DL-Selenomethionine Alleviates Oxidative Stress Induced by Zearalenone via Nrf2/Keap1 Signaling Pathway in IPEC-J2 Cells


Abstract
:1. Introduction
2. Results
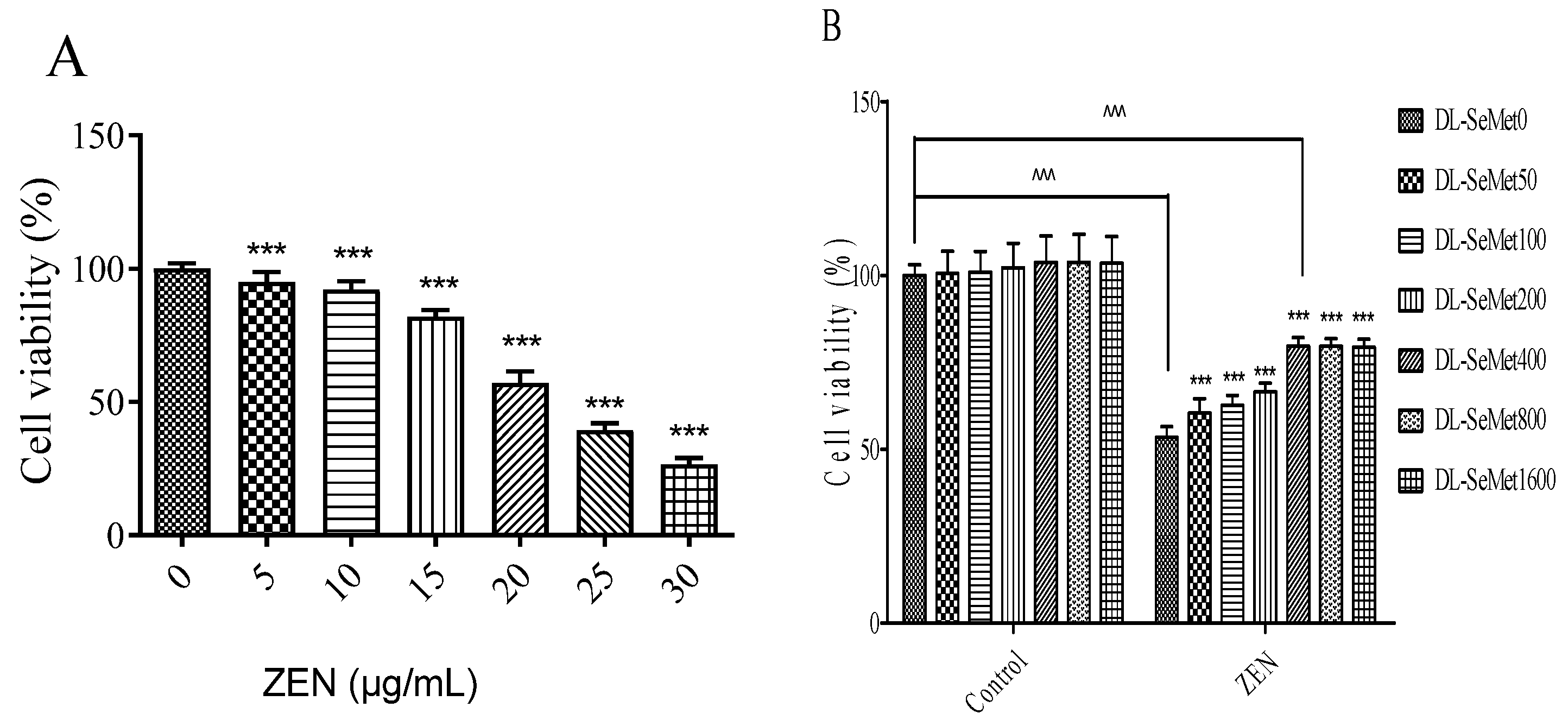
2.1. Experiment on Cell Viability
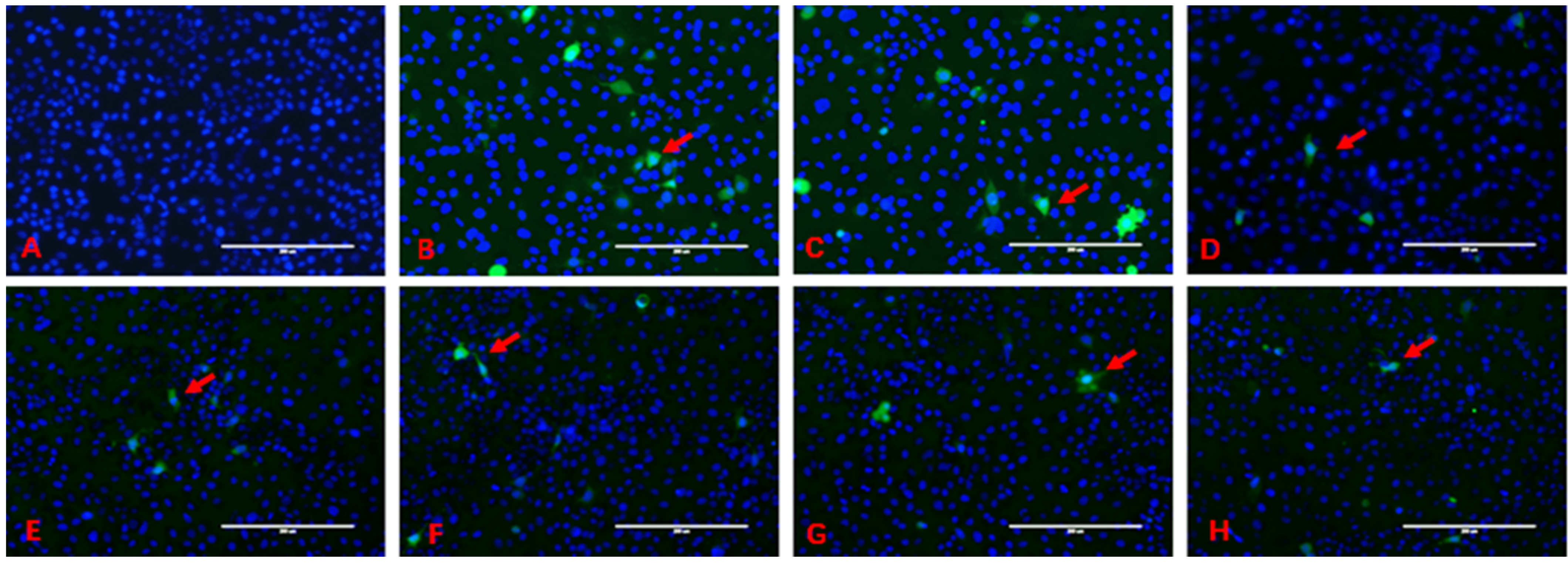
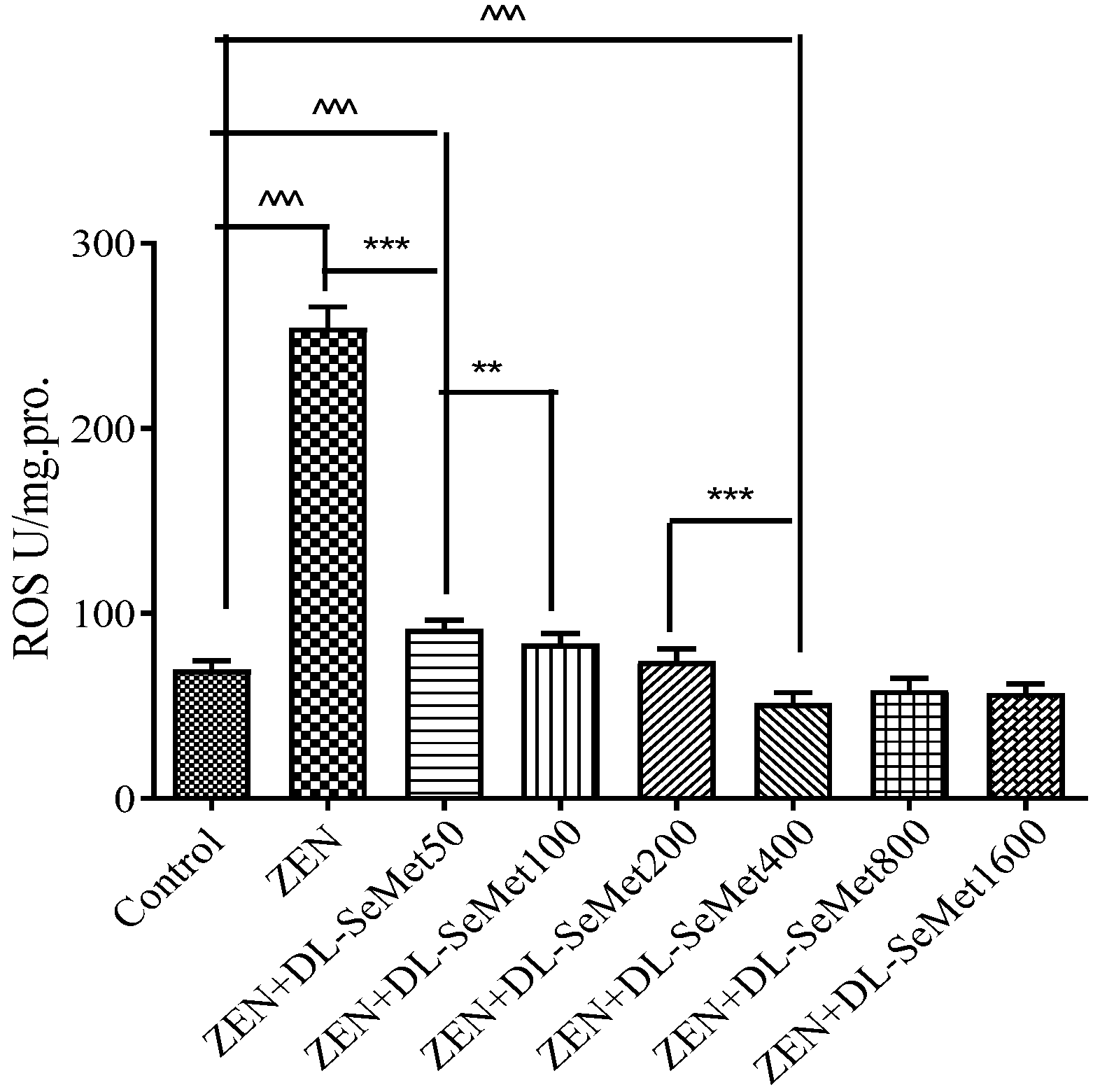
2.2. DL-SeMet Alleviates ZEN-Induced ROS Production
2.3. DL-SeMet Alleviates ZEN-Induced ROS Production
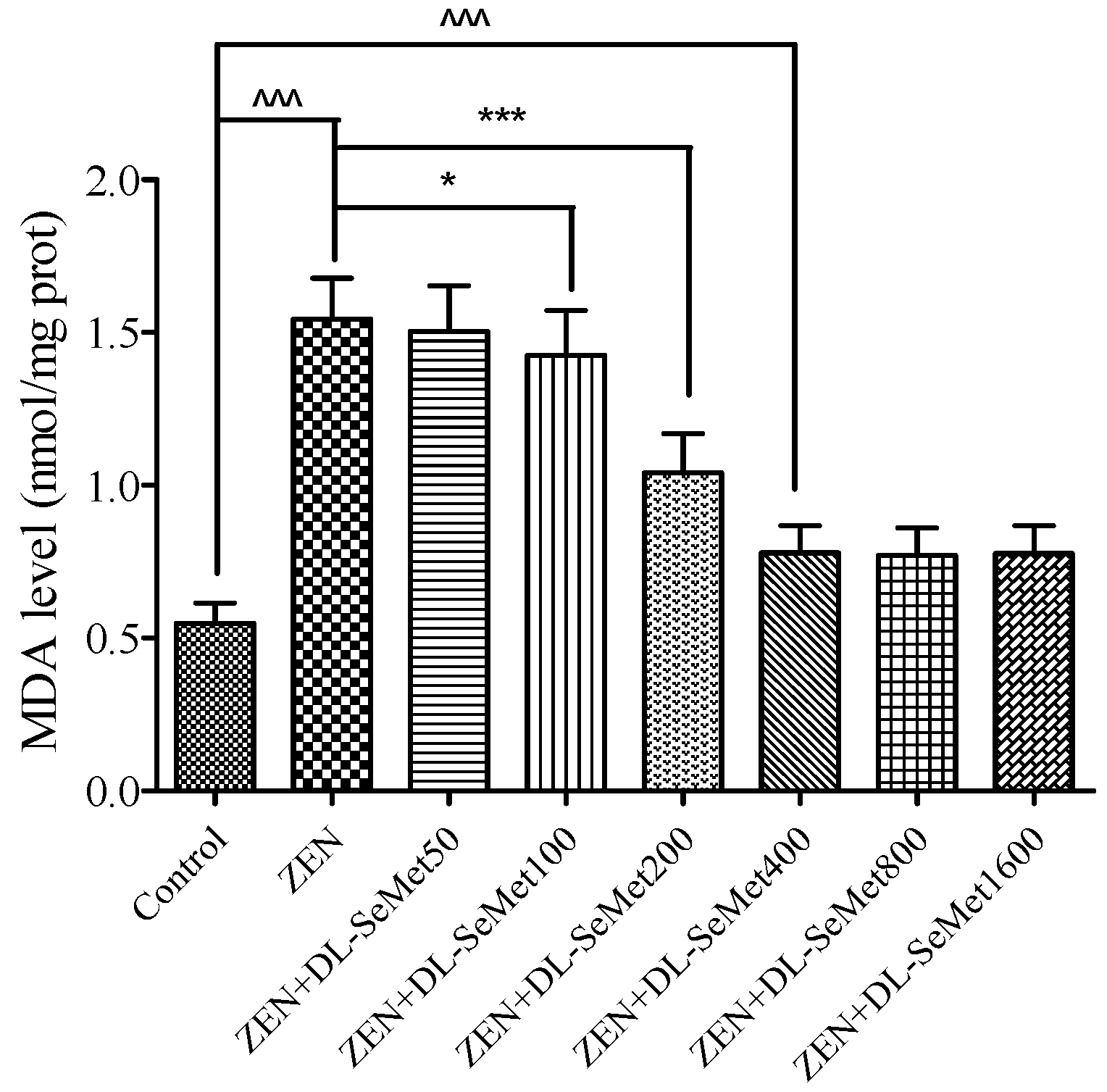
2.4. DL-SeMet Attenuates ZEN-Stimulated Lipid Peroxidation
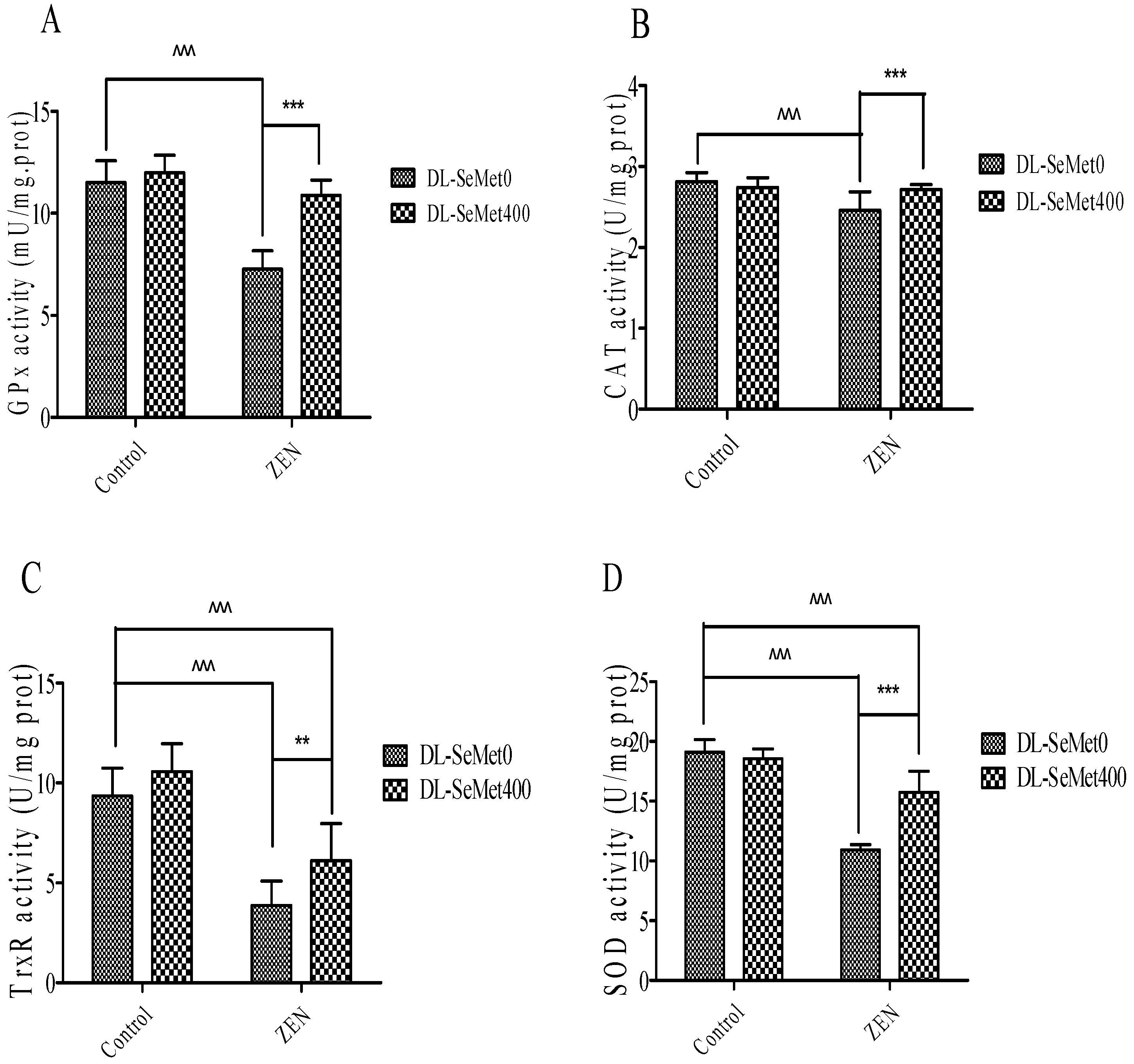
2.5. DL-SeMet Prevents ZEN-Induced Decrease in Enzymatic Antioxidants
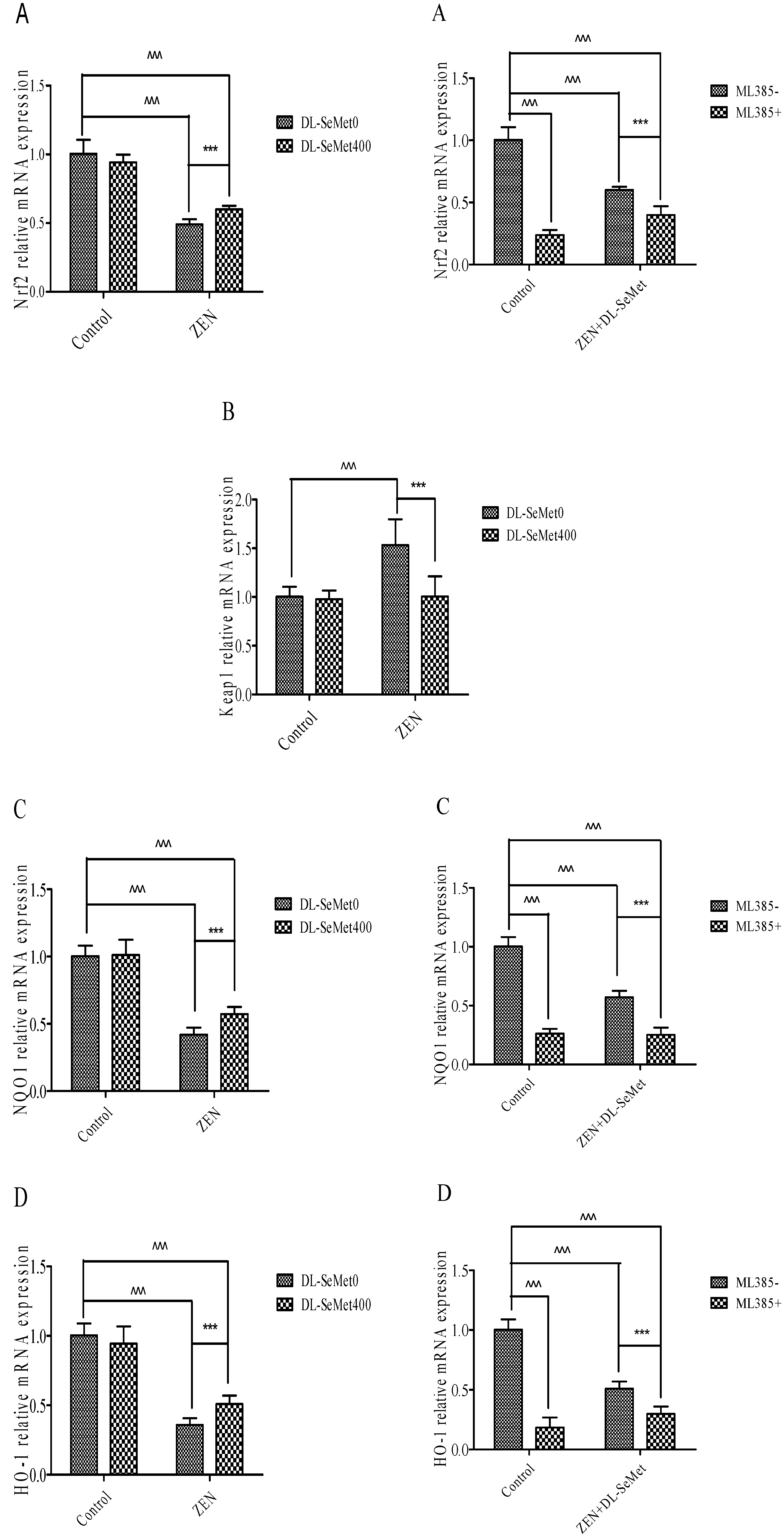
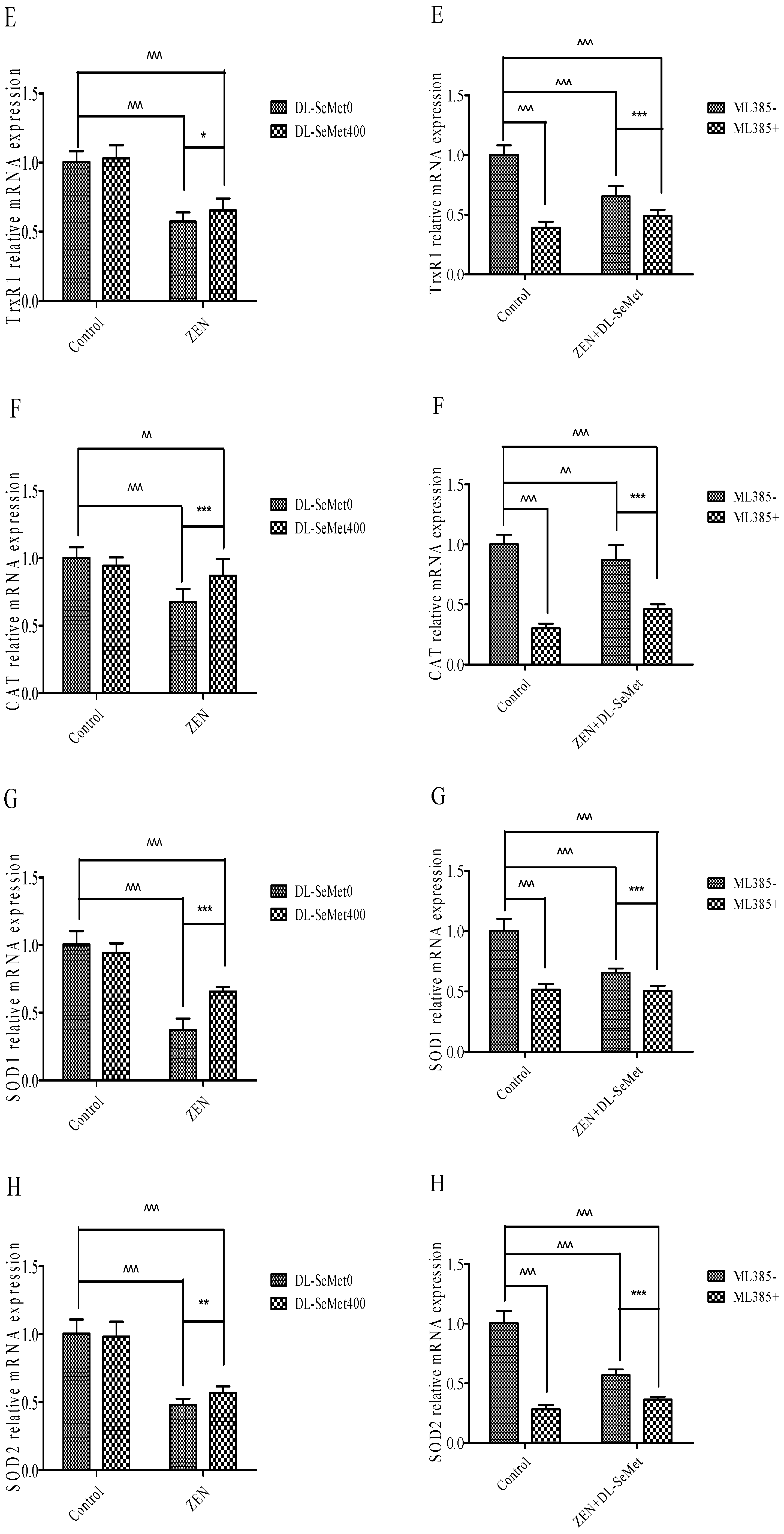
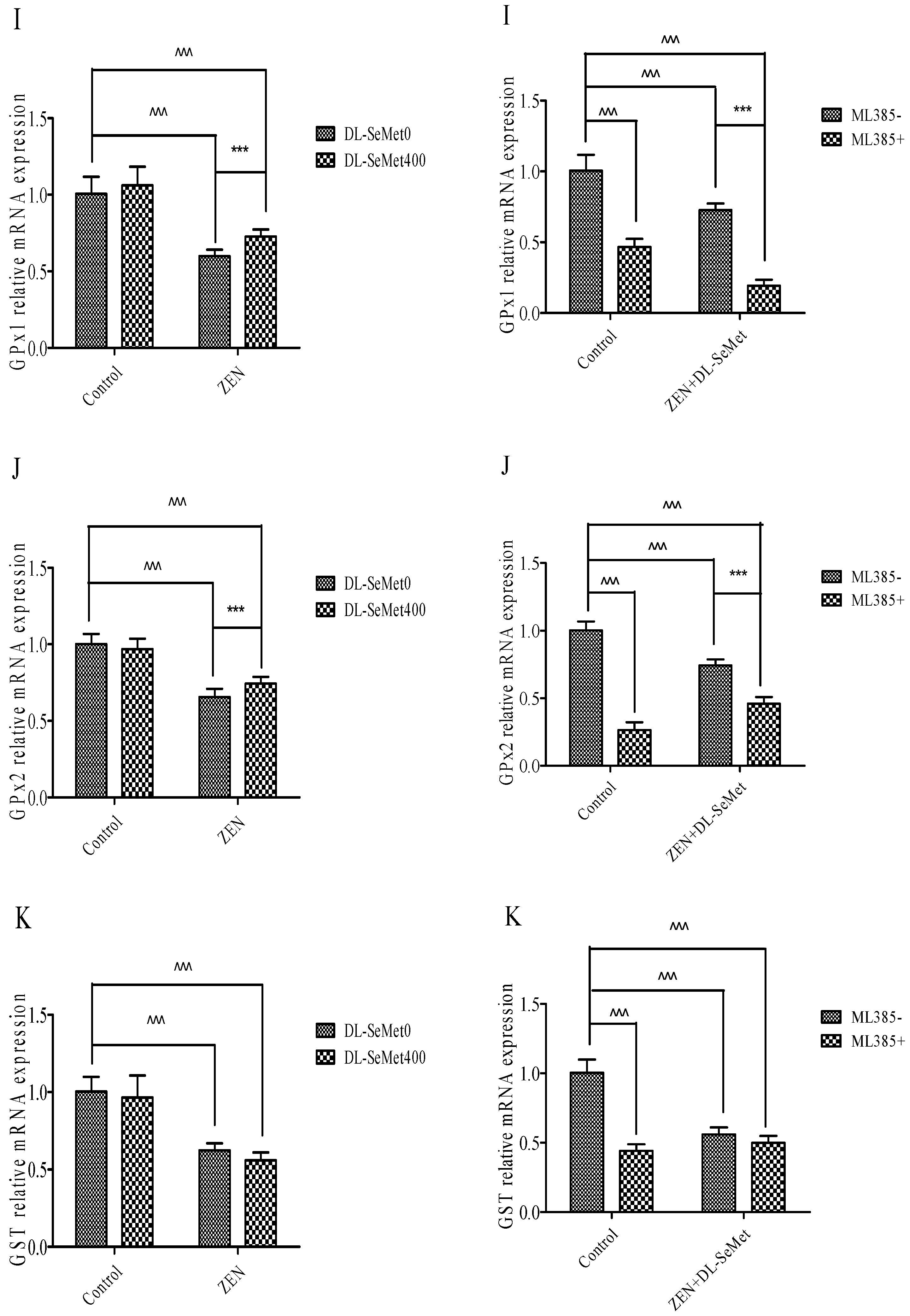
2.6. DL-SeMet Normalizes the mRNA Expression of Nrf2, Keap1 and Enzymatic Antioxidants in ZEN Treated IPEC-J2 Cells
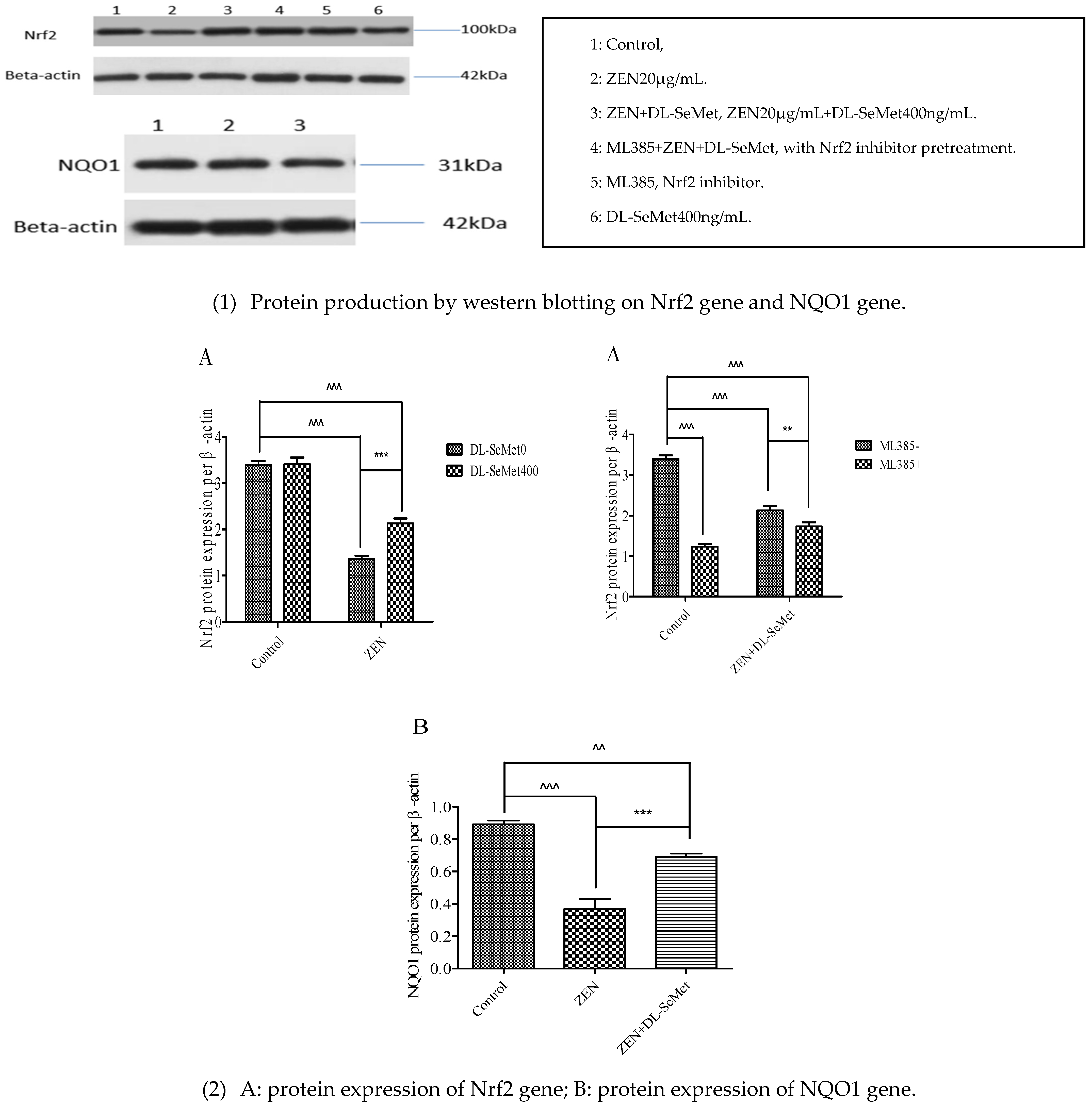
2.7. DL-SeMet Regularizes the Protein Expression of Nrf2 and NQO1 in ZEN Treated IPEC-J2 Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cell Culture and Treatment
5.3. Measurement of Cell Viability
5.4. Determination of Intracellular ROS
5.5. Evaluation of ROS Level
5.6. Lipid Peroxidation (MDA) Assay
5.7. Antioxidant Enzymes Activity Assays
5.8. Quantitative Real-Time PCR
5.9. Western Blot Analysis
