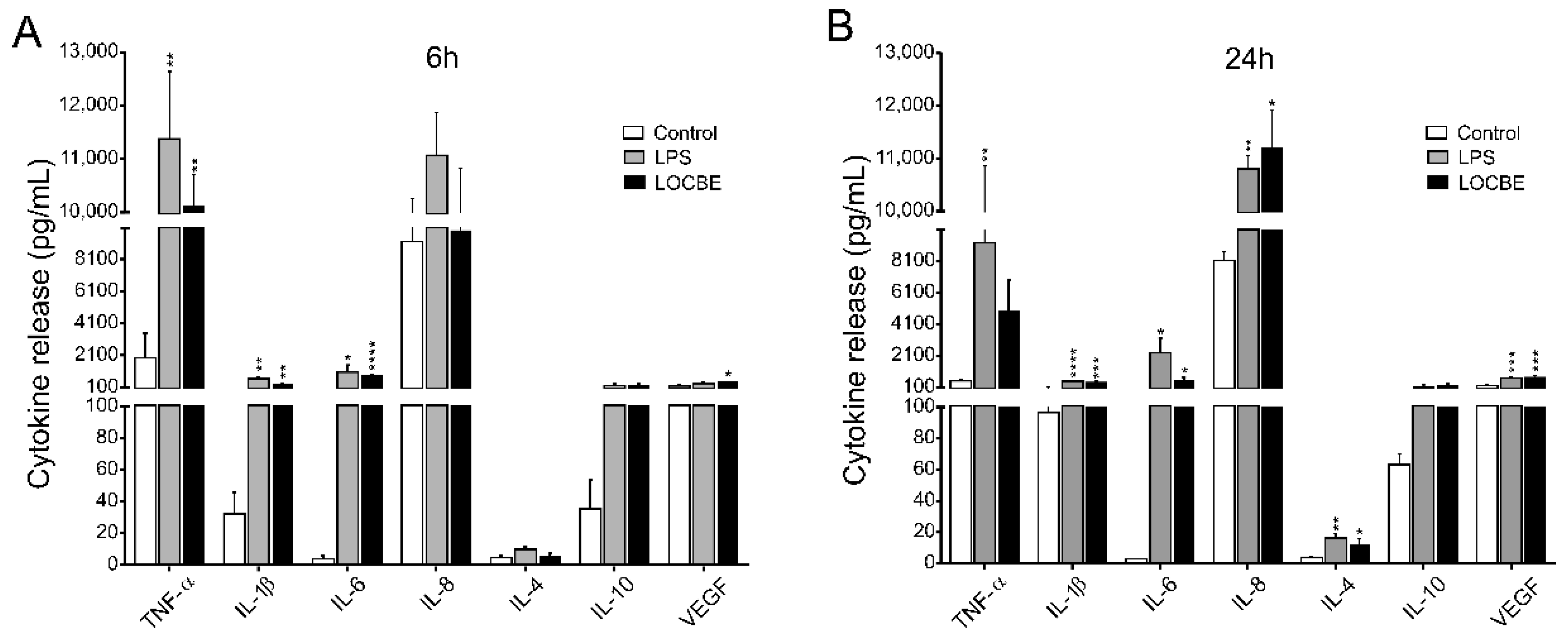
2.4. LOCBE Induces Inflammatory Cytokine and Chemokine Release
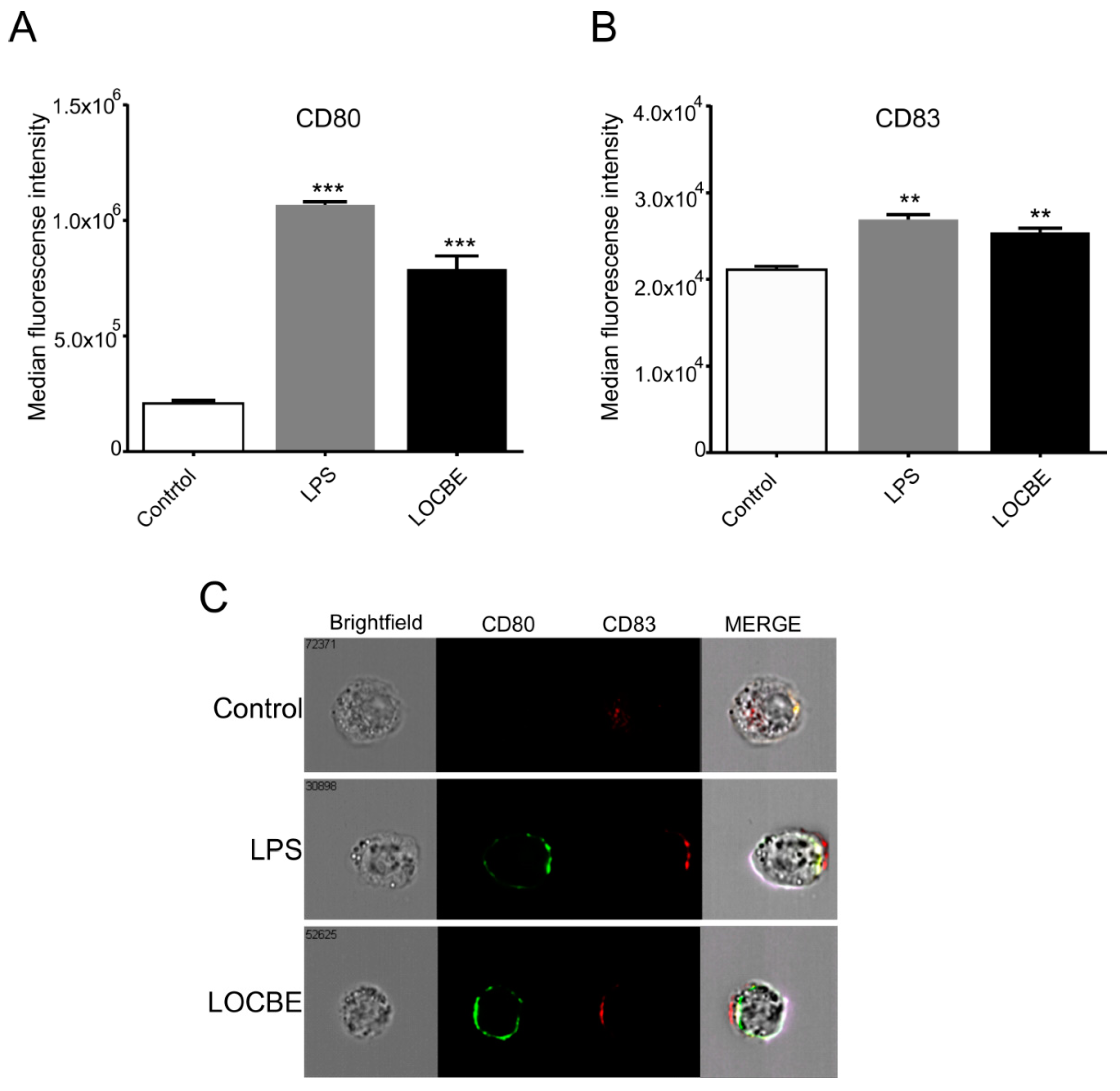
2.5. LOCBE Increases Expression of Membrane Markers
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Lonomia obliqua Crude Bristle Extract Obtention
5.2. Cell Culture and Differentiation
5.3. Cellular Viability and Cytotoxicity
5.4. NF-κB Pathway Activation
5.5. Gene Expression
5.6. Cytokine and Chemokine Profile Liberation
5.7. Cell Surface Proteins Expression
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Arocha-Piñango, C.L.; Guerrero, B. Hemorrhagic syndrome induced by caterpillars. Clinical and experimental studies. Review. Investig. Clin. 2003, 44, 155–163. [Google Scholar]
- Lemaire, C. Révision Du Genre Lonomia Walker [Lep. Attacidae]. Ann. Soc. Entomol. France 1972, 8, 767–861. [Google Scholar]
