Development of a Broad-Spectrum Antiserum against Cobra Venoms Using Recombinant Three-Finger Toxins


Abstract
:1. Introduction
2. Results
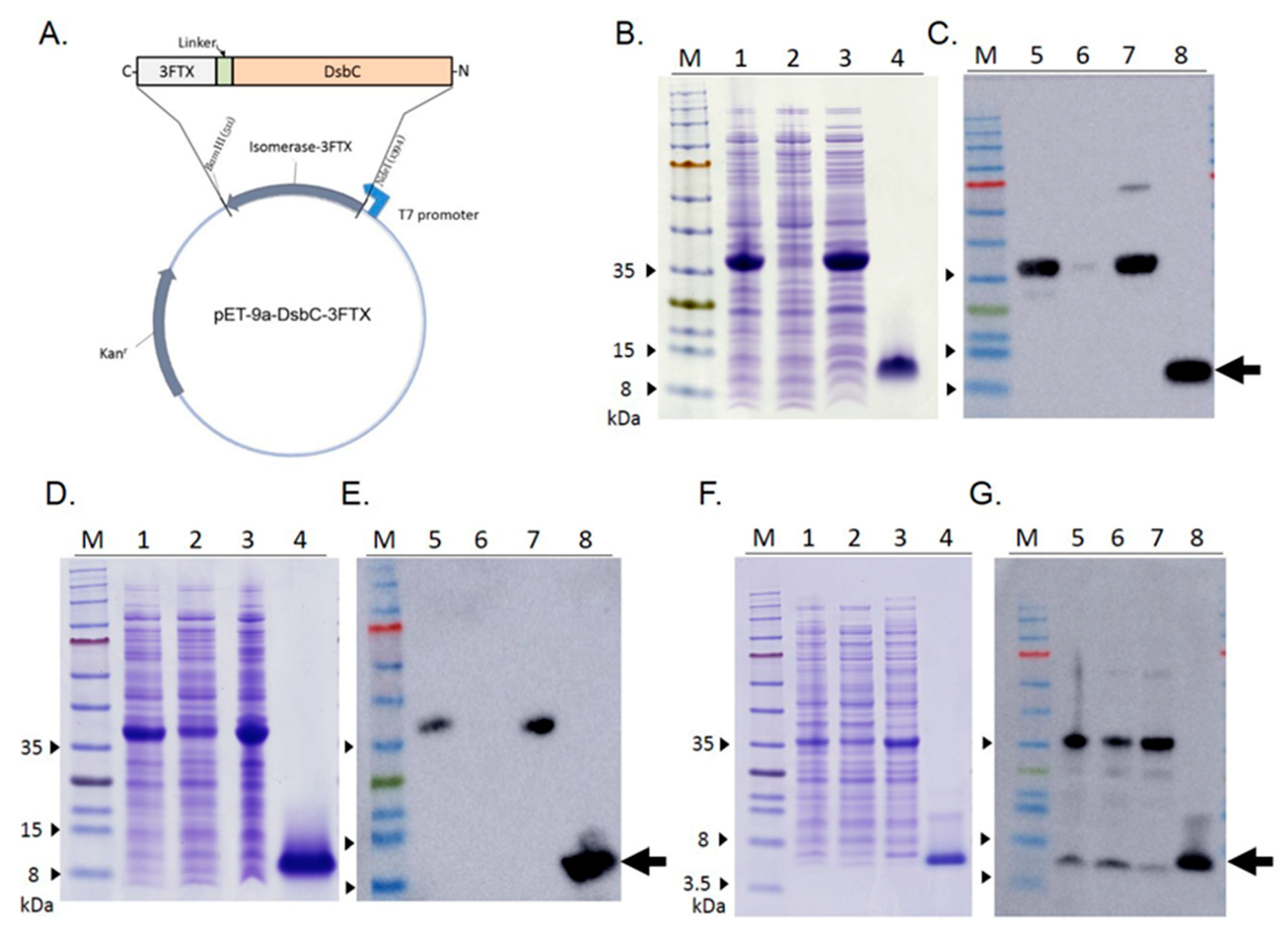
2.1. Expression of Recombinant Three-Finger Toxins (r3FTXs)
2.2. Structural Characterization of r3FTXs
2.3. Assessment of the Immunogenicity of Individual r3FTX in a Mouse Model
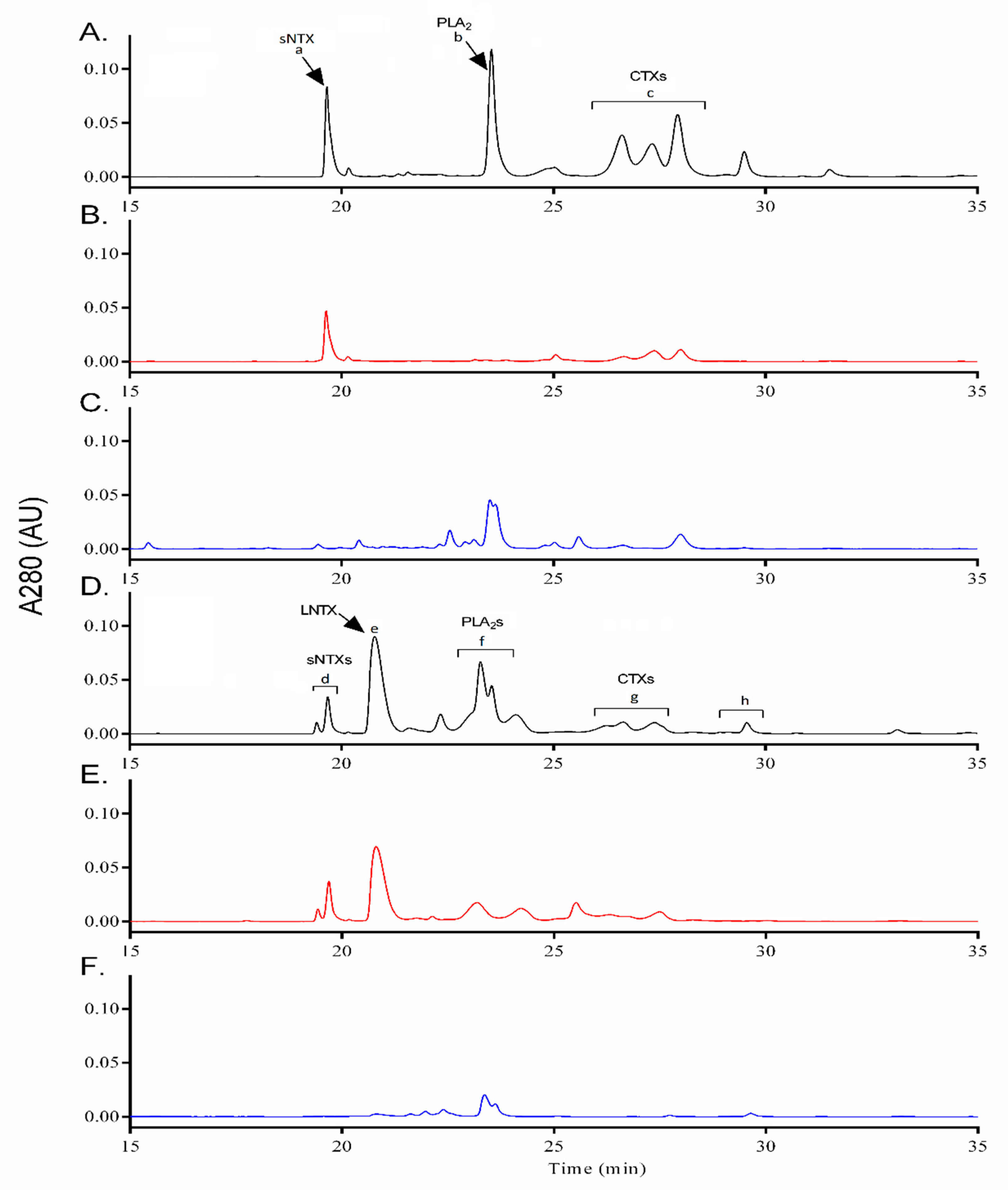
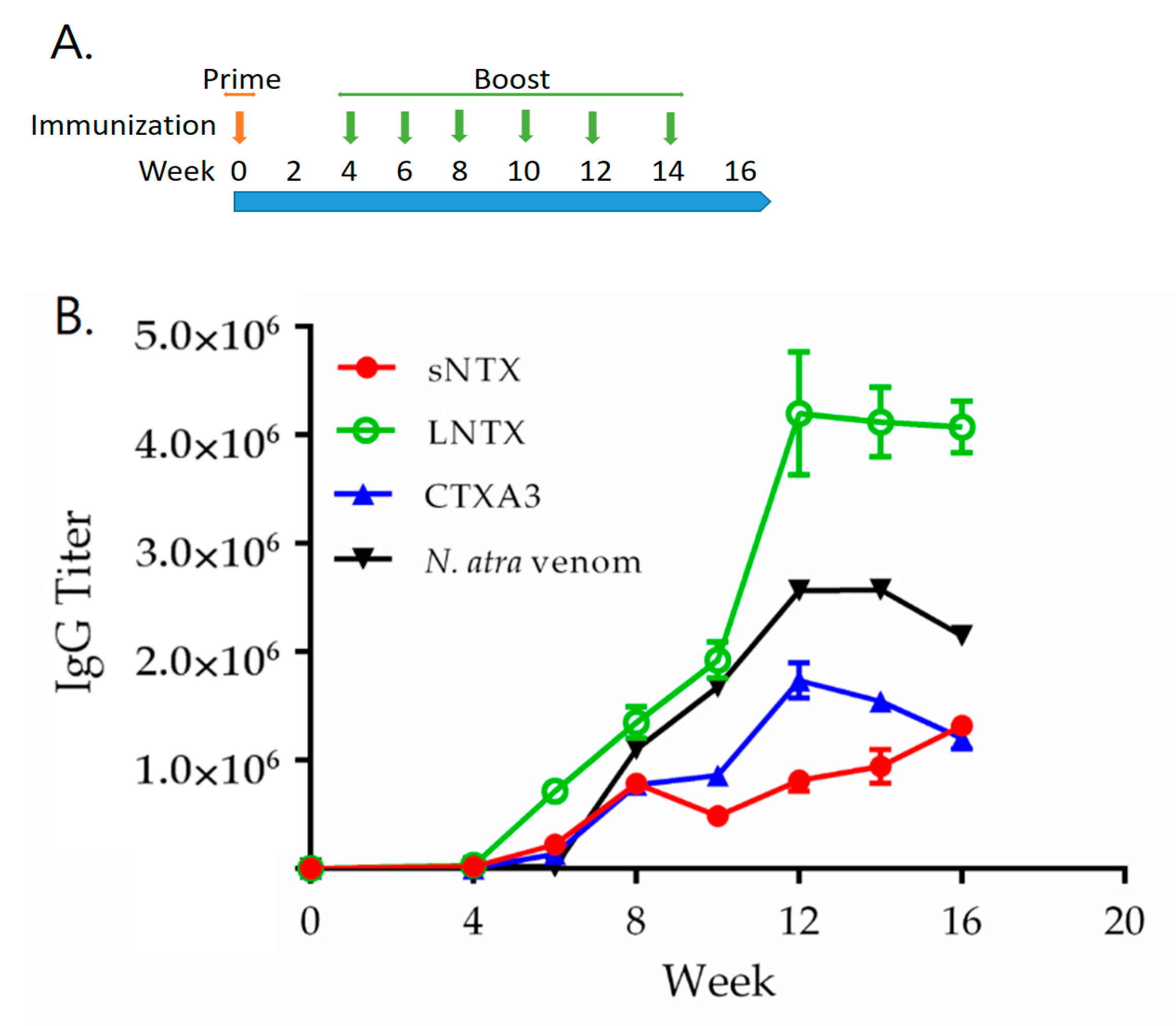
2.4. Assessment of Immunogenicity of r3FTX Mixture in Rabbit Model
2.5. Immunoreactivity and Neutralizing Potency of Rabbit Anti-r3FTXs Antibodies
2.6. Hyperimmunization with r3FTXs in Horse Model and Potency Evaluation of Horse Antiserum
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Venoms and Antivenoms
5.2. Expression of r3FTXs
5.3. Expression of Recombinant TEV Protease
5.4. Gel Electrophoresis and Immunoblotting
5.5. Circular Dichroism Spectroscopy Analysis
5.6. Analysis of Disulfide Bond Linkages Using Mass Spectrometry
5.7. Separation of Venom Proteins by RP-HPLC
5.8. Animal Experiments
5.8.1. Animals
5.8.2. Determination of Venom Lethality
5.8.3. Immunization of Mice with Individual r3FTX and Antibody Titration Analysis of Antiserum
5.8.4. Immunization with r3FTXs Mixture in Rabbit
5.8.5. Immunoreactivity of Rabbit Anti-r3FTXs Antibodies with Venom Components
5.8.6. Hyperimmunization of Horse with r3FTX Mixture
5.8.7. In Vivo Neutralization Assay
5.8.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chippaux, J.P. Snake-bites: Appraisal of the global situation. Bull World Health Organ. 1998, 76, 515–524. [Google Scholar] [PubMed]
- Su, H.Y.; Wang, M.J.; Li, Y.H.; Tang, C.N.; Tsai, M.J. Can surgical need in patients with Naja atra (Taiwan or Chinese cobra) envenomation be predicted in the emergency department? Hong Kong Med. J. 2016, 22, 435–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, L.L.; Gao, J.F.; Zhang, Y.X.; Shen, S.S.; He, Y.; Wang, J.; Ma, X.M.; Ji, X. Proteomic characterization and comparison of venoms from two elapid snakes (Bungarus multicinctus and Naja atra) from China. J. Proteom. 2016, 138, 83–94. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Ahmed, M.; Nadeem, A.; Mahajan, J.; Choudhary, A.; Pal, J. Emergency treatment of a snake bite: Pearls from literature. J. Emergencies Trauma Shock. 