Biological Transformation of Zearalenone by Some Bacterial Isolates Associated with Ruminant and Food Samples

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
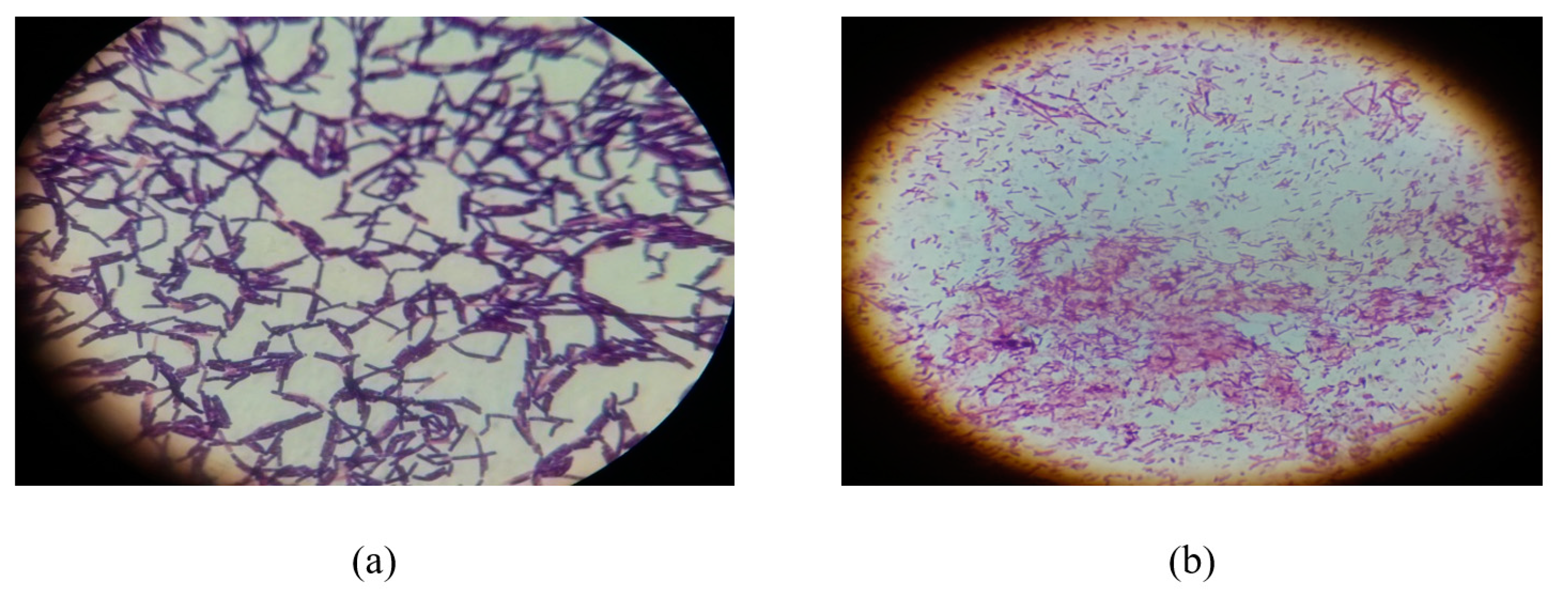
2.1. Gram’s Staining and Morphological Characterization
2.2. Biochemical Identification
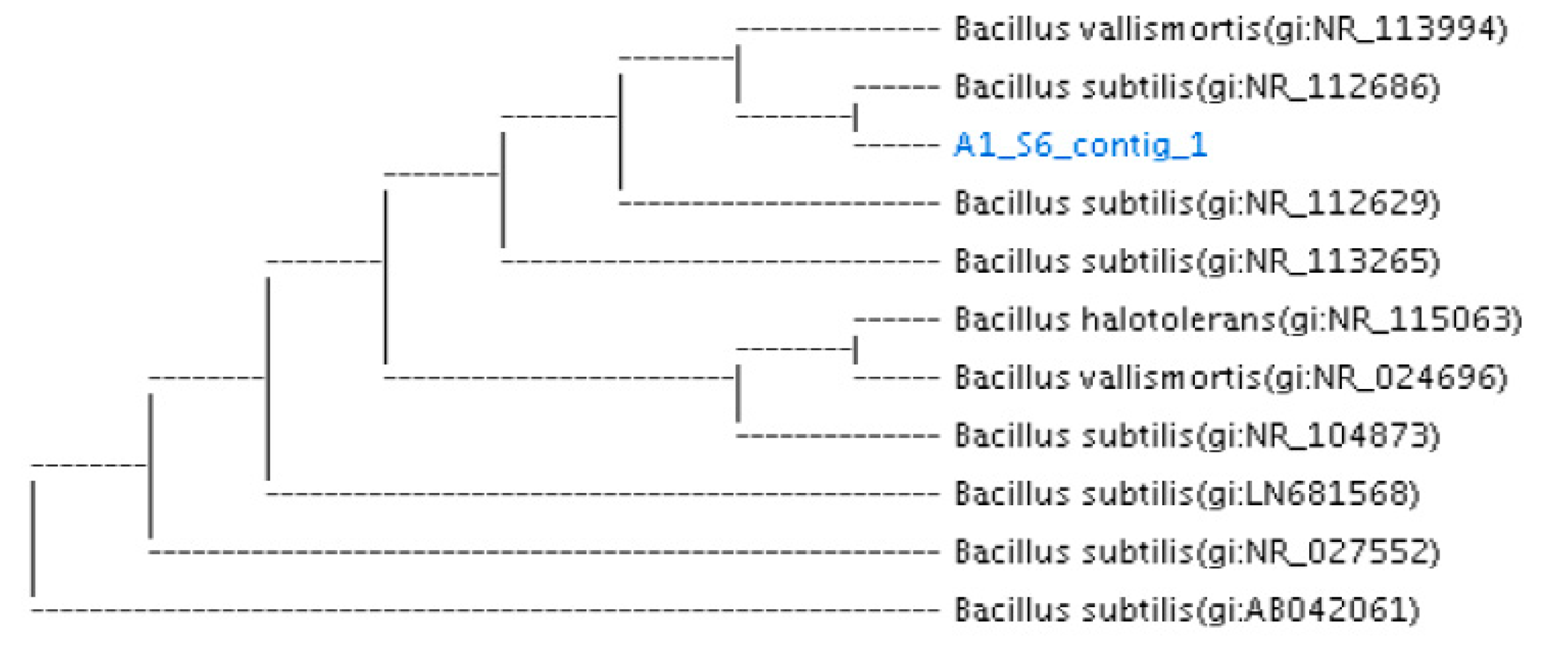
2.3. Molecular Identification
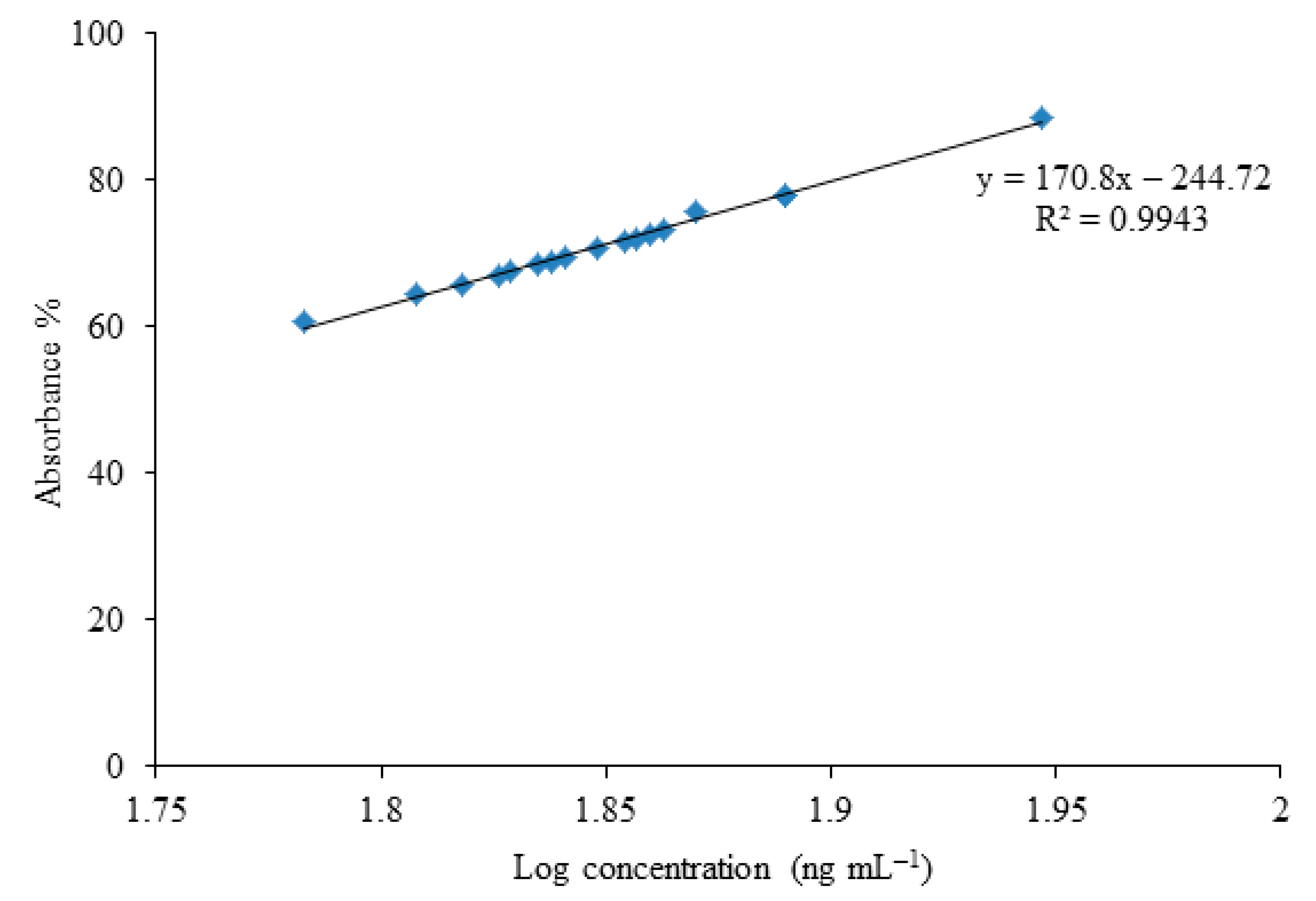
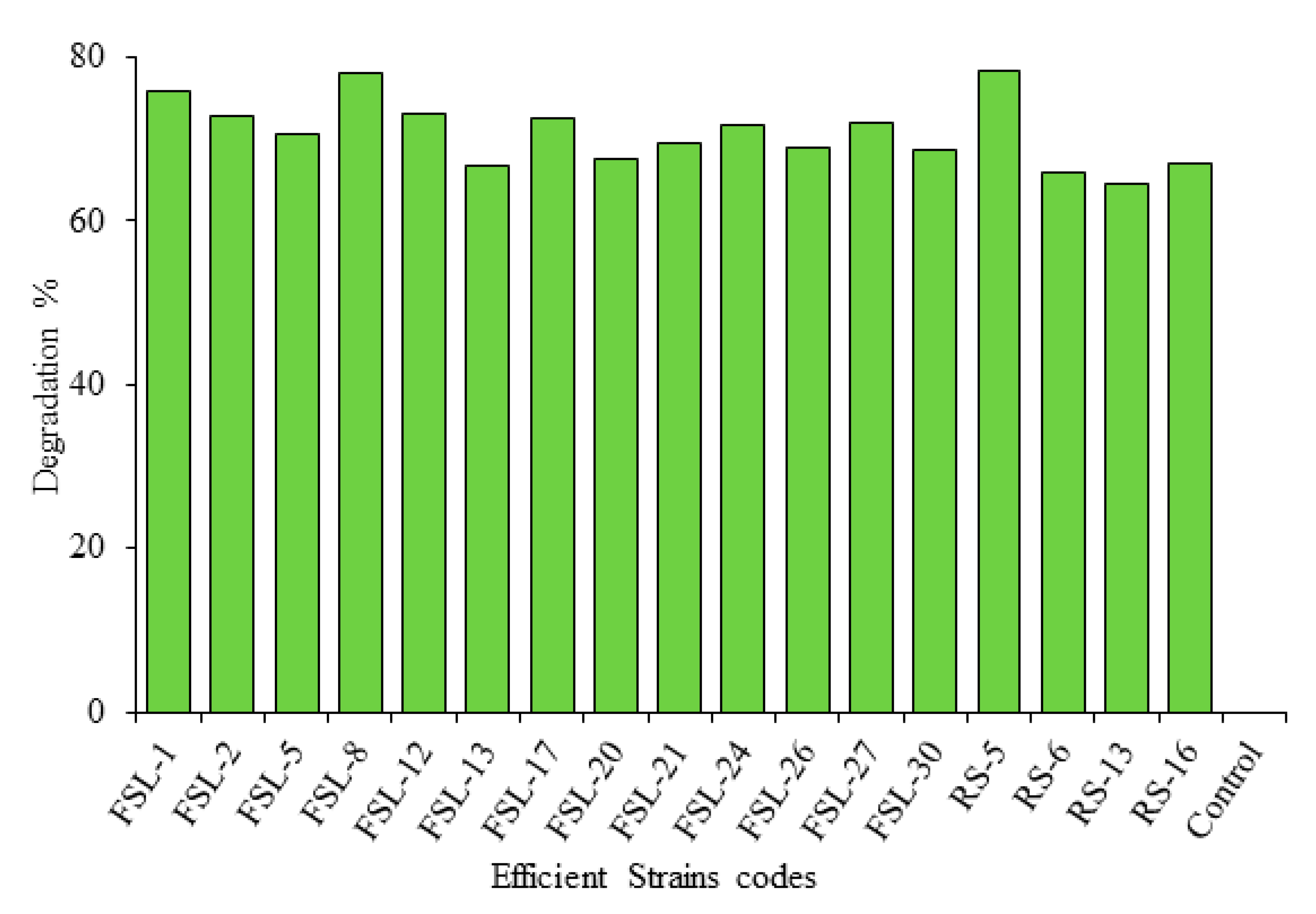
2.4. Screening of Zearalenone Biodegrading Bacterial Strains through ELISA
Detection of Zearalenone through ELISA
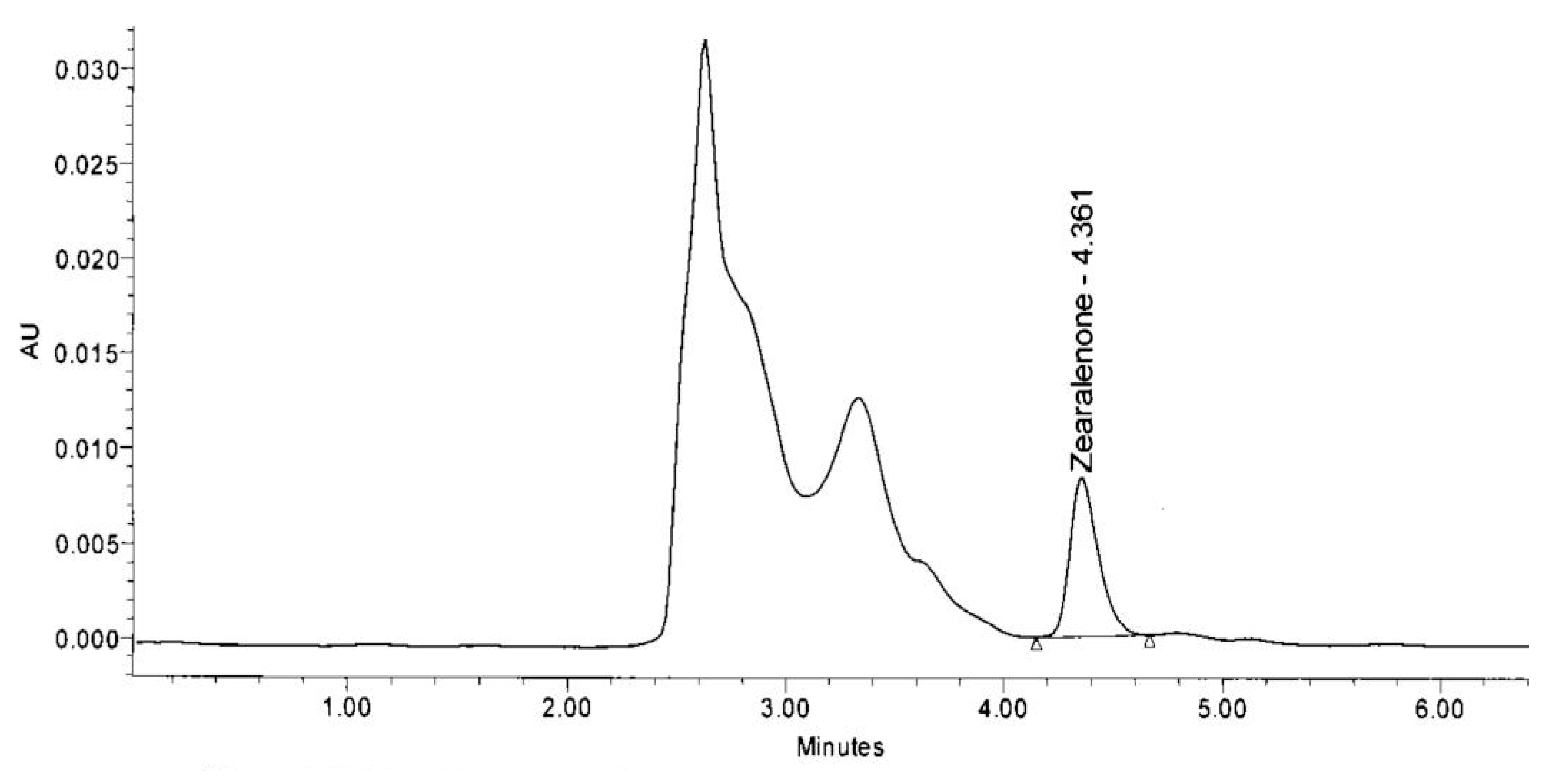
2.5. Analysis of Bio-Transforming Ability of Bacterial Isolates through HPLC
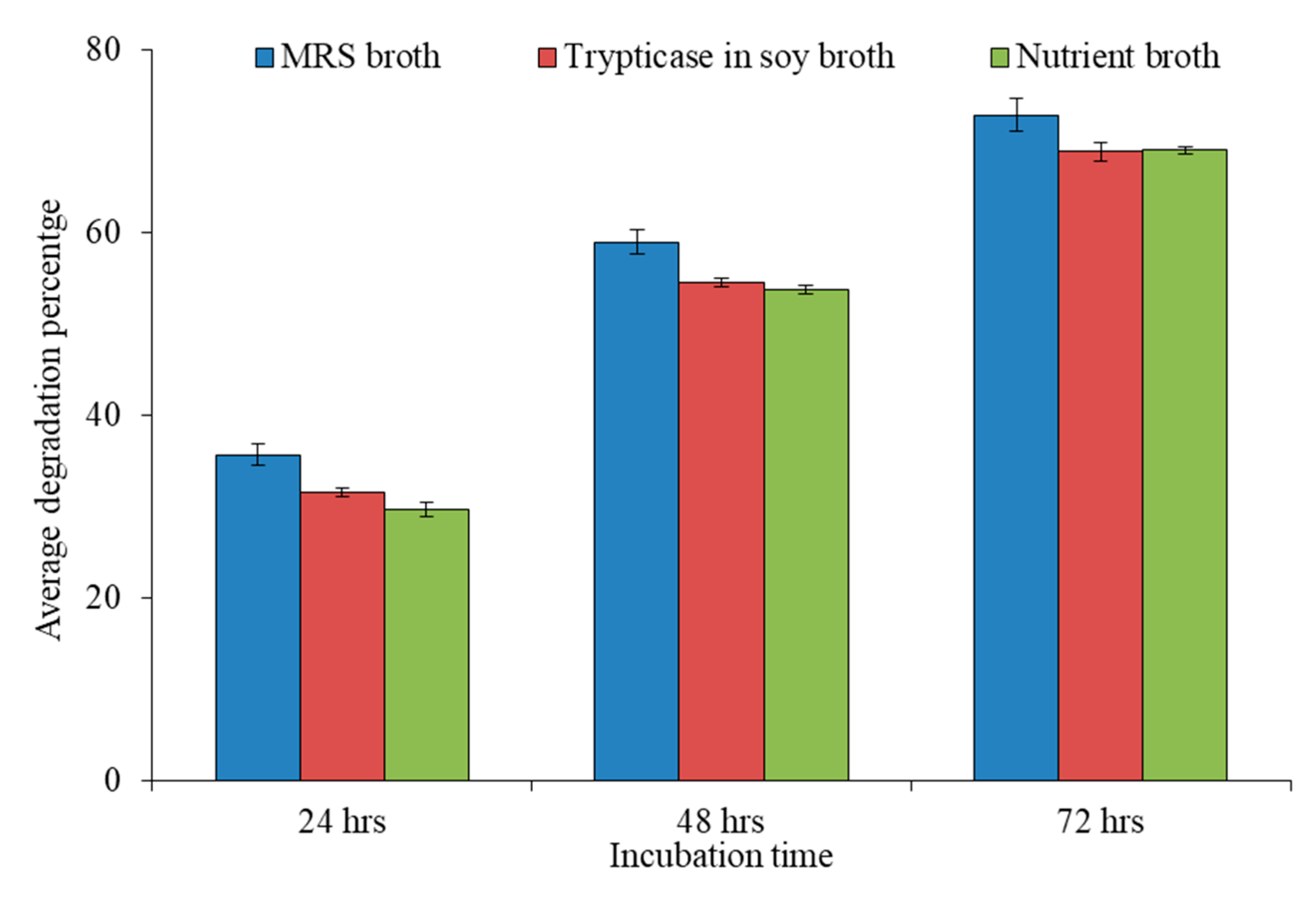
2.6. t-Test:/Statistical Analysis
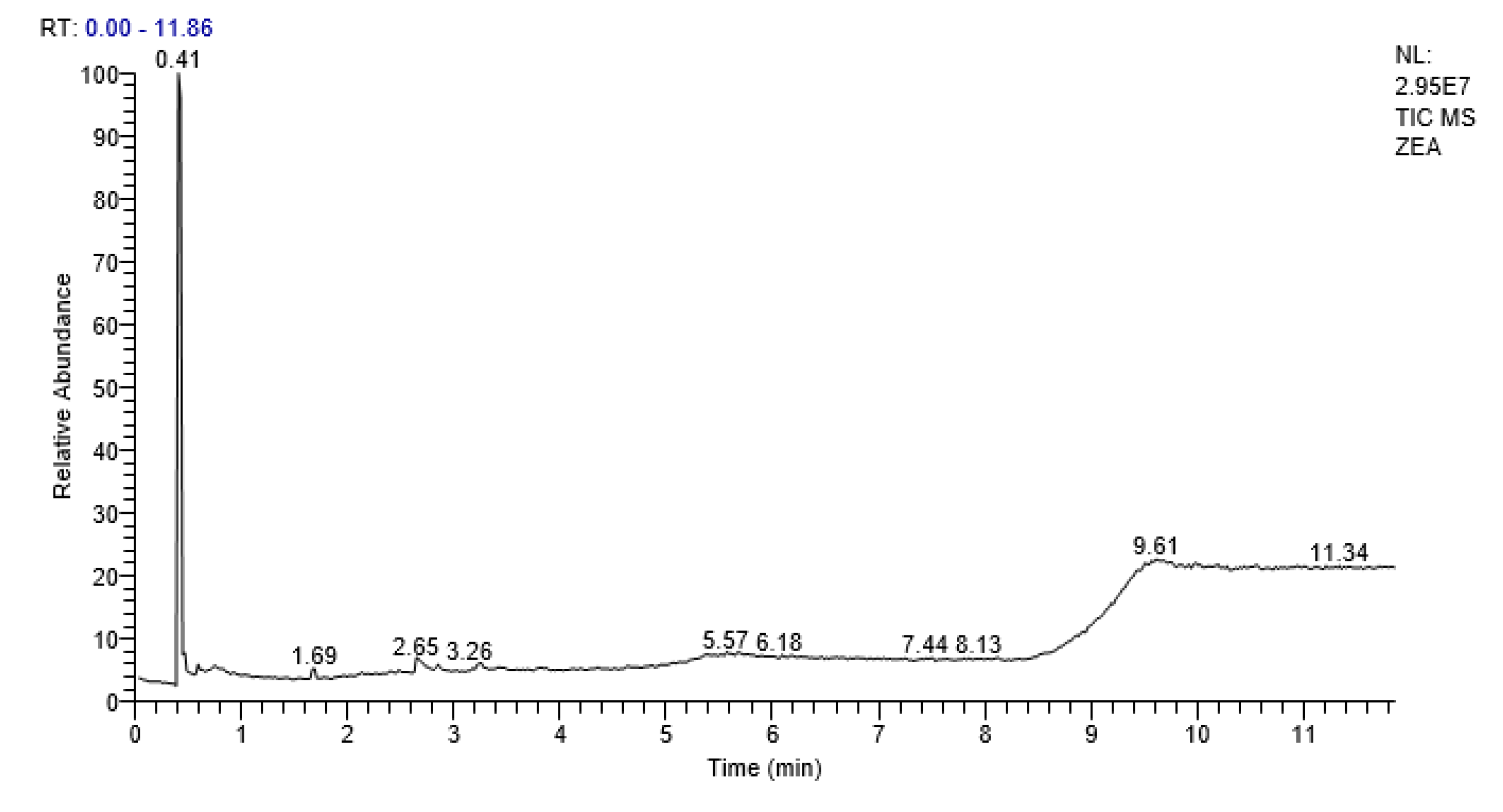
2.7. GCMS Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents Used in Current Research Study
5.2. Stock and Working Solution Preparation for ELISA and HPLC
5.3. Collection of Samples and Isolation of Bacteria
5.4. Microbial Identification
Molecular Identification
5.5. Analysis of ZEA Reduction Ability of Isolated Bacteria through ELISA
5.6. Analysis of ZEA Reduction Ability of Bacterial Isolates through HPLC
HPLC Conditions
5.7. Gas Chromatography-Mass Spectrometry
5.7.1. Sample Preparation and Zearalenone Extraction
5.7.2. GCMS Condition
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations, and intake of Zearalenone, An oestragenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Hestbjerg, H.; Nielsen, K.F.; Thrane, U.; Elmholt, S. Production of trichothecenes and other secondary metabolites by Fusarium culmorum and Fusarium equiseti on common laboratory media and a soil organic matter agar: An ecological interpretation. J. Agric. Food Chem. 2002, 50, 7593–7599. [Google Scholar] [CrossRef]
- Goliński, P.; Waśkiewicz, A.; Gromadzka, K. Mycotoxins and mycotoxicoses under climatic conditions of Poland. Pol. J. Vet. Sci. 2009, 12, 581–588. [Google Scholar]
