Deoxynivalenol (Vomitoxin)-Induced Anorexia Is Induced by the Release of Intestinal Hormones in Mice





 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
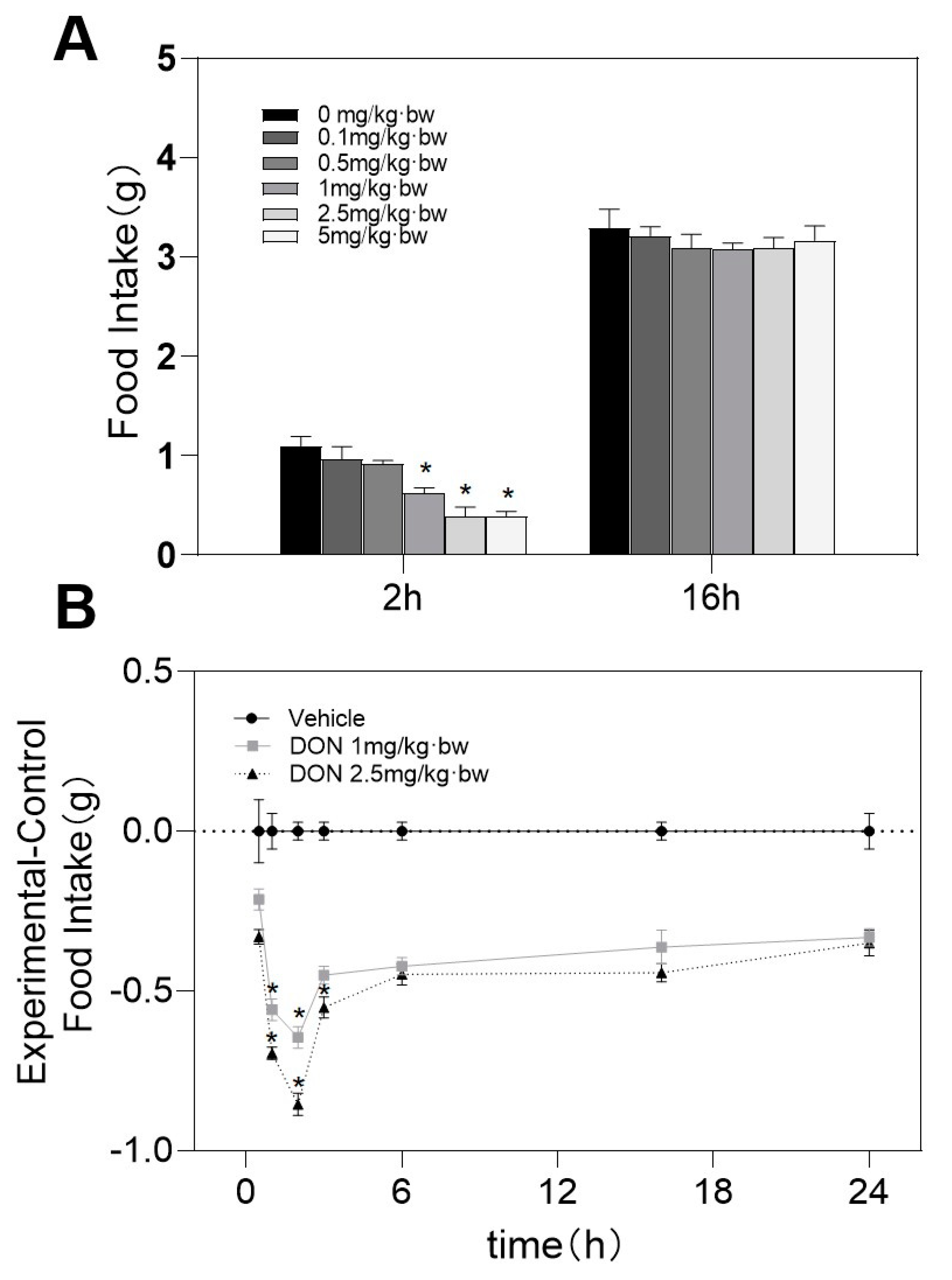
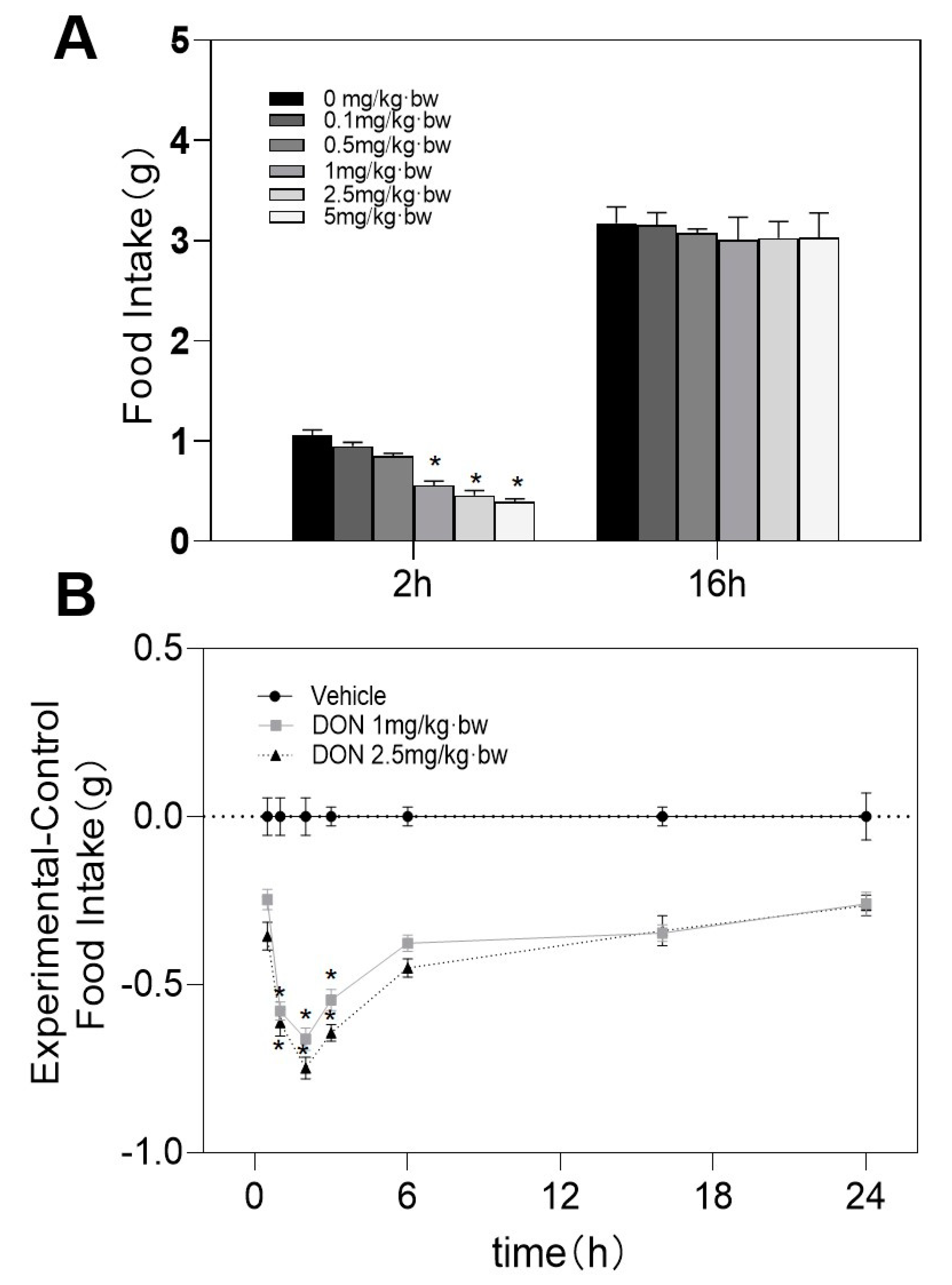
2.1. DON Induces Acute Anorectic Reaction in Mice
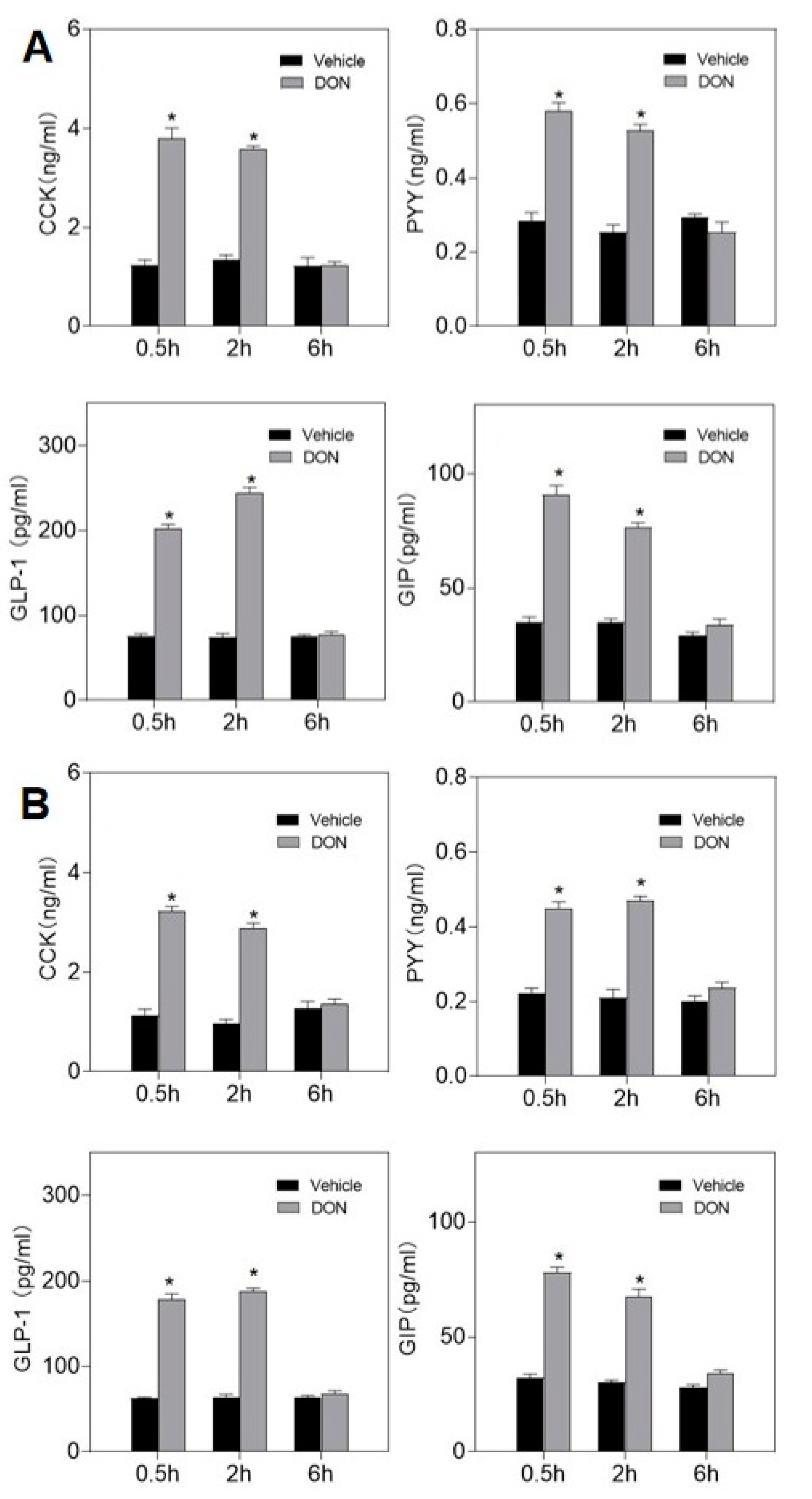
2.2. DON Evokes Plasma Intestinal Hormones Elevation
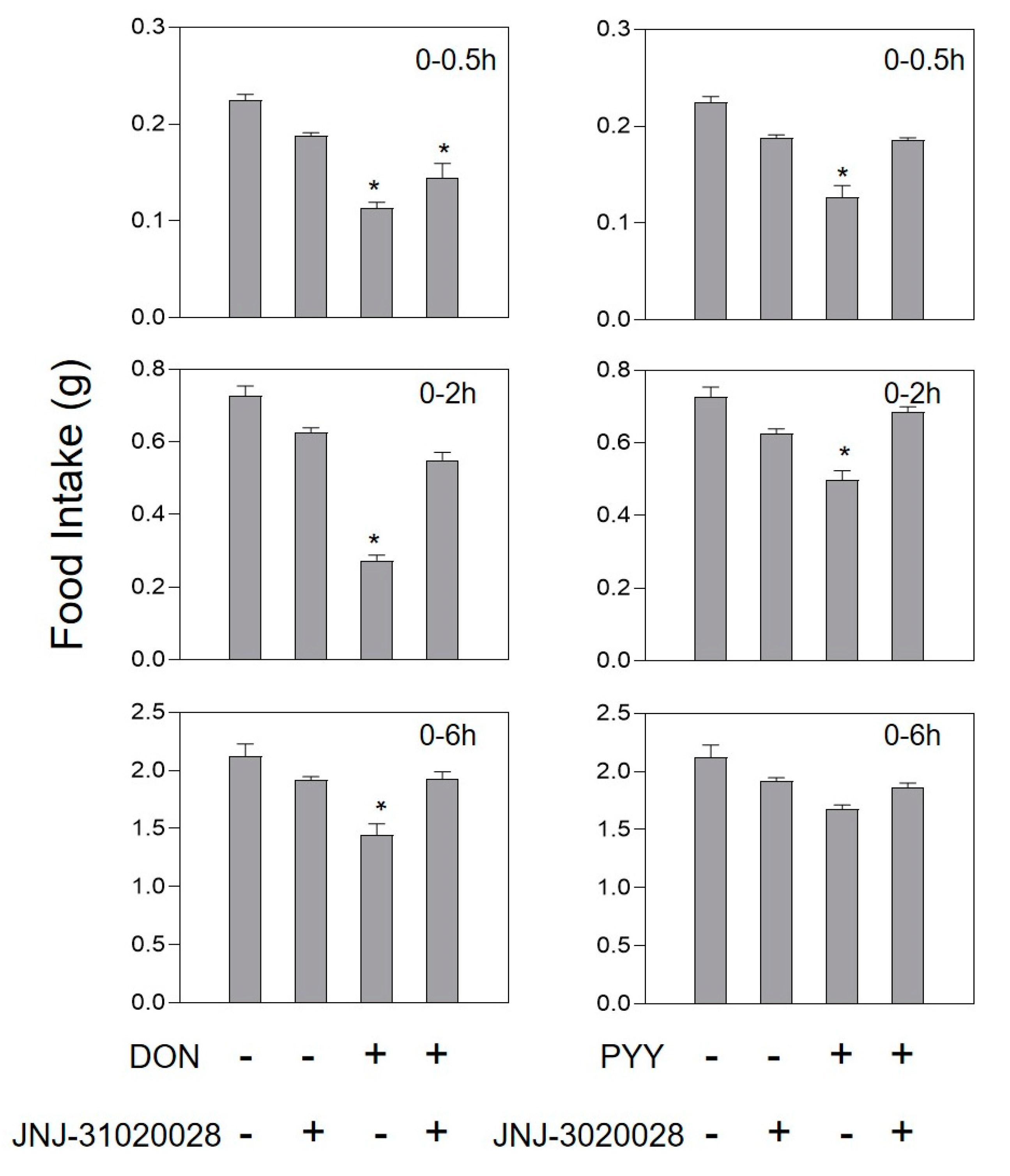
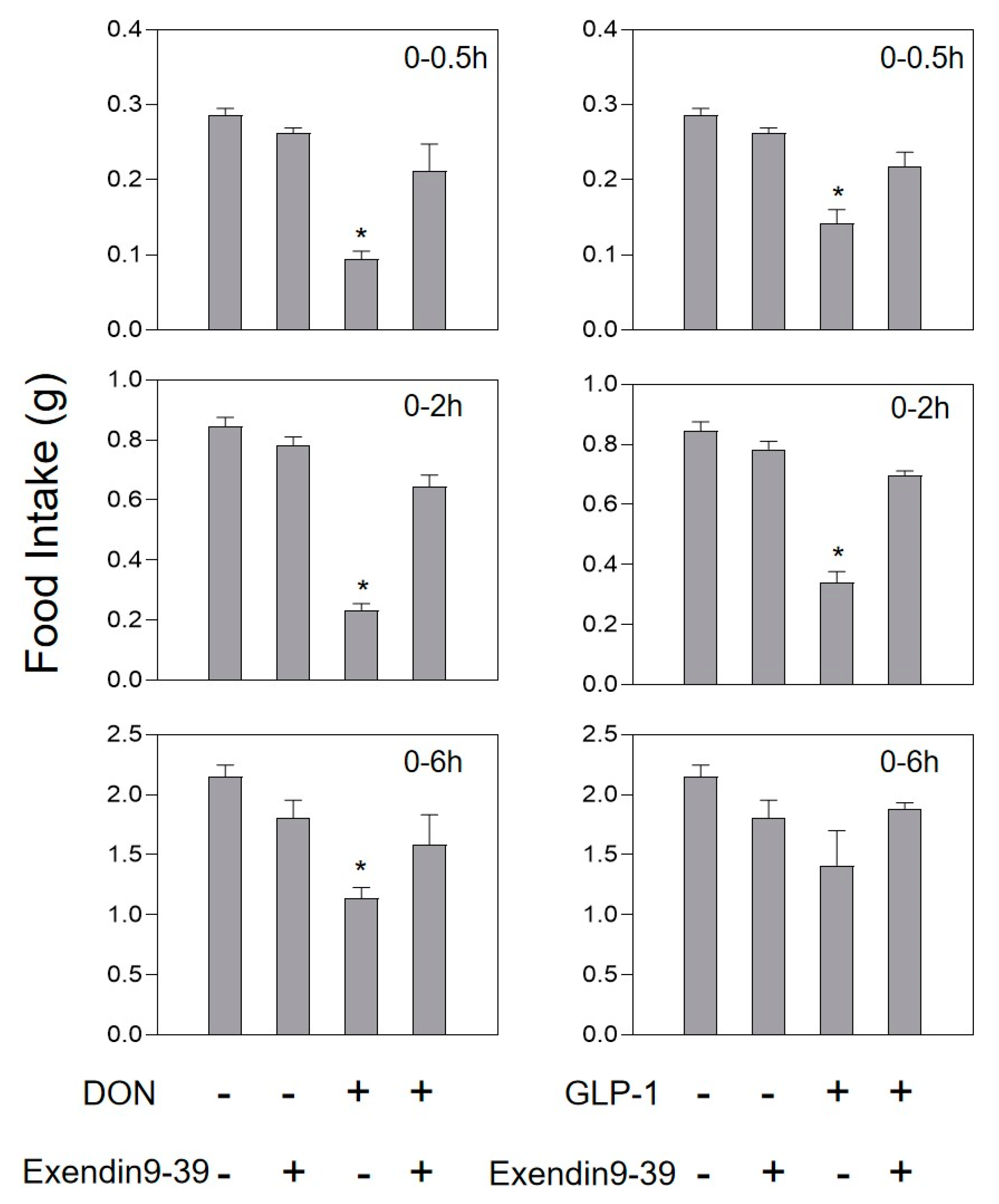
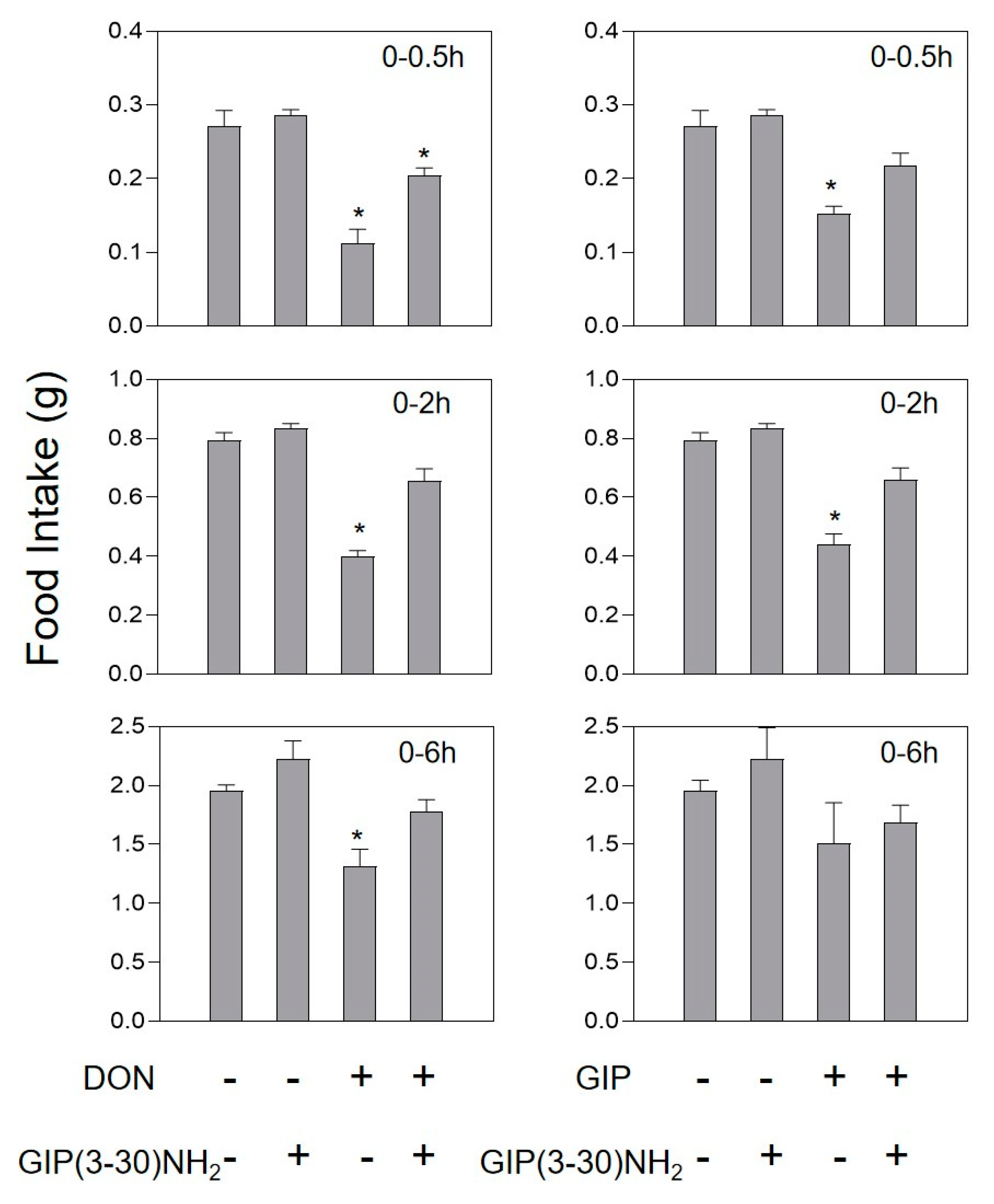
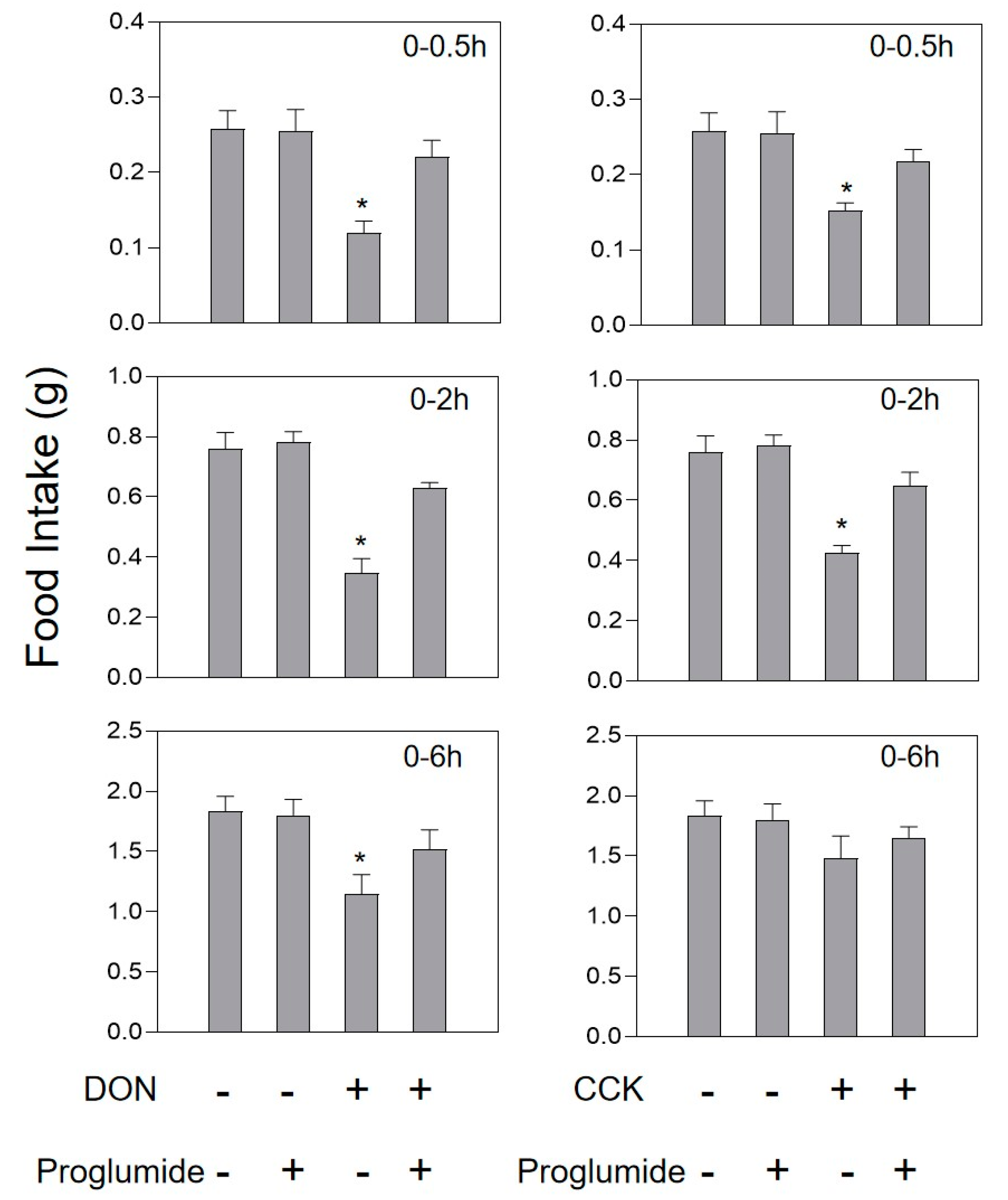
2.3. DON and Intestinal Hormone-Induced Anorexia Is Reduced by Intestinal Hormone Receptor Antagonists
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. Toxins and Chemicals
5.3. DON Induces Acute Anorexia in Mice
5.4. DON-Induced Plasma Hormone Measurement
5.5. Effect of Antagonists on Anorexia Caused by Hormones or DON
5.6. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tran, S.T.; Smith, T.K.; Girgis, G.N. A survey of free and conjugated deoxynivalenol in the 2008 corn crop in Ontario, Canada. J. Sci. Food Agric. 2012, 92, 37–41. [Google Scholar] [CrossRef] [PubMed]