- Lemaire, C. Descriptions d’Attacidae Nouveaux d’Amérique Centrale et Du Sud. (Lep.). I. Descriptions Preliminaires de 16 Espécies Nouvelles Du Genre Lonomia Walker. Bull. Soc. Entomol. 1972, 76, 197–205. [Google Scholar]
- Arocha-Piñango, C.L.; de Bosch, N.B.; Torres, A.; Goldstein, C.; Nouel, A.; Argüello, A.; Carvajal, Z.; Guerrero, B.; Ojeda, A.; Rodriguez, A. Six New Cases of a Caterpillar-Induced Bleeding Syndrome. Thromb. Haemost. 1992, 67, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.N.; Mignone Chagas, M.A.; Casertano, S.A.; Cavagnaro, L.E.; Peichoto, M.E. Accidents with caterpillar Lonomia obliqua (Walker, 1855). An emerging problem. Medicine (Buenos Aires) 2015, 75, 328–333. [Google Scholar]
- DIVE-Diretoria de Vigilância Epidemiológica Acidente Com Lagarta Lonomia obliqua Na Região Oeste de Santa Catarina. Available online: http://www.dive.sc.gov.br/index.php/arquivo-noticias/631-acidente-com-lagarta-lonomia-obliqua-na-regiao-oeste-de-santa-catarina (accessed on 11 January 2020).
