Mycotoxin Profile and Phylogeny of Pathogenic Alternaria Species Isolated from Symptomatic Tomato Plants in Lebanon

 , ,
, ,  ,
,  ,
, 

Abstract
:1. Introduction
2. Results
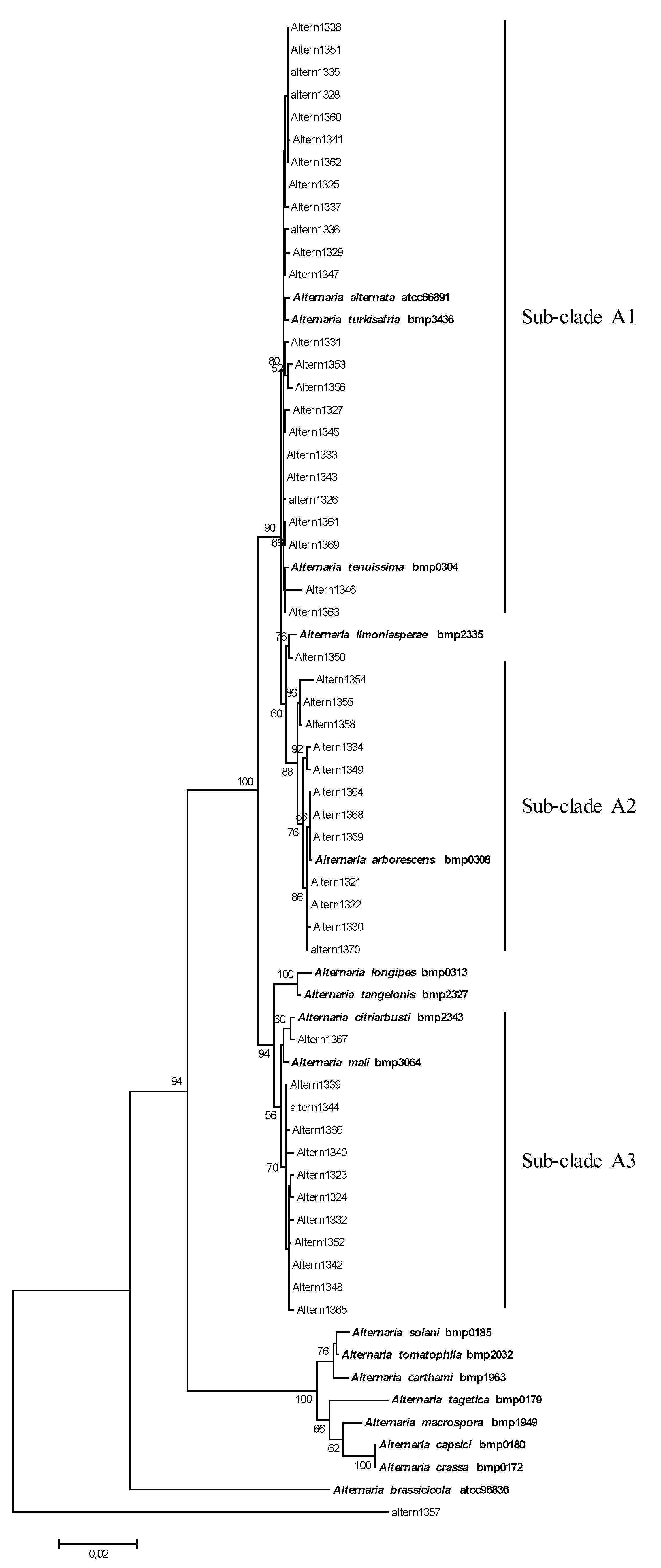
2.1. Phylogenetic Analysis
2.2. Mycotoxin Production
2.3. Pathogenicity Test
3. Discussion
4. Materials and Methods
4.1. Tomato Sampling and Fungal Isolation
4.2. DNA Extraction
4.3. PCR Amplification
4.4. Sequencing and Phylogenetic Analysis
4.5. Mycotoxin Production and Analysis
4.6. Pathogenicity Test
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization Statistics Division (FAOSTAT). Available online: www.fao.org/faostat (accessed on 13 June 2021).