5.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Cynthia, C.; Marthe, D.B.; Olusegun, A. The status of fusarium mycotoxins in sub-saharan africa: A review of emerging trends and post-harvest mitigation strategies towards food control. Toxins 2017, 9, 19. [Google Scholar]
- Mally, A.; Solfrizzo, M.; Degen, G.H. Biomonitoring of the mycotoxin zearalenone: Current state-of-the art and application to human exposure assessment. Arch. Toxicol. 2016, 90, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Calori-Domingues, M.A.; Bernardi, C.M.G.; Nardin, M.S. Co-occurrence and distribution of deoxynivalenol, nivalenol and zearalenone in wheat from Brazil. Food Addit. Contam. Part B 2016, 9, 142–151. [Google Scholar] [CrossRef]
- Oveisi, M.R.; Mannan, H.; Shadi, M. Determination of zearalenone in corn flour and a cheese snack product using high-performance liquid chromatography with fluorescence detection. Food Addit. Contam. 2005, 22, 6. [Google Scholar]
- Hassan, Z.U.; Thani, R.A.; Atia, F.A.; Meer, S.A.; Jaoua, S. Co-occurrence of mycotoxins in commercial formula milk and cereal-based baby food in qatar. Food Addit. Contam. Part B 2018, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Cojocneanu-Petric, R.; Jurj, A.; Taranu, I.; Gras, A.M. Microarray based gene expression analysis of sus scrofa duodenum exposed to zearalenone: Significance to human health. BMC Genom. 