Naturally Occurring Fusarium Species and Mycotoxins in Oat Grains from Manitoba, Canada

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Oat Sample Collections
4.2. Isolation and Identification of Fusarium Pathogens
4.3. Extraction and Quantification of Fusarium Genomic DNA in Oat Grains
4.4. Identification of Fusarium Chemotypes
4.5. Detection and Quantification of Mycotoxins
4.6. Sequencing of F. poae Tef-1α, Tri1, and Tri8 Genes
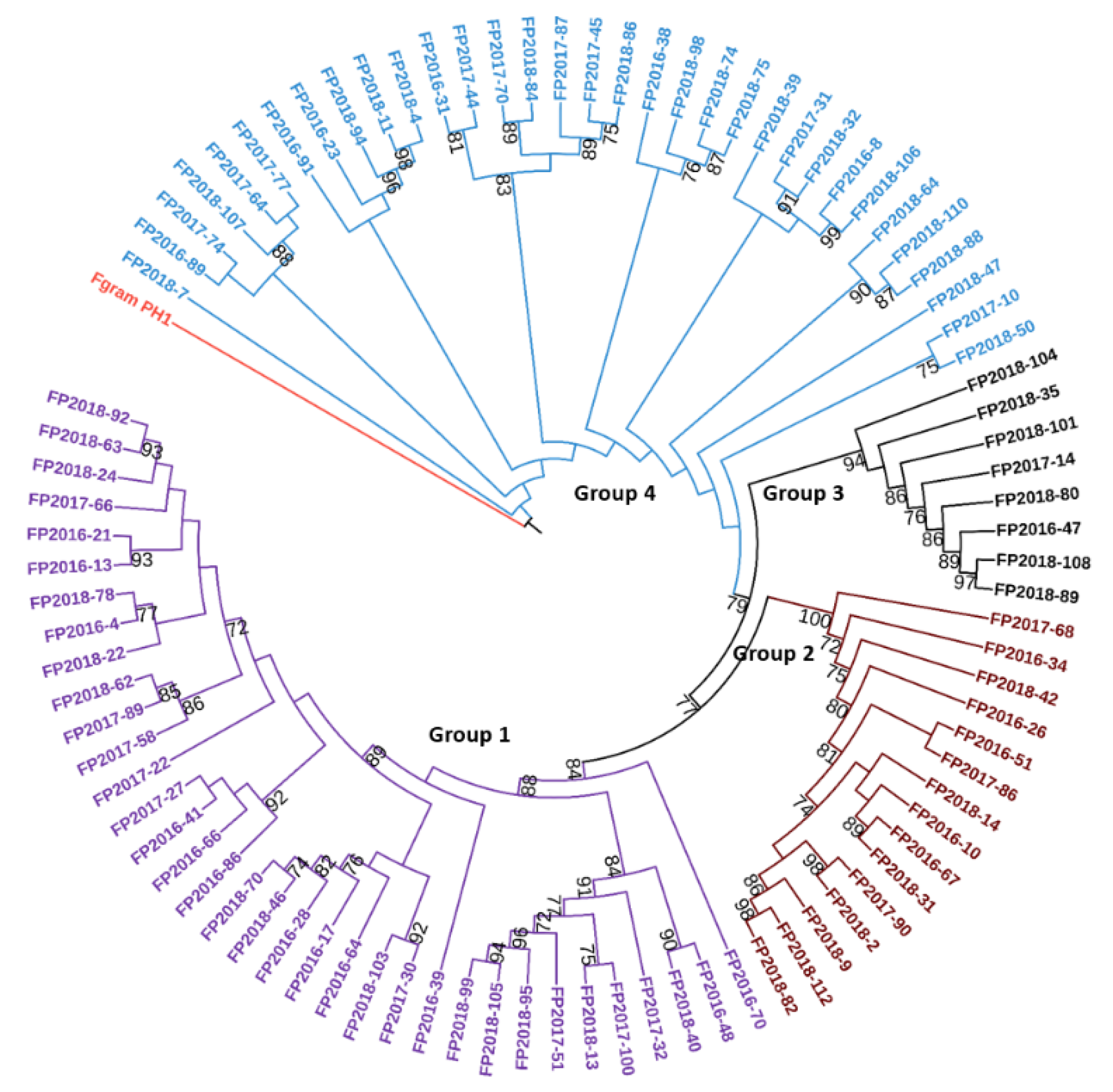
4.7. Phylogenetic Analysis of F. poae Strains Based on Tri1, Tri8, and Tef-1α Genes
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strychar, R. World oat production, trade, and usage. In Oats: Chemistry and Technology, 2nd ed.; American Association of Cereal Chemists, Inc. (AACC): St. Paul, MN, USA, 2016; pp. 1–10. [Google Scholar]
- Schöneberg, T.; Martin, C.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Keller, B.; Vogelgsang, S. Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 1608–1619. [Google Scholar] [CrossRef] [Green Version]
- Bjørnstad, Å.; Skinnes, H. Resistance to Fusarium infection in oats (Avena sativa L.). Cereal Res. Commun. 2008, 36, 57–62. [Google Scholar] [CrossRef]
- Tekauz, A.; Mitchell Fetch, J.; Rossnagel, B.G.; Savard, M.E. Progress in assessing the impact of Fusarium head blight on oat in western Canada and screening Avena germplasm for resistance. Cereal Res. Commun. 2008, 36 (Suppl. 6), 49–56. [Google Scholar] [CrossRef]
- Tekauz, A.; McCallum, B.; Ames, N.; Fetch, J.M. Fusarium head blight of oat—Current status in western Canada. Can. J. Plant Pathol. 2004, 26, 473–479. [Google Scholar] [CrossRef]
- Clear, R.M.; Patrick, S.K.; Gaba, D. Prevalence of fungi and fusariotoxins on oat seed from western Canada, 1995–1997. Can. J. Plant Pathol. 2000, 22, 310–314. [Google Scholar] [CrossRef]
- Tamburic-Ilincic, L. Fusarium species and mycotoxins associated with oat in southwestern Ontario, Canada. Can. J. Plant Sci. 2010, 90, 211–216. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.H.; Seifert, K.; Guo, W.; Blackwell, B.A.; Harris, L.J.; Overy, D.P. Prevalence of Fusarium species causing head blight of spring wheat, barley and oat in Ontario during 2001–2017. Can. J. Plant Pathol. 2019, 41, 392–402. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (SCAB) in small-grain cereals—A review. Plant Pathol. 2007, 44, 207–238. [Google Scholar] [CrossRef]
