Ciguatoxin-Producing Dinoflagellate Gambierdiscus in the Beibu Gulf: First Report of Toxic Gambierdiscus in Chinese Waters

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
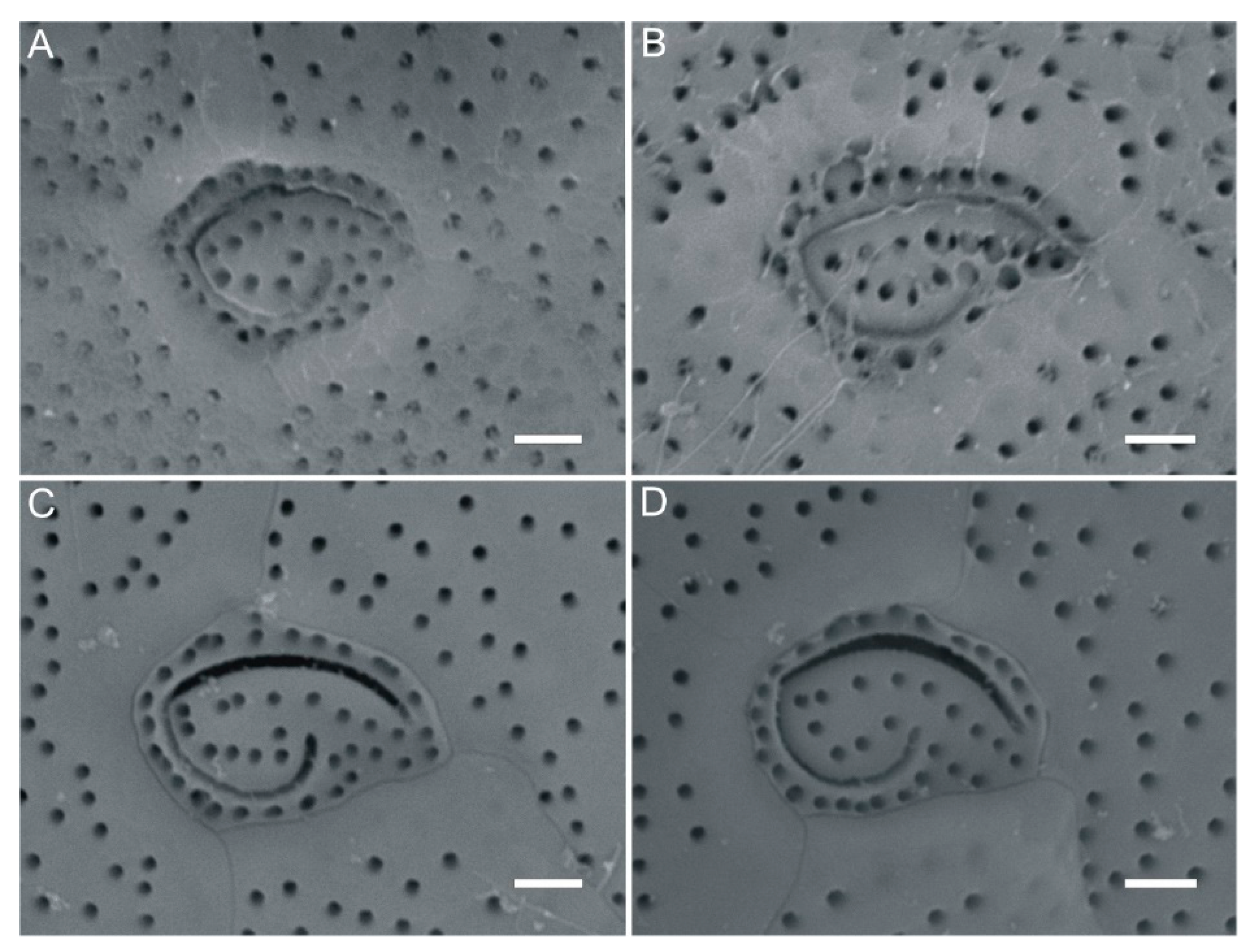
2.1. Morphological Analysis
2.2. Sequence Analysis
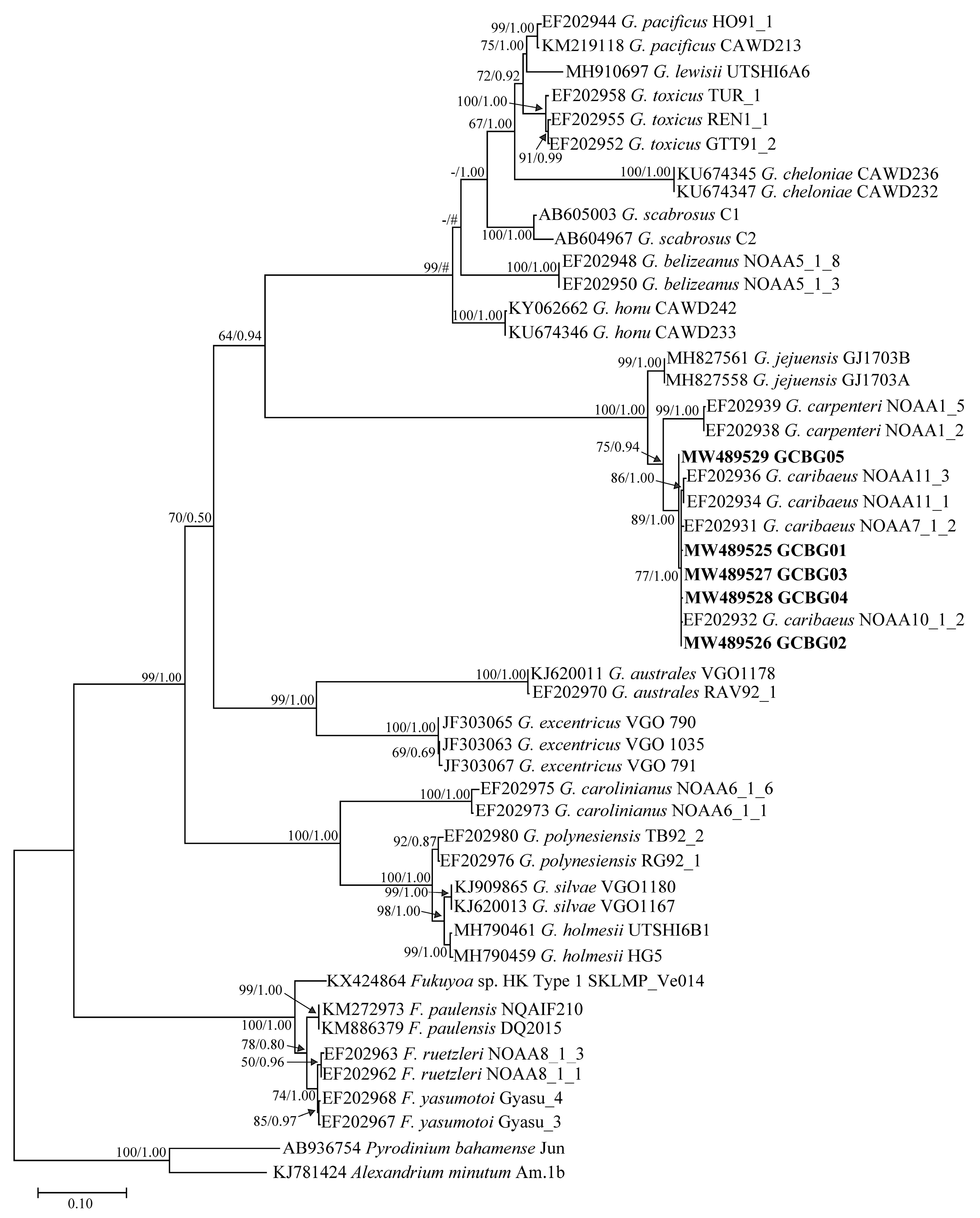
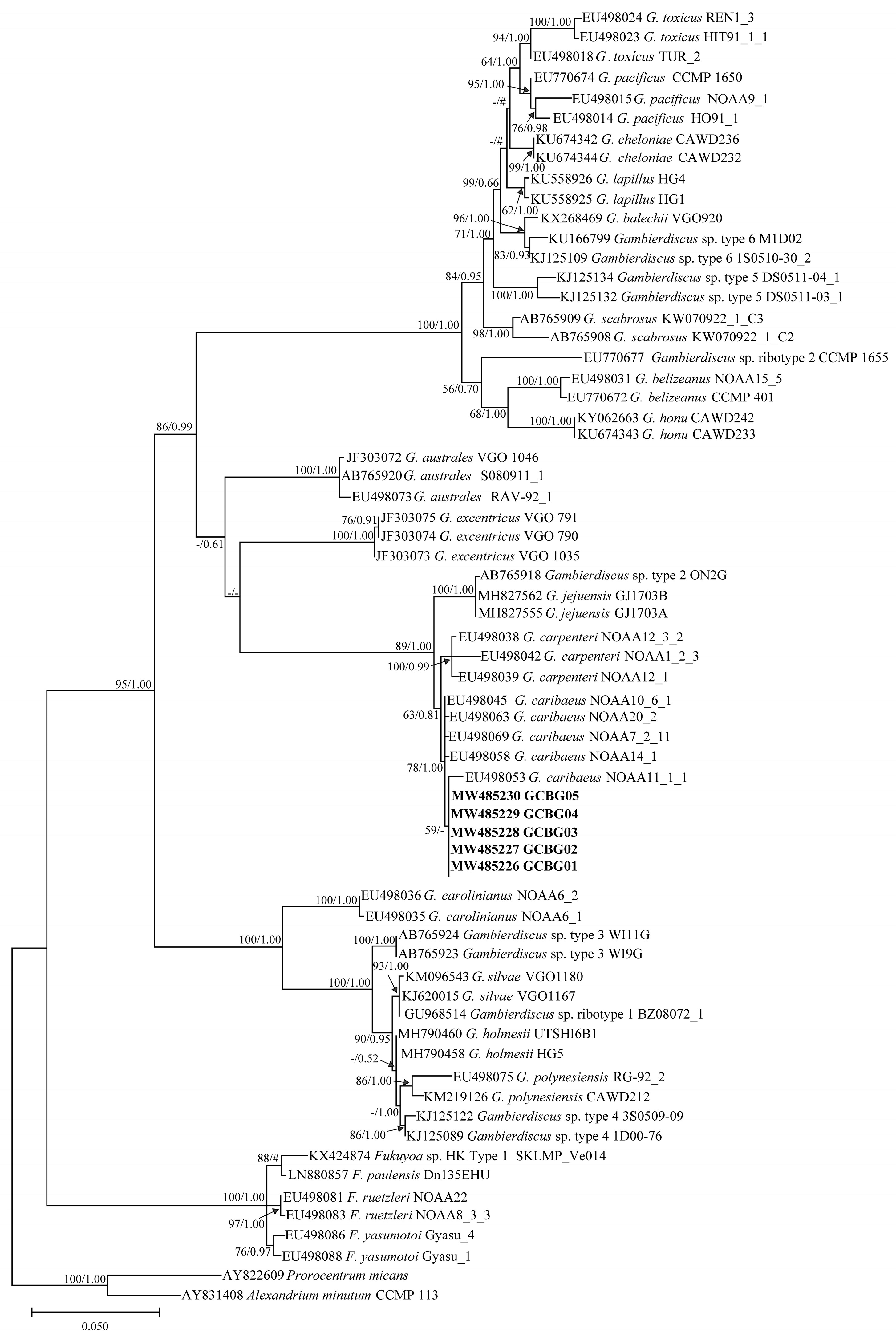
2.3. Phylogenetic Tree Analysis
2.4. Toxin Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
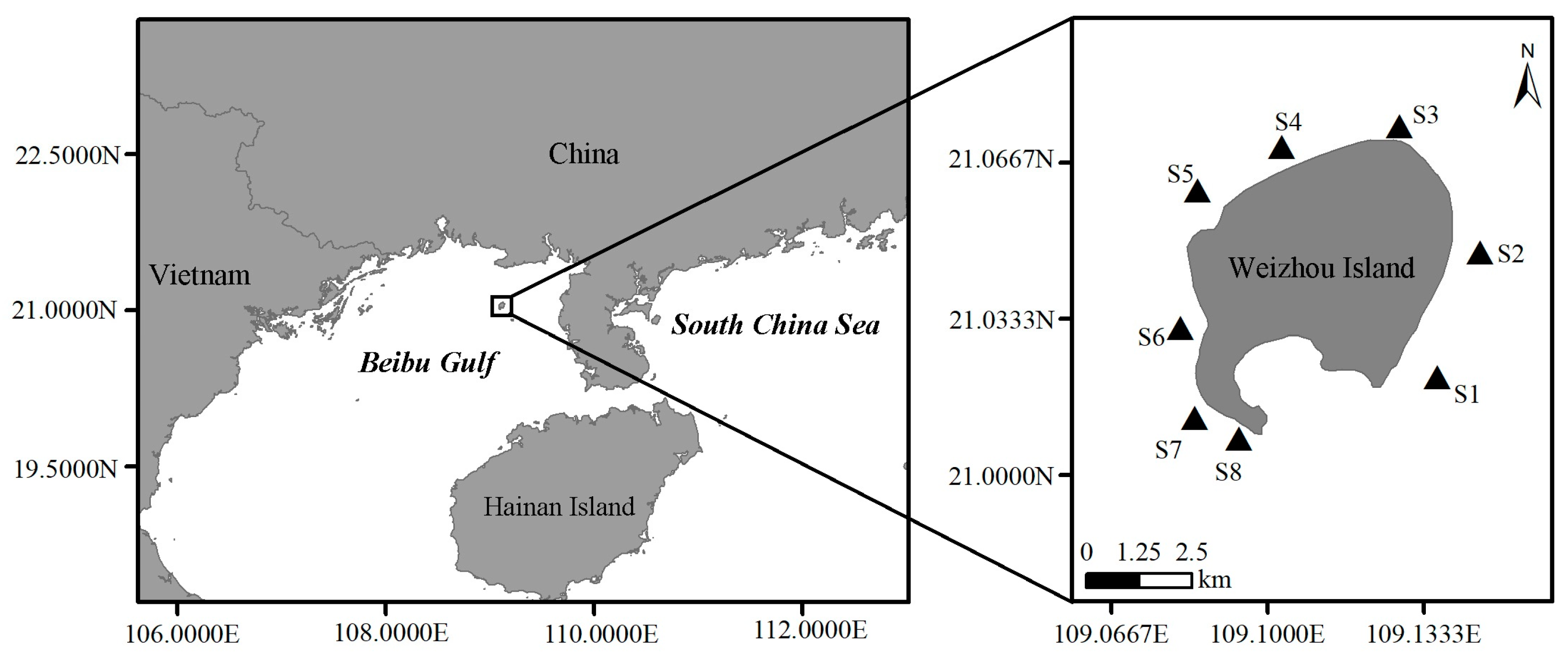
5.1. Study Area and Sample Collection
5.2. Culture Establishment
5.3. Scanning Electron Microscopy
5.4. DNA Extraction and PCR Amplification
5.5. Phylogeny and Sequence Analysis
5.6. CTX-Like Toxicity Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinain, M.; Gatti, C.M.I.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef] [PubMed]
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Chan, T.Y.K. Characteristic features and contributory factors in fatal Ciguatera Fish Poisoning—Implications for prevention and public education. Am. J. Trop. Med. Hyg. 2016, 94, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Randall, J.E. A review of ciguatera, tropical fish poisoning, with a tentative explanation of its cause. Bull. Mar. Sci. 1958, 8, 236–267. [Google Scholar]
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a dinoflagellate as a likely culprit of ciguatera. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Adachi, R.; Fukuyo, Y. The thecal structure of a marine toxic dinoflagellate Gambierdiscus toxicus gen. et sp. nov. collected in a ciguatera-endemic area. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Gómez, F.; Qiu, D.J.; Lopes, R.M.; Lin, S.J. Fukuyoa paulensis gen. et sp nov., a new genus for the globular species of the dinoflagellate Gambierdiscus (Dinophyceae). PLoS ONE 2015, 10, e0119676. [Google Scholar]
- Parsons, M.L.; Aligizaki, K.; Bottein, M.Y.D.; Fraga, S.; Morton, S.L.; Penna, A.; Rhodes, L. Gambierdiscus and Ostreopsis: Reassessment of the state of knowledge of their taxonomy, geography, ecophysiology, and toxicology. Harmful Algae 2012, 14, 107–129. [Google Scholar] [CrossRef]
- Xu, Y.; Richlen, M.L.; Liefer, J.D.; Robertson, A.; Kulis, D.; Smith, T.B.; Parsons, M.L.; Anderson, D.M. Influence of environmental variables on Gambierdiscus spp. (Dinophyceae) growth and distribution. PLoS ONE 2016, 11, e0153197. [Google Scholar] [CrossRef]