- Bullerman, L.B.; Bianchini, A. Stability of mycotoxins during food processing. Int. J. Food Micro. 2007, 119, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Hueza, I.; Raspantini, P.; Raspantini, L.; Latorre, A.; Górniak, S. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massart, F.; Meucci, V.; Saggese, G.; Soldani, G. High growth rate of girls with precocious puberty exposed to estrogenic mycotoxins. J. Pediatr. 2008, 152, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Jakimiuk, E.; Gajecka, M.; Jana, B.; Brzuzan, P.; Zielonka, L.; Skorska-Wyszynska, E.; Gajecki, M. Factors determining sensitivity of prepubertal gilts to hormonal influence of zearalenone. Pol. J. Vet. Sci. 2009, 12, 149–158. [Google Scholar] [PubMed]
- Liu, T.; Xu, X.; Leng, W.; Xue, Y.; Dong, J.; Jin, Q. Analysis of gene expression changes in Trichophyton rubrum after skin interaction. J. Med. Microbiol. 2014, 63 Pt 5, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Perczak, A.; Juś, K.; Marchwińska, K.; Gwiazdowska, D.; Waśkiewicz, A.; Goliński, P. Degradation of zearalenone by essential oils under in vitro conditions. Front. Microbiol. 2016, 7, 1224. [Google Scholar] [CrossRef] [Green Version]
- Abid-Essefi, S.; Ouanes, Z.; Hassen, W.; Baudrimont, I.; Creppy, E.; Bacha, H. Cytotoxicity, inhibition of DNA and protein syntheses and oxidative damage in cultured cells exposed to zearalenone. Toxicol. in Vitro 2004, 18, 467–474. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Asi, M.R.; Jinap, S.; Rashid, U. Detection of aflatoxins and zearalenone contamination in wheat derived products. Food Control 2014, 35, 223–226. [Google Scholar] [CrossRef]
- Van Bennekom, E.O.; Brouwer, L.; Laurant, E.H.M.; Hooijerink, H.; Nielen, M.W.F. Confirmatory analysis method for zeranol, its metabolites and related mycotoxins in urine by liquid chromatography-negative ion electrospray tandem mass spectrometry. Anal. Chim. Acta 2002, 473, 151–160. [Google Scholar] [CrossRef]
- Tiemann, U.; Tomek, W.; Schneider, F.; Vanselow, J. Effects of the mycotoxins α-and β-zearalenol on regulation of progesterone synthesis in cultured granulosa cells from porcine ovaries. Reprod. Toxicol. 2003, 17, 673–681. [Google Scholar] [CrossRef]
