Microbial Reduction of Fumonisin B1 by the New Isolate Serratia marcescens 329-2


Abstract
:1. Introduction
2. Results
2.1. Acclimatization and Isolation of Potential Fumonisin-Degrading Bacteria
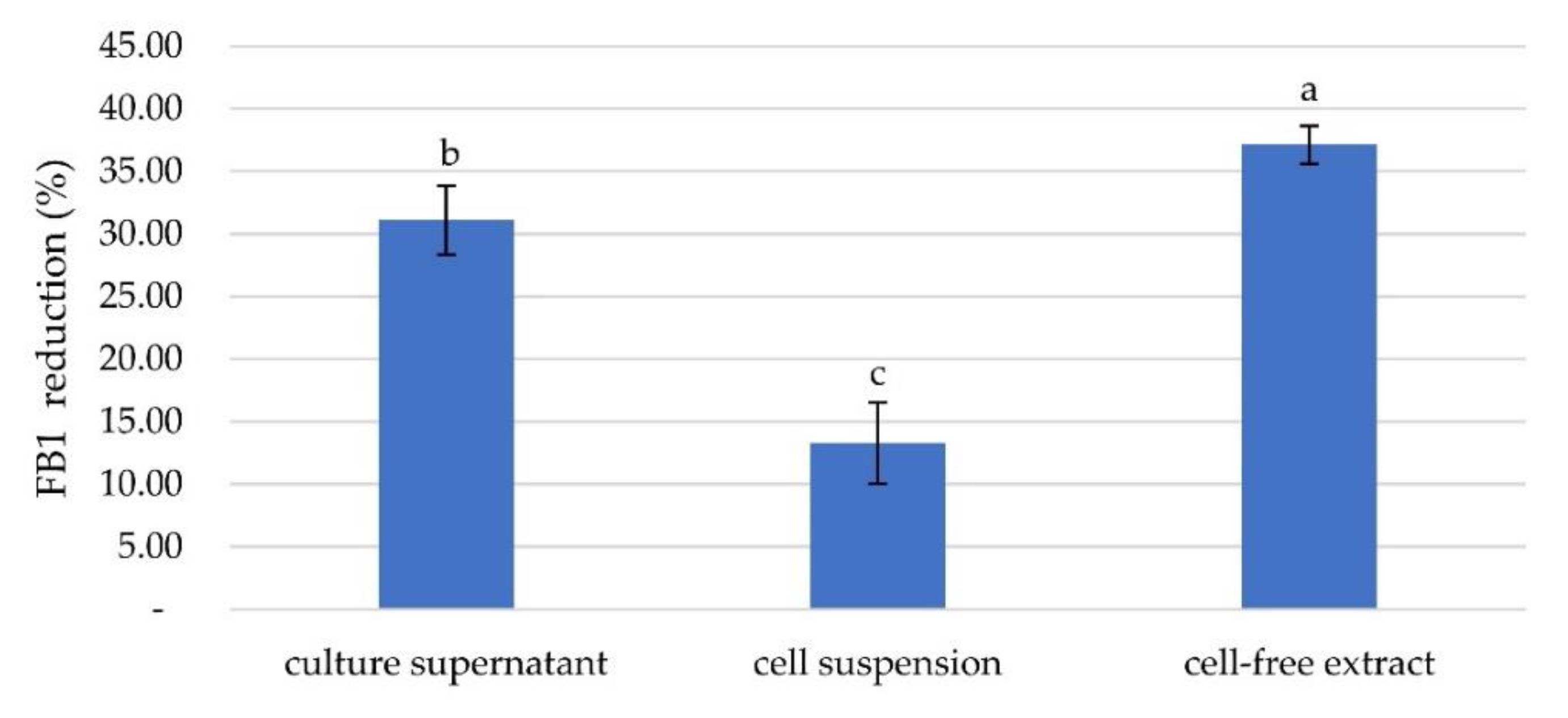
2.2. Fumonisin B1 Removal Activity by a Selected Bacterial Isolate and Determination of the Active Components
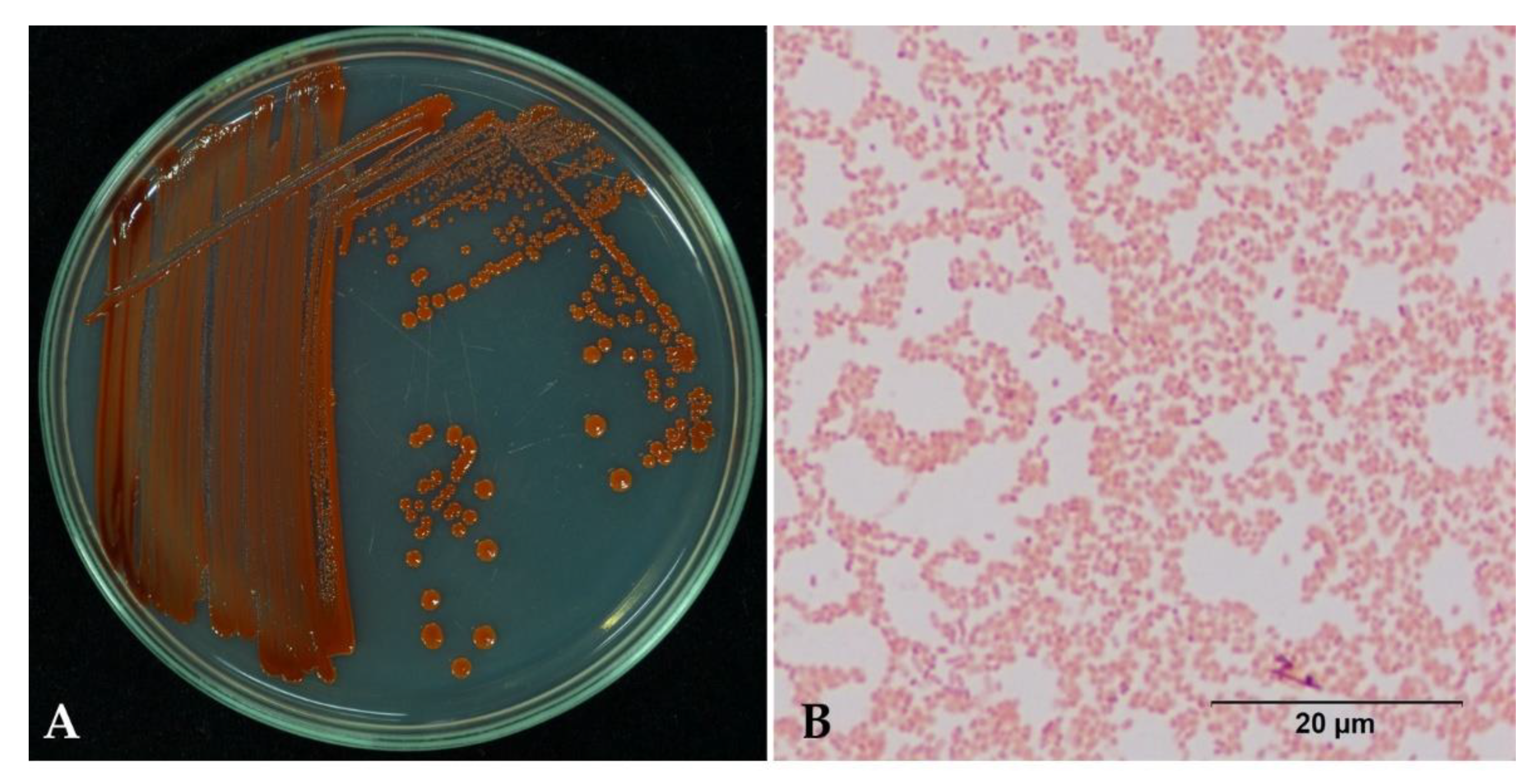
2.3. Bacterial Identification
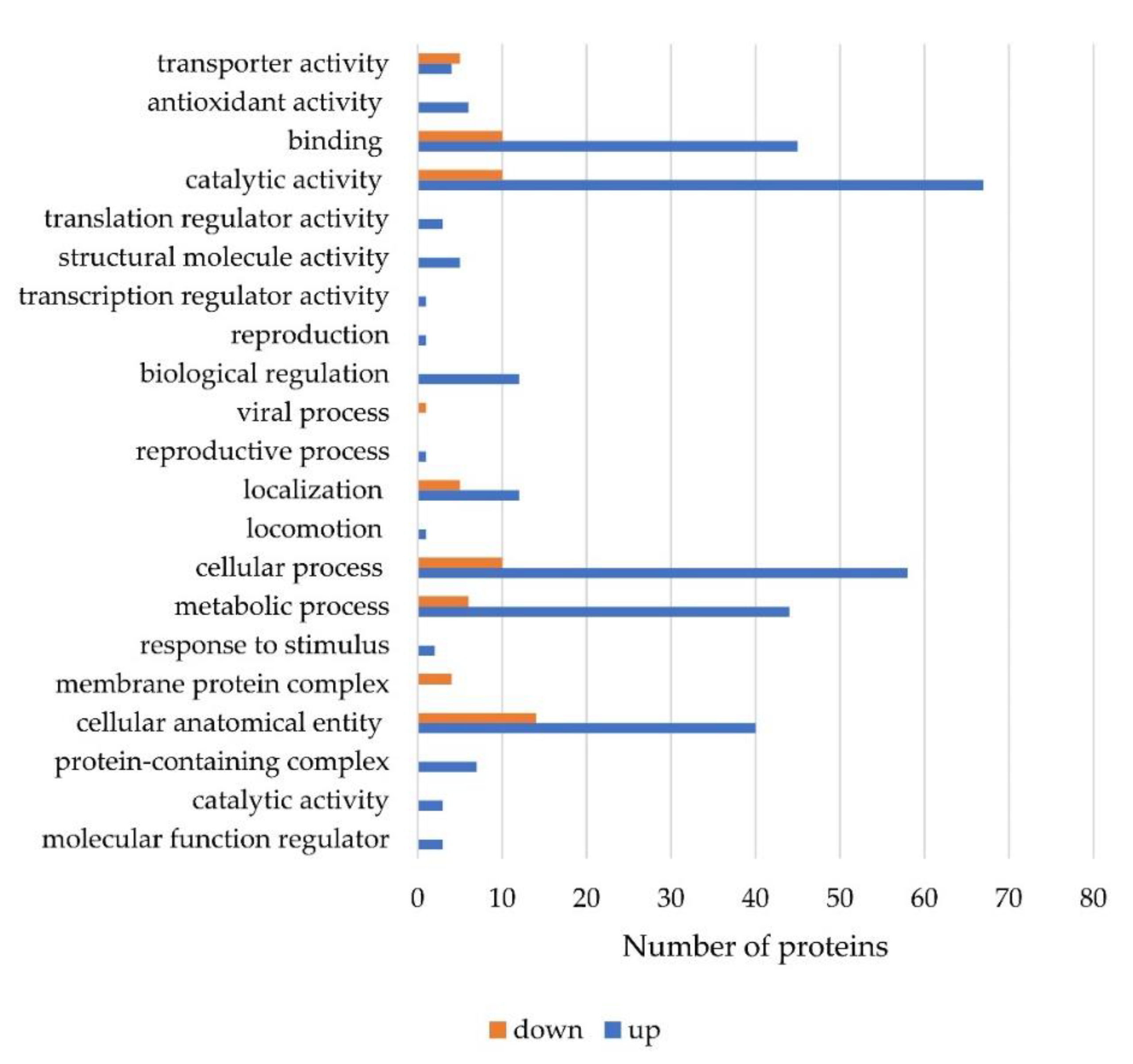
2.4. Proteins Expression during Fumonisin B1 Reduction by S. marcescens 329-2
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Acclimatization and Isolation of Potential Fumonisin-Degrading Bacteria
5.1.1. Acclimatization of Bacteria from Natural Resources
5.1.2. Isolation of Potential Fumonisin-Degrading Bacteria
5.2. Fumonisin B1 Removal Activity by a Selected Bacterial Isolate
5.3. Determination of the Active Components