- Carrijo-Carvalho, L.C.; Chudzinski-Tavassi, A.M. The Venom of the Lonomia Caterpillar: An Overview. Toxicon 2007, 49, 741–757. [Google Scholar] [CrossRef] [PubMed]
- Kelen, E.M.A.; Picarelli, Z.P.; Duarte, A.C. Hemorrhagic Syndrome Induced by Contact with Caterpillars of the Genus Lonomia (Saturniidae, Hemileucinae). J. Toxicol. Toxin Rev. 1995, 14, 283–308. [Google Scholar] [CrossRef]
- Kowacs, P.A.; Cardoso, J.; Entres, M.; Novak, E.M.; Werneck, L.C. Fatal Intracerebral Hemorrhage Secondary to Lonomia obliqua Caterpillar Envenoming: Case Report. Arq. Neuro-Psiquiatr. 2006, 64, 1030–1032. [Google Scholar] [CrossRef] [Green Version]
- Portela Gamborgi, G.; Brett Metcalf, E.; Barros, E.J.G. Acute Renal Failure Provoked by Toxin from Caterpillars of the Species Lonomia obliqua. Toxicon 2006, 47, 68–74. [Google Scholar] [CrossRef]
- Zannin, M.; Lourenço, D.M.; Motta, G.; Dalla Costa, L.R.; Grando, M.; Gamborgi, G.P.; Noguti, M.A.; Chudzinski-Tavassi, A.M. Blood Coagulation and Fibrinolytic Factors in 105 Patients with Hemorrhagic Syndrome Caused by Accidental Contact with Lonomia obliqua Caterpillar in Santa Catarina, Southern Brazil. Thromb. Haemost. 2003, 89, 355–364. [Google Scholar] [CrossRef]
- Sano-Martins, I.S.; Duarte, A.C.; Guerrero, B.; Moraes, R.H.P.; Barros, E.J.G.; Arocha-Piñango, C.L. Hemostatic Disorders Induced by Skin Contact with Lonomia obliqua (Lepidoptera, Saturniidae) Caterpillars. Rev. Inst. Med. Trop. Sao Paulo 2017, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, M.; Reck, J.; Terra, R.M.S.; da Silva, W.O.B.; Santi, L.; Pinto, A.F.M.; Vainstein, M.H.; Termignoni, C.; Guimarães, J.A. Lonomia obliqua Venomous Secretion Induces Human Platelet Adhesion and Aggregation. J. Thromb. Thrombolysis 2010, 30, 300–310. [Google Scholar] [CrossRef]
- Pinto, A.F.M.; Dragulev, B.; Guimarães, J.A.; Fox, J.W. Novel Perspectives on the Pathogenesis of Lonomia obliqua Caterpillar Envenomation Based on Assessment of Host Response by Gene Expression Analysis. Toxicon 2008, 51, 1119–1128. [Google Scholar] [CrossRef]
- Reis, C.V.; Andrade, S.A.; Ramos, O.H.P.; Ramos, C.R.R.; Ho, P.L.; de Fátima Correia Batista, I.; Chudzinski-Tavassi, A.M. Lopap, a Prothrombin Activator from Lonomia obliqua Belonging to the Lipocalin Family: Recombinant Production, Biochemical Characterization and Structure–Function Insights. Biochem. J. 2006, 398, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Alvarez Flores, M.P.; Fritzen, M.; Reis, C.V.; Chudzinski-Tavassi, A.M. Losac, a Factor X Activator from Lonomia obliqua Bristle Extract: Its Role in the Pathophysiological Mechanisms and Cell Survival. Biochem. Biophys. Res. Commun. 2006, 343, 1216–1223. [Google Scholar] [CrossRef]
- Reis, C.V.; Farsky, S.H.P.; Fernandes, B.L.; Santoro, M.L.; Oliva, M.L.V.; Mariano, M.; Chudzinski-Tavassi, A.M. In Vivo Characterization of Lopap, a Prothrombin Activator Serine Protease from the Lonomia obliqua Caterpillar Venom. Thromb. Res. 2001, 102, 437–443. [Google Scholar] [CrossRef]
- de Castro Bastos, L.; Veiga, A.B.G.; Guimarães, J.A.; Tonussi, C.R. Nociceptive and Edematogenic Responses Elicited by a Crude Bristle Extract of Lonomia obliqua Caterpillars. Toxicon 2004, 43, 273–278. [Google Scholar] [CrossRef]