- Gilbert, J.; Tekauz, A. Review: Recent developments in research on fusarium head blight of wheat in Canada. Can. J. Plant Pathol. 2000, 22, 1–8. [Google Scholar] [CrossRef]
- Rohácik, T.; Hudec, K. Influence of agro-environmental factors on fusarium infestation and population structure in wheat kernels. Ann. Agric. Environ. Med. 2005, 12, 39–45. [Google Scholar]
- Stenglein, S. Fusarium poae: A pathogen that needs more attention. J. Plant Pathol. 2009, 91, 25–36. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in gereal grains—An update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, M.S.; Decundo, J.; Martinez, M.; Dieguez, S.N.; Moreyra, F.; Moreno, M.V.; Stenglein, S.A. Natural contamination with mycotoxins produced by Fusarium graminearum and Fusarium poae in malting barley in Argentina. Toxins 2018, 10, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinolfo, M.J.; Stenglein, S. Fusarium poae and mycotoxins: Potential risk for consumers. Bol. De La Soc. Argent. De Bot. 2014, 49, 5–20. [Google Scholar] [CrossRef]
- Stenglein, S.A.; Dinolfo, M.I.; Barros, G.; Bongiorno, F.; Chulze, S.N.; Moreno, M.V. Fusarium poae pathogenicity and mycotoxin accumulation on selected wheat and barley genotypes at a single location in Argentina. Plant Dis. 2014, 98, 1733–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, H. Deoxynivalenol and nivalenol toxicities in cultured cells: A review of comparative studies. Food Saf. 2018, 6, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Yagen, B.; Joffe, A.Z. Screeing of toxic isolates of Fusarium poae and Fusarium sporotrichiodes involved in causing alimentary toxic aleukia. Appl. Environ. Microbiol. 1976, 32, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Foroud, N.A.; Eudes, F. Trichothecenes in cereal grains. Int. J. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef] [Green Version]
- Tekle, S.; Dill-Macky, R.; Skinnes, H.; Tronsmo, A.M.; Bjcirnstad, C. Infection process of Fusarium graminearum in oats (Avena sativa L.). Eur. J. Plant Pathol. 2012, 132, 431–442. [Google Scholar] [CrossRef]