- De Saeger, S.; Sibanda, L.; Van Peteghem, C. Analysis of zearalenone and α-zearalenol in animal feed using high-performance liquid chromatography. Anal. Chim. Acta 2003, 487, 137–143. [Google Scholar] [CrossRef]
- El-Nezami, H.; Polychronaki, N.; Salminen, S.; Mykkänen, H. Binding rather than metabolism may explain the interaction of two food-grade Lactobacillus strains with Zearalenone and Its Derivative ά-Zearalenol. Appl. Environ. Microbiol. 2002, 68, 3545–3549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabak, B.; Dobson, A.D.; Var, I.I.L. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci.Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Bata, Á.; Lásztity, R. Detoxification of mycotoxin-contaminated food and feed by microorganisms. Trends Food Sci. Technol. 1999, 10, 223–228. [Google Scholar] [CrossRef]
- Takahashi-Ando, N.; Ohsato, S.; Shibata, T.; Hamamoto, H.; Yamaguchi, I.; Kimura, M. Metabolism of zearalenone by genetically modified organisms expressing the detoxification gene from Clonostachys rosea. Appl. Environ. Microbiol. 2004, 70, 3239–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosawang, C.; Karlsson, M.; Vélëz, H.; Rasmussen, P.H.; Collinge, D.B.; Jensen, B.; Jensen, D.F. Zearalenone detoxification by zearalenone hydrolase is important for the antagonistic ability of Clonostachys rosea against mycotoxigenic Fusarium graminearum. Fung. Biol. 2014, 118, 364–373. [Google Scholar] [CrossRef]
- Król, A.; Pomastowski, P.; Rafińska, K.; Railean-Plugaru, V.; Walczak, J.; Buszewski, B. Microbiology neutralization of zearalenone using Lactococcus lactis and Bifidobacterium sp. Anal. Bioanal. Chem. 2018, 410, 943–952. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Walczak, J.; Railean-Plugaru, V.; Rudnicka, J.; Buszewski, B. Investigation of Zearalenone adsorption and biotransformation by microorganisms cultured under cellular stress conditions. Toxins 2019, 11, 463. [Google Scholar] [CrossRef] [Green Version]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of Fumonisin B1 and Zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef]
