Effect of Different Species of Prorocentrum Genus on the Japanese Oyster Crassostrea gigas Proteomic Profile


Abstract
:1. Introduction
2. Results
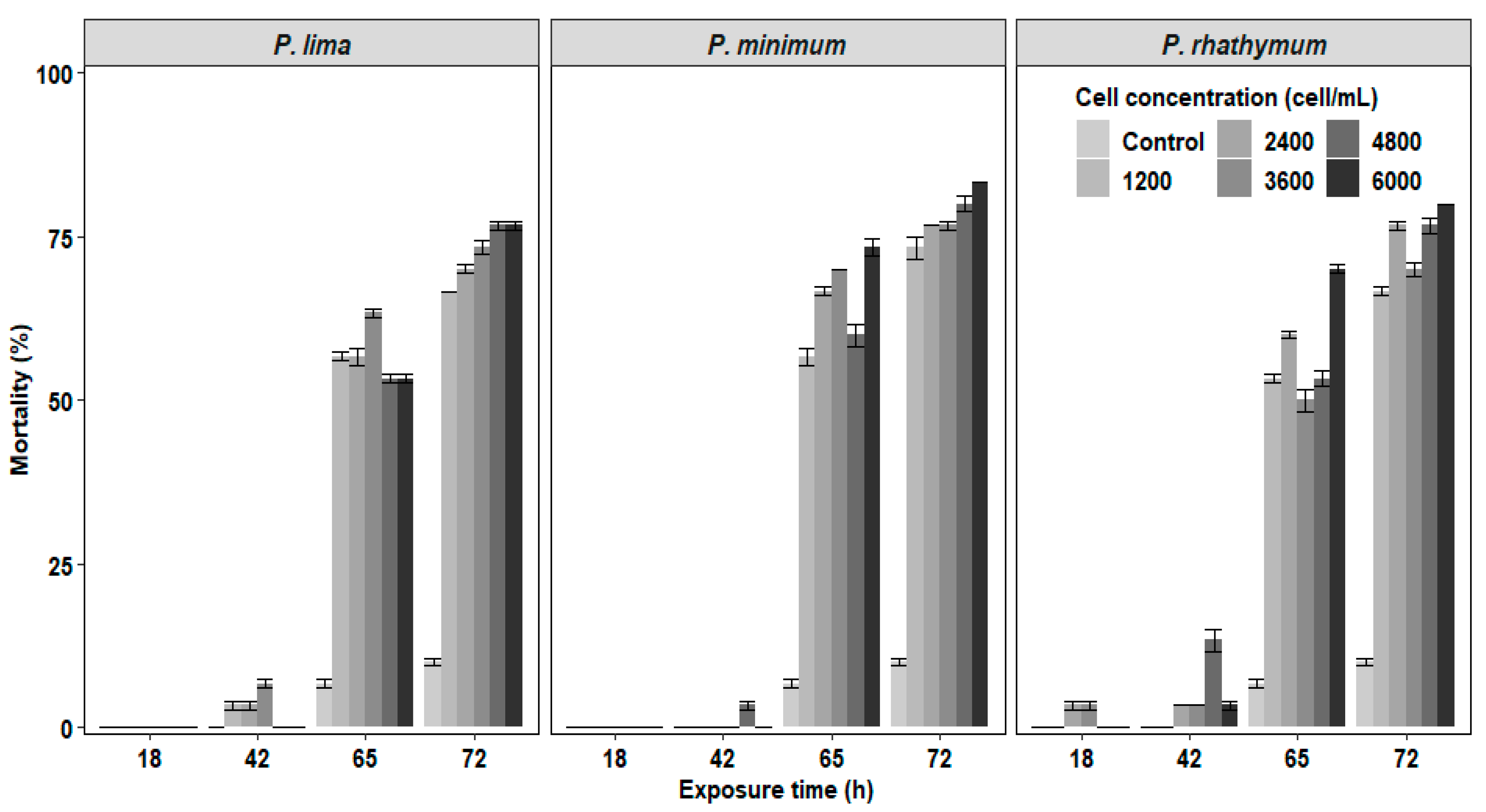
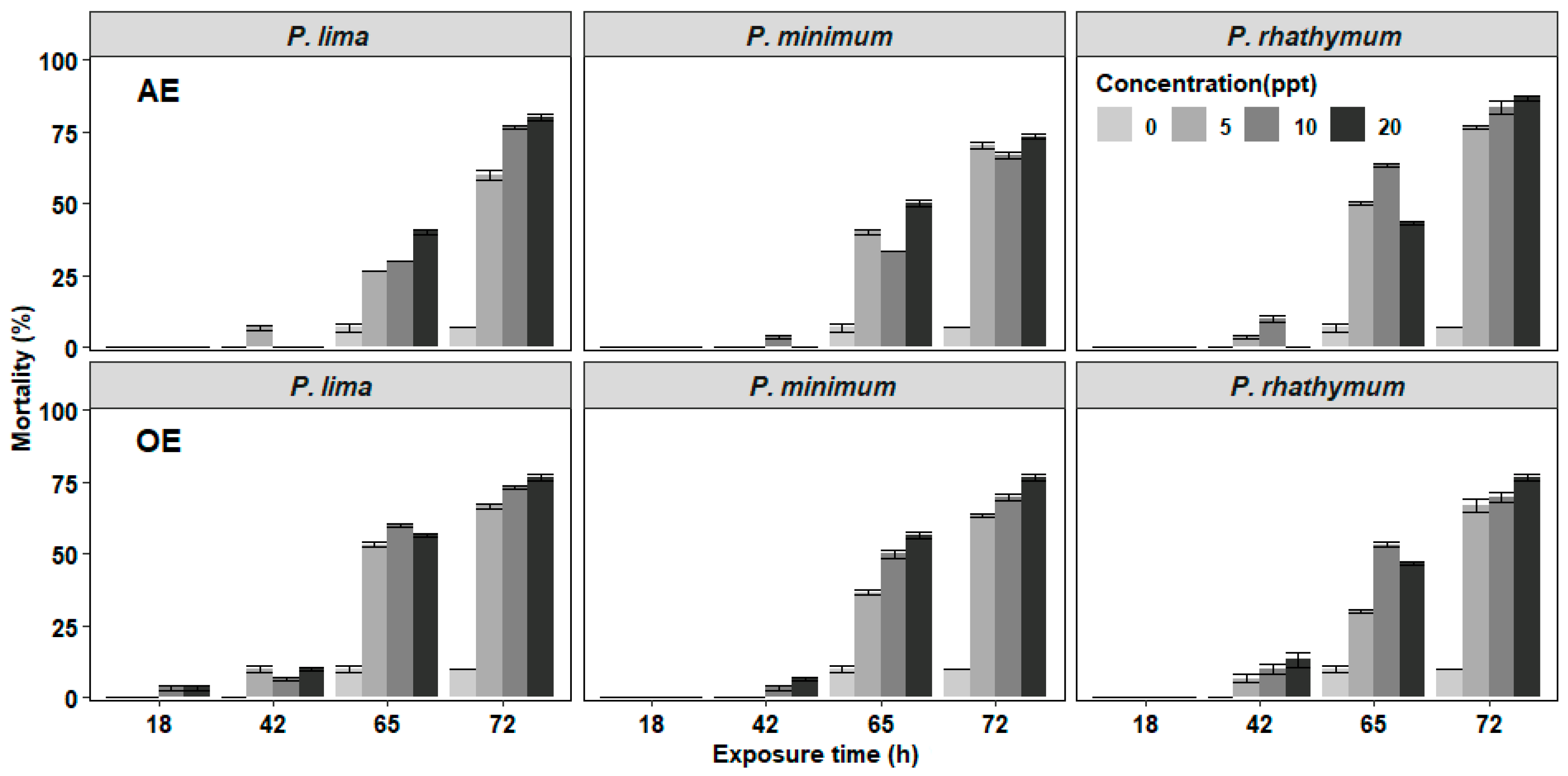
2.1. Mortality of C. gigas Exposed to Prorocentrum Complete Cells or Extracts (Time, Number, and Concentration)
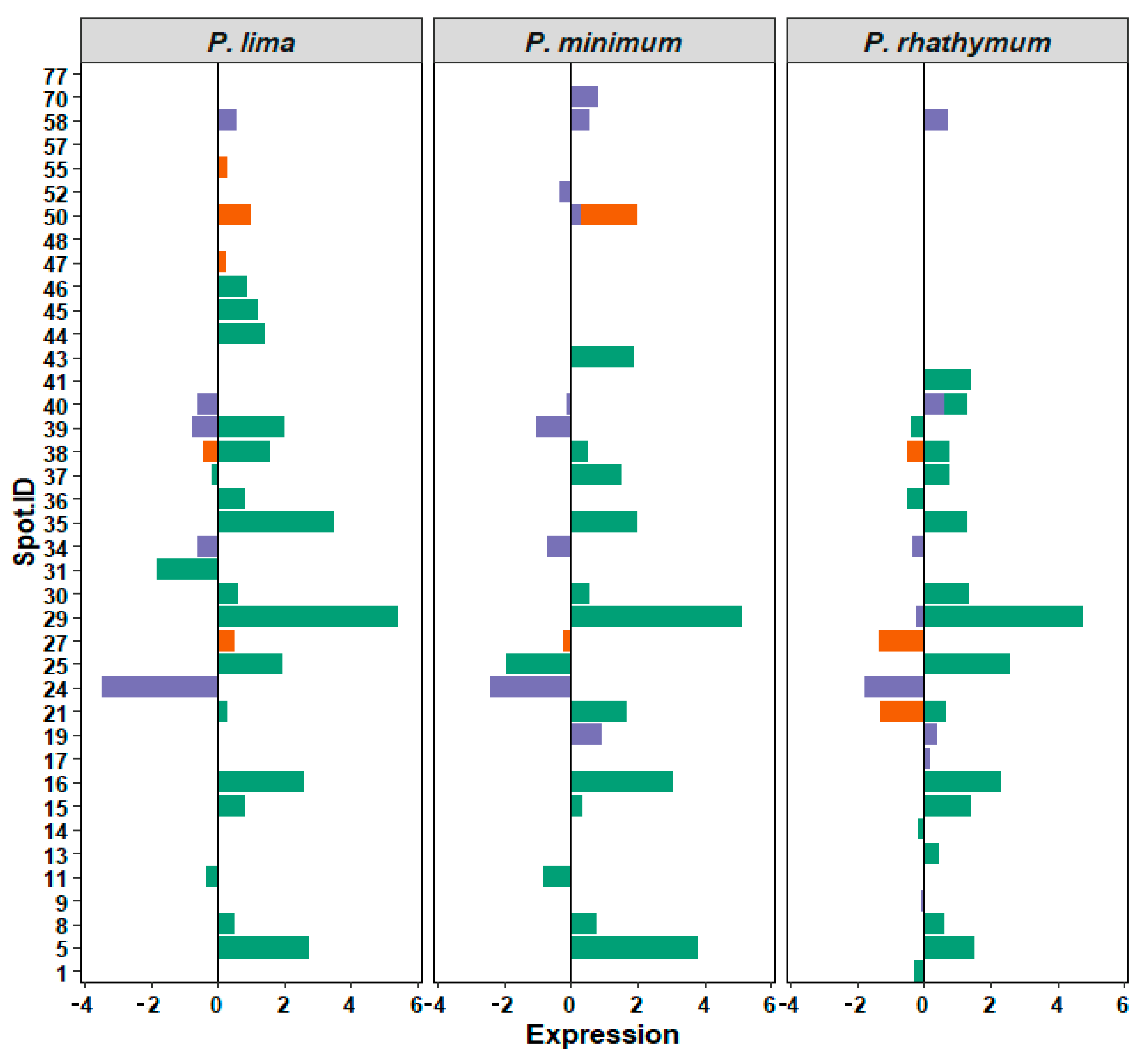
2.2. Proteomic Response
2.3. Protein Identification
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Biological Material
5.2. Extract Preparation
5.3. Exposure Experiments
5.3.1. Exposure of C. gigas to Organic (OE) and Aqueous (AE) Extracts
5.3.2. Exposure of C. gigas to Whole Live Cells of Prorocentrum spp.
5.4. Protein Extraction and Quantification
5.5. Proteomic Analysis
5.5.1. Isoelectric Focusing (IEF, First Dimension)
5.5.2. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE, Second Dimension)
5.5.3. Image Analysis and Spot Detection
5.5.4. Identification of Differentially Expressed Proteins
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, A.; Gautam, S.; Kumar, S. Phycotoxins. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 25–29. ISBN 978-0-12384-733-1. [Google Scholar]
- Rossini, G.P.; Hess, P. Phycotoxins: Chemistry, mechanisms of action and shellfish poisoning. In Molecular, Clinical and Environmental Toxicology; Luch, A., Ed.; Birkhäuser: Basel, Switzerland, 2010; Volume 100, pp. 65–122. ISBN 978-3-7643-8338-1. [Google Scholar]
- Philip, F.S.; Val, R. Beasley Phycotoxins. In Handbook of Toxicologic Pathology; Wanda, M.H., Colin, G.R., Matthew, A.W., Eds.; Academic Press: Cambridge, MA, USA, 2002; Volume 1, pp. 631–643. ISBN 978-0-12-330215-1. [Google Scholar]
- Gerssen, A.; Pol-Hofstad, I.E.; Poelman, M.; Mulder, P.P.J.; Van Den Top, H.J.; De Boer, J. Marine Toxins: Chemistry, Toxicity, Occurrence and Detection, with Special Reference to the Dutch Situation. Toxins 2010, 2, 878–904. [Google Scholar] [CrossRef] [Green Version]
- Visciano, P.; Schirone, M.; Berti, M.; Milandri, A.; Tofalo, R.; Suzzi, G. Marine Biotoxins: Occurrence, Toxicity, Regulatory Limits and Reference Methods. Front. Microbiol. 2016, 7, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D.; Guiry, G.M. Algaebase. Available online: https://www.algaebase.org (accessed on 19 July 2020).
- Hoppenrath, M.; Chomérat, N.; Horiguchi, T.; Schweikert, M.; Nagahama, Y.; Murray, S. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae)—A proposal and review. Harmful Algae 2013, 27, 1–28. [Google Scholar] [CrossRef]
- Verma, A.; Kazandjian, A.; Sarowar, C.; Harwood, D.T.; Murray, J.S.; Pargmann, I.; Hoppenrath, M.; Murray, S.A. Morphology and Phylogenetics of Benthic Prorocentrum Species (Dinophyceae) from Tropical Northwestern Australia. Toxins 2019, 11, 571. [Google Scholar] [CrossRef] [Green Version]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Paśaro, E.; Meńdez, J.; Laffon, B. Okadaic Acid: More than a diarrheic toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hégaret, H.; Wikfors, G.H.; Shumway, S.E. Diverse Feeding Responses of Five Species of Bivalve Mollusc When Exposed to Three Species of Harmful Algae. J. Shellfish Res. 2007, 26, 549–559. [Google Scholar] [CrossRef]