- Tini, F.; Beccari, G.; Onofri, A.; Ciavatta, E.; Gardiner, D.M.; Covarelli, L. Fungicides may have differential efficacies towards the main causal agents of Fusarium head blight of wheat. Pest Manag. Sci. 2020, 76, 3738–3748. [Google Scholar] [CrossRef]
- Tan, J.; De Zutter, N.; De Saeger, S.; De Boevre, M.; Tran, T.M.; van der Lee, T.; Waalwijk, C.; Willems, A.; Vandamme, P.; Ameye, M.J.F.I.P.S. Presence of the weakly pathogenic Fusarium poae in the Fusarium head blight disease complex hampers biocontrol and chemical control of the virulent Fusarium graminearum Pathogen. Front. Plant Sci. 2021, 12, 216. [Google Scholar] [CrossRef]
- Audenaert, K.; Landschoot, S.; Vanheule, A.; Waegeman, W.; De Baets, B.; Haesaert, G. Impact of fungicide timing on the composition of the Fusarium head blight disease complex and the presence of deoxynivalenol (DON) in wheat. In Fungicides—Beneficial and Harmful Aspects; Thajuddin, N., Ed.; IntechOpen: London, UK, 2011; pp. 79–98. [Google Scholar]
- Gilbert, J.; Clear, R.M.; Ward, T.J.; Gaba, D.; Tekauz, A.; Turkington, T.K.; Woods, S.M.; Nowicki, T.; O’Donnell, K. Relative aggressiveness and production of 3- or 15-acetyl deoxynivalenol and deoxynivalenol by Fusarium graminearum in spring wheat. Can. J. Plant Pathol. 2010, 32, 146–152. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Tittlemier, S.A.; Fernando, W.G.D. Nivalenol-producing Fusarium cerealis associated with fusarium head blight in winter wheat in Manitoba, Canada. Plant Pathol. 2015, 64, 988–995. [Google Scholar] [CrossRef]
- Amarasinghe, C.; Tamburic-Ilincic, L.; Gilbert, J.; Brûlé-Babel, A.L.; Dilantha Fernando, W. Evaluation of different fungicides for control of Fusarium head blight in wheat inoculated with 3-ADON and 15-ADON chemotypes of Fusarium graminearum in Canada. Can. J. Plant Pathol. 2013, 35, 200–208. [Google Scholar] [CrossRef]
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef]