- Tester, P.A.; Litaker, R.W.; Berdalet, E. Climate change and harmful benthic microalgae. Harmful Algae 2020, 91, 101655. [Google Scholar] [CrossRef]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Vázquez, E.J.; Almazán-Becerril, A.; López-Cortés, D.J.; Heredia-Tapia, A.; Hernández-Sandoval, F.E.; Band-Schmidt, C.J.; Bustillos-Guzmán, J.J.; Gárate-Lizárraga, I.; García-Mendoza, E.; Salinas-Zavala, C.A.; et al. Ciguatera in Mexico (1984–2013). Mar. Drugs 2019, 17, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdalet, E.; Bravo, I.; Evans, J.; Fraga, S.; Kibler, S.; Kudela, M.; Larsen, J.; Litaker, W.; Penna, A.; Tester, P.; et al. Global Ecology and Oceanography of Harmful Algal Blooms, GEOHAB Core Research Project: HABs in Benthic Systems; Intergovernmental Oceanographic Commission: Lausanne, Switzerland, 2012. [Google Scholar]
- Berdalet, E.; Tester, P.A. Key Questions and Recent Research Advances on Harmful Algal Blooms in Benthic Systems. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Springer: Cham, Switzerland, 2018; Volume 232, pp. 261–286. [Google Scholar]
- Hua, Z. Overview of studies on ciguatoxin. Mar. Environ. Sci. 1994, 13, 57–63. [Google Scholar]
- Chan, T.Y.K. Emergence and epidemiology of ciguatera in the coastal cities of southern China. Mar. Drugs 2015, 13, 1175–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, T.Y.K. Ciguatera fish poisoning in east Asia and southeast Asia. Mar. Drugs 2015, 13, 3466–3478. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Jin, X. Toxic fish in China. Chin. J. Zool. 1977, 38–40. [Google Scholar]
- Chan, T.Y.K.; Wang, A.Y.M. Life-threatening bradycardia and hypotension in a patient with ciguatera fish poisoning. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 71. [Google Scholar] [CrossRef]
- Zhong, W.; Wu, G. Clinical manifestations of cardiovascular system in patients with ciguatoxin poisoning. J. New Med. 2006, 37, 316–317. [Google Scholar]
- Xu, Y.; Wang, A.; Hu, R.; Jiang, T.; Jiang, T. Ciguatera toxins in coral fishes along the southern coast of China. China Environ. Sci. 2012, 32, 330–336. [Google Scholar]
- Chen, G. Studies on the dinoflagellata in adjacent waters of the Xisha Islands III. some rare tropical ocean species. Oceanol. Limnol. Sin. 1989, 20, 230–237. [Google Scholar]