2016, 17, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Zhang, S.; Zhang, M.; Yang, L.; Cheng, B.; Li, J. Individual and combined effects of Fusarium toxins on apoptosis in PK15 cells and the protective role of N-acetylcysteine. Food Chem. Toxicol. 2017, 111, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, M.; Zhang, W.; Gu, A.; Dong, J.; Li, J. Protective effect of N-acetylcysteine against oxidative stress induced by zearalenone via mitochondrial apoptosis pathway in SIEC02 cells. Toxins 2018, 10, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatay, E.; Font, G.; Ruiz, M.J. Cytotoxic effects of zearalenone and its metabolites and antioxidant cell defense in CHO-K1 cells. Food Chem. Toxicol. 2016, 96, 43–49. [Google Scholar] [CrossRef]
- Maher, J.; Yamamoto, M. The rise of antioxidant signaling e the evolution and hormetic actions of Nrf2. Toxicol. Appl. Pharmacol. 2010, 244, 4–15. [Google Scholar] [CrossRef]
- Wastney, M.E.; Combs, G.F.; Canfield, W.K. A human model of selenium that integrates metabolism from selenite and selenomethionine. J. Nutr. 2011, 141, 708–717. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.Z.; Tiwary, A.K.; Stegelmeier, B.L. Comparative oral dose toxicokinetics of sodium selenite and selenomethionine. J. Appl. Toxicol. 2016, 37, 2. [Google Scholar] [CrossRef]
- Nickel, A.; Kottra, G.; Schmidt, G. Characteristics of transport of selenoamino acids by epithelial amino acid transporters. Chem. Biol. Interact. 2009, 177, 234–241. [Google Scholar] [CrossRef]