- Valverde-Bogantes, E.; Bianchini, A.; Herr, J.R.; Rose, D.J.; Wegulo, S.N.; Hallen-Adams, H.E. Recent population changes of Fusarium head blight pathogens: Drivers and implications. Can. J. Plant Pathol. 2020, 42, 315–329. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.H.; Marchand, G.; Guo, W.; Ren, C.Z.; Savard, M.; McElroy, R.B. Timing of inoculation and Fusarium species affect the severity of Fusarium head blight on oat. Can. J. Plant Sci. 2015, 95, 517–524. [Google Scholar] [CrossRef]
- Beccari, G.; Arellano, C.; Covarelli, L.; Tini, F.; Sulyok, M.; Cowger, C. Effect of wheat infection timing on Fusarium head blight causal agents and secondary metabolites in grain. Int. J. Food Microbiol. 2019, 290, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Cooke, B.M.; Doohan, F.M.; Brennan, J.; Monaghan, S.; Moretti, A.; Mule, G.; et al. Relationship between the fungal complex causing Fusarium head blight of wheat and environmental conditions. Phytopathology 2008, 98, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Ameye, M.; Audenaert, K.; De Zutter, N.; Steppe, K.; Vanhaecke, L.; De Vleesschauwer, D.; Haesaert, G.; Smagghe, G. Priming of wheat with the green leaf volatile Z-3-hexenyl acetate enhances defense against Fusarium graminearum but boosts deoxynivalenol production. Plant Physiol. 2015, 167, 1671–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; Ameye, M.; Landschoot, S.; De Zutter, N.; De Saeger, S.; De Boevre, M.; Abdallah, M.F.; Van der Lee, T.; Waalwijk, C.; Audenaert, K. At the scene of the crime: New insights into the role of weakly pathogenic members of the fusarium head blight disease complex. Mol. Plant Pathol. 2020, 21, 1559–1572. [Google Scholar] [CrossRef]
- Gao, T.; Zhou, H.; Zhou, W.; Hu, L.; Chen, J.; Shi, Z.Q. The fungicidal activity of thymol against Fusarium graminearum via inducing lipid peroxidation and disrupting ergosterol biosynthesis. Molecules 2016, 21, 770. [Google Scholar] [CrossRef] [Green Version]
- Gräfenhan, T.; Patrick, S.K.; Roscoe, M.; Trelka, R.; Gaba, D.; Chan, J.M.; McKendry, T.; Clear, R.M.; Tittlemier, S.A. Fusarium damage in cereal grains from Western Canada. 1. Phylogenetic analysis of moniliformin-producing Fusarium species and their natural occurrence in mycotoxin-contaminated wheat, oats, and rye. J. Agric. Food Chem. 2013, 61, 5425–5437. [Google Scholar] [CrossRef] [PubMed]