- García, G.R.; Payros, D.; Pinton, P.; Dogi, C.A.; Laffitte, J.; Neves, M.; González Pereyra, M.L.; Cavaglieri, L.R.; Oswald, I.P. Intestinal toxicity of deoxynivalenol is limited by Lactobacillus rhamnosus RC007 in pig jejunum explants. Arch. Toxicol. 2018, 92, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Oswald, I.P. Effect of deoxynivalenol and other Type B trichothecenes on the intestine: A review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef] [PubMed]
- van der Fels-Klerx, H.J. Evaluation of performance of predictive models for deoxynivalenol in wheat. Risk Anal. 2014, 34, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Zielonka, Ł.; Goliński, P.; Gajęcki, M. Deoxynivalenol in the gastrointestinal tract of immature gilts under per os toxin application. Toxins 2014, 6, 973–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, M.W. Central nervous system regulation of food intake. Obesity 2006, 14, 1s–8s. [Google Scholar] [CrossRef] [PubMed]
- Sjölund, K.; Sandén, G.; Håkanson, R.; Sundler, F. Endocrine cells in human intestine: An immunocytochemical study. Gastroenterology 1983, 85, 1120–1130. [Google Scholar] [CrossRef]
- Santos-Hernández, M.; Miralles, B.; Amigo, L.; Recio, I. Intestinal Signaling of Proteins and Digestion-Derived Products Relevant to Satiety. J. Agric. Food Chem. 2018, 66, 10123–10131. [Google Scholar] [CrossRef] [Green Version]