5.4. Bacterial Identification
5.5. Proteins Expression during Fumonisin B1 Reduction by S. marcescens 329-2
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shibamoto, T.; Bjeldanes, L.F. Chapter 6-Fungal Toxins Occurring in Foods; Taylor, S.L., Ed.; Academic Press: San Diego, CA, USA, 1993; pp. 97–116. [Google Scholar]
- Pohland, A.E. Mycotoxins in review. Food Addit. Contam. 1993, 10, 17–28. [Google Scholar] [CrossRef]
- Ayalew, A. Mycotoxins and surface and internal fungi of maize from Ethiopia. Afr. J. Food Agric. Nutr. Dev. 2010, 10. [Google Scholar] [CrossRef]
- Tola, M.; Kebede, B.; Yildiz, F. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Hoboken, NJ, USA, 2006. [Google Scholar]
- EFSA. Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed. EFSA J. 2018, 16, e05242. [Google Scholar] [CrossRef]
- Rheeder, J.P.; Marasas, W.F.O.; Vismer, H.F. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [Green Version]
- Sydenham, E.W.; Shephard, G.S.; Thiel, P.G.; Marasas, W.F.O.; Stockenstrom, S. Fumonisin contamination of commercial corn-based human foodstuffs. J. Agric. Food Chem. 1991, 39, 2014–2018. [Google Scholar] [CrossRef]
- Presello, D.A.; Botta, G.; Iglesias, J.; Eyhérabide, G.H. Effect of disease severity on yield and grain fumonisin concentration of maize hybrids inoculated with Fusarium verticillioides. Crop Prot. 2008, 27, 572–576. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Desjardins, A.E. Fumonisins in maize: Can we reduce their occurrence? Plant Dis. 1997, 81, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biomin. BIOMIN World Mycotoxin Survey 2020. Available online: https://www.biomin.net/downloads/2020-biomin-world-mycotoxin-survey-report/#c41851 (accessed on 15 May 2021).