- Tittlemier, S.A.; Blagden, R.; Chan, J.M.; Roscoe, M.; Pleskach, K. A multi-year survey of mycotoxins and ergosterol in Canadian oats. Mycotoxin Res. 2020, 36, 103–114. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 401/2006 of 23, Laying down the Methods of Sampling and Analysis for the Official Control of the Levels of Mycotoxins in Food Stuffs, February 2006; European Commission: Brussels, Belgium, 2006; Volume 70, pp. 12–34. [Google Scholar]
- Campbell, H.; Choo, T.M.; Vigier, B.; Underhill, L. Mycotoxins in barley and oat samples from eastern Canada. Can. J. Plant Sci. 2000, 80, 977–980. [Google Scholar] [CrossRef]
- Visconti, A.; Minervini, F.; Lucivero, G.; Gambatesa, V. Cytotoxic and immunotoxic effects of Fusarium mycotoxins using a rapid colorimetric bioassay. Mycopathologia 1991, 113, 181–186. [Google Scholar] [CrossRef]
- Minervini, F.; Fornelli, F.; Flynn, K.M. Toxicity and apoptosis induced by the mycotoxins nivalenol, deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line. Toxicol. Vitr. 2004, 18, 21–28. [Google Scholar] [CrossRef]
- Yang, Y.X.; Yu, S.; Tan, Y.L.; Liu, N.; Wu, A. Individual and combined cytotoxic effects of co-occurring deoxynivalenol family mycotoxins on human gastric epithelial cells. Toxins 2017, 9, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheat, S.; Pinton, P.; Cossalter, A.; Cognie, J.; Vilariño, M.; Callu, P.; Raymond-Letron, I.; Oswald, I.P.; Kolf-Clauw, M. The mycotoxins deoxynivalenol and nivalenol show in vivo synergism on jejunum enterocytes apoptosis. Food Chem. Toxicol. 2016, 87, 45–54. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Paavanen-Huhtala, S.; Jestoi, M.; Parikka, P.; Hietaniemi, V.; Gagkaeva, T.; Sarlin, T.; Haikara, A.; Laaksonen, S.; Rizzo, A. Real-time PCR detection and quantification of Fusarium poae, F. graminearum, F. sporotrichioides and F. langsethiae in cereal grains in Finland and Russia. Arch. Phytopathol. Plant Prot. 2008, 41, 243–260. [Google Scholar] [CrossRef]