- Lu, S.H.; Hodgkiss, I.J. An unusual year for the occurrence of harmful algae. Harmful Algal News 1999, 18, 1–3. [Google Scholar]
- Lu, S.H.; Hodgkiss, I.J. Harmful algal bloom causative collected from Hong Kong waters. Hydrobiologia 2004, 512, 231–238. [Google Scholar] [CrossRef]
- Lei, A.; Chen, H.; Chen, J.; Hu, Z. Research on species composition and biomass of Phytoplankton in Daya Bay artificial reef area. J. Ocean Technol. 2009, 28, 83–88. [Google Scholar]
- Zhang, H.; Wu, Z.; Cen, J.; Li, Y.; Wang, H.; Lu, S. First report of three benthic dinoflagellates, Gambierdiscus pacificus, G. australes and G. caribaeus (Dinophyceae), from Hainan Island, South China Sea. Phycol. Res. 2016, 64, 259–273. [Google Scholar] [CrossRef]
- Zhao, W.; Wei, H.; Guo, K. The community structure and the seasonal changes of phytoplankton in Dayao Bay in Dalian Coast from 2006 to 2007. J. Dalian Ocean Univ. 2011, 26, 291–298. [Google Scholar]
- Wu, H.; Luan, Q.; Guo, M.; Gu, H.; Zhai, Y.; Tan, Z. Phycotoxins in scallops (Patinopecten yessoensis) in relation to source, composition and temporal variation of phytoplankton and cysts in North Yellow Sea, China. Mar. Pollut. Bull. 2018, 135, 1198–1204. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, Y.; Kang, W.; Liang, J.; Gu, Y.; Jiang, X. Germination of phytoplankton resting cells from surface sediments in two areas of the Southern Chinese coastal waters. Mar. Ecol. 2013, 34, 218–232. [Google Scholar] [CrossRef]
- Leung, P.T.Y.; Yan, M.; Lam, V.; Yiu, S.K.F.; Chen, C.Y.; Murray, J.S.; Harwood, D.T.; Rhodes, L.L.; Lam, P.K.S.; Wai, T.C. Phylogeny, morphology and toxicity of benthic dinoflagellates of the genus Fukuyoa (Goniodomataceae, Dinophyceae) from a subtropical reef ecosystem in the South China Sea. Harmful Algae 2018, 74, 78–97. [Google Scholar] [CrossRef]
- Ho, T.V.; Bing, H.Y.N. First report of Gambierdiscus caribaeus and G. carpenteri (Dinophyceae) from Nha Trang Bay, South Central Vietnam. J. Mar. Biol. Ass. India 2018, 60, 5–11. [Google Scholar] [CrossRef]
- Wong, C.K.; Hung, P.; Lo, J.Y. Ciguatera fish poisoning in Hong Kong-A 10-year perspective on the class of ciguatoxins. Toxicon 2014, 86, 96–106. [Google Scholar] [CrossRef]
- Lv, S.; Li, Y. Research on the prevalence of ciguatera fish poisoning in China. China J. Public Health 2006, 22, 226–227. [Google Scholar]