2008, 1, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Viravan, C.; Looareesuwan, S.; Kosakarn, W.; Wuthiekanun, V.; McCarthy, C.J.; Stimson, A.F.; Bunnag, D.; Harinasuta, T.; Warrell, D.A. A national hospital-based survey of snakes responsible for bites in Thailand. Trans. R Soc. Trop. Med. Hyg. 1992, 86, 100–106. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Management of Snake-Bites; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Tan, C.H.; Liew, J.L.; Tan, K.Y.; Tan, N.H. Assessing SABU (Serum Anti Bisa Ular), the sole Indonesian antivenom: A proteomic analysis and neutralization efficacy study. Sci. Rep. 2016, 6, 37299. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.H.; Palasuberniam, P.; Blanco, F.B.; Tan, K.Y. Immunoreactivity and neutralization capacity of Philippine cobra antivenom against Naja philippinensis and Naja samarensis venoms. Trans. R. Soc. Trop. Med. Hyg. 2020. [Google Scholar] [CrossRef]
- Deka, A.; Gogoi, A.; Das, D.; Purkayastha, J.; Doley, R. Proteomics of Naja kaouthia venom from North East India and assessment of Indian polyvalent antivenom by third generation antivenomics. J. Proteom. 2019, 207, 103463. [Google Scholar] [CrossRef]
- Liu, C.C.; You, C.H.; Wang, P.J.; Yu, J.S.; Huang, G.J.; Liu, C.H.; Hsieh, W.C.; Lin, C.C. Analysis of the efficacy of Taiwanese freeze-dried neurotoxic antivenom against Naja kaouthia, Naja siamensis and Ophiophagus hannah through proteomics and animal model approaches. PLoS Negl. Trop. Dis. 2017, 11, e0006138. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.Y.; Tan, C.H.; Sim, S.M.; Fung, S.Y.; Tan, N.H. Geographical venom variations of the Southeast Asian monocled cobra (Naja kaouthia): Venom-induced neuromuscular depression and antivenom neutralization. CBP 2016, 185–186, 77–86. [Google Scholar] [CrossRef]
- Williams, D.J.; Gutiérrez, J.M.; Calvete, J.J.; Wüster, W.; Ratanabanangkoon, K.; Paiva, O.; Brown, N.I.; Casewell, N.R.; Harrison, R.A.; Rowley, P.D.; et al. Ending the drought: New strategies for improving the flow of affordable, effective antivenoms in Asia and Africa. J. Proteom. 2011, 74, 1735–1767. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Rabies and Envenomings: A Neglected Public Health Issue: Report of a Consultative Meeting; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- The, L. Snake-bite envenoming: A priority neglected tropical disease. Lancet 2017, 390, 2. [Google Scholar] [CrossRef] [Green Version]