- Martin, C.; Schöneberg, T.; Vogelgsang, S.; Mendes Ferreira, C.S.; Morisoli, R.; Bertossa, M.; Bucheli, T.D.; Mauch-Mani, B.; Mascher, F. Responses of oat grains to Fusarium poae and F. langsethiae Infections and mycotoxin contaminations. Toxins 2018, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martos, P.; Thompson, W.; Diaz, G. Multi residue mycotoxin analysis in wheat, barley, oats, rye and maize grain by high-performance liquid chromatography-tandem mass spectrometry. World Mycotoxin J. 2010, 3, 205–223. [Google Scholar] [CrossRef]
- Pageau, D.; Lafond, J.; Lajeunesse, J.; Savard, M. Impact of previous cultivation and nitrogen fertilization on deoxynivalenol content in barley. Can. J. Plant Pathol. 2008, 30, 397–403. [Google Scholar] [CrossRef]
- Beyer, M.; Klix, M.B.; Klink, H.; Verreet, J.A. Quantifying the effects of previous crop, tillage, cultivar and triazole fungicides on the deoxynivalenol content of wheat grain—A review. J. Plant Dis. Prot. 2006, 113, 241–246. [Google Scholar] [CrossRef]
- Knutsen, A.K.; Torp, M.; Holst-Jensen, A. Phylogenetic analyses of the Fusarium poae, Fusarium sporotrichioides and Fusarium langsethiae species complex based on partial sequences of the translation elongation factor-1 alpha gene. Int. J. Food Microbiol. 2004, 95, 287–295. [Google Scholar] [CrossRef]
- Dinolfo, M.I.; Stenglein, S.A.; Moreno, M.V.; Nicholson, P.; Jennings, P.; Salerno, G.L. ISSR markers detect high genetic variation among Fusarium poae isolates from Argentina and England. Eur. J. Plant Pathol. 2010, 127, 483–491. [Google Scholar] [CrossRef]
- Vanheule, A.; De Boevre, M.; Moretti, A.; Scauflaire, J.; Munaut, F.; De Saeger, S.; Bekaert, B.; Haesaert, G.; Waalwijk, C.; van der Lee, T.; et al. Genetic divergence and chemotype diversity in the Fusarium head blight pathogen Fusarium poae. Toxins 2017, 9, 255. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Enkerli, J.; Jenny, E.; Roffler, S.; Widmer, F. Characterization of Fusarium poae microsatellite markers on strains from Switzerland and other countries. J. Phytopathol. 2011, 159, 197–200. [Google Scholar] [CrossRef]
- Vanheule, A.; Audenaert, K.; Warris, S.; van de Geest, H.; Schijlen, E.; Höfte, M.; De Saeger, S.; Haesaert, G.; Waalwijk, C.; van der Lee, T. Living apart together: Crosstalk between the core and supernumerary genomes in a fungal plant pathogen. BMC Genom. 2016, 17, 670. [Google Scholar] [CrossRef] [Green Version]