- Wikfors, G.H.; Smolowitz, R.M. Experimental and Histological Studies of Four Life- History Stages of the Eastern Oyster, Crassostrea virginica, Exposed to a Cultured Strain of the Dinoflagellate Prorocentrum minimum. Biol. Bull. 1995, 188, 313–328. [Google Scholar] [CrossRef]
- Galimany, E.; Sunila, I.; Hégaret, H.; Ramón, M.; Wikfors, G.H. Pathology and immune response of the blue mussel (Mytilus edulis L.) after an exposure to the harmful dinoflagellate Prorocentrum minimum. Harmful Algae 2008, 7, 630–638. [Google Scholar] [CrossRef]
- Li, Y.; Sunila, I.; Wikfors, G.H. Bioactive effects of Prorocentrum minimum on juvenile bay scallops (Argopecten irradians irradians) are dependent upon algal physiological status. Bot. Mar. 2012, 55, 19–29. [Google Scholar] [CrossRef]
- Bauder, A.; Cembella, A.; Bricelj, V.; Quilliam, M. Uptake and fate of diarrhetic shellfish poisoning toxins from the dinoflagellate Prorocentrum lima in the bay scallop Argopecten irradians. Mar. Ecol. Prog. Ser. 2001, 213, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Kameneva, P.A.; Krasheninina, E.A.; Slobodskova, V.V.; Kukla, S.P.; Orlova, T.Y. Accumulation and tissue distribution of dinophysitoxin-1 and dinophysitoxin-3 in the mussel crenomytilus grayanus feeding on the benthic dinoflagellate prorocentrum foraminosum. Mar. Drugs 2017, 15, 330. [Google Scholar] [CrossRef] [Green Version]
- Prado-Alvarez, M.; Flórez-Barrós, F.; Méndez, J.; Fernandez-Tajes, J. Effect of okadaic acid on carpet shell clam (Ruditapes decussatus) haemocytes by in vitro exposure and harmful algal bloom simulation assays. Cell Biol. Toxicol. 2013, 29, 189–197. [Google Scholar] [CrossRef]
- Huang, L.; Liu, S.L.; Zheng, J.W.; Li, H.Y.; Liu, J.S.; Yang, W.D. P-glycoprotein and its inducible expression in three bivalve species after exposure to Prorocentrum lima. Aquat. Toxicol. 2015, 169, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Dou, M.; Jiao, Y.H.; Zheng, J.W.; Zhang, G.; Li, H.Y.; Liu, J.S.; Yang, W.D. De novo transcriptome analysis of the mussel Perna viridis after exposure to the toxic dinoflagellate Prorocentrum lima. Ecotoxicol. Environ. Saf. 2020, 192, 110265. [Google Scholar] [CrossRef]
- Romero-Geraldo, R.d.J.; Hernández-Saavedra, N.Y. Stress Gene Expression in Crassostrea gigas (Thunberg, 1793) in response to experimental exposure to the toxic dinoflagellate Prorocentrum lima (Ehrenberg) Dodge, 1975. Aquac. Res. 2014, 45, 1512–1522. [Google Scholar] [CrossRef]
- Prego-Faraldo, M.V.; Martínez, L.; Méndez, J. RNA-Seq Analysis for Assessing the Early Response to DSP Toxins in Mytilus galloprovincialis Digestive Gland and Gill. Toxins 2018, 10, 417. [Google Scholar] [CrossRef] [Green Version]