- Wang, X.; Li, G. The status and prospect of researches on coral reef in Weizhou Island. J. Guangxi Acad. Sci. 2009, 25, 72–75. [Google Scholar]
- Wang, W.; Yu, K.; Wang, Y. A review on the research of coral reefs in the Weizhou Island, Beibu Gulf. Trop. Geogr. 2016, 36, 72–79. [Google Scholar]
- Zhou, H.; Wang, X.; Liang, W. Reflections on the characteristics, evolution and protection and rehabilitation strategies of the coral reefs at Weizhou Island. J. Guangxi Acad. Sci. 2020, 36, 228–236. [Google Scholar]
- Wang, Y. Assessment of Coral Reef Ecosystem Health and Ecological Assets in Past Three Decades at Weizhou Island Reef, Beibu Gulf. Master’s Thesis, Guangxi University, Guangxi, China, 2020. [Google Scholar]
- Mustapa, N.I.; Yong, H.L.; Lee, L.K.; Lim, Z.F.; Lim, H.C.; Teng, S.T.; Luo, Z.; Gu, H.; Leaw, C.P.; Lim, P.T. Growth and epiphytic behavior of three Gambierdiscus species (Dinophyceae) associated with various macroalgal substrates. Harmful Algae 2019, 89, 101671. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Chinain, M.; Holmes, M.J.; Holland, W.C.; Tester, P.A. Taxonomy of Gambierdiscus including four new species, Gambierdiscus caribaeus, Gambierdiscus carolinianus, Gambierdiscus carpenteri and Gambierdiscus ruetzleri (Gonyaulacales, Dinophyceae). Phycologia 2009, 48, 344–390. [Google Scholar] [CrossRef]
- Kretzschmar, A.L.; Larsson, M.E.; Hoppenrath, M.; Doblin, M.A.; Murray, S.A. Characterisation of two toxic Gambierdiscus spp. (Gonyaulacales, Dinophyceae) from the great barrier reef (Australia): G. lewisii sp. nov. and G. holmesii sp. nov. Protist 2019, 170, 125699. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.H.; Jeong, H.J.; Du, Y.Y. Gambierdiscus jejuensis sp. nov., an epiphytic dinoflagellate from the waters of Jeju Island, Korea, effect of temperature on the growth, and its global distribution. Harmful Algae 2018, 80, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Vacarizas, J.; Benico, G.; Austero, N.; Azanza, R. Taxonomy and toxin production of Gambierdiscus carpenteri (Dinophyceae) in a tropical marine ecosystem: The first record from the Philippines. Mar. Pollut. Bull. 2018, 137, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Tawong, W.; Nishimura, T.; Sakanari, H.; Sato, S.; Yamaguchi, H.; Adachi, M. Characterization of Gambierdiscus and Coolia (Dinophyceae) isolates from Thailand based on morphology and phylogeny. Phycol. Res. 2015, 63, 125–133. [Google Scholar] [CrossRef]