- Zhao, Z.; Zhang, Y.; Gong, A.; Liu, N.; Chen, S.; Zhao, X.; Li, X.; Chen, L.; Zhou, C.; Wang, J. Biodegradation of mycotoxin fumonisin B1 by a novel bacterial consortium SAAS79. Appl. Microbiol. Biotechnol. 2019, 103, 7129–7140. [Google Scholar] [CrossRef] [PubMed]
- Lyagin, I.; Efremenko, E. Enzymes for detoxification of various mycotoxins: Origins and mechanisms of catalytic action. Molecules 2019, 24, 2362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.; Zhou, T. Strategies and methodologies for developing microbial detoxification systems to mitigate mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mulè, G. Mycotoxin biotransformation by native and commercial enzymes: Present and future perspectives. Toxins 2017, 9, 111. [Google Scholar] [CrossRef]
- Duvick, J.; Rood, T.; Maddox, J.; Gilliam, J. Detoxification of mycotoxins in planta as a strategy for improving grain quality and disease resistance: Identification of fumonisin-degrading microbes from maize. In Molecular Genetics of Host-Specific Toxins in Plant Disease: Proceedings of the 3rd Tottori International Symposium on Host-Specific Toxins, Daisen, Tottori, Japan, August 24–29, 1997; Kohmoto, K., Yoder, O.C., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 369–381. [Google Scholar]
- Blackwell, B.A.; Gilliam, J.; Savard, M.; Miller, J.; Duvick, J. Oxidative deamination of hydrolyzed fumonisin B1 (AP1) by cultures of Exophiala spinifera. Nat. Toxins 1999, 7, 31–38. [Google Scholar] [CrossRef]
- Collins, T.F.; Sprando, R.L.; Black, T.N.; Olejnik, N.; Eppley, R.M.; Shackelford, M.E.; Howard, P.C.; Rorie, J.I.; Bryant, M.; Ruggles, D.I. Effects of aminopentol on in utero development in rats. Food Chem. Toxicol. 2006, 44, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, R.; Nazzi, F.; Locci, R.; Firrao, G. Degradation of fumonisin B1 by a bacterial strain isolated from soil. Biodegradation 2006, 17, 31–38. [Google Scholar] [CrossRef]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Vekiru, E.; Krska, R.; Schatzmayr, G.; Moll, W.-D.; Grabherr, R. Degradation of fumonisin B1 by the consecutive action of two bacterial enzymes. J. Biotechnol. 2010, 145, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Masching, S.; Naehrer, K.; Schwartz-Zimmermann, H.E.; Sarandan, M.; Schaumberger, S.; Dohnal, I.; Nagl, V.; Schatzmayr, D. Gastrointestinal degradation of fumonisin B(1) by carboxylesterase FumD prevents fumonisin induced alteration of sphingolipid metabolism in turkey and swine. Toxins 2016, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.; Bottalico, A.; Mulé, G.; Moretti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some mediterranean crops. Eur. J. Plant Pathol. 2003, 109, 645–667. [Google Scholar] [CrossRef]
- Voss, K.A.; Riley, R.T. Fumonisin toxicity and mechanism of action: Overview and current perspectives. Food Saf. 2013, 1, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Medina, Á.; González-Jartín, J.M.; Sainz, M.J. Impact of global warming on mycotoxins. Curr. Opin. Food Sci. 2017, 18, 76–81. [Google Scholar] [CrossRef]
- Niderkorn, V.; Morgavi, D.P.; Pujos, E.; Tissandier, A.; Boudra, H. Screening of fermentative bacteria for their ability to bind and biotransform deoxynivalenol, zearalenone and fumonisins in an in vitro simulated corn silage model. Food Addit. Contam. 2007, 24, 406–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalie, D.K.; Deschamps, A.M.; Atanasova-Penichon, V.; Richard-Forget, F. Potential of Pediococcus pentosaceus (L006) isolated from maize leaf to suppress fumonisin-producing fungal growth. J. Food Prot. 2010, 73, 1129–1137. [Google Scholar] [CrossRef]