- Bohrer, C.B.; Reck Junior, J.; Fernandes, D.; Sordi, R.; Guimarães, J.A.; Assreuy, J.; Termignoni, C. Kallikrein–Kinin System Activation by Lonomia obliqua Caterpillar Bristles: Involvement in Edema and Hypotension Responses to Envenomation. Toxicon 2007, 49, 663–669. [Google Scholar] [CrossRef]
- Nascimento-Silva, V.; da Silva, G.R.; Moraes, J.A.; Cyrino, F.Z.; Seabra, S.H.; Bouskela, E.; Guimarães, J.A.; Barja-Fidalgo, C. A Pro-Inflammatory Profile of Endothelial Cell in Lonomia obliqua Envenomation. Toxicon 2012, 60, 50–60. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.C.; Balz, D.; de Souza, J.B.D.; Morsch, V.M.; Corrêa, M.C.; Zanetti, G.D.; Manfron, M.P.; Schetinger, M.R.C. Inhibition of NTPDase, 5′-Nucleotidase, Na+/K+-ATPase and Acetylcholinesterase Activities by Subchronic Treatment with Casearia Sylvestris. Phytomedicine 2006, 13, 509–514. [Google Scholar] [CrossRef]
- Berger, M.; de Moraes, J.A.; Beys-da-Silva, W.O.; Santi, L.; Terraciano, P.B.; Driemeier, D.; Cirne-Lima, E.O.; Passos, E.P.; Vieira, M.A.R.; Barja-Fidalgo, T.C.; et al. Renal and Vascular Effects of Kallikrein Inhibition in a Model of Lonomia obliqua Venom-Induced Acute Kidney Injury. PLoS Negl. Trop. Dis. 2019, 13, e0007197. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Beys-da-Silva, W.O.; Santi, L.; de Oliveira, I.M.; Jorge, P.M.; Henriques, J.A.P.; Driemeier, D.; Vieira, M.A.R.; Guimarães, J.A. Acute Lonomia obliqua Caterpillar Envenomation-Induced Physiopathological Alterations in Rats: Evidence of New Toxic Venom Activities and the Efficacy of Serum Therapy to Counteract Systemic Tissue Damage. Toxicon 2013, 74, 179–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraes, J.; Rodrigues, G.; Nascimento-Silva, V.; Renovato-Martins, M.; Berger, M.; Guimarães, J.; Barja-Fidalgo, C. Effects of Lonomia obliqua Venom on Vascular Smooth Muscle Cells: Contribution of NADPH Oxidase-Derived Reactive Oxygen Species. Toxins 2017, 9, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chudzinski-Tavassi, A.M.; Schattner, M.; Fritzen, M.; Pozner, R.G.; Reis, C.V.; Lourenço, D.; Lazzari, M.A. Effects of Lopap on Human Endothelial Cells and Platelets. Pathophysiol. Haemost. Thromb. 2001, 31, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Fritzen, M.; Flores, M.P.A.; Reis, C.V.; Chudzinski-Tavassi, A.M. A Prothrombin Activator (Lopap) Modulating Inflammation, Coagulation and Cell Survival Mechanisms. Biochem. Biophys. Res. Commun. 2005, 333, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.J.; DiPietro, L.A. Inflammation and Wound Healing: The Role of the Macrophage. Expert Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef] [Green Version]
- Hume, D.A. The Many Alternative Faces of Macrophage Activation. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS–) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D.C. NF-ΚB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Zen, K. Molecular Mechanisms That Influence the Macrophage M1–M2 Polarization Balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.K.; Mantovani, A. Macrophage Plasticity and Interaction with Lymphocyte Subsets: Cancer as a Paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef]