- Prego-Faraldo, M.V.; Valdiglesias, V.; Laffon, B.; Mendez, J.; Eirin Lopez, J.M. Early genotoxic and cytotoxic effects of the toxic dinoflagellate Prorocentrum lima in the Mussel Mytilus galloprovincialis. Toxins 2016, 8, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, S.; Förlin, L. Intracellular effects of okadaic acid in the blue mussel Mytilus edulis, and rainbow trout Oncorhynchus mykiss. Mar. Environ. Res. 1998, 46, 449–452. [Google Scholar] [CrossRef]
- Li, A.; Li, L.; Wang, W.; Zhang, G. Acetylome Analysis Reveals Population Differentiation of the Pacific Oyster Crassostrea gigas in Response to Heat Stress. Mar. Biotechnol. 2020, 22, 233–245. [Google Scholar] [CrossRef]
- De Lorgeril, J.; Zenagui, R.; Rosa, R.D.; Piquemal, D.; Bachère, E. Whole transcriptome profiling of successful immune response to vibrio infections in the oyster crassostrea gigas by digital gene expression analysis. PLoS ONE 2011, 6, e23142. [Google Scholar] [CrossRef] [Green Version]
- De Lorgeril, J.; Petton, B.; Lucasson, A.; Perez, V.; Stenger, P.L.; Dégremont, L.; Montagnani, C.; Escoubas, J.M.; Haffner, P.; Allienne, J.F.; et al. Differential basal expression of immune genes confers Crassostrea gigas resistance to Pacific oyster mortality syndrome. BMC Genom. 2020, 21, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Tomanek, L. Environmental Proteomics: Changes in the Proteome of Marine Organisms in Response to Environmental Stress, Pollutants, Infection, Symbiosis, and Development. Ann. Rev. Mar. Sci. 2011, 3, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Olsson, B.; Bradley, B.P.; Gilek, M.; Reimer, O.; Shepard, J.L.; Tedengren, M. Physiological and proteomic responses in Mytilus edulis exposed to PCBs and PAHs extracted from Baltic Sea sediments. Hydrobiologia 2004, 514, 15–27. [Google Scholar] [CrossRef]
- Shepard, J.L.; Bradley, B.P. Protein expression signatures and lysosomal stability in Mytilus edulis exposed to graded copper concentrations. Mar. Environ. Res. 2000, 50, 457–463. [Google Scholar] [CrossRef]
- Shepard, J.L.; Olsson, B.; Tedengren, M.; Bradley, B.P. Protein expression signatures identified in Mytilus edulis exposed to PCBs, copper and salinity stress. Mar. Environ. Res. 2000, 50, 337–340. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, M.J.; Grøsvik, B.E.; Rodríguez-Ariza, A.; Goksøyr, A.; López-Barea, J. Changes in protein expression profiles in bivalve molluscs (Chamaelea gallina) exposed to four model environmental pollutants. Proteomics 2003, 3, 1535–1543. [Google Scholar] [CrossRef]
- Wei, L.; Wang, Q.; Wu, H.; Ji, C.; Zhao, J. Proteomic and metabolomic responses of Pacific oyster Crassostrea gigas to elevated pCO2 exposure. J. Proteom. 2015, 112, 83–94. [Google Scholar] [CrossRef]
- Campos, A.; Tedesco, S.; Vasconcelos, V.; Cristobal, S. Proteomic research in bivalves: Towards the identification of molecular markers of aquatic pollution. J. Proteom. 2012, 75, 4346–4359. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, D.-Z.; Wang, W.-X. Cadmium-induced changes in trace element bioaccumulation and proteomics perspective in four marine bivalves. Environ. Toxicol. Chem. 2012, 31, 1292–1300. [Google Scholar] [CrossRef]