- Deepthi, B.V.; Poornachandra Rao, K.; Chennapa, G.; Naik, M.K.; Chandrashekara, K.T.; Sreenivasa, M.Y. Antifungal attributes of Lactobacillus plantarum MYS6 against fumonisin producing Fusarium proliferatum associated with poultry feeds. PLoS ONE 2016, 11, e0155122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, B.; Bresch, H.; Schillinger, U.; Thiel, P.G. The effect of fumonisin B1 on the growth of bacteria. World J. Microbiol. Biotechnol. 1997, 13, 539–543. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, C.; Zhang, D.; Zhao, M.; Zheng, D.; Lyu, Y.; Cheng, W.; Guo, P.; Cui, Z. Effective degradation of aflatoxin B(1) using a novel thermophilic microbial consortium TADC7. Bioresour. Technol. 2017, 224, 166–173. [Google Scholar] [CrossRef]
- John, G.H.; Noel, R.K.; Peter, H.A.S.; James, T.S.; Stanly, T.W. Bergey’s Manual of Determinative Bacteriology; Williams & Wilkins: Baltimore, MD, USA, 1994. [Google Scholar]
- Li, B.; Yu, R.; Liu, B.; Tang, Q.; Zhang, G.; Wang, Y.; Xie, G.; Sun, G. Characterization and comparison of Serratia marcescens isolated from edible cactus and from silkworm for virulence potential and chitosan susceptibility. Braz. J. Microbiol. 2011, 42, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.-Q.; Liu, Z.-Q.; Zheng, Y.-G.; Shen, Y.-C. Identification and characterization of Serratia marcescens ZJB-09104, a nitrile-converting bacterium. World J. Microbiol. Biotechnol. 2010, 26, 817–823. [Google Scholar] [CrossRef]
- Patel, J.B. 16S rRNA gene sequencing for bacterial pathogen identification in the clinical laboratory. Mol. Diagn. 2001, 6, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Woese, C.R. Bacterial evolution. Microbiol. Rev. 1987, 51, 221–271. [Google Scholar] [CrossRef]
- Pandey, A.K.; Chaudhary, P.; Singh, S.B.; Arora, A.; Kumar, K.; Chaudhry, S.; Nain, L. Deciphering the traits associated with PAH degradation by a novel Serratia marcesencs L-11 strain. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2012, 47, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Stancu, M.M. Highly solvent tolerance in Serratia marcescens IBBPo15. Braz. Arch. Biol. Technol. 2016, 59. [Google Scholar] [CrossRef]
- Neville, S.A.; LeCordier, A.; Ziochos, H.; Chater, M.J.; Gosbell, I.B.; Maley, M.W.; van Hal, S.J. Utility of matrix-assisted laser desorption ionization–time of flight mass spectrometry following introduction for routine laboratory bacterial identification. J. Clin. Microbiol. 2011, 49, 2980–2984. [Google Scholar] [CrossRef] [Green Version]
- Timperio, A.M.; Gorrasi, S.; Zolla, L.; Fenice, M. Evaluation of MALDI-TOF mass spectrometry and MALDI biotyper in comparison to 16S rDNA sequencing for the identification of bacteria isolated from Arctic sea water. PLoS ONE 2017, 12, e0181860. [Google Scholar] [CrossRef] [PubMed]
- Sauget, M.; Valot, B.; Bertrand, X.; Hocquet, D. Can MALDI-TOF mass spectrometry reasonably type bacteria? Trends Microbiol. 2017, 25, 447–455. [Google Scholar] [CrossRef]
- Li, Y.; Shan, M.; Zhu, Z.; Mao, X.; Yan, M.; Chen, Y.; Zhu, Q.; Li, H.; Gu, B. Application of MALDI-TOF MS to rapid identification of anaerobic bacteria. BMC Infect. Dis. 2019, 19, 941. [Google Scholar] [CrossRef] [Green Version]
- Dingle, T.C.; Butler-Wu, S.M. Maldi-tof mass spectrometry for microorganism identification. Clin. Lab. Med. 2013, 33, 589–609. [Google Scholar] [CrossRef]