- Tudó, À.; Gaiani, G.; Rey Varela, M.; Tsumuraya, T.; Andree, K.B.; Fernández-Tejedor, M.; Campàs, M.; Diogène, J. Further advance of Gambierdiscus species in the Canary Islands, with the first report of Gambierdiscus belizeanus. Toxins 2020, 12, 692. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef] [PubMed]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, e0185776. [Google Scholar] [CrossRef] [PubMed]
- Rossignoli, A.E.; Tudó, A.; Bravo, I.; Díaz, P.A.; Diogène, J.; Riobó, P. Toxicity characterisation of Gambierdiscus species from the Canary Islands. Toxins 2020, 12, 134. [Google Scholar] [CrossRef] [Green Version]
- Longo, S.; Sibat, M.; Viallon, J.; Darius, H.T.; Hess, P.; Chinain, M. Intraspecific variability in the toxin production and toxin profiles of In vitro cultures of Gambierdiscus polynesiensis (Dinophyceae) from French Polynesia. Toxins 2019, 11, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roeder, K.; Erler, K.; Kibler, S.; Tester, P.; Nguyen-Ngoc, L.; Gerdts, G.; Luckas, B. Characteristic profiles of ciguatera toxins in different strains of Gambierdiscus spp. Toxicon 2010, 56, 731–738. [Google Scholar] [CrossRef]
- Tawong, W.; Yoshimatsu, T.; Yamaguchi, H.; Adachi, M. Temperature and salinity effects and toxicity of Gambierdiscus caribaeus (Dinophyceae) from Thailand. Phycologia 2016, 55, 274–278. [Google Scholar] [CrossRef]
- Lewis, R.J.; Inserra, M.; Vetter, I.; Holland, W.C.; Hardison, D.R.; Tester, P.A.; Litaker, R.W. Rapid extraction and identification of maitotoxin and ciguatoxin-like toxins from Caribbean and Pacific Gambierdiscus using a new functional bioassay. PLoS ONE 2016, 11, e0160006. [Google Scholar] [CrossRef]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Séchet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Asencio, L.; Clausing, R.J.; Vandersea, M.; Chamero-Lago, D.; Gómez-Batista, M.; Hernández-Albernas, J.I.; Chomérat, N.; Rojas-Abrahantes, G.; Litaker, R.W.; Tester, P.; et al. Ciguatoxin occurrence in food-web components of a Cuban coral reef ecosystem: Risk-assessment implications. Toxins 2019, 11, 722. [Google Scholar] [CrossRef] [Green Version]