- Meng, J.; Wang, W.; Li, L.; Yin, Q.; Zhang, G. Cadmium effects on DNA and protein metabolism in oyster (Crassostrea gigas) revealed by proteomic analyses. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef]
- Miserazzi, A.; Perrigault, M.; Sow, M.; Gelber, C.; Ciret, P.; Lomenech, A.M.; Dalens, J.M.; Weber, C.; Le Floch, S.; Lacroix, C.; et al. Proteome changes in muscles, ganglia, and gills in Corbicula fluminea clams exposed to crude oil: Relationship with behavioural disturbances. Aquat. Toxicol. 2020, 223, 105482. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, M.; Wang, W.; Liu, Z.; Xu, J.; Jia, Z.; Chen, H.; Qiu, L.; Lv, Z.; Wang, L.; et al. Transcriptional changes of Pacific oyster Crassostrea gigas reveal essential role of calcium signal pathway in response to CO2-driven acidification. Sci. Total Environ. 2020, 741, 140177. [Google Scholar] [CrossRef]
- Dineshram, R.; Quan, Q.; Sharma, R.; Chandramouli, K.; Yalamanchili, H.K.; Chu, I.; Thiyagarajan, V. Comparative and quantitative proteomics reveal the adaptive strategies of oyster larvae to ocean acidification. Proteomics 2015, 15, 4120–4134. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Wang, Q.; Yang, D.; Liu, Y.; Ran, W.; Qu, Y.; Wu, H.; Cong, M.; Li, F.; Ji, C.; et al. CO2-induced ocean acidification impairs the immune function of the Pacific oyster against Vibrio splendidus challenge: An integrated study from a cellular and proteomic perspective. Sci. Total Environ. 2018, 625, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Dineshram, R.; Wong, K.K.W.W.; Xiao, S.; Yu, Z.; Qian, P.Y.; Thiyagarajan, V. Analysis of Pacific oyster larval proteome and its response to high-CO2. Mar. Pollut. Bull. 2012, 64, 2160–2167. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zou, Y.; Weng, H.W.; Li, H.Y.; Liu, J.S.; Yang, W.D. Proteomic profile in Perna viridis after exposed to Prorocentrum lima, a dinoflagellate producing DSP toxins. Environ. Pollut. 2015, 196, 350–357. [Google Scholar] [CrossRef]
- Fernández, M.L.; Richard, D.J.A.; Cembella, A.D. In vivo assays for phycotoxins. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Enevoldsen, H.O., Eds.; UNESCO: Paris, France, 2003; pp. 297–346. ISBN 978-9-23103-871-6. [Google Scholar]
- Blanchard, M.; Pechenik, J.A.; Giudicelli, E.; Connan, J.P.; Robert, R. Competition for food in the larvae of two marine molluscs, Crepidula fornicata and Crassostrea gigas. Aquat. Living Resour. 2008, 21, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Dodge, J.D. Marine Dinoflagellates of the British Isles; Dodge, J.D., Ed.; Govt. Bookshops: London, UK, 1982; Volume 13, ISBN 011-2411-967. [Google Scholar]
- Hernández-Becerril, D.U.; Cortés Altamirano, R.; Alonso, R.R. The dinoflagellate genus Prorocentrum along the coasts of the Mexican Pacific. Hydrobiologia 2000, 418, 111–121. [Google Scholar] [CrossRef]