- Minghui, R.; Malecela, M.N.; Cooke, E.; Abela-Ridder, B. WHO’s Snakebite Envenoming Strategy for prevention and control. Lancet Glob. Health 2019, 7, e837–e838. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Liu, J.; Xu, K. Novel genes encoding six kinds of three-finger toxins in Ophiophagus hannah (king cobra) and function characterization of two recombinant long-chain neurotoxins. Biochem. J. 2006, 398, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Nirthanan, S.; Gwee, M.C. Three-finger alpha-neurotoxins and the nicotinic acetylcholine receptor, forty years on. J. Pharmacol. Sci. 2004, 94, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Fernández, J.; Alape-Girón, A.; Angulo, Y.; Sanz, L.; Gutiérrez, J.M.; Calvete, J.J.; Lomonte, B. Venomic and antivenomic analyses of the Central American coral snake, Micrurus nigrocinctus (Elapidae). J. Proteome Res. 2011, 10, 1816–1827. [Google Scholar] [CrossRef]
- Castro, K.L.; Duarte, C.G.; Ramos, H.R.; Machado de Avila, R.A.; Schneider, F.S.; Oliveira, D.; Freitas, C.F.; Kalapothakis, E.; Ho, P.L.; Chavez-Olortegui, C. Identification and characterization of B-cell epitopes of 3FTx and PLA(2) toxins from Micrurus corallinus snake venom. Toxicon Off. J. Int. Soc. Toxinol. 2015, 93, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.Y.; Tan, C.H.; Tan, N.H. Venom and Purified Toxins of the Spectacled Cobra (Naja naja) from Pakistan: Insights into Toxicity and Antivenom Neutralization. Am. J. Trop. Med. Hyg. 2016, 94, 1392–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, C.; Solano, S.; Segura, Á.; Herrera, M.; Estrada, R.; Villalta, M.; Vargas, M.; Gutiérrez, J.M.; León, G. Cross-reactivity and cross-immunomodulation between venoms of the snakes Bothrops asper, Crotalus simus and Lachesis stenophrys, and its effect in the production of polyspecific antivenom for Central America. Toxicon Off. J. Int. Soc. Toxinol. 2017, 138, 43–48. [Google Scholar] [CrossRef]
- Ratanabanangkoon, K.; Tan, K.Y.; Eursakun, S.; Tan, C.H.; Simsiriwong, P.; Pamornsakda, T.; Wiriyarat, W.; Klinpayom, C.; Tan, N.H. A Simple and Novel Strategy for the Production of a Pan-specific Antiserum against Elapid Snakes of Asia. PLoS Negl. Trop. Dis. 2016, 10, e0004565. [Google Scholar] [CrossRef] [Green Version]