- Rödel, J.; Mellmann, A.; Stein, C.; Alexi, M.; Kipp, F.; Edel, B.; Dawczynski, K.; Brandt, C.; Seidel, L.; Pfister, W.; et al. Use of MALDI-TOF mass spectrometry to detect nosocomial outbreaks of Serratia marcescens and Citrobacter freundii. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Othman, M.A.; El-Zamik, F.I.; Hegazy, M.I.; Salama, A.S.A. Isolation and identification of egyptian strains of Serratia marcescens producing antibacterial and antioxidant prodigiosin pigment. Zagazig J. Agric. Res. 2019, 46, 1573–1582. [Google Scholar] [CrossRef]
- Ng, L.S.Y.; Sim, J.H.C.; Eng, L.C.; Menon, S.; Tan, T.Y. Comparison of phenotypic methods and matrix-assisted laser desorption ionisation time-of-flight mass spectrometry for the identification of aero-tolerant Actinomyces spp. isolated from soft-tissue infections. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1749–1752. [Google Scholar] [CrossRef] [PubMed]
- Someya, N.; Kataoka, N.; Komagata, T.; Hirayae, K.; Hibi, T.; Akutsu, K. Biological control of cyclamen soilborne diseases by Serratia marcescens strain B2. Plant Dis. 2000, 84, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, H.; Sato, M.; Sato, Z.; Isaka, M. Biocontrol of Phytophthora capsici by Serratia marcescens F-1-1 and analysis of biocontrol mechanisms using transposon-insertion mutants. Jpn. J. Phytopathol. 1998, 64, 287–293. [Google Scholar] [CrossRef]
- Jaiganesh, V.; Eswaran, A.; Balabaskar, P.; Kannan, C. Antagonistic activity of Serratia marcescens against Pyricularia oryzae. Not. Bot. Horti Agrobot. Cluj. Napoca 2007, 35. [Google Scholar] [CrossRef]
- Press, C.M.; Loper, J.E.; Kloepper, J.W. Role of iron in rhizobacteria-mediated induced systemic resistance of cucumber. Phytopathology 2001, 91, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, X.; Wu, J.; Yu, J.; Xu, M.; Chen, D.; Zhang, Z.; Li, X.; Chi, Y.; Wan, S. In vitro inhibitory effect of the bacterium Serratia marcescens on Fusarium proliferatum growth and fumonisins production. Biol. Control. 2020, 143, 104188. [Google Scholar] [CrossRef]
- Carbonell, G.V.; Della Colleta, H.H.M.; Yano, T.; Darini, A.L.C.; Levy, C.E.; Fonseca, B.A.L. Clinical relevance and virulence factors of pigmented Serratia marcescens. FEMS Immunol. Med. Microbiol. 2000, 28, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, D.; Heinl, S.; Schwartz, H.E.; Grabherr, R.; Schatzmayr, G.; Haltrich, D.; Moll, W.-D. Enhancement of solubility in Escherichia coli and purification of an aminotransferase from Sphingopyxis sp. MTA144 for deamination of hydrolyzed fumonisin B(1). Microb. Cell Factories 2010, 9, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devlin, T.M. Textbook of Biochemistry; John Wiley: New York, NY, USA, 2002. [Google Scholar]
- McKee, T.; McKee, J.R. Biochemistry; McGraw-Hill: Boston, MA, USA, 2003. [Google Scholar]
- Montella, I.R.; Schama, R.; Valle, D. The classification of esterases: An important gene family involved in insecticide resistance-A review. Mem. Inst. Oswaldo Cruz. 2012, 107, 437–449. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (2006/576/EC). Off. J. Eur. Union 2006, L229, 7–9. [Google Scholar]
- Wang, J.Q.; Yang, F.; Yang, P.L.; Liu, J.; Lv, Z.H. Microbial reduction of zearalenone by a new isolated Lysinibacillus sp. ZJ-2016-1. World Mycotoxin J. 2018, 11, 571–578. [Google Scholar] [CrossRef]
- Davies, J.A.; Anderson, G.K.; Beveridge, T.J.; Clark, H.C. Chemical mechanism of the gram stain and synthesis of a new electron-opaque marker for electron microscopy which replaces the iodine mordant of the stain. J. Bacteriol. 1983, 156, 837–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Turner, S.; Pryer, K.M.; Miao, V.P.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Source | Number of Screened Samples | Potential Degrading Samples |
|---|---|---|
| Maize | 37 | 4 |
| Rice | 12 | 0 |
| Soil | 8 | 0 |
| Fermented fluid | 38 | 1 |