- Tinyiro, S.E.; Wokadala, C.; Xu, D.; Yao, W. Adsorption snd degradation of zearalenone by Bacillus strains. Folia Microbiol. 2011, 56, 321–327. [Google Scholar] [CrossRef]
- Lei, Y.P.; Zhao, L.H.; Ma, Q.G.; Zhang, J.Y.; Zhou, T.; Gao, C.Q.; Ji, C. Degradation of zearalenone in swine feed and feed ingredients by Bacillus subtilis ANSB01G. World Mycotoxin J. 2014, 7, 143–151. [Google Scholar] [CrossRef]
- Wang, J.Q.; Yang, F.; Yang, P.L.; Liu, J.; Lv, Z.H. Microbial reduction of zearalenone by a new isolated Lysinibacillus sp. ZJ-2016-1. World Mycotoxin J. 2018, 11, 571–578. [Google Scholar] [CrossRef]
- Altalhi, A.D.; El-Deeb, B. Localization of zearalenone detoxification gene(s) in pZEA-1 plasmid of Pseudomonas putida ZEA-1 and expressed in Escherichia coli. J. Hazard. Mater. 2009, 161, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Złoch, M.; Pomastowski, P.; Peer, M.; Sparbier, K.; Kostrzewa, M.; Buszewski, B. Study on carbapenemase-producing bacteria by matrix-assisted laser desorption/ionization approach. PLoS ONE 2021, 16, e0247369. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.S.; Yu, D.; Liu, N.; Wu, A. Degrading Ochratoxin A and Zearalenone Mycotoxins using a multifunctional recombinant enzyme. Toxins 2019, 11, 301. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.J.; Kang, J.S.; Cho, W.T.; Lee, C.H.; Ha, J.K.; Song, K.B. In vitro degradation of Zearalenone by Bacillus subtilis. Biotechnol. Lett. 2010, 32, 1921–1924. [Google Scholar] [CrossRef] [PubMed]
- Ser, H.L.; Palanisamy, U.D.; Yin, W.F.; Abd Malek, S.N.; Chan, K.G.; Goh, B.H.; Lee, L.H. Presence of antioxidative agent, Pyrrolo[ 1,2-a]pyrazine-1,4-dione, hexahydro-in newly isolated Streptomyces mangrovisoli sp.nov. Front. Microbiol. 2015, 6, 854. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Vujanovic, V. Biodegradation and biodetoxification of Fusarium mycotoxins by Sphaerodes mycoparasitica. AMB Express 2017, 7, 145. [Google Scholar] [CrossRef]