- Dolimbek, B.Z.; Atassi, M.Z. Protection against alpha-bungarotoxin poisoning by immunization with synthetic toxin peptides. Mol. Immunol. 1996, 33, 681–689. [Google Scholar] [CrossRef]
- Léonetti, M.; Pillet, L.; Maillère, B.; Lamthanh, H.; Frachon, P.; Couderc, J.; Ménez, A. Immunization with a peptide having both T cell and conformationally restricted B cell epitopes elicits neutralizing antisera against a snake neurotoxin. J. Immunol. 1990, 145, 4214–4221. [Google Scholar] [PubMed]
- Wagstaff, S.C.; Laing, G.D.; Theakston, R.D.; Papaspyridis, C.; Harrison, R.A. Bioinformatics and multiepitope DNA immunization to design rational snake antivenom. PLoS Med. 2006, 3, e184. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Garzón, J.F.; Bénard-Valle, M.; Restano-Cassulini, R.; Zamudio, F.; Corzo, G.; Alagón, A.; Olvera-Rodríguez, A. Cloning and sequencing of three-finger toxins from the venom glands of four Micrurus species from Mexico and heterologous expression of an alpha-neurotoxin from Micrurus diastema. Biochimie 2018, 147, 114–121. [Google Scholar] [CrossRef]
- de la Rosa, G.; Corrales-García, L.L.; Rodriguez-Ruiz, X.; López-Vera, E.; Corzo, G. Short-chain consensus alpha-neurotoxin: A synthetic 60-mer peptide with generic traits and enhanced immunogenic properties. Amino Acids 2018, 50, 885–895. [Google Scholar] [CrossRef] [PubMed]
- de la Rosa, G.; Olvera, F.; Archundia, I.G.; Lomonte, B.; Alagón, A.; Corzo, G. Horse immunization with short-chain consensus α-neurotoxin generates antibodies against broad spectrum of elapid venomous species. Nat. Commun. 2019, 10, 3642. [Google Scholar] [CrossRef] [Green Version]
- Laustsen, A.H.; Gutiérrez, J.M.; Lohse, B.; Rasmussen, A.R.; Fernández, J.; Milbo, C.; Lomonte, B. Snake venomics of monocled cobra (Naja kaouthia) and investigation of human IgG response against venom toxins. Toxicon Off. J. Int. Soc. Toxinol. 2015, 99, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.Y.; Wei, T.Y.; Liu, B.S.; Lin, M.H.; Chiang, S.K.; Chen, S.F.; Sung, W.C. Monitoring the Disulfide Bonds of Folding Isomers of Synthetic CTX A3 Polypeptide Using MS-Based Technology. Toxins 2019, 11, 52. [Google Scholar] [CrossRef] [Green Version]