| Total | 95 | 5 |
| Rank | Quality | Matched Pattern | Score |
|---|---|---|---|
| 1 | +++ | Serratia marcescens DSM 12481 DSM | 2.361 |
| 2 | +++ | Serratia marcescens DSM 12485 DSM | 2.345 |
| 3 | +++ | Serratia marcescens 13103_1 CHB | 2.319 |
| 4 | +++ | Serratia marcescens subsp. marcescens DSM 30121T DSM | 2.306 |
| 5 | ++ | Serratia marcescens subsp. sakuensis CIP 107489T HAM | 2.240 |
| 6 | ++ | Serratia ureilytica DSM 16952T DSM | 2.155 |
| 7 | ++ | Serratia marcescens DSM 30122 DSM | 2.147 |
| 8 | ++ | Serratia marcescens (PX) 24086109 MLD | 2.062 |
| 9 | ++ | Serratia marcescens DSM 12483 DSM | 2.039 |
| 10 | + | Serratia entomophila DSM 12358T DSM | 1.942 |
| Entry | Protein Names | Gene Names | Fold Change |
|---|---|---|---|
| A0A6G9UZ48 | ABC transporter substrate-binding protein | HCG50_10660 | 8.20 |
| A0A6N0D898 | Porin OmpC | ompC | 7.94 |
| A0A3E2ENK9 | Amino acid ABC transporter substrate-binding protein | gltI | 7.12 |
| V5YV29 | Maltodextrin-binding protein | malE | 6.89 |
| A0A080V044 | Universal stress protein | uspA | 5.42 |
| A0A6M5HVT2 | 4-hydroxy-tetrahydrodipicolinate synthase | dapA | 5.33 |
| A0A6G9UQE2 | ABC transporter substrate-binding protein | HCG50_08880 | 5.33 |
| A0A1Q4NZ53 | Superoxide dismutase | BHU62_14220 | 4.99 |
| A0A6G9UU24 | Phosphate-binding protein PstS | pstS | 4.98 |
| A0A6N3ZRH0 | Fumarylacetoacetate hydrolase family protein | G3M84_1332 | 4.91 |
| A0A656VL53 | Alpha/beta hydrolase | AB868_03825 | 4.90 |
| A0A6M5HYD0 | Phenylacetate-CoA oxygenase/reductase subunit PaaK | paaK | 4.78 |
| A0A3E2EF40 | Malate dehydrogenase | mdh | 4.68 |
| A0A6N0D0Q5 | Neutral metalloproteinase | F0335_18215 | 4.49 |
| A0A086FBX8 | Transcription termination/antitermination protein NusG | nusG | 4.39 |
| A0A2V4FJ05 | Peptide deformylase | def | 4.35 |
| V5YU98 | Extracellular solute-binding protein | E4655_11925 | 4.27 |
| A0A5Q8BY15 | YtfJ family protein | EGJ31_19890 | 4.20 |
| A0A1Q5WAZ4 | Oligopeptide ABC transporter substrate-binding protein OppA | A8A12_03045 | 4.18 |
| A0A6H1E4N5 | Acetylornithine/succinyldiaminopimelate aminotransferase | argD | 4.10 |
| A0A6N3ZYZ9 | 2,3-diphosphoglycerate-dependent phosphoglycerate mutase | gpmA | 4.06 |
| A0A0P0Q8S3 | ABC transporter substrate-binding protein | AR325_02675 | 4.06 |
| A0A6N0CVJ7 | Superoxide dismutase | sodB | 4.03 |
| A0A5Q8C0J8 | Organic hydroperoxide resistance protein | EGJ31_14360 | 4.02 |
| V5YUS0 | Periplasmic serine endoprotease DegP-like | degQ | 4.01 |
| Q6MXC8 | Methyltransferase | SMR0272 | 3.94 |
| A0A2S4XAJ8 | Surface composition regulator | glgS | 3.89 |
| A0A1Q5WEW3 | Antibiotic biosynthesis monooxygenase | A8A12_06980 | 3.87 |
| A0A6I4GZS8 | Hydrolase | GMA22_24835 | 3.80 |
| A0A221FKL4 | UPF0234 protein BVG93_01845 | BVG93_01845 | 3.75 |
| A0A6H3S2C0 | ATP-dependent protease subunit HslV | hslV | 3.71 |
| A0A6M5I193 | MBL fold metallo-hydrolase | HMI62_20840 | 3.64 |
| A0A6N0DB57 | Protein deglycase HchA | hchA | 3.62 |
| A0A2V4G7I4 | Amino acid ABC transporter substrate-binding protein | glnH | 3.62 |
| A0A5C7CH16 | VOC family protein | FOT62_15570 | 3.58 |
| A0A0G8B4P9 | Peptidyl-prolyl cis-trans isomerase | fkpA | 3.58 |
| A0A656VU86 | Long-chain fatty acid transport protein | AB868_00683 | 3.55 |
| A0A2S4 × 857 | Histidine ABC transporter substrate-binding protein HisJ | hisJ | 3.47 |
| A0A656V5R8 | 5-methyltetrahydropteroyltriglutamate--homocysteine methyltransferase | metE | 3.46 |
| A0A0G8BFE1 | Cystine ABC transporter substrate-binding protein | tcyJ | 3.43 |
| A0A1C3HIZ5 | Aconitate hydratase B | acnB | 3.35 |
| A0A0U6KIH4 | GTP cyclohydrolase 1 | folE | 3.34 |