- De Jesuś Romero-Geraldo, R.; Garciá-Lagunas, N.; Hernańdez-Saavedra, N.Y. Effects of in vitro exposure to diarrheic toxin producer Prorocentrum lima on gene expressions related to cell cycle regulation and immune response in Crassostrea gigas. PLoS ONE 2014, 9, e97181. [Google Scholar] [CrossRef] [Green Version]
- Romero-Geraldo, R.d.J.; García-Lagunas, N.; Hernández-Saavedra, N.Y. Crassostrea gigas exposure to the dinoflagellate Prorocentrum lima: Histological and gene expression effects on the digestive gland. Mar. Environ. Res. 2016, 120, 93–102. [Google Scholar] [CrossRef]
- García-Lagunas, N.; Romero-Geraldo, R.; Kao-Godinez, A.K.; Hernández-Saavedra, N.Y. Differential expression of immune response genes in Pacific oyster, Crassostrea gigas spat, fed with dinoflagellates Gymnodinium catenatum and Prorocentrum lima. Lat. Am. J. Aquat. Res. 2019, 47, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, A.L. Efecto de la Exposición de Juveniles del Ostión Japonés Crassostrea Gigas (Thunberg, 1793) a Dinoflagelados Tóxicos Productores de Toxinas Dsp y Psp; Centro de Investigaciones Biológicas del Noroeste, S.C.: La Paz, Mexico, 2013. [Google Scholar]
- Pearce, I.; Handlinger, J.H.; Hallegraeff, G.M. Histopathology in Pacific oyster (Crassostrea gigas) spat caused by the dinoflagellate Prorocentrum rhathymum. Harmful Algae 2005, 4, 61–74. [Google Scholar] [CrossRef]
- De las Heras, S.S. Efecto de Dinoflagelados Tóxicos del Género Prorocentrum (Dinophyceae) en el Ostión Japonés Crassostrea Gigas (Thunberg, 1793); Universidad Autónoma De Baja California Sur: La Paz, Mexico, 2009. [Google Scholar]
- Landsberg, J.H. The Effects of Harmful Algal Blooms on Aquatic Organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Fernández Robledo, J.A.; Yadavalli, R.; Allam, B.; Pales Espinosa, E.; Gerdol, M.; Greco, S.; Stevick, R.J.; Gómez-Chiarri, M.; Zhang, Y.; Heil, C.A.; et al. From the raw bar to the bench: Bivalves as models for human health. Dev. Comp. Immunol. 2019, 92, 260–282. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.; Goetz, G.; White, S.; Goetz, F. Analysis of genes isolated from plated hemocytes of the Pacific oyster, Crassostreas gigas. Mar. Biotechnol. 2009, 11, 24–44. [Google Scholar] [CrossRef] [PubMed]
- Gueguen, Y.; Cadoret, J.P.; Flament, D.; Barreau-Roumiguière, C.; Girardot, A.L.; Garnier, J.; Hoareau, A.; Bachère, E.; Escoubas, J.M. Immune gene discovery by expressed sequence tags generated from hemocytes of the bacteria-challenged oyster, Crassostrea gigas. Gene 2003, 303, 139–145. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, D.; Jiang, J.; Cui, S.; Pu, H.; Jiang, S. Molecular characterization and expression analysis of cathepsin L1 cysteine protease from pearl oyster Pinctada fucata. Fish Shellfish Immunol. 2010, 29, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Qiu, L.; Liu, Z.; Wang, W.; Chen, H.; Jia, Y.; Jia, Z.; Jiang, S.; Wang, L.; Song, L. Molecular characterization of a cathepsin L1 highly expressed in phagocytes of pacific oyster Crassostrea gigas. Dev. Comp. Immunol. 