- Gaiani, G.; Leonardo, S.; Tudó, À.; Toldrà, A.; Rey, M.; Andree, K.B.; Tsumuraya, T.; Hirama, M.; Diogène, J.; O’Sullivan, C.K.; et al. Rapid detection of ciguatoxins in Gambierdiscus and Fukuyoa with immunosensing tools. Ecotoxicol. Environ. Saf. 2020, 204, 111004. [Google Scholar] [CrossRef] [PubMed]
- Lartigue, J.; Jester, E.L.E.; Dickey, R.W.; Villareal, T.A. Nitrogen source effects on the growth and toxicity of two strains of the ciguatera-causing dinoflagellate Gambierdiscus toxicus. Harmful Algae 2009, 8, 781–791. [Google Scholar] [CrossRef]
- Liefer, J.D.; Richlen, M.L.; Smith, T.B.; DeBose, J.L.; Xu, Y.; Anderson, D.M.; Robertson, A. Asynchrony of Gambierdiscus spp. Abundance and Toxicity in the U.S. Virgin Islands: Implications for Monitoring and Management of Ciguatera. Toxins 2021, 13, 413. [Google Scholar] [CrossRef] [PubMed]
- Morton, S.; Bomber, J.; Tindall, D.; Aikman, K. Response of Gambierdiscus toxicus to light: Cell physiology and toxicity. In Toxic Phytoplankton Blooms in the Sea; Smayda, T., Shimizu, Y., Eds.; Elsevier: New York, NY, USA, 1993; pp. 541–546. [Google Scholar]
- Chinain, M.; Germain, M.; Deparis, X.; Pauillac, S.; Legrand, A.M. Seasonal abundance and toxicity of the dinoflagellate Gambierdiscus spp. (Dinophyceae), the causative agent of ciguatera in Tahiti, French Polynesia. Mar. Biol. 1999, 135, 259–267. [Google Scholar] [CrossRef]
- Reverté, L.; Toldrà, A.; Andree, K.B.; Fraga, S.; De Falco, G.; Campàs, M.; Diogène, J. Assessment of cytotoxicity in ten strains of Gambierdiscus australes from Macaronesian Islands by neuro-2a cell-based assays. J. Appl. Phycol. 2018, 30, 2447–2461. [Google Scholar] [CrossRef]
- Kibler, S.R.; Litaker, R.W.; Holland, W.C.; Vandersea, M.W.; Tester, P.A. Growth of eight Gambierdiscus (Dinophyceae) species: Effects of temperature, salinity and irradiance. Harmful Algae 2012, 19, 1–14. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef]
- Morton, S.; Norris, D. The role of temperature, salinity, and light on the seasonality of Prorocentrum lima. In Toxic Marine Phytoplankton; Graneli, E., Ed.; Elsevier: New York, NY, USA, 1990; pp. 201–205. [Google Scholar]