- Shafik, N.M.; El Batsh, M.M. Protective effects of combined selenium and punica granatum treatment on some inflammatory and oxidative stress markers in arsenic-induced hepatotoxicity in rats. Biol. Trace Elem. Res. 2016, 169, 121–128. [Google Scholar] [CrossRef]
- Zhu, L.; Yuan, H.; Guo, C. Zearalenone induces apoptosis and necrosis in porcine granulosa cells via a caspase-3-and caspase-9-dependent mitochondrial signaling pathway. J. Cell. Physiol. 2012, 227, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.Y.; Zheng, Z.H.; Son, Y.O. Mycotoxin zearalenone induces AIF-and ROS-mediated cell death through p53-and MAPK-dependent signaling pathways in RAW264. 7 macrophages. Toxicol. In Vitro 2011, 25, 1654–1663. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Selenium: Significance and outlook for supplementation. Nutrition 2013, 29, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cheng, Y.; Li, X. Biogenic nanoselenium particles effectively attenuate oxidative stress-induced intestinal epithelial barrier injury by activating the Nrf2 antioxidant pathway. ACS Appl. Mater. Interfaces 2017, 9, 14724–14740. [Google Scholar] [CrossRef]
- Hou, L.; Zhou, X.; Gan, F. Combination of selenomethionine and N-acetylcysteine alleviates the joint toxicities of aflatoxin B1 and ochratoxin A by ERK MAPK signal pathway in porcine alveolar macrophages. J. Agric. Food Chem. 2018, 66, 5913–5923. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Hong, X.; Yu, H. Selenium alleviates porcine nephrotoxicity of ochratoxin a by improving selenoenzyme expression in vitro. PLoS ONE 2015, 10, e0119808. [Google Scholar]