| A0A6N0CW48 | Branched-chain amino acid ABC transporter substrate-binding protein | F0335_15805 | 3.32 |
| A0A6N3ZXZ8 | ABC transporter substrate-binding protein | G3M84_09620 | 3.32 |
| A0A6G8TTH4 | Autoinducer 2-binding protein LsrB | G5643_21680 | 3.29 |
| A0A656VPU2 | Uncharacterized protein | AB868_00798 | 3.26 |
| A0A1C3HHX7 | Nitrogen regulatory protein P-II | glnB | 3.23 |
| A0A1Q5WH71 | Thiol:disulfide interchange protein | dsbA | 3.22 |
| A0A0G8B466 | 2-dehydro-3-deoxygluconokinase | AR325_02155 | 3.22 |
| A0A0M5K334 | Transaldolase | tal | 3.18 |
| A0A080UWJ0 | Peptidyl-prolyl cis-trans isomerase | fklB | 3.15 |
| A0A6N0D450 | Two-component system response regulator BaeR | baeR | 3.13 |
| A0A6N0CZA0 | Glucose-6-phosphate isomerase | pgi | 3.10 |
| A0A0F6KTS7 | 2-dehydro-3-deoxy-phosphogluconate aldolase | eda | 3.10 |
| V5YUY6 | Stringent starvation protein A | sspA | 3.09 |
| A0A6M5HTX8 | Uncharacterized protein | HMI62_14785 | 3.08 |
| A0A1Q4NZT3 | DUF1471 domain-containing protein | BHU62_12635 | 3.03 |
| A0A086FJA0 | 50S ribosomal protein L9 | rplI | 3.02 |
| Entry | Protein Names | Gene Names | Fold Change |
|---|---|---|---|
| A0A514I8F1 | DUF3251 domain-containing protein | FG174_04965 | 0.50 |
| A0A086FBJ7 | Outer membrane protein assembly factor BamD | bamD | 0.48 |
| A0A6G8TR73 | Outer membrane lipoprotein RcsF | rcsF | 0.48 |
| A0A080UZC4 | Tol-Pal system protein TolR | tolR | 0.48 |
| A0A2V4H3Z0 | Ribose-phosphate pyrophosphokinase | prs | 0.47 |
| V5YU02 | Divisome-associated lipoprotein YraP | yraP | 0.47 |
| A0A0M4S0F9 | Glutamine synthetase | glnA | 0.47 |
| A0A379ZG33 | Phage shock protein A | pspA | 0.47 |
| Q5J5B8 | Outer membrane protein assembly factor BamC | nlpbsm | 0.46 |
| A0A379ZA39 | Outer membrane protein slp | slp | 0.46 |
| A0A080V8M2 | Heat shock chaperone IbpB | ibpB | 0.46 |
| A0A6I4HK57 | Outer membrane protein assembly factor BamB | bamB | 0.46 |
| A0A0P0QBU0 | Glycoprotein-polysaccharide metabolism protein | A8A12_17600 | 0.46 |
| A0A6I6ZSG9 | ATP synthase subunit alpha | atpA | 0.46 |
| A0A6N0CYE2 | Phosphate acetyltransferase | pta | 0.45 |
| A0A6N0CUX7 | Glycine-tRNA ligase subunit beta | glyS | 0.43 |
| A0A1Q5WD75 | NAD(P)H dehydrogenase (quinone) | wrbA | 0.42 |
| A0A6I6ZPJ4 | Terminase | GV243_19590 | 0.40 |
| A0A3E2EPF1 | Outer membrane | ompA | 0.39 |
| A0A1Q4NYR4 | ATP synthase gamma chain | atpG | 0.38 |
| A0A6M5IGG0 | SPOR domain-containing protein | HMI62_24000 | 0.35 |
| A0A1Q4P2F3 | Fructose-1,6-bisphosphatase class 1 | fbp | 0.34 |
| A0A6H3SAN4 | Glyceraldehyde-3-phosphate dehydrogenase | gapA | 0.34 |
| A0A0M3UJK8 | ATP synthase subunit beta | atpD | 0.30 |
| A0A6G8TKN9 | Uncharacterized protein | Uncharacterized protein | 0.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keawmanee, P.; Rattanakreetakul, C.; Pongpisutta, R. Microbial Reduction of Fumonisin B1 by the New Isolate Serratia marcescens 329-2. Toxins 2021, 13, 638. https://doi.org/10.3390/toxins13090638
Keawmanee P, Rattanakreetakul C, Pongpisutta R. Microbial Reduction of Fumonisin B1 by the New Isolate Serratia marcescens 329-2. Toxins. 2021; 13(9):638. https://doi.org/10.3390/toxins13090638
Chicago/Turabian StyleKeawmanee, Pisut, Chainarong Rattanakreetakul, and Ratiya Pongpisutta. 2021. "Microbial Reduction of Fumonisin B1 by the New Isolate Serratia marcescens 329-2" Toxins 13, no. 9: 638. https://doi.org/10.3390/toxins13090638
APA StyleKeawmanee, P., Rattanakreetakul, C., & Pongpisutta, R. (2021). Microbial Reduction of Fumonisin B1 by the New Isolate Serratia marcescens 329-2. Toxins, 13(9), 638. https://doi.org/10.3390/toxins13090638