- Chinain, M.; Faust, M.A.; Pauillac, S. Morphology and molecular analyses of three toxic species of Gambierdiscus (Dinophyceae): G. pacificus, sp. nov., G. australes, sp. nov., and G. polynesiensis, sp. nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Xu, Y.; Richlen, M.L.; Morton, S.L.; Mak, Y.L.; Chan, L.L.; Tekiau, A.; Anderson, D.M. Distribution, abundance and diversity of Gambierdiscus spp. from a ciguatera-endemic area in Marakei, Republic of Kiribati. Harmful Algae 2014, 34, 56–68. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Analysis Length | Average Content (%) | Conserved Site | Variable Site | Parsimonious Information Site | Monomorphic Site | Conversion/Transversion Ratio | |||
|---|---|---|---|---|---|---|---|---|---|---|
| A | T | G | C | |||||||
| D1–D3 LSU rDNA | 1035 | 25.3 | 25.1 | 30.8 | 18.8 | 323 | 696 | 661 | 35 | 0.9 |
| D8–D10 LSU rDNA | 914 | 26.1 | 25.8 | 27.8 | 20.3 | 532 | 369 | 281 | 87 | 1.5 |
| SSU rDNA | 1774 | 24.9 | 27.1 | 28.1 | 19.9 | 1016 | 728 | 578 | 149 | 1.6 |
| Strains | Species | Toxicity |
|---|---|---|
| GCBG01 | G. caribaeus | 5.4 |
| GCBG02 | G. caribaeus | ND |
| GCBG03 | G. caribaeus | ND |
| GCBG04 | G. caribaeus | ND |
| GCBG05 | G. caribaeus | ND |
| Strain | Locality | Toxicity | Methodology | References |
|---|---|---|---|---|
| NOAA 7 (CCMP 1652) | Mataiva, Tahiti, South Pacific | 2,3-dihydroxy CTX3C * | LC-MS/MS | [49] |
| NOAA 20 (CCMP 1651) | Grand Cayman Island, Caribbean | 2,3-dihydroxy CTX3C * | LC-MS/MS | [49] |
| CCMP 401 | St. Barthelmy Island, Caribbean | 2,3-dihydroxy CTX3C * | LC-MS/MS | [49] |
| CCMP 1733 | Carry Bow Cay, Belize, Caribbean | 2,3-dihydroxy CTX3C * | LC-MS/MS | [49] |
| TF9G | Koh Wai, Trat, Thailand | ≥100 × 10−4 MU/1000 cells | MBA | [43,50] |
| Gam 19 | Carrie Bowe Caye Belize, Caribbean | <LOD | CBA | [51] |
| Pat HI Jar 2 Gam 2 | Big Island, Hawaii, USA, Pacific | <LOD | CBA | [51] |
| CCMP1733 | Carrie Bow Cay, Belize, Caribbean | 0.80 ± 0.43 fg CTX3C eq cell−1 | CBA | [46] |
| CCMP 1651 | Grand Cayman Island, Caribbean | 0.48 ± 0.04 fg CTX3C eq cell−1 | CBA | [46] |
| SW gam 5 | Southwater Cay, Belize, Caribbean | 1.52 ± 0.26 fg CTX3C eq cell−1 | CBA | [46] |
| CBC gam1 | Carrie Bow Cay, Belize, Caribbean | 0.62 ± 0.12 fg CTX3C eq cell−1 | CBA | [46] |
| Mexico Algae 1 | Cancun, Mexico | 1.29 ± 0.40 fg CTX3C eq cell−1 | CBA | [46] |
| Dive 1 FA | Carrie Bow Cay, Belize, Caribbean | 0.69 ± 0.19 fg CTX3C eq cell−1 | CBA | [46] |
| Keys Jar 7 | Florida Keys, USA | 0.19 ± 0.03 fg CTX3C eq cell−1 | CBA | [46] |
| Bill Hi Gam8 | Waikiki Beach, Honolulu, Hawaii | 1.60 ± 1.00 fg CTX3C eq cell−1 | CBA | [52] |
| CUB4A5 | Cienfuegos coast, south-central Cuba | <LOD | RBA | [53] |