- Prusky, D.; Eshel, D.; Kobiler, I.; Yakoby, N.; Beno-Moualem, D.; Ackerman, M.; Zuthji, Y.; Ben Arie, R. Postharvest chlorine treatments for the control of the persimmon black spot disease caused by Alternaria alternata. Postharvest Biol. Technol. 2001, 22, 271–277. [Google Scholar] [CrossRef]
- Smith, J.E.; Mengesha, B.; Tang, H.; Mengiste, T.; Bluhm, B.H. Resistance to Botrytis cinerea in Solanum lycopersicoides involves widespread transcriptional reprogramming. BMC Genom. 2014, 15, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derbalah, A.; Shenashen, M.; Hamza, A.; Mohamed, A.; El Safty, S. Antifungal activity of fabricated mesoporous silica nanoparticles against early blight of tomato. Egypt. J. Basic Appl. Sci. 2018, 5, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.F.; Moretti, A.; Solfrizzo, M. Alternaria toxins and plant diseases: An overview of origin, occurrence and risks. World Mycotoxin J. 2009, 2, 129–140. [Google Scholar] [CrossRef]
- Mamgain, A.; Roychowdhury, R.; Tah, J. Alternaria pathogenicity and its strategic controls. Res. J. Biol. 2013, 1, 1–9. [Google Scholar]
- Rotem, J. The Genus Alternaria: Biology, Epidemiology and Pathogenicity; APS Press: St. Paul, MN, USA, 1994. [Google Scholar]
- Pegg, G.S.; Giblin, F.R.; McTaggart, A.; Guymer, G.P.; Taylor, H.; Ireland, K.; Shivas, R.G.; Perry, S. Puccinia psidiiin Queensland, Australia: Disease symptoms, distribution and impact. Plant Pathol. 2013, 63, 1005–1021. [Google Scholar] [CrossRef] [Green Version]
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Stammler, G.; Bohme, F.; Philippi, J.; Miessner, S.; Tegge, V. Pathogenicity of Alternaria species on potatoes and tomatoes. Fourteenth Euroblight Workshop PPO Spec. Rep. 2014, 16, 85–96. [Google Scholar]
- Caldas, E.; Jones, A.D.; Ward, B.; Winter, C.K.; Gilchrist, D.G. Structural characterization of three new AAL toxins produced by Alternaria alternata f. sp. lycopersici. J. Agric. Food Chem. 1994, 42, 327–333. [Google Scholar] [CrossRef]
- Soleimani, M.; Esmailzadeh, M.; Rouhani, H. Exogenous Applications of Salicylic Acid for Inducing Systemic Acquired Resistance Against Tomato Stem Canker Disease. J. Biol. Sci. 2008, 8, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Somma, S.; Pose, G.; Pardo, A.; Mulè, G.; Pinto, V.F.; Moretti, A.; Logrieco, A.F. AFLP variability, toxin production, and pathogenicity of Alternaria species from Argentinean tomato fruits and puree. Int. J. Food Microbiol. 2011, 145, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Berbee, M.; Simmons, E.; Cardoso, C.; Reis, A.; Maffia, L.; Mizubuti, E. First report of Alternaria tomatophila and A. grandis causing early blight on tomato and potato in Brazil. New Dis. Rep. 2010, 22, 28. [Google Scholar] [CrossRef] [Green Version]
- Bessadat, N.; Berruyer, R.; Hamon, B.; Kihal, M.; Henni, D.E.; Simoneau, P.; Bataille-Simoneau, N.; Benichou, S. Alternaria species associated with early blight epidemics on tomato and other Solanaceae crops in northwestern Algeria. Eur. J. Plant Pathol. 2016, 148, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Kokaeva, L.Y.; Belosokhov, A.F.; Doeva, L.Y.; Skolotneva, E.S.; Elansky, S.N. Distribution of Alternaria species on blighted potato and tomato leaves in Russia. J. Plant Dis. Prot. 2017, 125, 1–8. [Google Scholar] [CrossRef]
- Turkensteen, J.; Spoelder, J.; Mulder, A. Will the real Alternaria stand up please—Experiences with Alternaria-like diseases on potatoes during the 2009 season in the Netherlands. In PPO-Special Report no. 14. DLO Foundation; Schepers, H.T.A.M., Ed.; DLO Foundation: Wageningen, The Netherlands, 2010; pp. 165–170. [Google Scholar]