- Lobstein, J.; Emrich, C.A.; Jeans, C.; Faulkner, M.; Riggs, P.; Berkmen, M. SHuffle, a novel Escherichia coli protein expression strain capable of correctly folding disulfide bonded proteins in its cytoplasm. Microb. Cell Fact. 2012, 11, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchetto, J.; Sequeira, A.F.; Ramond, L.; Peysson, F.; Bras, J.L.; Saez, N.J.; Duhoo, Y.; Blemont, M.; Guerreiro, C.I.; Quinton, L.; et al. High-throughput expression of animal venom toxins in Escherichia coli to generate a large library of oxidized disulphide-reticulated peptides for drug discovery. Microb. Cell Fact. 2017, 16, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozach, H.; Fruchart-Gaillard, C.; Fenaille, F.; Beau, F.; Ramos, O.H.; Douzi, B.; Saez, N.J.; Moutiez, M.; Servent, D.; Gondry, M.; et al. High throughput screening identifies disulfide isomerase DsbC as a very efficient partner for recombinant expression of small disulfide-rich proteins in E. coli. Microb. Cell Fact. 2013, 12, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moise, L.; Piserchio, A.; Basus, V.J.; Hawrot, E. NMR structural analysis of alpha-bungarotoxin and its complex with the principal alpha-neurotoxin-binding sequence on the alpha 7 subunit of a neuronal nicotinic acetylcholine receptor. J. Biol. Chem. 2002, 277, 12406–12417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betzel, C.; Lange, G.; Pal, G.P.; Wilson, K.S.; Maelicke, A.; Saenger, W. The refined crystal structure of alpha-cobratoxin from Naja naja siamensis at 2.4-A resolution. J. Biol. Chem. 1991, 266, 21530–21536. [Google Scholar] [CrossRef]
- Panagides, N.; Jackson, T.N.; Ikonomopoulou, M.P.; Arbuckle, K.; Pretzler, R.; Yang, D.C.; Ali, S.A.; Koludarov, I.; Dobson, J.; Sanker, B.; et al. How the Cobra Got Its Flesh-Eating Venom: Cytotoxicity as a Defensive Innovation and Its Co-Evolution with Hooding, Aposematic Marking, and Spitting. Toxins 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Ramos, H.R.; Junqueira-de-Azevedo Ide, L.; Novo, J.B.; Castro, K.; Duarte, C.G.; Machado-de-Ávila, R.A.; Chavez-Olortegui, C.; Ho, P.L. A Heterologous Multiepitope DNA Prime/Recombinant Protein Boost Immunisation Strategy for the Development of an Antiserum against Micrurus corallinus (Coral Snake) Venom. PLoS Negl. Trop. Dis. 2016, 10, e0004484. [Google Scholar] [CrossRef] [Green Version]