- Atalayer, D.; Astbury, N.M. Anorexia of aging and gut hormones. Aging Dis. 2013, 4, 264–275. [Google Scholar] [CrossRef]
- Schalla, M.A.; Stengel, A. The Role of Ghrelin in Anorexia Nervosa. Int. J. Mol. Sci. 2018, 19, 2117. [Google Scholar] [CrossRef] [Green Version]
- Terciolo, C.; Maresca, M.; Pinton, P.; Oswald, I.P. Review article: Role of satiety hormones in anorexia induction by Trichothecene mycotoxins. Food Chem. Toxicol. 2018, 121, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.; Zhang, H.; Li, Y.; Wu, W.; Zhang, H. Gut satiety hormones cholecystokinin and glucagon-like Peptide-1(7-36) amide mediate anorexia induction by trichothecenes T-2 toxin, HT-2 toxin, diacetoxyscirpenol and neosolaniol. Toxicol. Appl. Pharmacol. 2017, 335, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, J.; Heng, J.; Newsholme, P.; Carlessi, R. Pleiotropic Effects of GLP-1 and Analogs on Cell Signaling, Metabolism, and Function. Front. Endocrinol. 2018, 9, 672. [Google Scholar] [CrossRef] [Green Version]
- Harding, R.K.; McDonald, T.J. Identification and characterization of the emetic effects of peptide YY. Peptides 1989, 10, 21–24. [Google Scholar] [CrossRef]