- Simmons, E.G. Alternaria taxonomy: Current status, viewpoint, challenge. In Alternaria Biology, Plant Diseases and Metabolites; Chelkowski, J., Visconti, A., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1992; pp. 1–35. [Google Scholar]
- Simmons, E.G. Alternaria: An. Identification Manual; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Lawrence, D.P.; Rotondo, F.; Gannibal, P.B. Biodiversity and taxonomy of the pleomorphic genus Alternaria. Mycol. Prog. 2015, 15, 1–22. [Google Scholar] [CrossRef]
- Somma, S.; Amatulli, M.T.; Masiello, M.; Moretti, A.; Logrieco, A.F. Alternaria species associated to wheat black point identified through a multilocus sequence approach. Int. J. Food Microbiol. 2019, 293, 34–43. [Google Scholar] [CrossRef]
- Woudenberg, J.; Seidl, M.F.; Groenewald, J.; de Vries, M.; Stielow, J.; Thomma, B.; Crous, P. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, D.P.; Gannibal, P.B.; Dugan, F.M.; Pryor, B.M. Characterization of Alternaria isolates from the infectoria species-group and a new taxon from Arrhenatherum, Pseudoalternaria arrhenatheria sp. nov. Mycol. Prog. 2013, 13, 257–276. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.; Peever, T.L.; Pryor, B.M. The sections of Alternaria: Formalizing species-group concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.P.; Park, M.S.; Pryor, B.M. Nimbya and Embellisia revisited, with nov. comb for Alternaria celosiae and A. perpunctulata. Mycol. Prog. 2011, 11, 799–815. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [Green Version]
- Al Ghafri, A.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Al-Saady, N.A.; Al-Sadi, A.M. A new section and a new species of Alternaria encountered from Oman. Phytotaxa 2019, 405, 279–289. [Google Scholar] [CrossRef]
- Patriarca, A.; Cabral, L.D.C.; Pavicich, M.A.; Nielsen, K.F.; Andersen, B. Secondary metabolite profiles of small-spored Alternaria support the new phylogenetic organization of the genus. Int. J. Food Microbiol. 2019, 291, 135–143. [Google Scholar] [CrossRef]
- Hasan, H.A.H. Alternaria toxins in black rot lesion of tomato fruit: Conditions and regulation of their production. Mycopathologia 1995, 130, 171–177. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 2nd ed.; Blackie Academic and Professional: London, UK, 1997. [Google Scholar]
- Van de Perre, E.; Deschuyffeleer, N.; Jacxsens, L.; Vekeman, F.; Van Der Hauwaert, W.; Asam, S.; Rychlik, M.; Devlieghere, F.; De Meulenaer, B. Screening of moulds and mycotoxins in tomatoes, bell peppers, onions, soft red fruits and derived tomato products. Food Control 2014, 37, 165–170. [Google Scholar] [CrossRef]
- Rosenzweig, C.I.A.; Yang, X.; Epstein, P.R.; Chivian, E. Climate change and extreme weather events: Implications for food production, plant diseases and pests. Glob. Chang. Hum. Health 2001, 2, 90–104. [Google Scholar] [CrossRef]
- Magan, N.; Medina-Vaya, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre- and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Wang, W.; Jones, C.; Ciacci-Zanella, J.; Holt, T.; Gilchrist, D.G.; Dickman, M.B. Fumonisins and Alternaria alternata lycopersici toxins: Sphinganine analog mycotoxinsinduce apoptosis in monkey kidney cells. Proc. Natl. Acad. Sci. USA 1996, 93, 3461–3465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomma, B.P.H.J. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant. Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Frisvad, J. Natural Occurrence of Fungi and Fungal Metabolites in Moldy Tomatoes. J. Agric. Food Chem. 2004, 52, 7507–7513. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Bottalico, A.; Mulè, G.; Moretti, A.; Perrone, G. Epidemiology of Toxigenic Fungi and Their Associated Mycotoxins for Some Mediterranean Crops; Springer: Dordrecht, The Netherlands, 2003; pp. 645–667. [Google Scholar] [CrossRef]