- Vanhoutte, I.; Audenaert, K.; De Gelder, L. Biodegradation of Mycotoxins: Tales from known and unexplored worlds. Front. Microbiol. 2016, 7, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafay, S.; Gil-Izquierdo, A. Bioavailability of phenolic acids. Phytochem. Rev. 2008, 7, 301–311. [Google Scholar] [CrossRef]
- Aneja, M.K.; Sharma, S.; Fleischmann, F.; Stich, S.; Heller, W.; Bahnweg, G.; Munch, J.C.; Schloter, M. Microbial colonization of beech and spruce litter-influence of decomposition site and plant litter species on the diversity of microbial community. Microb. Ecol. 2006, 52, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Fawole, M.O.; Oso, B.A. Characterization of Bacteria: Laboratory Manual of Microbiology; Spectrum Book Ltd.: Ibadan, Nigeria, 2004; pp. 24–33. [Google Scholar]
- Cheesbrough, M. District Laboratory Practice in Tropical Countries; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Mirocha, C.J.; Kolaczkowski, E.; Xie, W.; Yu, H.; Jelen, H. Analysis of deoxynivalenol and its derivatives (Batch and Single Kernel) Using Gas Chromatography/Mass Spectrometry. J. Agric. Food Chem. 1998, 46, 1414–1418. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Strain Code | Catalase Test | Oxidase Test | Amylase Test | Indole Test | Carbohydrate Fermentation Test | Gram Staining |
|---|---|---|---|---|---|---|---|
| 1 | FSL-1 | – | – | + | – | + | Gram positive rods |
| 2 | FSL-2 | + | + | + | – | + | Gram negative rods |
| 3 | FSL-5 | + | + | – | – | + | Gram Positive short rods |
| 4 | FSL-8 | – | – | – | – | + | Gram positve rods |
| 5 | FSL-12 | – | – | + | – | + | Gram positive rods |
| 6 | FSL-13 | + | + | – | – | – | Gram positive short rods |
| 7 | FSL-17 | + | + | + | – | + | Gram negative rods |
| 8 | FSL-18 | – | – | + | – | + | Gram positive rods |
| 9 | FSL-20 | + | + | + | – | + | Gram negative rods |
| 10 | FSL-21 | + | + | – | – | – | Gram positive short rods |
| 11 | FSL-24 | – | – | + | – | + | Gram positive rods |