- Lankisch, P.G.; Lege, L.; Oldigs, H.D.; Vogt, W. Binding of phospholipase A to the cirect lytic factor revealed by the interaction of Ca2+ with the haemolytic effect. Biochim. Biophys. Acta 1971, 239, 267–272. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics, lethality and neutralization of Naja kaouthia (monocled cobra) venoms from three different geographical regions of Southeast Asia. J. Proteom. 2015, 120, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Yap, M.K.; Fung, S.Y.; Tan, K.Y.; Tan, N.H. Proteomic characterization of venom of the medically important Southeast Asian Naja sumatrana (Equatorial spitting cobra). Acta Trop 2014, 133, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Tan, C.H.; Tan, K.Y.; Quraishi, N.H.; Tan, N.H. Elucidating the biogeographical variation of the venom of Naja naja (spectacled cobra) from Pakistan through a venom-decomplexing proteomic study. J. Proteom. 2018, 175, 156–173. [Google Scholar] [CrossRef]
- Lauridsen, L.P.; Laustsen, A.H.; Lomonte, B.; Gutierrez, J.M. Exploring the venom of the forest cobra snake: Toxicovenomics and antivenom profiling of Naja melanoleuca. J. Proteom. 2017, 150, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Tan, N.H.; Wong, K.Y.; Tan, C.H. Venomics of Naja sputatrix, the Javan spitting cobra: A short neurotoxin-driven venom needing improved antivenom neutralization. J. Proteom. 2017, 157, 18–32. [Google Scholar] [CrossRef]
- Petras, D.; Sanz, L.; Segura, A.; Herrera, M.; Villalta, M.; Solano, D.; Vargas, M.; Leon, G.; Warrell, D.A.; Theakston, R.D.; et al. Snake venomics of African spitting cobras: Toxin composition and assessment of congeneric cross-reactivity of the pan-African EchiTAb-Plus-ICP antivenom by antivenomics and neutralization approaches. J. Proteome Res. 2011, 10, 1266–1280. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.W.; Liu, B.S.; Chien, K.Y.; Chiang, L.C.; Huang, S.Y.; Sung, W.C.; Wu, W.G. Cobra venom proteome and glycome determined from individual snakes of Naja atra reveal medically important dynamic range and systematic geographic variation. J. Proteom. 2015, 128, 92–104. [Google Scholar] [CrossRef]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Bulyaki, E.; Kun, J.; Moussong, E.; Lee, Y.H.; Goto, Y.; Refregiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucleic Acids Res. 2018, 46, W315–W322. [Google Scholar] [CrossRef]