- Yamagishi, D.; Akamatsu, H.; Otani, H.; Kodama, M. Pathological evaluation of host-specific AAL-toxins and fumonisin mycotoxins produced by Alternaria and Fusarium species. J. Gen. Plant. Pathol. 2006, 72, 323–327. [Google Scholar] [CrossRef]
- Yekeler, H.; Bitmiş, K.; Ozçelik, N.; Doymaz, M.Z.; Çalta, M. Analysis of Toxic Effects of Alternaria Toxins on Esophagus of Mice by Light and Electron Microscopy. Toxicol. Pathol. 2001, 29, 492–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asam, S.; Habler, K.; Rychlik, M. Determination of tenuazonic acid in human urine by means of a stable isotope dilution assay. Anal. Bioanal. Chem. 2013, 405, 4149–4158. [Google Scholar] [CrossRef] [Green Version]
- Ostrý, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Solhaug, A.; Wisbech, C.; Christoffersen, T.E.; Hult, L.; Lea, T.; Eriksen, G.S.; Holme, J.A. The mycotoxin alternariol induces DNA damage and modify macrophage phenotype and inflammatory responses. Toxicol. Lett. 2015, 239, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Huybrechts, I.; De Ruyck, K.; De Saeger, S.; De Boevre, M. Uniting large-scale databeses to unravel the impact of chronic multi-mycotoxins exposures on colorectal cancer incidence in Europe. In Proceedings of the 2nd MycoKey International Conference, Wuhan, China, 16–18 September 2018; China Agricultural Science and Technology Press: Beijing, China, 2018; pp. 181–183. [Google Scholar]
- Patriarca, A. Alternaria in food products. Curr. Opin. Food Sci. 2016, 11, 1–9. [Google Scholar] [CrossRef]
- Cabral, L.D.C.; Terminiello, L.; Pinto, V.F.; Nielsen, K.F.; Patriarca, A. Natural occurrence of mycotoxins and toxigenic capacity of Alternaria strains from mouldy peppers. Int. J. Food Microbiol. 2016, 236, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Laemmlen, F. Alternaria Diseases. University of California Cooperative Extension Farm Advisor, Santa Barbara and San Luis Obispo Counties. 2001. Available online: https://anrcatalog.ucanr.edu/pdf/8040.pdf. (accessed on 15 October 2019).
- Nash, A.F.; Gardner, R.G. Tomato early blight resistance in a breeding line derived from Lycopersicon hirsutum PI 126445. Plant Dis. 1988, 72, 206–209. [Google Scholar] [CrossRef]
- Hassanein, N.; Zeid, M.; Youssef, K.; Mahmoud, D. Control of tomato early blight and wilt using aqueous extract of neem leaves. Phytopathol. Mediterr. 2010, 49, 143–151. [Google Scholar]
- Singh, H.K.; Srivastava, S.; Singh, R.B.; Singh, A.K. Management of Alternaria blight of rapeseed-mustard. J. Plant Dis. Prot. 2013, 8, 131–136. [Google Scholar]
- Andersen, B.; Sørensen, J.L.; Nielsen, K.F.; Ende, B.G.V.D.; De Hoog, S. A polyphasic approach to the taxonomy of the Alternaria infectoria species–group. Fungal Genet. Biol. 2009, 46, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Zwickel, T.; Kahl, S.M.; Rychlik, M.; Müller, M.E.H. Chemotaxonomy of Mycotoxigenic Small-Spored Alternaria Fungi—Do Multitoxin Mixtures Act as an Indicator for Species Differentiation? Front. Microbiol. 2018, 9, 1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Gobashy, S.F.; Mikhail, W.; Ismail, A.M.; Zekry, A.; Moretti, A.; Susca, A.; Soliman, A.S. Phylogenetic, toxigenic and virulence profiles of Alternaria species causing leaf blight of tomato in Egypt. Mycol. Prog. 2018, 17, 1269–1282. [Google Scholar] [CrossRef]
- Ramires, F.A.; Masiello, M.; Somma, S.; Villani, A.; Susca, A.; Logrieco, A.F.; Luz, C.; Meca, G.; Moretti, A. Phylogeny and Mycotoxin Characterization of Alternaria Species Isolated from Wheat Grown in Tuscany, Italy. Toxins 2018, 10, 472. [Google Scholar] [CrossRef] [Green Version]