- Wang, X.; Zuo, Z.; Zhao, C. Protective role of selenium in the activities of antioxidant enzymes in piglet splenic lymphocytes exposed to deoxynivalenol. Environ. Toxicol. Pharmacol. 2016, 47, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Zanelatto, L.C.; Marques, L.A.; Brianese, R.C. BIO044 chemoprotective action of L-selenomethionine in HepG2 cells: Evaluation of stress genes expression induced by H2O2. Rev. Eletrônica Farmácia 2012, 9, 1. [Google Scholar]
- Hao, S.; Hu, J.; Song, S. Selenium alleviates aflatoxin b₁-induced immune toxicity through improving glutathione peroxidase 1 and selenoprotein S expression in primary porcine splenocytes. J. Agric. Food Chem. 2016, 64, 1–9. [Google Scholar] [CrossRef]
- Venkataramana, M.; Nayaka, S.C.; Anand, T. Zearalenone induced toxicity in SHSY-5Y cells: The role of oxidative stress evidenced by N-acetyl cysteine. Food Chem. Toxicol. 2014, 65, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Jornot, L.; Junod, A.F. Differential regulation of glutathione peroxidase by selenomethionine and hyperoxia in endothelial cells. Biochem. J. 1995, 306, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I. Glutathione peroxidases in poultry biology: Part 2. Modulation of enzymatic activities. World’s Poult. Sci. J. 2018, 74, 239–250. [Google Scholar] [CrossRef]