| 12 | FSL-26 | – | – | + | – | + | Gram negative rods |
| 13 | FSL-27 | – | – | + | – | + | Gram positive rods |
| 14 | RS-5 | + | + | + | – | + | Gram positive rods |
| 15 | RS-6 | + | + | – | – | – | Gram Positive short rods |
| 16 | RS-13 | + | + | – | – | + | Gram positive rods |
| 17 | RS-16 | + | + | + | – | + | Gram positive rods |
| S.No | Strain Code | Absorbance Percentage (Y-Axis) | Log Concentration (X-Axis) | Initial Concentration (ng mL−1) | Final Concentration (ng mL−1) | Degraded Amount (ng mL−1) |
|---|---|---|---|---|---|---|
| 1 | FSL-1 | 75.7 | 1.87 | 25 | 6.07 | 18.93 |
| 2 | FSL-2 | 72.8 | 1.86 | 25 | 5.55 | 19.45 |
| 3 | FSL-5 | 70.6 | 1.848 | 25 | 7.35 | 17.65 |
| 4 | FSL-8 | 78 | 1.89 | 25 | 5.5 | 19.50 |
| 5 | FSL-12 | 73.1 | 1.863 | 25 | 6.72 | 18.28 |
| 6 | FSL-13 | 66.8 | 1.81 | 25 | 8.9 | 16.10 |
| 7 | FSL-17 | 72.6 | 1.86 | 25 | 6.85 | 18.15 |
| 8 | FSL-20 | 67.5 | 1.829 | 25 | 8.1 | 19.50 |
| 9 | FSL-21 | 69.5 | 1.841 | 25 | 7.62 | 17.38 |
| 10 | FSL-24 | 71.6 | 1.854 | 25 | 7.22 | 17.78 |
| 11 | FSL-26 | 69 | 1.838 | 25 | 7.75 | 17.25 |
| 12 | FSL-27 | 72 | 1.857 | 25 | 7.0 | 18.00 |
| 13 | FSL-30 | 68.5 | 1.835 | 25 | 7.8 | 17.20 |
| 14 | RS-5 | 78.3 | 1.89 | 25 | 2.9 | 22.10 |
| 15 | RS-6 | 65.9 | 1.818 | 25 | 8.75 | 16.25 |
| 16 | RS-13 | 64.4 | 1.808 | 25 | 8.9 | 16.10 |
| 17 | RS-16 | 67 | 1.826 | 25 | 8.25 | 16.75 |
| 18 | Control | 0 | 1.39 | 25 | 25 | 0 |
| S.No | Strain Code | Degradation % 24 h | Degradation % 48 h | Degradation % 72 h | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MRS Broth | Trypticase in Soy Broth | Nutrient Broth | MRS Broth | Trypticase in Soy Broth | Nutrient Broth | MRS Broth | Trypticase in Soy Broth | Nutrient Broth | ||
| 1 | FSL-1 | 39.3 ± 0.04 | 35.3 ± 0.01 | 32.1 ± 0.04 | 61.7 ± 0.02 | 58.8 ± 0.02 | 57.3 ± 0.02 | 75.7 ± 0.01 | 72.8 ± 0.02 | 74.9 ± 0.04 |
| 2 | FSL-2 | 39.0 ± 0.01 | 32.8 ± 0.02 | 32.5 ± 0.02 | 61.0 ± 0.01 | 58.3 ± 0.04 | 58.0 ± 0.01 | 72.8 ± 0.01 | 70.9 ± 0.03 | 70.8 ± 0.02 |