- Masiello, M.; Somma, S.; Susca, A.; Ghionna, V.; Logrieco, A.F.; Franzoni, M.; Ravaglia, S.; Meca, G.; Moretti, A. Molecular Identification and Mycotoxin Production by Alternaria Species Occurring on Durum Wheat, Showing Black Point Symptoms. Toxins 2020, 12, 275. [Google Scholar] [CrossRef] [Green Version]
- Chalbi, A.; Sghaier-Hammami, B.; Meca, G.; Quiles, J.M.; Abdelly, C.; Marangi, C.; Logrieco, A.F.; Moretti, A.; Masiello, M. Characterization of mycotoxigenic Alternaria species isolated from the Tunisian halophyte Cakile maritima. Phytopathol. Mediterr. 2020, 59, 107–118. [Google Scholar] [CrossRef]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S.T.; Ippolito, A.; Sanzani, S.M.; Fischer, R. Alternariol as virulence and colonization factor of Alternaria alternata during plant infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef]
- Graf, E.; Schmidt-Heydt, M.; Geisen, R. HOG MAP kinase regulation of alternariol biosynthesis in Alternaria alternata is important for substrate colonization. Int. J. Food Microbiol. 2012, 157, 353–359. [Google Scholar] [CrossRef]
- Wojciechowska, E.; Weinert, C.; Egert, B.; Trierweiler, B.; Schmidt-Heydt, M.; Horneburg, B.; Graeff-Hönninger, S.; Kulling, S.E.; Geisen, R. Chlorogenic acid, a metabolite identified by untargeted metabolome analysis in resistant tomatoes, inhibits the colonization by Alternaria alternata by inhibiting alternariol biosynthesis. Eur. J. Plant Pathol. 2014, 139, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Soukup, S.T.; Kohn, B.N.; Pfeiffer, E.; Geisen, R.; Metzler, M.; Bunzel, M.; Kulling, S.E. Sulfoglucosides as Novel Modified Forms of the Mycotoxins Alternariol and Alternariol Monomethyl Ether. J. Agric. Food Chem. 2016, 64, 8892–8901. [Google Scholar] [CrossRef] [Green Version]
- Garganese, F.; Schena, L.; Siciliano, I.; Prigigallo, M.I.; Spadaro, D.; De Grassi, A.; Ippolito, A.; Sanzani, S.M. Characterization of Citrus-Associated Alternaria Species in Mediterranean Areas. PLoS ONE 2016, 11, e0163255. [Google Scholar] [CrossRef] [Green Version]
- Crudo, F.; Varga, E.; Aichinger, G.; Galaverna, G.; Marko, D.; Dall’Asta, C.; Dellafiora, L. Co-Occurrence and Combinatory Effects of Alternaria Mycotoxins and other Xenobiotics of Food Origin: Current Scenario and Future Perspectives. Toxins 2019, 11, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, E.G.; Roberts, R. Alternaria themes and variations (73). Mycotaxon 1993, 48, 109–140. [Google Scholar]
- Murray, M.; Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Li, F.-Q.; Yoshizawa, T.; Kawamura, O.; Luo, X.-Y.; Li, Y.-W. Aflatoxins and Fumonisins in Corn from the High-Incidence Area for Human Hepatocellular Carcinoma in Guangxi, China. J. Agric. Food Chem. 2001, 49, 4122–4126. [Google Scholar] [CrossRef]
- Rubert, J.; Dzuman, Z.; Vaclavikova, M.; Zachariasova, M.; Soler, C.; Hajslova, J. Analysis of mycotoxins in barley using ultra high liquid chromatography high resolution mass spectrometry: Comparison of efficiency and efficacy of different extraction procedures. Talanta 2012, 99, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Myresiotis, C.K.; Testempasis, S.; Vryzas, Z.; Karaoglanidis, G.S.; Papadopoulou-Mourkidou, E. Determination of mycotoxins in pomegranate fruits and juices using a QuEChERS-based method. Food Chem. 2015, 182, 81–88. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Strain | Alternaria Species | Mycotoxin (mg·kg−1) | |||
|---|---|---|---|---|---|
| AOH | AME | ALT | TA | ||
| Altern1325 | A. alternata | 26 | 20 | 1 | 4442 |
| Altern1327 * | A. alternata | 908 | 6900 | 1538 | 2658 |