- Challis, B.G.; Pinnock, S.B.; Coll, A.P.; Carter, R.N.; Dickson, S.L.; O’Rahilly, S. Acute effects of PYY3-36 on food intake and hypothalamic neuropeptide expression in the mouse. Biochem. Biophys. Res. Commun. 2003, 311, 915–919. [Google Scholar] [CrossRef]
- Batterham, R.L.; Cowley, M.A.; Small, C.J.; Herzog, H.; Cohen, M.A.; Dakin, C.L.; Wren, A.M.; Brynes, A.E.; Low, M.J.; Ghatei, M.A.; et al. Gut hormone PYY(3-36) physiologically inhibits food intake. Nature 2002, 418, 650–654. [Google Scholar] [CrossRef]
- Kanoski, S.E.; Fortin, S.M.; Arnold, M.; Grill, H.J.; Hayes, M.R. Peripheral and central GLP-1 receptor populations mediate the anorectic effects of peripherally administered GLP-1 receptor agonists, liraglutide and exendin-4. Endocrinology 2011, 152, 3103–3112. [Google Scholar] [CrossRef] [Green Version]
- van Bloemendaal, L.; Ten Kulve, J.S.; la Fleur, S.E.; Ijzerman, R.G.; Diamant, M. Effects of glucagon-like peptide 1 on appetite and body weight: Focus on the CNS. J. Endocrinol. 2014, 221, T1–T16. [Google Scholar] [CrossRef] [Green Version]
- Seino, Y.; Yabe, D. Glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1: Incretin actions beyond the pancreas. J. Diabetes Investig. 2013, 4, 108–130. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Zhou, H.R.; Bursian, S.J.; Pan, X.; Link, J.E.; Berthiller, F.; Adam, G.; Krantis, A.; Durst, T.; Pestka, J.J. Comparison of anorectic and emetic potencies of deoxynivalenol (vomitoxin) to the plant metabolite deoxynivalenol-3-glucoside and synthetic deoxynivalenol derivatives EN139528 and EN139544. Toxicol. 2014, 142, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Wu, W.D.; Lu, X.; Zhang, J.; He, C.H.; Zhang, H.B. Role of Glucagon-Like Peptide-1 and Gastric Inhibitory Peptide in Anorexia Induction Following Oral Exposure to the Trichothecene Mycotoxin Deoxynivalenol (Vomitoxin). Toxicol. Sci. 2017, 159, 16–24. [Google Scholar] [CrossRef]
- Flannery, B.M.; Clark, E.S.; Pestka, J.J. Anorexia induction by the trichothecene deoxynivalenol (vomitoxin) is mediated by the release of the gut satiety hormone peptide YY. Toxicol. Sci. 2012, 130, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Zhou, H.R.; He, K.; Pan, X.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Role of cholecystokinin in anorexia induction following oral exposure to the 8-ketotrichothecenes deoxynivalenol, 15-acetyldeoxynivalenol, 3-acetyldeoxynivalenol, fusarenon X, and nivalenol. Toxicol. Sci. 2014, 138, 278–289. [Google Scholar] [CrossRef]
- Flannery, B.M.; Wu, W.; Pestka, J.J. Characterization of deoxynivalenol-induced anorexia using mouse bioassay. Food Chem. Toxicol. 2011, 49, 1863–1869. [Google Scholar] [CrossRef] [Green Version]