- Witte, T.; Harris, L.J.; Nguyen, H.; Hermans, A.; Johnston, A.; Sproule, A.; Dettman, J.; Boddy, C.; Overy, D. Apicidin biosynthesis ss linked to accessory chromosomes in Fusarium poae isolates. BMC Genom. 2021, 22, 1–18. [Google Scholar] [CrossRef]
- Abbas, H.K.; Mirocha, C.J. Isolation and purification of a hemorrhagic factor (wortmannin) from Fusarium oxysporum (N17B). Appl. Environ. Microbiol. 1988, 54, 1268–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; McCormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New tricks of an old enemy: Isolates of Fusarium graminearum produce a type A trichothecene mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; McCormick, S.P.; Waalwijk, C.; van der Lee, T.; Proctor, R.H. The genetic basis for 3-ADON and 15-ADON trichothecene chemotypes in Fusarium. Fungal Genet. Biol. 2011, 48, 485–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Blackwell Pub.: Ames, IA, USA, 2006. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
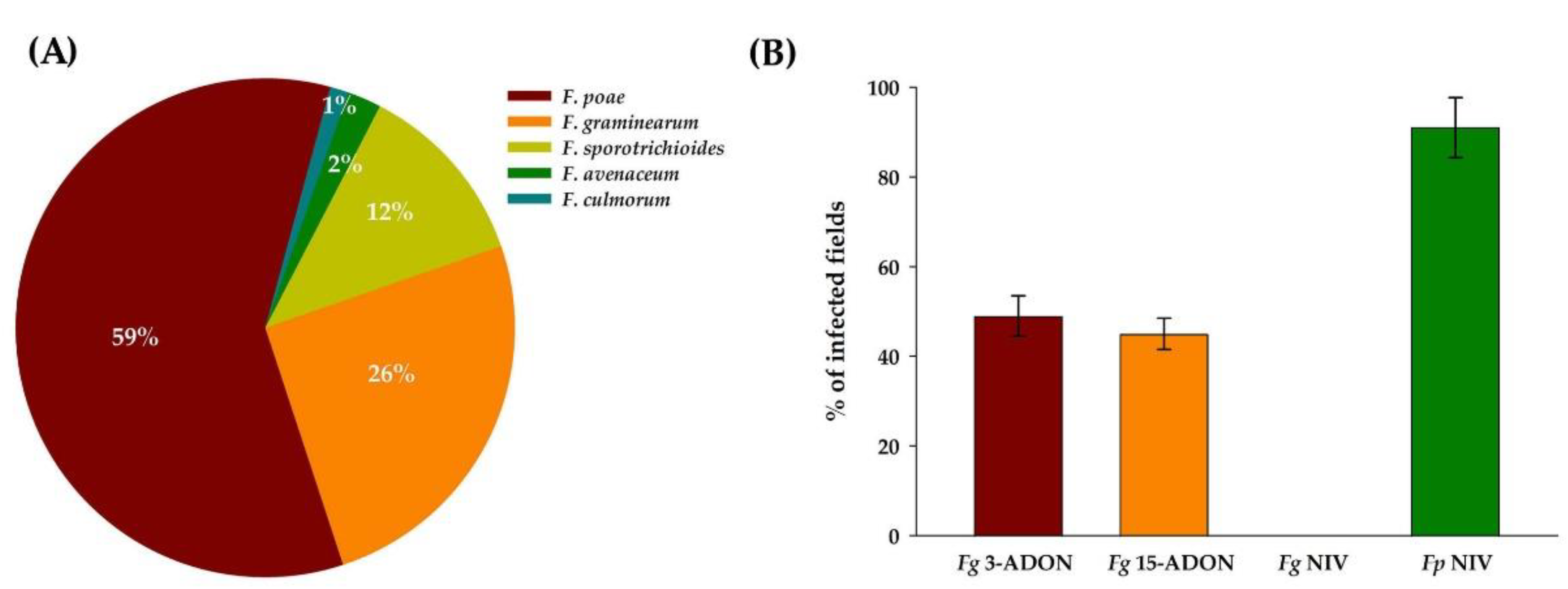
| Fusarium Species | % of Infected Fields | % of Infected Kernels | ||||||
|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | Three-Year Mean | 2016 | 2017 | 2018 | Three-Year Mean | |
| F. poae | 72.0 | 65.0 | 69.0 | 68.7 ± 3.5 | 61.0 | 62.0 | 53.0 | 58.7 ± 4.9 |
| F. graminearum | 28.0 | 25.0 | 48.0 | 33.7 ± 9.6 | 22.3 | 14.8 | 12.8 | 16.6 ± 5.0 |
| F. sporotrichioides | 9.0 | 7.0 | 15.0 | 10.3 ± 4.2 | 8.0 | 22.0 | 12.0 | 14.0 ± 7.2 |
| F. avenaceum | 2.3 | 3.3 | 1.3 | 2.3 ± 11 | 0.4 | 0.5 | 0.2 | 0.4 ± 0.2 |
| F. culmorum | 2.3 | 1.7 | 0.0 | 1.3 ± 1.2 | 0.4 | 0.3 | 0.0 | 0.2 ± 0.2 |
| Total no. of fields/kernels | 43 | 60 | 65 | 2150 | 3000 | 3250 | ||
| Year | 2016 | 2017 | 2018 | |||
|---|---|---|---|---|---|---|
| n-Samples/Fields | 43 | 60 | 65 | |||
| Species | Range | Mean | Range | Mean | Range | Mean |
| F. poae | 0.01–1.58 * | 0.13 ± 0.05 a ** | 0.02–1.35 | 0.12 ± 0.07 a | 0.00–1.37 | 0.08 ± 0.01 a |
| F. graminearum | 0.01–1.93 | 0.05 ± 0.01 b | 0.01–0.80 | 0.07 ± 0.01 a | 0.01–0.31 | 0.07 ± 0.01 a |
| F. sporotrichioides | 0.01 -0.38 | 0.02 ± 0.01 b | 0.01–0.05 | 0.01 ± 0.01 b | 0.00–0.15 | 0.01 ± 0.01 b |
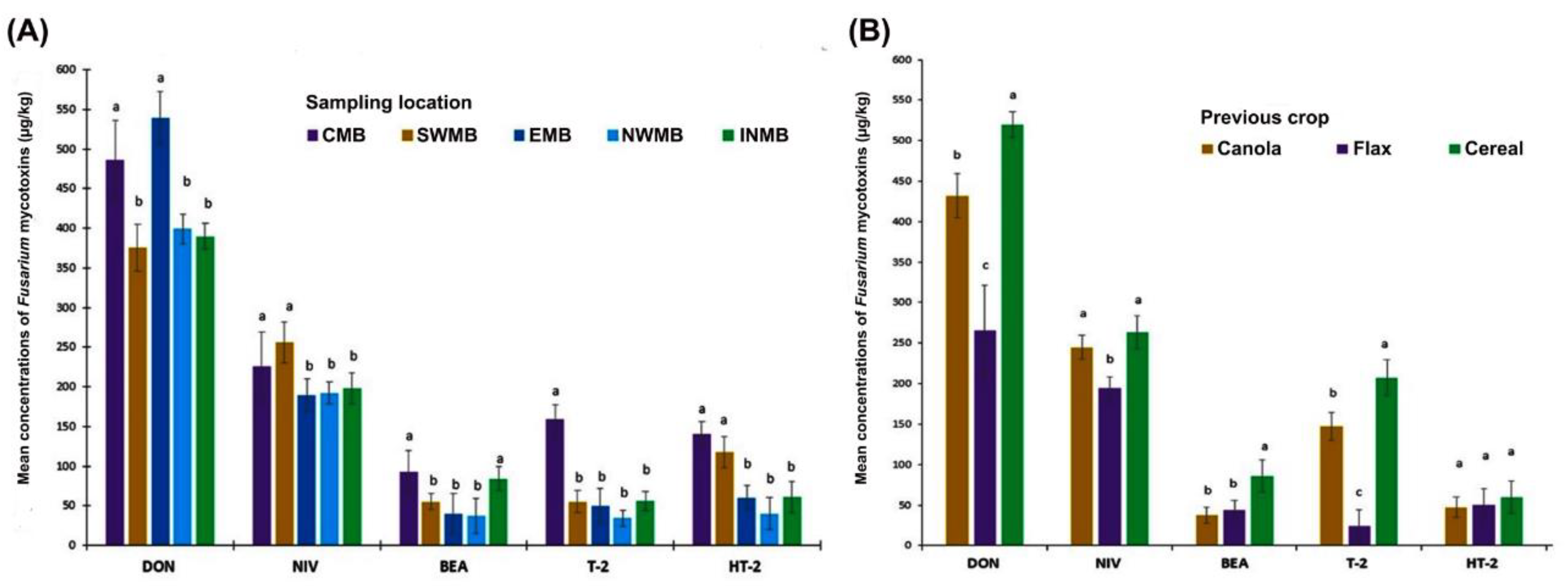