- Nioi, P.; Nguyen, T.; Sherratt, P.J.; Pickett, C.B. The carboxy-terminal Neh3 domain of Nrf2 is required for transcriptional activation. Mol. Cell. Biol. 2005, 25, 10895–10906. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Kyo, M.; Kamiya, T. Predictive base substitution rules that determine the binding and transcriptional specificity of Maf recognition elements. Genes Cells 2006, 11, 575–591. [Google Scholar] [CrossRef] [Green Version]
- Ak, T.; Lhami, G. Antioxidant and radical scavenging properties of curcumin. Chem. Biol. Interact. 2008, 174, 27–37. [Google Scholar] [CrossRef]
- Périne, D.; Aurélie, B.; Vasseur, P. Molecular cloning and expression study of pi-class glutathione S-transferase (pi-GST) and selenium-dependent glutathione peroxidase (Se-GPx) transcripts in the freshwater bivalve dreissena polymorpha. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2008, 147, 69–77. [Google Scholar]
- Wang, C.; Yang, Y.; Gao, N.; Lan, J.; Dou, X.; Li, J.; Shan, A. L-Threonine upregulates the expression of β-defensins by activating the NF-κB signaling pathway and suppressing SIRT1 expression in porcine intestinal epithelial cells. Food Funct. 2021, 12, 5821–5836. [Google Scholar] [CrossRef]