| 3 | FSL-5 | 36.5 ± 0.03 | 31.8 ± 0.03 | 31.0 ± 0.04 | 57.0 ± 0.03 | 53.0 ± 0.04 | 51.8 ± 0.03 | 70.6 ± 0.04 | 66.5 ± 0.01 | 67.0 ± 0.03 |
| 4 | FSL-8 | 37.1 ± 0.02 | 33.8 ± 0.02 | 28.6 ± 0.01 | 62.0 ± 0.04 | 58.4 ± 0.02 | 56.2 ± 0.04 | 78.0 ± 0.02 | 72.5 ± 0.01 | 70.6 ± 0.01 |
| 5 | FSL-17 | 38.6 ± 0.03 | 29.8 ± 0.02 | 27.9 ± 0.01 | 60.3 ± 0.02 | 56.3 ± 0.01 | 55.8 ± 0.03 | 72.6 ± 0.04 | 69.6 ± 0.04 | 69.0 ± 0.01 |
| 6 | FSL-20 | 32.3 ± 0.02 | 30.8 ± 0.01 | 28.6 ± 0.01 | 51.2 ± 0.01 | 46.4 ± 0.01 | 48.2 ± 0.02 | 67.5 ± 0.03 | 61.8 ± 0.02 | 65.0 ± 0.03 |
| 7 | FSL-21 | 31.1 ± 0.04 | 27.3 ± 0.03 | 26.7 ± 0.02 | 56.9 ± 0.04 | 52.1 ± 0.04 | 51.0 ± 0.01 | 69.5 ± 0.03 | 65.7 ± 0.03 | 65.5 ± 0.03 |
| 8 | FSL-24 | 36.8 ± 0.03 | 28.5 ± 0.04 | 29.0 ± 0.04 | 60.7 ± 0.02 | 54.0 ± 0.03 | 53.3 ± 0.04 | 71.6 ± 0.01 | 66.7 ± 0.01 | 66.8 ± 0.01 |
| 9 | FSL-27 | 31.7 ± 0.01 | 27.0 ± 0.01 | 26.4 ± 0.04 | 53.2 ± 0.03 | 48.0 ± 0.04 | 46.6 ± 0.03 | 72.0 ± 0.01 | 67.8 ± 0.01 | 67.3 ± 0.02 |
| 10 | RS-5 | 34.3 ± 0.01 | 38.6 ± 0.02 | 33.6 ± 0.01 | 65.6 ± 0.04 | 59.3 ± 0.01 | 59.0 ± 0.03 | 78.3 ± 0.01 | 73.8 ± 0.04 | 72.8 ± 0.01 |
| Strain Codes | ELISA % | HPLC % |
|---|---|---|
| FSL-1 | 75.7 | 71.2 |
| FSL-2 | 72.8 | 72.3 |
| FSL-5 | 70.6 | 69.7 |
| FSL-8 | 78.0 | 72.4 |
| FSL-12 | 73.1 | 70.3 |
| FSL-13 | 66.8 | 65.3 |
| FSL-17 | 72.6 | 70.2 |
| FSL-20 | 67.5 | 69.0 |
| FSL-21 | 69.5 | 69.2 |
| FSL-24 | 71.6 | 69.8 |
| FSL-26 | 69.0 | 68.5 |
| FSL-27 | 72.0 | 70.0 |
| FSL-30 | 68.5 | 68.0 |
| RS-5 | 78.3 | 74.3 |
| RS-6 | 65.9 | 64.2 |
| RS-13 | 64.4 | 64.0 |
| RS-17 | 67.0 | 66.4 |
| Average | 70.77 | 69.11 |
| Standard Deviation (±) | 3.935 | 2.768 |
| p value | p < 0.05 | |
| S.No | Compound | Retention Time (RT) | Area % | Probability |
|---|---|---|---|---|
| 1 | 1 propene,3,3ꞌ-oxy bis | 0.43 | 40.56 | 21.01 |
| 2 | 4,8 Dioxaspiro[2.5] oct-1-ene 6,6 dimethyl | 0.43 | 40.56 | 7.04 |
| 3 | 3-pentyn-1-ol | 0.43 | 40.56 | 5.25 |
| 4 | 3,5,9-trioxa-5-phosphahepatacos-8-en-1-aminium | 1.69 | 0.46 | 18.53 |
| 5 | pyrrolo[1.2-q] pyrazine-1,4-dione,hexahydro | 2.69 | 6.17 | 56.34 |