| Altern1328 | A. alternata | 4567 | 4967 | 1025 | Nd ** |
| Altern1329 * | A. alternata | 1271 | 4967 | 558 | Nd |
| Altern1331 | A. alternata | 102 | 516 | 462 | 603 |
| Altern1333 | A. alternata | 84 | 82 | 1959 | 1331 |
| Altern1336 | A. alternata | 126 | 855 | 74 | 532 |
| Altern1337 | A. alternata | 676 | 632 | 4097 | Nd |
| Altern1341 * | A. alternata | 4002 | 9172 | 214 | 3938 |
| Altern1343 * | A. alternata | 131 | 660 | 2687 | Nd |
| Altern1345 | A. alternata | 1646 | 11,021 | 394 | Nd |
| Altern1351 | A. alternata | 3090 | 8678 | 1114 | 145 |
| Altern1353 | A. alternata | 441 | 2718 | 1238 | 2066 |
| Altern1360 | A. alternata | Nd | Nd | Nd | 2035 |
| Altern1362 | A. alternata | 3062 | 15,070 | 5156 | 1155 |
| Altern1346 | A. alternata | 1199 | 2385 | 1950 | Nd |
| Mean value | 1422 | 4576 | 1498 | 1890 | |
| Min | 26 | 20 | 1 | 145 | |
| Max | 4567 | 15,070 | 5156 | 4442 | |
| Altern1350 | A. limoniasperae | Nd | 102.51 | 3.86 | Nd |
| Altern1321 | A. arborescens | 27 | 25 | 42 | 2083 |
| Altern1322 | A. arborescens | 333 | 249 | 134 | 283 |
| Altern1330 * | A. arborescens | 2059 | 6395 | 570 | Nd |
| Altern1334 | A. arborescens | 610 | 405 | 387 | 2713 |
| Altern1349 | A. arborescens | 5634 | 16,006 | 735 | 1251 |
| Altern1354 | A. arborescens | 3501 | Nd | Nd | Nd |
| Altern1355 * | A. arborescens | 4612 | 5737 | 597 | 1008 |
| Altern1358 * | A. arborescens | 264 | 142 | 28 | 456 |
| Altern1359 | A. arborescens | 1786 | 3026 | 550 | 754 |
| Altern1364 | A. arborescens | 3271 | 6662 | 1560 | 496 |
| Altern1368 | A. arborescens | 69 | 860 | 328 | 75 |
| Altern1370 | A. arborescens | 2731 | 12,868 | 894 | 3777 |
| Mean value | 2075 | 4761 | 529 | 1290 | |
| Min | 27 | 25 | 28 | 75 | |
| Max | 5634 | 16,006 | 1560 | 3777 | |
| Altern1323 * | A. mali | 92 | 23 | 286 | 4507 |
| Altern1332 | A. mali | 254 | 10,182 | 1153 | 649 |
| Altern1340 * | A. mali | 384 | 893 | 2046 | 1896 |
| Altern1342 | A. mali | 2367 | 10,998 | 2722 | Nd |
| Altern1344 | A. mali | 796 | 6593 | Nd | Nd |
| Altern1352 * | A. mali | 915 | 7111 | 1326 | 452 |
| Mean value | 802 | 5967 | 1507 | 1876 | |
| Min | 92 | 23 | 286 | 452 | |
| Max | 2367 | 10,998 | 2722 | 4507 | |
| Mycotoxin | LOD (mg/kg) | LOQ (mg/kg) | Recovery (%) | ME (%) |
|---|---|---|---|---|
| AOH | 0.01 | 0.1 | 71 | 79 |
| AME | 0.01 | 0.1 | 70 | 78 |
| ALT | 0.01 | 0.1 | 62 | 75 |
| TA | 0.01 | 0.1 | 65 | 73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habib, W.; Masiello, M.; El Ghorayeb, R.; Gerges, E.; Susca, A.; Meca, G.; Quiles, J.M.; Logrieco, A.F.; Moretti, A. Mycotoxin Profile and Phylogeny of Pathogenic Alternaria Species Isolated from Symptomatic Tomato Plants in Lebanon. Toxins 2021, 13, 513. https://doi.org/10.3390/toxins13080513
Habib W, Masiello M, El Ghorayeb R, Gerges E, Susca A, Meca G, Quiles JM, Logrieco AF, Moretti A. Mycotoxin Profile and Phylogeny of Pathogenic Alternaria Species Isolated from Symptomatic Tomato Plants in Lebanon. Toxins. 2021; 13(8):513. https://doi.org/10.3390/toxins13080513
Chicago/Turabian StyleHabib, Wassim, Mario Masiello, Romy El Ghorayeb, Elvis Gerges, Antonia Susca, Giuseppe Meca, Juan M. Quiles, Antonio F. Logrieco, and Antonio Moretti. 2021. "Mycotoxin Profile and Phylogeny of Pathogenic Alternaria Species Isolated from Symptomatic Tomato Plants in Lebanon" Toxins 13, no. 8: 513. https://doi.org/10.3390/toxins13080513
APA StyleHabib, W., Masiello, M., El Ghorayeb, R., Gerges, E., Susca, A., Meca, G., Quiles, J. M., Logrieco, A. F., & Moretti, A. (2021). Mycotoxin Profile and Phylogeny of Pathogenic Alternaria Species Isolated from Symptomatic Tomato Plants in Lebanon. Toxins, 13(8), 513. https://doi.org/10.3390/toxins13080513