- Ayed-Boussema, I.; Bouaziz, C.; Rjiba, K. The mycotoxin zearalenone induces apoptosis in human hepatocytes (HepG2) via p53-dependent mitochondrial signaling pathway. Toxicol. Vitr. 2008, 22, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Feng, Y.; Deng, J.; Zhang, N.Y.; Sun, L.H. Selenium deficiency aggravates aflatoxin b1-induced immunotoxicity in chick spleen by regulating 6 selenoprotein genes and redox/inflammation/apoptotic signaling. J. Nutr. 2019, 149, 19. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xiao, Z.H.; Liu, M.; Zhang, N.Y.; Sun, L.H. Dietary silymarin supplementation alleviates zearalenone-induced hepatotoxicity and reproductive toxicity in rats. J. Nutr. 2018, 148, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences (5′ → 3′) | Fragments Size (bp) | Accession Number |
|---|---|---|---|
| GAPDH | FP: GGAAGCTGTGGCGTGATGGC RP: TTCTCCAGGCGGCAGGTCAG | 175 | NM-001206359.1 |
| Nrf2 | FP: CCAATTCAGCCAGCACAACACATC RP: GACTGAGCCTGGTTAGGAGCAATG | 149 | XM_021075133.1 |
| Keap1 | FP: GGAGGACCACACCAAGCAAGC RP: GGATGAAGCCAGCACCACCTTG | 142 | NM-001114671.1 |
| CAT | FP: ACGCCTGTGTGAGAACATTG RP: GTCCAGAAGAGCCTGAATGC | 124 | NM_214301.2 |
| HO-1 | FP: AGGCTGAGAATGCCGAGTTC RP: TGTGGTACAAGGACGCCATC | 90 | NM_001004027.1 |
| NQO1 | FP: AGTATCCTGCCGAGACTGCTCTG RP: CACAAGGTCTGCGGCTTCCAC | 95 | NM-001159613.1 |
| TrxR1 | FP: GCTCAAGTGCGGACTGACCAAG RP: AGCAACCGGCCTGGAGGATG | 128 | NM_214154.3 |
| SOD1 | FP: GCGAGTCATGGCGACGAAGG RP: GACCTGCACTGGTACAGCCTTG | 191 | NM_001190422.1 |