- Grohmann, U.; Belladonna, M.L.; Vacca, C.; Bianchi, R.; Fallarino, F.; Orabona, C.; Fioretti, M.C.; Puccetti, P. Positive Regulatory Role of IL-12 in Macrophages and Modulation by IFN-γ. J. Immunol. 2001, 167, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamunaa, A.; Vejayan, J.; Halijah, I.; Sharifah, S.; Ambu, S. Cytotoxicity of Southeast Asian Snake Venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Heinen, T.E.; de Farias, C.B.; Abujamra, A.L.; Mendonça, R.Z.; Roesler, R.; da Veiga, A.B.G. Effects of Lonomia obliqua Caterpillar Venom upon the Proliferation and Viability of Cell Lines. Cytotechnology 2014, 66, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Flores, M.P.; Hébert, A.; Gouelle, C.; Geller, S.; Chudzinski-Tavassi, A.M.; Pellerin, L. Neuroprotective Effect of RLosac on Supplement-Deprived Mouse Cultured Cortical Neurons Involves Maintenance of Monocarboxylate Transporter MCT2 Protein Levels. J. Neurochem. 2019, 148, 80–96. [Google Scholar] [CrossRef] [Green Version]
- Bosch, R.V.; Alvarez-Flores, M.P.; Maria, D.A.; Chudzinski-Tavassi, A.M. Hemolin Triggers Cell Survival on Fibroblasts in Response to Serum Deprivation by Inhibition of Apoptosis. Biomed. Pharmacother. 2016, 82, 537–546. [Google Scholar] [CrossRef]
- Waismam, K.; Chudzinski-Tavassi, A.M.; Carrijo-Carvalho, L.C.; Fernandes Pacheco, M.T.; Farsky, S.H.P. Lopap: A Non-Inflammatory and Cytoprotective Molecule in Neutrophils and Endothelial Cells. Toxicon 2009, 53, 652–659. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Darnell, J.; Kerr, I.; Stark, G. Jak-STAT Pathways and Transcriptional Activation in Response to IFNs and Other Extracellular Signaling Proteins. Science 1994, 264, 1415–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Overmeire, E.; Laoui, D.; Keirsse, J.; Bonelli, S.; Lahmar, Q.; Van Ginderachter, J.A. STAT of the Union: Dynamics of Distinct Tumor-Associated Macrophage Subsets Governed by STAT1: Highlights. Eur. J. Immunol. 2014, 44, 2238–2242. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, A.P.; Diez, D.; Miranda-Saavedra, D. The IL-10/STAT3-Mediated Anti-Inflammatory Response: Recent Developments and Future Challenges. Brief. Funct. Genom. 2013, 12, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Metzemaekers, M.; Vanheule, V.; Janssens, R.; Struyf, S.; Proost, P. Overview of the Mechanisms That May Contribute to the Non-Redundant Activities of Interferon-Inducible CXC Chemokine Receptor 3 Ligands. Front. Immunol. 2018, 8, 1970. [Google Scholar] [CrossRef]
- Levings, M.K.; Schrader, J.W. IL-4 Inhibits the Production of TNF-α and IL-12 by STAT6-Dependent and -Independent Mechanisms. J. Immunol. 1999, 162, 5224–5229. [Google Scholar] [PubMed]
- Onderdijk, A.J.; Baerveldt, E.M.; Kurek, D.; Kant, M.; Florencia, E.F.; Debets, R.; Prens, E.P. IL-4 Downregulates IL-1β and IL-6 and Induces GATA3 in Psoriatic Epidermal Cells: Route of Action of a Th2 Cytokine. J. Immunol. 2015, 195, 1744–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrante, C.J.; Pinhal-Enfield, G.; Elson, G.; Cronstein, B.N.; Hasko, G.; Outram, S.; Leibovich, S.J. The Adenosine-Dependent Angiogenic Switch of Macrophages to an M2-Like Phenotype Is Independent of Interleukin-4 Receptor Alpha (IL-4Rα) Signaling. Inflammation 2013, 36, 921–931. [Google Scholar] [CrossRef]