- Pestka, J.J. Deoxynivalenol-induced proinflammatory gene expression: Mechanisms and pathological sequelae. Toxins 2010, 2, 1300–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.R.; Pestka, J.J. Deoxynivalenol (Vomitoxin)-Induced Cholecystokinin and Glucagon-Like Peptide-1 Release in the STC-1 Enteroendocrine Cell Model Is Mediated by Calcium-Sensing Receptor and Transient Receptor Potential Ankyrin-1 Channel. Toxicol. Sci. 2015, 145, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Brothers, S.P.; Saldanha, S.A.; Spicer, T.P.; Cameron, M.; Mercer, B.A.; Chase, P.; McDonald, P.; Wahlestedt, C.; Hodder, P.S. Selective and brain penetrant neuropeptide y y2 receptor antagonists discovered by whole-cell high-throughput screening. Mol. Pharmacol. 2010, 77, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sloth, B.; Holst, J.J.; Flint, A.; Gregersen, N.T.; Astrup, A. Effects of PYY1-36 and PYY3-36 on appetite, energy intake, energy expenditure, glucose and fat metabolism in obese and lean subjects. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1062–E1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, S.; Sternini, C.; Raybould, H.E. Role of CCK1 and Y2 receptors in activation of hindbrain neurons induced by intragastric administration of bitter taste receptor ligands. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R33–R38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawley, J.N.; Corwin, R.L. Biological actions of cholecystokinin. Peptides 1994, 15, 731–755. [Google Scholar] [CrossRef]
- Shillabeer, G.; Davison, J.S. Proglumide, a cholecystokinin antagonist, increases gastric emptying in rats. Am. J. Physiol. 1987, 252, R353–R360. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.S.; Sparre-Ulrich, A.H.; Christensen, M.; Knop, F.K.; Hartmann, B.; Holst, J.J.; Rosenkilde, M.M. N-terminally and C-terminally truncated forms of glucose-dependent insulinotropic polypeptide are high-affinity competitive antagonists of the human GIP receptor. Br. J. Pharmacol. 2016, 173, 826–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Bates, M.A.; Bursian, S.J.; Flannery, B.; Zhou, H.-R.; Link, J.E.; Zhang, H.; Pestka, J.J. Peptide YY3–36 and 5-Hydroxytryptamine Mediate Emesis Induction by Trichothecene Deoxynivalenol (Vomitoxin). Toxicol. Sci. 2013, 133, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparre-Ulrich, A.H.; Gabe, M.N.; Gasbjerg, L.S.; Christiansen, C.B.; Svendsen, B.; Hartmann, B.; Holst, J.J.; Rosenkilde, M.M. GIP(3-30)NH(2) is