| 6 | Glycyl-L-proline | 2.69 | 6.17 | 30.75 |
| 7 | Pyrrolo[1,2-a] pyrazine-1,4 dione, hexahydro-3-(2-methyl propyl)- | 3.26 | 2.85 | 40.66 |
| 8 | 9-octadecenoic acid, (2-phenyl-1,3-dioxolan-4-y) methyl ester, cis | 3.81 | 0.48 | 36.79 |
| 9 | Hexasiloxane,1,1,3,3,5,5,7,7,9,9,11,11 dodecamethyl | 5.61 | 8.21 | 35.62 |
| 10 | 1-monolinoleoylglycerol trymethylsilyl ether | 7.47 | 0.12 | 53.03 |
| 11 | 1-monolinoleoylglycerol trymethylsilyl ether | 8.05 | 0.25 | 54.44 |
| 12 | Octasiloxane,1,1,3,3,5,5,7,7,9,9,11,11,13,13,15,15-hexadecamethyl- | 9.57 | 40.27 | 41.45 |
| 13 | Octasiloxane,1,1,3,3,5,5,7,7,9,9,11,11,13,13,15,15-hexadecamethyl- | 10.51 | 0.26 | 40.70 |
| 14 | Octasiloxane,1,1,3,3,5,5,7,7,9,9,11,11,13,13,15,15-hexadecamethyl- | 10.94 | 0.24 | 50.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zada, S.; Alam, S.; Ayoubi, S.A.; Shakeela, Q.; Nisa, S.; Niaz, Z.; Khan, I.; Ahmed, W.; Bibi, Y.; Ahmed, S.; et al. Biological Transformation of Zearalenone by Some Bacterial Isolates Associated with Ruminant and Food Samples. Toxins 2021, 13, 712. https://doi.org/10.3390/toxins13100712
Zada S, Alam S, Ayoubi SA, Shakeela Q, Nisa S, Niaz Z, Khan I, Ahmed W, Bibi Y, Ahmed S, et al. Biological Transformation of Zearalenone by Some Bacterial Isolates Associated with Ruminant and Food Samples. Toxins. 2021; 13(10):712. https://doi.org/10.3390/toxins13100712
Chicago/Turabian StyleZada, Sharif, Sadia Alam, Samha Al Ayoubi, Qismat Shakeela, Sobia Nisa, Zeeshan Niaz, Ibrar Khan, Waqas Ahmed, Yamin Bibi, Shehzad Ahmed, and et al. 2021. "Biological Transformation of Zearalenone by Some Bacterial Isolates Associated with Ruminant and Food Samples" Toxins 13, no. 10: 712. https://doi.org/10.3390/toxins13100712
APA StyleZada, S., Alam, S., Ayoubi, S. A., Shakeela, Q., Nisa, S., Niaz, Z., Khan, I., Ahmed, W., Bibi, Y., Ahmed, S., & Qayyum, A. (2021). Biological Transformation of Zearalenone by Some Bacterial Isolates Associated with Ruminant and Food Samples. Toxins, 13(10), 712. https://doi.org/10.3390/toxins13100712