- Hefetz-Sela, S.; Stein, I.; Klieger, Y.; Porat, R.; Sade-Feldman, M.; Zreik, F.; Nagler, A.; Pappo, O.; Quagliata, L.; Dazert, E.; et al. Acquisition of an Immunosuppressive Protumorigenic Macrophage Phenotype Depending on C-Jun Phosphorylation. Proc. Natl. Acad. Sci. USA 2014, 111, 17582–17587. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T.; Natoli, G. Transcriptional Regulation of Macrophage Polarization: Enabling Diversity with Identity. Nat. Rev. Immunol. 2011, 11, 750–761. [Google Scholar] [CrossRef]
- Moro-García, M.A.; Mayo, J.C.; Sainz, R.M.; Alonso-Arias, R. Influence of Inflammation in the Process of T Lymphocyte Differentiation: Proliferative, Metabolic, and Oxidative Changes. Front. Immunol. 2018, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Johns, D.G.; Webb, R.C. TNF-α-Induced Endothelium-Independent Vasodilation: A Role for Phospholipase A2-Dependent Ceramide Signaling. Am. J. Physiol. Heart Circ. Physiol. 1998, 275, H1592–H1598. [Google Scholar] [CrossRef]
- Huang, Y.; Yan, L.; Rong, S.; Haller, H.; Kirch, T. TNF-α Induces Endothelial Dysfunction via PKC-ζ-Dependent NADPH Oxidase Activation. J. Huazhong Univ. Sci. Technol. Med. Sci. 2012, 32, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Kobayashi, K.; Yamanaka, H.; Okubo, M.; Noguchi, K. Microglial TNFα Induces COX2 and PGI2 Synthase Expression in Spinal Endothelial Cells during Neuropathic Pain. eNeuro 2017, 4, ENEURO.0064-17.2017. [Google Scholar] [CrossRef]
- Barrios, M.; Taylor, P.; Rodríguez-Acosta, A.; Sánchez, E.E.; Arocha-Piñango, C.L.; Gil, A.; Salazar, A.M.; Carvajal, Z.; Abad, M.J.; Guerrero, B. A Mouse Model to Study the Alterations in Haemostatic and Inflammatory Parameters Induced by Lonomia Achelous Caterpillar Haemolymph. Toxicon 2012, 59, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Nery, N.M.; Setúbal, S.S.; Boeno, C.N.; Lopes, J.A.; Paloschi, M.V.; Pontes, A.S.; Luna, K.P.; Zuliani, J.P. Bothrops Erythromelas Venom and Its Action on Isolated Murine Macrophages. Toxicon 2020, 185, 156–163. [Google Scholar] [CrossRef]
- Petricevich, V.L.; Hernández Cruz, A.; Coronas, F.I.V.; Possani, L.D. Toxin Gamma from Tityus Serrulatus Scorpion Venom Plays an Essential Role in Immunomodulation of Macrophages. Toxicon 2007, 50, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Salazar, E.; Salazar, A.M.; Taylor, P.; Urdanibia, I.; Pérez, K.; Rodríguez-Acosta, A.; Sánchez, E.E.; Guerrero, B. Contribution of Endothelial Cell and Macrophage Activation in the Alterations Induced by the Venom of Micrurus Tener Tener in C57BL/6 Mice. Mol. Immunol. 2019, 116, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Setubal, S.; Pontes, A.; Furtado, J.; Kayano, A.; Stábeli, R.; Zuliani, J. Effect of Bothrops Alternatus Snake Venom on Macrophage Phagocytosis and Superoxide Production: Participation of Protein Kinase C. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17. [Google Scholar] [CrossRef]