- Huang, S.Y.; Chen, S.F.; Chen, C.H.; Huang, H.W.; Wu, W.G.; Sung, W.C. Global disulfide bond profiling for crude snake venom using dimethyl labeling coupled with mass spectrometry and RADAR algorithm. Anal. Chem. 2014, 86, 8742–8750. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.S.; Wu, W.G.; Lin, M.H.; Li, C.H.; Jiang, B.R.; Wu, S.C.; Leng, C.H.; Sung, W.C. Identification of Immunoreactive Peptides of Toxins to Simultaneously Assess the Neutralization Potency of Antivenoms against Neurotoxicity and Cytotoxicity of Naja atra Venom. Toxins 2017, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Chippaux, J.P. Guidelines for the production, control and regulation of snake antivenom immunoglobulins. Biol. Aujourd’hui 2010, 204, 87–91. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Disulfide-Linked Peptide | ||||||
|---|---|---|---|---|---|---|---|
| Toxin | Expected MW (Da) 1 | Observed MW (Da) | Toxicity | Secondary Structure 3 | Peptide Sequence | Mass Ion (m/z) | Linkage Site(s) |
| LNTX | 7815.66 | 7815.57 | LD50 2: 0.15 | 33.5% β-sheet | TWC26DAFC30SIR | 616.2 (2+) | C26-30 |
| (0.12–0.19) | 9.8% α-helix | C3FITPDITSK | 846.9 (7+) | C3-C14 | |||
| 10.5% Turn | DC14PNGHVC20YTK | C20-C41 | |||||
| VDLGC41AATC45PTVK | C45-C56 | ||||||
| TGVDIQC56C57STDNC62NPFPTR | C57-C62 | ||||||
| rLNTX | 7872.68 | 7872.58 | LD50: 0.21 | 34.7% β-sheet | TWC27DAFC31SIR | 616.2 (2+) | C27-31 |
| (0.18–0.24) | 9.2% α-helix | C4FITPDITSK | 846.9 (7+) | C4-C15 | |||
| 9.9% Turn | DC15PNGHVC21YTK | C21-C42 | |||||
| VDLGC42AATC46PTVK | C46-C57 | ||||||
| TGVDIQC57C58STDNC63NPFPTR | C58-C63 | ||||||
| sNTX | 6945.04 | 6944.97 | LD50: 0.23 | 54.5% β-sheet | LEC3HNQQSSQTPTTTGC17SGGETNC24YK | 899.9 (6+) | C3-C24 |
| (0.17–0.26) | 0.0% α-helix | GC41GC43PSVK | C17-C41 | ||||
| 7.4% Turn | NGIEINC54C55TTDR | C43-C54 | |||||
| C60NN | C55-C60 | ||||||
| rsNTX | 7001.06 | 7000.99 | LD50: 0.25 | 53.4% β-sheet | GLEC4HNQQSSQTPTTTGC18SGGETNC25YK | 909.3 (6+) | C4-C25 |
| (0.19–0.33) | 0.0% α-helix | GC42GC44PSVK | C18-C42 | ||||
| 8.1% Turn | NGIEINC55C56TTDR | C44-C55 | |||||
| C61NN | C56-C61 | ||||||
| CTXA3 | 6734.43 | 6735.36 | IC50 4: 1.93 | 54.5% β-sheet | LKC3NK | 701.9 (2+) | C3-C21 |
| (1.67–2.23) | 0.0% α-helix | NLC21YK | |||||