2018, 89, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Venier, P.; De Pittà, C.; Pallavicini, A.; Marsano, F.; Varotto, L.; Romualdi, C.; Dondero, F.; Viarengo, A.; Lanfranchi, G. Development of mussel mRNA profiling: Can gene expression trends reveal coastal water pollution? Mutat. Res. Fundam. Mol. Mech. Mutagen. 2006, 602, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Corporeau, C.; Tamayo, D.; Pernet, F.; Quéré, C.; Madec, S. Proteomic signatures of the oyster metabolic response to herpesvirus OsHV-1 μVar infection. J. Proteom. 2014, 109, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Zhang, Q.; Kong, X.; Huang, H.; Ke, C. Differential effects of bisphenol A toxicity on oyster (Crassostrea angulata) gonads as revealed by label-free quantitative proteomics. Chemosphere 2017, 176, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Huan, P.; Wang, H.; Liu, B. Transcriptomic Analysis of the Clam Meretrix meretrix on Different Larval Stages. Mar. Biotechnol. 2012, 14, 69–78. [Google Scholar] [CrossRef]
- Payton, L.; Perrigault, M.; Hoede, C.; Massabuau, J.C.; Sow, M.; Huvet, A.; Boullot, F.; Fabioux, C.; Hegaret, H.; Tran, D. Remodeling of the cycling transcriptome of the oyster Crassostrea gigas by the harmful algae Alexandrium minutum. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Thompson, E.L.; Taylor, D.A.; Nair, S.V.; Birch, G.; Haynes, P.A.; Raftos, D.A. A proteomic analysis of the effects of metal contamination on Sydney Rock Oyster (Saccostrea glomerata) haemolymph. Aquat. Toxicol. 2011, 103, 241–249. [Google Scholar] [CrossRef]
- Thompson, E.L.; Taylor, D.A.; Nair, S.V.; Birch, G.; Haynes, P.A.; Raftos, D.A. Proteomic discovery of biomarkers of metal contamination in Sydney Rock oysters (Saccostrea glomerata). Aquat. Toxicol. 2012, 109, 202–212. [Google Scholar] [CrossRef]
- Cao, T.; Thongam, U.; Jin, J.P. Invertebrate troponin: Insights into the evolution and regulation of striated muscle contraction. Arch. Biochem. Biophys. 2019, 666, 40–45. [Google Scholar] [CrossRef]
- Nishita, K.; Tanaka, H.; Ojima, T. Amino Acid Sequence of Troponin C from Scallop Striated Adductor Muscle. J. Biol. Chem. 1994, 269, 3464–3468. [Google Scholar] [CrossRef]
- Yuasa, H.J.; Takagi, T. The genomic structure of the scallop, Patinopecten yessoensis, troponin C gene: A hypothesis for the evolution of troponin C. Gene 2000, 245, 275–281. [Google Scholar] [CrossRef]
- Umasuthan, N.; Elvitigala, D.A.S.; Revathy, K.S.; Lee, Y.; Whang, I.; Park, M.-A.; Lee, J. Identification and in silico analysis of a novel troponin C like gene from Ruditapes philippinarum (Bivalvia: Veneridae) and its transcriptional response for calcium challenge. Gene 2012. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Choi, Y.H. Regulation of adductor muscle growth by the IGF-1/AKT pathway in the triploid Pacific oyster, Crassostrea gigas. Fish. Aquat. Sci. 2019, 22, 19. [Google Scholar] [CrossRef] [Green Version]