- Khemili, D.; Valenzuela, C.; Laraba-Djebari, F.; Hammoudi-Triki, D. Differential Effect of Androctonus Australis Hector Venom Components on Macrophage KV Channels: Electrophysiological Characterization. Eur. Biophys. J. 2019, 48, 1–13. [Google Scholar] [CrossRef]
- Zoccal, K.F.; da Silva Bitencourt, C.; Secatto, A.; Sorgi, C.A.; de Castro Figueredo Bordon, K.; Sampaio, S.V.; Arantes, E.C.; Faccioli, L.H. Tityus Serrulatus Venom and Toxins Ts1, Ts2 and Ts6 Induce Macrophage Activation and Production of Immune Mediators. Toxicon 2011, 57, 1101–1108. [Google Scholar] [CrossRef]
- Lund, M.E.; To, J.; O’Brien, B.A.; Donnelly, S. The Choice of Phorbol 12-Myristate 13-Acetate Differentiation Protocol Influences the Response of THP-1 Macrophages to a pro-Inflammatory Stimulus. J. Immunol. Methods 2016, 430, 64–70. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Buchser, W.; Collins, M.; Garyantes, T.; Guha, R.; Haney, S.; Lemmon, V.; Li, Z.; Trask, O.J. Assay Development Guidelines for Image-Based High Content Screening, High Content Analysis and High Content Imaging. In Assay Guidance Manual; Sittampalam, G.S., Grossman, A., Brimacombe, K., Arkin, M., Auld, D., Austin, C.P., Baell, J., Bejcek, B., Caaveiro, J.M.M., Chung, T.D.Y., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte (pg/mL) | Control | LPS | LOCBE | |||
|---|---|---|---|---|---|---|
| 6 h | 24 h | 6 h | 24 h | 6 h | 24 h | |
| CXCL10 | 219.8 ± 101 | 2670 ± 200.3 | 13,831 ± 2379 ** | 16,729 ± 503.3 **** | 11,888 ± 1362 ** | 17,275 ± 107.6 **** |
| CCL22 | 17.53 ± 1.2 | 114.1 ± 23.1 | 69.20 ± 15.7 * | 2405 ± 612.6 * | 36.23 ± 12.5 | 2283 ± 263.5 *** |
| CCL2 | 5810 ± 1826 | 455.6 ± 51.9 | 8678 ± 341.5 | 8448 ± 217.3 **** | 7399 ± 979.9 | 6015 ± 1471 * |
| CCL3 | 5587 ± 1329 | 450.9 ± 110 | 7887 ± 312.8 | 8108 ± 92.5 **** | 7294 ± 732 | 8200↑ |
| CCL4 | 5802 ± 1462 | 5161 ± 784.3 | 8643↑ | 8643↑ | 8090 ± 553.5 | 8643↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, D.S.; de Souza, J.G.; Alvarez-Flores, M.P.; Cunegundes, P.S.; DeOcesano-Pereira, C.; Lobba, A.M.; Gomes, R.N.; Chudzinski-Tavassi, A.M. Lonomia obliqua Venom Induces NF-κB Activation and a Pro-Inflammatory Profile in THP-1-Derived Macrophage. Toxins 2021, 13, 462. https://doi.org/10.3390/toxins13070462
Oliveira DS, de Souza JG, Alvarez-Flores MP, Cunegundes PS, DeOcesano-Pereira C, Lobba AM, Gomes RN, Chudzinski-Tavassi AM. Lonomia obliqua Venom Induces NF-κB Activation and a Pro-Inflammatory Profile in THP-1-Derived Macrophage. Toxins. 2021; 13(7):462. https://doi.org/10.3390/toxins13070462
Chicago/Turabian StyleOliveira, Douglas Souza, Jean Gabriel de Souza, Miryam Paola Alvarez-Flores, Priscila S. Cunegundes, Carlos DeOcesano-Pereira, Aline Maia Lobba, Renata N. Gomes, and Ana Marisa Chudzinski-Tavassi. 2021. "Lonomia obliqua Venom Induces NF-κB Activation and a Pro-Inflammatory Profile in THP-1-Derived Macrophage" Toxins 13, no. 7: 462. https://doi.org/10.3390/toxins13070462
APA StyleOliveira, D. S., de Souza, J. G., Alvarez-Flores, M. P., Cunegundes, P. S., DeOcesano-Pereira, C., Lobba, A. M., Gomes, R. N., & Chudzinski-Tavassi, A. M. (2021). Lonomia obliqua Venom Induces NF-κB Activation and a Pro-Inflammatory Profile in THP-1-Derived Macrophage. Toxins, 13(7), 462. https://doi.org/10.3390/toxins13070462