| Year | 2016 | 2017 | 2018 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| N-Samples/Fields | 43 | 60 | 65 | ||||||
| % of Fields Above LOD | Maximum (µg/kg) | Mean (µg/kg) | % of Fields Above LOD | Maximum (µg/kg) | Mean (µg/kg) | % of Fields Above LOD | Maximum (µg/kg) | Mean (µg/kg) | |
| DON | 85 | 4143 | 604 ± 87 | 41 | 1881 | 569 ± 104 | 40 | 632 | 253 ± 20 |
| NIV | 100 | 865 | 252 ± 27 | 78 | 795 | 234 ± 16 | 98 | 581 | 145 ± 12 |
| T-2 | 77 | 1155 | 43 ± 18 | 81 | 973 | 53 ± 16 | 25 | 794 | 22 ± 12 |
| HT-2 | 35 | 1100 | 60 ± 29 | 13 | 419 | 26 ± 10 | 17 | 654 | 23 ± 11 |
| BEA | 90 | 119 | 25 ± 4 | 80 | 159 | 41 ± 4 | 71 | 169 | 24 ± 5 |
| MON | 32 | 349 | 31 ± 11 | 20 | 533 | 30 ± 11 | 11 | 209 | 12 ± 5 |
| DAS | 15 | 25 | 8 ± 1.1 | 7 | 25 | 7 ± 0.6 | 5 | 25 | 6 ± 0.5 |
| ENNs | 43 | 1605 | 65 ± 6 | 39 | 93 | 16 ± 2 | 17 | 128 | 10 ± 1 |
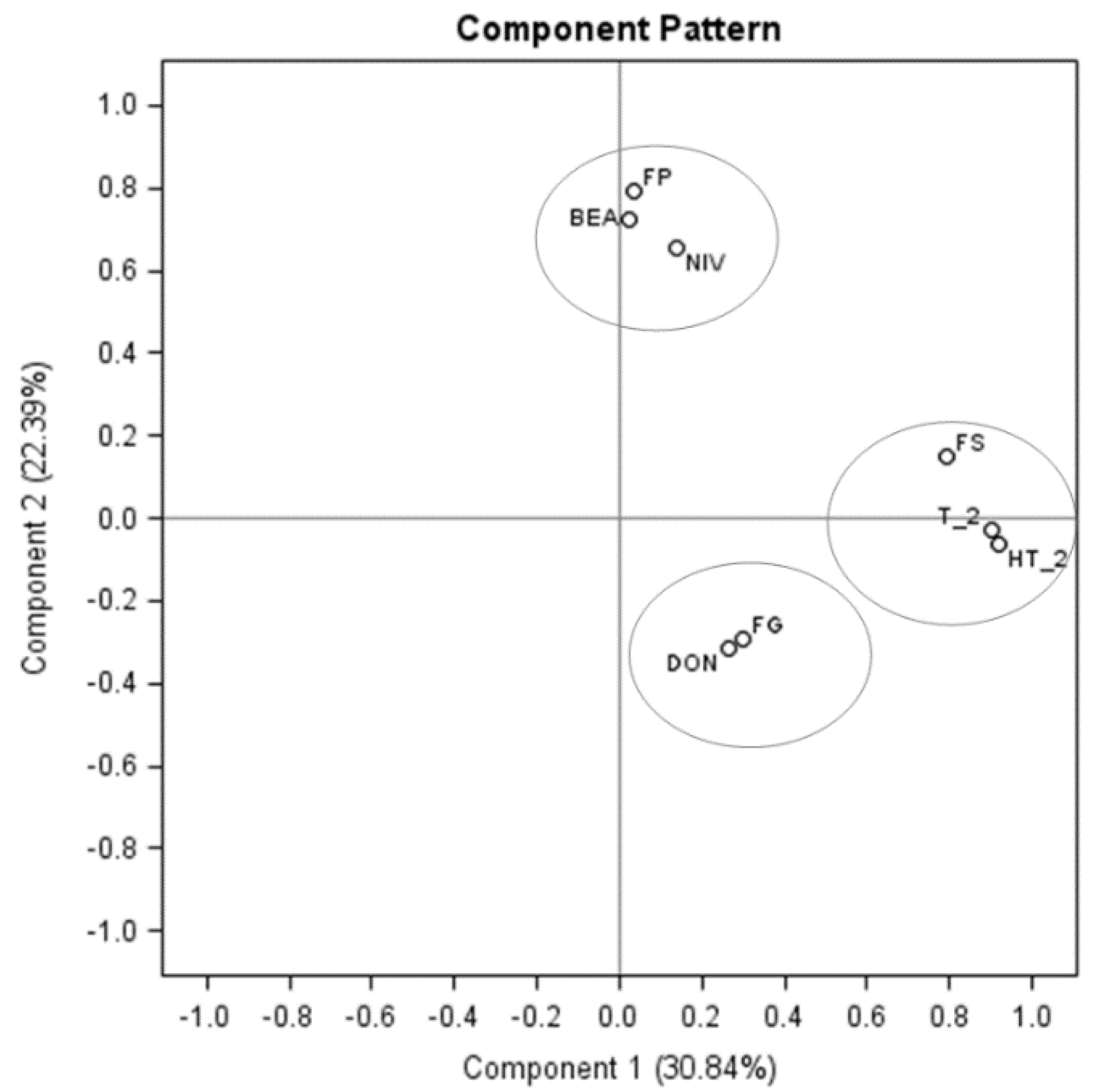
| †p-Value | |||||
|---|---|---|---|---|---|
| Fusarium Mycotoxin Levels | |||||
| Source of variation | DON | NIV | BEA | T-2 | HT-2 |
| Sample locations | * | * | * | ns | * |
| Crop rotations | * | ns | * | * | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.N.; Tabassum, M.; Banik, M.; Daayf, F.; Fernando, W.G.D.; Harris, L.J.; Sura, S.; Wang, X. Naturally Occurring Fusarium Species and Mycotoxins in Oat Grains from Manitoba, Canada. Toxins 2021, 13, 670. https://doi.org/10.3390/toxins13090670
Islam MN, Tabassum M, Banik M, Daayf F, Fernando WGD, Harris LJ, Sura S, Wang X. Naturally Occurring Fusarium Species and Mycotoxins in Oat Grains from Manitoba, Canada. Toxins. 2021; 13(9):670. https://doi.org/10.3390/toxins13090670
Chicago/Turabian StyleIslam, M. Nazrul, Mourita Tabassum, Mitali Banik, Fouad Daayf, W. G. Dilantha Fernando, Linda J. Harris, Srinivas Sura, and Xiben Wang. 2021. "Naturally Occurring Fusarium Species and Mycotoxins in Oat Grains from Manitoba, Canada" Toxins 13, no. 9: 670. https://doi.org/10.3390/toxins13090670
APA StyleIslam, M. N., Tabassum, M., Banik, M., Daayf, F., Fernando, W. G. D., Harris, L. J., Sura, S., & Wang, X. (2021). Naturally Occurring Fusarium Species and Mycotoxins in Oat Grains from Manitoba, Canada. Toxins, 13(9), 670. https://doi.org/10.3390/toxins13090670