| VGO1362 | La Gomera Porto-Playa Santiago, Canary Islands | 6.00 fg CTX3C eq cell−1 | CBA | [47] |
| VGO1364 | La Gomera Porto-Playa Santiago, Canary Islands | 25.9 ± 5.0 fg CTX3C eq cell−1 | CBA | [47] |
| VGO1365 | La Gomera Porto-Playa Santiago, Canary Islands | <LOD | CBA | [47] |
| VGO1366 | La Gomera Porto-Playa Santiago, Canary Islands | 5.00 fg CTX3C eq cell−1 | CBA | [47] |
| VGO1367 | La Gomera, San Sebastián-Playa la Cueva, Canary Islands | 903.7 ± 158.9 fg CTX3C eq cell−1 | CBA | [47] |
| IRTA-SMM-17-03 | Tamaduste, El Hierro, Canary Islands | <LOD | CBA | [44] |
| IRTA-SMM-17_03 | Tamaduste, El Hierro, Canary Islands | 0.13–0.21 fg 51-hydroxy CTX3C eq cell−1 | Immunoassay | [54] |
| IRTA-SMM-17_03 | Tamaduste, El Hierro, Canary Islands | 1.3–2.4 fg CTX3C eq cell−1 | Immunoassay | [54] |
| GCBG01 | Weizhou Island, Beibu Gulf, China | 5.40 fg CTX3C eq cell−1 | CBA | This study |
| GCBG02 | Weizhou Island, Beibu Gulf, China | <LOD | CBA | This study |
| GCBG03 | Weizhou Island, Beibu Gulf, China | <LOD | CBA | This study |
| GCBG04 | Weizhou Island, Beibu Gulf, China | <LOD | CBA | This study |
| GCBG05 | Weizhou Island, Beibu Gulf, China | <LOD | CBA | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; He, X.; Lee, W.H.; Chan, L.L.; Lu, D.; Wang, P.; Tao, X.; Li, H.; Yu, K. Ciguatoxin-Producing Dinoflagellate Gambierdiscus in the Beibu Gulf: First Report of Toxic Gambierdiscus in Chinese Waters. Toxins 2021, 13, 643. https://doi.org/10.3390/toxins13090643
Xu Y, He X, Lee WH, Chan LL, Lu D, Wang P, Tao X, Li H, Yu K. Ciguatoxin-Producing Dinoflagellate Gambierdiscus in the Beibu Gulf: First Report of Toxic Gambierdiscus in Chinese Waters. Toxins. 2021; 13(9):643. https://doi.org/10.3390/toxins13090643
Chicago/Turabian StyleXu, Yixiao, Xilin He, Wai Hin Lee, Leo Lai Chan, Douding Lu, Pengbin Wang, Xiaoping Tao, Huiling Li, and Kefu Yu. 2021. "Ciguatoxin-Producing Dinoflagellate Gambierdiscus in the Beibu Gulf: First Report of Toxic Gambierdiscus in Chinese Waters" Toxins 13, no. 9: 643. https://doi.org/10.3390/toxins13090643
APA StyleXu, Y., He, X., Lee, W. H., Chan, L. L., Lu, D., Wang, P., Tao, X., Li, H., & Yu, K. (2021). Ciguatoxin-Producing Dinoflagellate Gambierdiscus in the Beibu Gulf: First Report of Toxic Gambierdiscus in Chinese Waters. Toxins, 13(9), 643. https://doi.org/10.3390/toxins13090643