a potent competitive antagonist of the GIP receptor and effectively inhibits GIP-mediated insulin, glucagon, and somatostatin release. Biochem. Pharmacol. 2017, 131, 78–88. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, J.; Guo, D.; Gao, X.; Wang, J.; Nepovimova, E.; Wu, W.; Kuca, K. Deoxynivalenol (Vomitoxin)-Induced Anorexia Is Induced by the Release of Intestinal Hormones in Mice. Toxins 2021, 13, 512. https://doi.org/10.3390/toxins13080512
Yue J, Guo D, Gao X, Wang J, Nepovimova E, Wu W, Kuca K. Deoxynivalenol (Vomitoxin)-Induced Anorexia Is Induced by the Release of Intestinal Hormones in Mice. Toxins. 2021; 13(8):512. https://doi.org/10.3390/toxins13080512
Chicago/Turabian StyleYue, Jianming, Dawei Guo, Xiuge Gao, Jiacai Wang, Eugenie Nepovimova, Wenda Wu, and Kamil Kuca. 2021. "Deoxynivalenol (Vomitoxin)-Induced Anorexia Is Induced by the Release of Intestinal Hormones in Mice" Toxins 13, no. 8: 512. https://doi.org/10.3390/toxins13080512
APA StyleYue, J., Guo, D., Gao, X., Wang, J., Nepovimova, E., Wu, W., & Kuca, K. (2021). Deoxynivalenol (Vomitoxin)-Induced Anorexia Is Induced by the Release of Intestinal Hormones in Mice. Toxins, 13(8), 512. https://doi.org/10.3390/toxins13080512