- Funabara, D.; Ishikawa, D.; Urakawa, Y.; Kanoh, S. Ca2+-Induced Conformational Change of Troponin C from the Japanese Pearl Oyster, Pinctada fucata. Am. J. Mol. Biol. 2018, 8, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Funabara, D.; Urakawa, Y.; Kanoh, S. Molecular Cloning and Tissue Distribution of Troponin C from the Japanese Pearl Oyster, Pinctada fucata. Am. J. Mol. Biol. 2018, 8, 166–177. [Google Scholar] [CrossRef] [Green Version]
- Morquecho, L. Listado de Cepas. Available online: https://www.cibnor.gob.mx/investigacion/colecciones-biologicas/codimar/listado-de-cepas (accessed on 30 June 2021).
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Arredondo, V.B.O.; Band, C.J.S. Aislamiento Mantenimiento y Cultivo de Microalgas Nocivas. Manual de Curso de Cultivos; Centro de Investigaciones Biologicas de Noroeste: La Paz, Mexico, 2000. [Google Scholar]
- Lagos, N. Principales toxinas de origenfitoplanctónico: Identificación y cuantificaciónmediante cromatografía líquida de alta resolucion (HPLC). In Floraciones Algales Nocivas en el Cono Sur Américano; Sar, E.A., Ferrario, M.E., Reguera, B., Eds.; Instituto Español de Oceanografía: Madrid, Spain, 2002; Volume 1, pp. 57–72. ISBN 84-95877-01-5. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | P. lima | P. minimum | P. rhathymum | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exposure Time (h) | |||||||||||||
| 18 | 42 | 65 | 72 | 18 | 42 | 65 | 72 | 18 | 42 | 65 | 72 | ||
| WC (cell/mL) | Control | 0 | 0 | 6.6 ± 0.6 | 3.3 ± 0.6 | 0 | 0 | 6.6 ± 0.6 | 3.3 ± 0.6 | 0 | 0 | 6.6 ± 0.6 | 3.3 ± 0.6 |
| 1200 | 0 | 3.3 ± 0.6 | 53.3 ± 0.6 | 10.0 | 0 | 0 | 56.6 ± 1.3 | 16.6± 1.7 | 0 | 0 | 53.3 ± 0.6 | 13.3 ± 0.6 | |
| 2400 | 0 | 3.3 ± 0.6 | 53.3 ± 1.3 | 13.3 ± 0.6 | 0 | 0 | 66.6 ± 0.6 | 10.0 | 3.33 ± 0.6 | 0 | 56.6 ± 0.6 | 16.6 ± 0.6 | |
| 3600 | 0 | 6.6 ± 0.6 | 56.6 ± 0.6 | 10.0 ± 1.1 | 0 | 0 | 70.0 | 6.67 ± 0.6 | 3.33 ± 0.6 | 0 | 46.6 ± 1.7 | 20.0 ± 1.1 | |
| 4800 | 0 | 0 | 53.3 ± 0.6 | 23.3 ± 0.6 | 0 | 3.3 ± 0.6 | 56.6 ± 1.7 | 20.0 ± 1.1 | 0 | 13.3 ± 1.7 | 40.0 ± 1.1 | 23.3 ± 1.3 | |
| 6000 | 0 | 0 | 53.3 ± 0.6 | 23.3 ± 0.6 | 0 | 0 | 73.3 ± 1.3 | 10.0 | 0 | 3.3 ± 0.6 | 66.6 ± 0.6 | 10.0 | |
| AE (ppt) | Control | 0 | 0 | 6.6 ± 1.3 | 0 | 0 | 0 | 6.6 ± 1.3 | 0 | 0 | 0 | 6.6 ± 1.3 | 0 |
| 20 | 0 | 0 | 40.0 ± 1.1 | 40.0 ± 1.1 | 0 | 0 | 50.0 ± 1.1 | 23.3 ± 0.6 | 0 | 0 | 43.3 ± 0.6 | 43.3 ± 0.6 | |
| 10 | 0 | 0 | 30.0 | 46.6 ± 0.6 | 0 | 3.3 ± 0.6 | 30.0 | 33.3 ± 1.23 | 0 | 10.0 ± 1.1 | 53.3 ± 0.6 | 20.0 ± 2.2 | |
| 5 | 0 | 6.6 ± 0.6 | 20.0 | 33.3 ± 1.7 | 0 | 0 | 40.0 ± 1.1 | 10 ± 1.1 | 0 | 3.3 ± 0.6 | 46.6 ± 0.6 | 26.6 ± 0.6 | |
| OE (ppt) | Control | 0 | 0 | 10.0 ± 1.1 | 0 | 0 | 0 | 0 | 10.0 ± 1.1 | 0 | 0 | 10.0 ± 1.1 | 0 |
| 20 | 3.3 ± 0.6 | 6.6 ± 0.6 | 46.6 ± 0.6 | 20.0 ± 1.1 | 0 | 6.6 ± 3.33 | 50.0 ± 1.1 | 20.0 ± 1.1 | 0 | 13.3 ± 2.6 | 33.3 ± 0.6 | 30 ± 1.1 | |
| 10 | 3.3 ± 0.6 | 3.3 ± 0.6 | 53.3 ± 0.6 | 13.3 ± 0.6 | 0 | 3.3 ± 0.6 | 46.6 ± 1.3 | 20.0 ± 1.1 | 0 | 10.0 ± 1.9 | 43.3 ± 0.6 | 16.6± 1.7 | |