| SOD2 | FP: TGTATCCGTCGGCGTCCAAGG RP: TCCTGGTTAGAACAAGCGGCAATC | 93 | NM_214127.2 |
| GPX1 | FP: GCGTCGCTCTGAGGCACAAC RP: GGTCGGACGTACTTGAGGCAATTC | 167 | NM_214201.1 |
| GPX2 | FP: ATTCTTCCTGGCTCCTCCTTCCTC RP: AGGCTGATAGCACTGAGGTCGTAG | 136 | NM-001115136.1 |
| GST | FP: GCCGAGGCAGAATGGAGTGTATC RP: GGTGGCGATGTAGTTGAGGATGG | 197 | NM-214389.2 |
| GSH-Px | FP: CGTGTAACCAGTTCGGACATCAGG RP: CGCCATTCACCTCACACTTCTCG | 129 | AJ010340.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Zhang, M.; Li, J.; Shan, A. DL-Selenomethionine Alleviates Oxidative Stress Induced by Zearalenone via Nrf2/Keap1 Signaling Pathway in IPEC-J2 Cells. Toxins 2021, 13, 557. https://doi.org/10.3390/toxins13080557
Sun H, Zhang M, Li J, Shan A. DL-Selenomethionine Alleviates Oxidative Stress Induced by Zearalenone via Nrf2/Keap1 Signaling Pathway in IPEC-J2 Cells. Toxins. 2021; 13(8):557. https://doi.org/10.3390/toxins13080557
Chicago/Turabian StyleSun, Haoyang, Meiling Zhang, Jianping Li, and Anshan Shan. 2021. "DL-Selenomethionine Alleviates Oxidative Stress Induced by Zearalenone via Nrf2/Keap1 Signaling Pathway in IPEC-J2 Cells" Toxins 13, no. 8: 557. https://doi.org/10.3390/toxins13080557
APA StyleSun, H., Zhang, M., Li, J., & Shan, A. (2021). DL-Selenomethionine Alleviates Oxidative Stress Induced by Zearalenone via Nrf2/Keap1 Signaling Pathway in IPEC-J2 Cells. Toxins, 13(8), 557. https://doi.org/10.3390/toxins13080557