| 7.4% Turn | TC14PAGK | 561.4 (5+) | C14-C38 | ||||
| GC38IDVC42PK | C42-C53 | ||||||
| YVC53C54NTDR | C54-C59 | ||||||
| C59N | |||||||
| rCTXA3 | 6791.45 | 6792.38 | IC50 4: 2.02 | 53.4% β-sheet | GLKC4NK | 487.3 (3+) | C4-C22 |
| (1.88–2.15) | 0.0 % α-helix | NLC22YK | |||||
| 8.1% Turn | TC15PAGK | 561.5 (5+) | C15-C39 | ||||
| GC39IDVC43PK | C43-C54 | ||||||
| YVC54C55NTDR | C55-C60 | ||||||
| C60N | |||||||
| Immunogen 1/Assays | Antibody Titer 2 | Potency | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Anti-sNTX | Anti-LNTX | Anti-CTXA3 | Naja atra3 × LD50 | Naja kaouthia 5 × LD50 | Naja siamensis 3 × LD50 | ||||
| ED50 4 (mg) | P 5 (mg/g) | ED50 (mg) | P (mg/g) | ED50 (mg) | P (mg/g) | ||||
| Venoms | 1.2 × 105 | 6.6 × 105 | 1.3 × 105 | 3.3 | 8.3 | 4.9 | 4.7 | 1.8 | 6.5 |
| r3FTXs | 1.9 × 105 | 6.0 × 105 | 2.9 × 105 | <2.5 | >20 | 3.8 | 6.1 | 1.4 | 8.2 |
| Preimmune | <103 | <103 | <103 | N.E. 3 | N.E. | N.E. | N.E. | N.E. | N.E. |
| N. atra 3 × LD50 | N. kaouthia 5 × LD50 | N. siamensis 3 × LD50 | ||||
|---|---|---|---|---|---|---|
| Horse anti-r3FTXs antibodies | ED50 (mg) 2 | P (mg/g) | ED50 (mg) | P (mg/g) | ED50 (mg) | P (mg/g) |
| 1.26 (0.58–2.76) 1 | 21.33 | 3.78 (2.50–5.71) 1 | 6.10 | 0.45 | 25.78 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.-S.; Jiang, B.-R.; Hu, K.-C.; Liu, C.-H.; Hsieh, W.-C.; Lin, M.-H.; Sung, W.-C. Development of a Broad-Spectrum Antiserum against Cobra Venoms Using Recombinant Three-Finger Toxins. Toxins 2021, 13, 556. https://doi.org/10.3390/toxins13080556
Liu B-S, Jiang B-R, Hu K-C, Liu C-H, Hsieh W-C, Lin M-H, Sung W-C. Development of a Broad-Spectrum Antiserum against Cobra Venoms Using Recombinant Three-Finger Toxins. Toxins. 2021; 13(8):556. https://doi.org/10.3390/toxins13080556
Chicago/Turabian StyleLiu, Bing-Sin, Bo-Rong Jiang, Kai-Chieh Hu, Chien-Hsin Liu, Wen-Chin Hsieh, Min-Han Lin, and Wang-Chou Sung. 2021. "Development of a Broad-Spectrum Antiserum against Cobra Venoms Using Recombinant Three-Finger Toxins" Toxins 13, no. 8: 556. https://doi.org/10.3390/toxins13080556
APA StyleLiu, B. -S., Jiang, B. -R., Hu, K. -C., Liu, C. -H., Hsieh, W. -C., Lin, M. -H., & Sung, W. -C. (2021). Development of a Broad-Spectrum Antiserum against Cobra Venoms Using Recombinant Three-Finger Toxins. Toxins, 13(8), 556. https://doi.org/10.3390/toxins13080556