| 5 | 0 | 10.0 ± 1.1 | 43.3 ± 0.6 | 13.3 ± 0.6 | 0 | 0 | 36.6 ± 0.6 | 26.6 ± 0.6 | 0 | 6.6 ± 1.3 | 23.3 ± 0.6 | 36.6 ± 2.3 | |
| WC Treatment (434 Spots; 26 Regulated) | Species | ||
|---|---|---|---|
| Type of Regulation | P. lima | P. minimum | P. rhathymum |
| Upregulated | 14 | 10 | 13 |
| Downregulated | 3 | 5 | 4 |
| Suppressed | 3 | 5 | 3 |
| Induced | 3 | 1 | 2 |
| AE Treatment (476 spots; 7 regulated) | |||
| Upregulated | 2 | 1 | 0 |
| Downregulated | 1 | 0 | 3 |
| Suppressed | 2 | 4 | 2 |
| Induced | 2 | 1 | 0 |
| OE Treatment (623 spots; 13 regulated) | |||
| Upregulated | 1 | 3 | 3 |
| Downregulated | 4 | 6 | 4 |
| Suppressed | 6 | 3 | 3 |
| Induced | 1 | 2 | 1 |
| Spot ID | Protein | AAs 1 | Query Cover 2 | Protein Score 3 | NP 4 | Accession No 5 | Expression Type 6 | ||
|---|---|---|---|---|---|---|---|---|---|
| Pl1 | Pm 2 | Pr 3 | |||||||
| Whole Cells (WC) | |||||||||
| 11 | Toll-interacting protein isoform X4 | 281 | 14 | 62.4 | 2 | XP_011451181.1 | - | - | + |
| Dihydropteridine reductase | 237 | 15 | 60.8 | 2 | XP_011421352.1 | ||||
| 14 | Peptidase inhibitor 15-A isoform X1 | 325 | 57 | 53.4 | 4 | XP_011454703.1 | 0 | 0 | - |
| Cathepsin L1 | 330 | 27 | 43.1 | 2 | XP_011432380.1 | ||||
| Periostin | 289 | 41 | 37 | 2 | XP_011452718.1 | ||||
| 15 | Tropomyosin isoform 2 | 284 | 99 | 111 | 14 | NP_001295835.2 | + | + | + |
| 31 | Myosin essential light chain, striated adductor muscle | 157 | 94 | 62.4 | 4 | XP_011411901.1 | - | 0 | 0 |
| Aqueous Extract (AE) | |||||||||
| 27 | Actin cytoplasmic | 535 | 85 | 139 | 8 | XP_034314733.1 | + | - | - |
| Organic Extract (OE) | |||||||||
| 17 | Troponin C isoform X2 | 150 | 100 | 36.2 | 2 | XP_011429059.1 | + | + | + |
| 58 | Retrograde protein of 51 kDa isoform X5 | 581 | 16 | 67.7 | 5 | XP_019924109.1 | 1 | 1 | 1 |
| Mammalian ependymin-related protein 1 | 193 | 8 | 62 | 2 | XP_011413901.1 | ||||
| Actin-2 | 376 | 26 | 57.8 | 4 | XP_011444815.1 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matus Hernández, M.A.; Hernández Saavedra, N.Y. Effect of Different Species of Prorocentrum Genus on the Japanese Oyster Crassostrea gigas Proteomic Profile. Toxins 2021, 13, 504. https://doi.org/10.3390/toxins13070504
Matus Hernández MA, Hernández Saavedra NY. Effect of Different Species of Prorocentrum Genus on the Japanese Oyster Crassostrea gigas Proteomic Profile. Toxins. 2021; 13(7):504. https://doi.org/10.3390/toxins13070504
Chicago/Turabian StyleMatus Hernández, Miguel Angel, and Norma Yolanda Hernández Saavedra. 2021. "Effect of Different Species of Prorocentrum Genus on the Japanese Oyster Crassostrea gigas Proteomic Profile" Toxins 13, no. 7: 504. https://doi.org/10.3390/toxins13070504
APA StyleMatus Hernández, M. A., & Hernández Saavedra, N. Y. (2021). Effect of Different Species of Prorocentrum Genus on the Japanese Oyster Crassostrea gigas Proteomic Profile. Toxins, 13(7), 504. https://doi.org/10.3390/toxins13070504