Insect Body Defence Reactions against Bee Venom: Do Adipokinetic Hormones Play a Role?

 , , ,
, , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
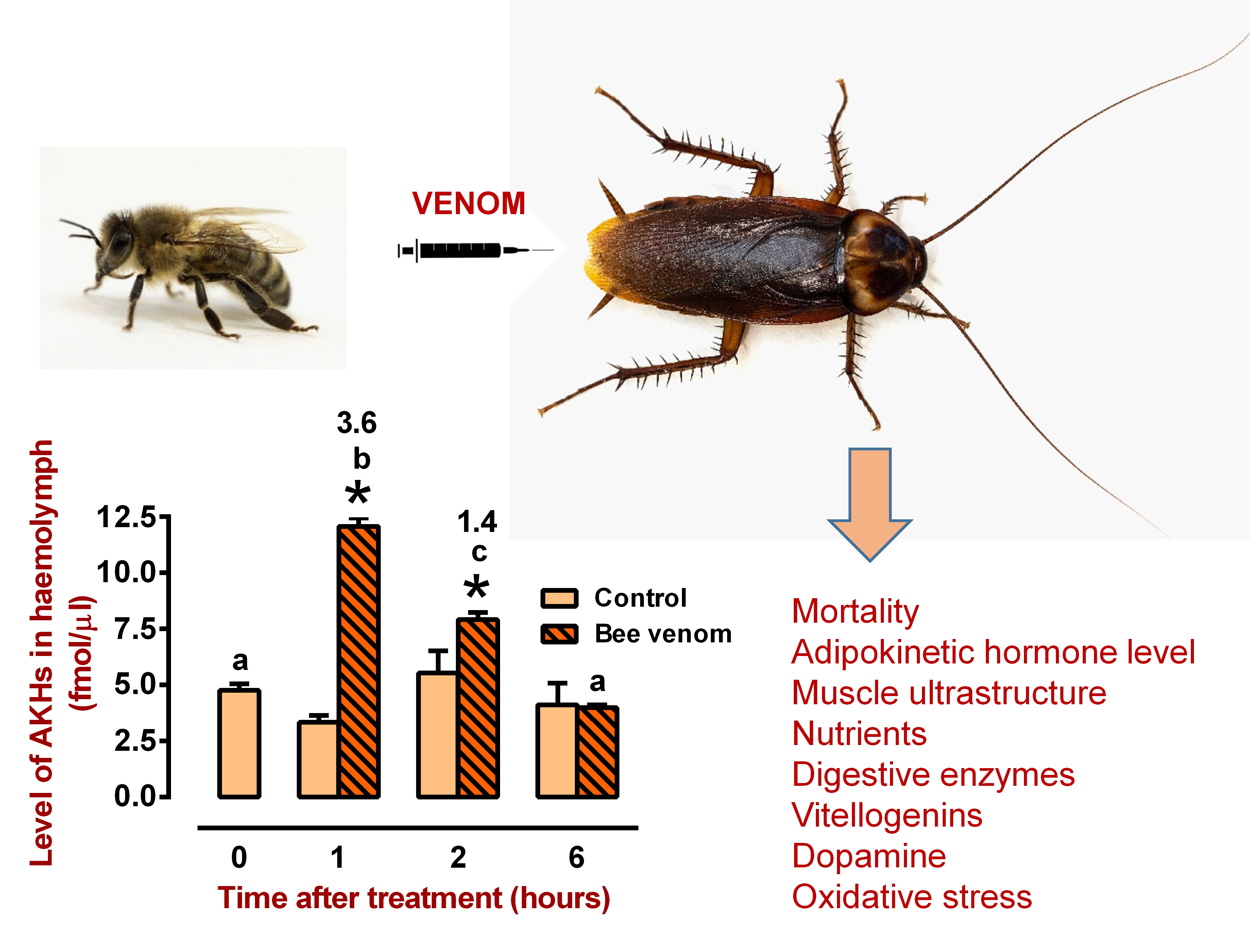
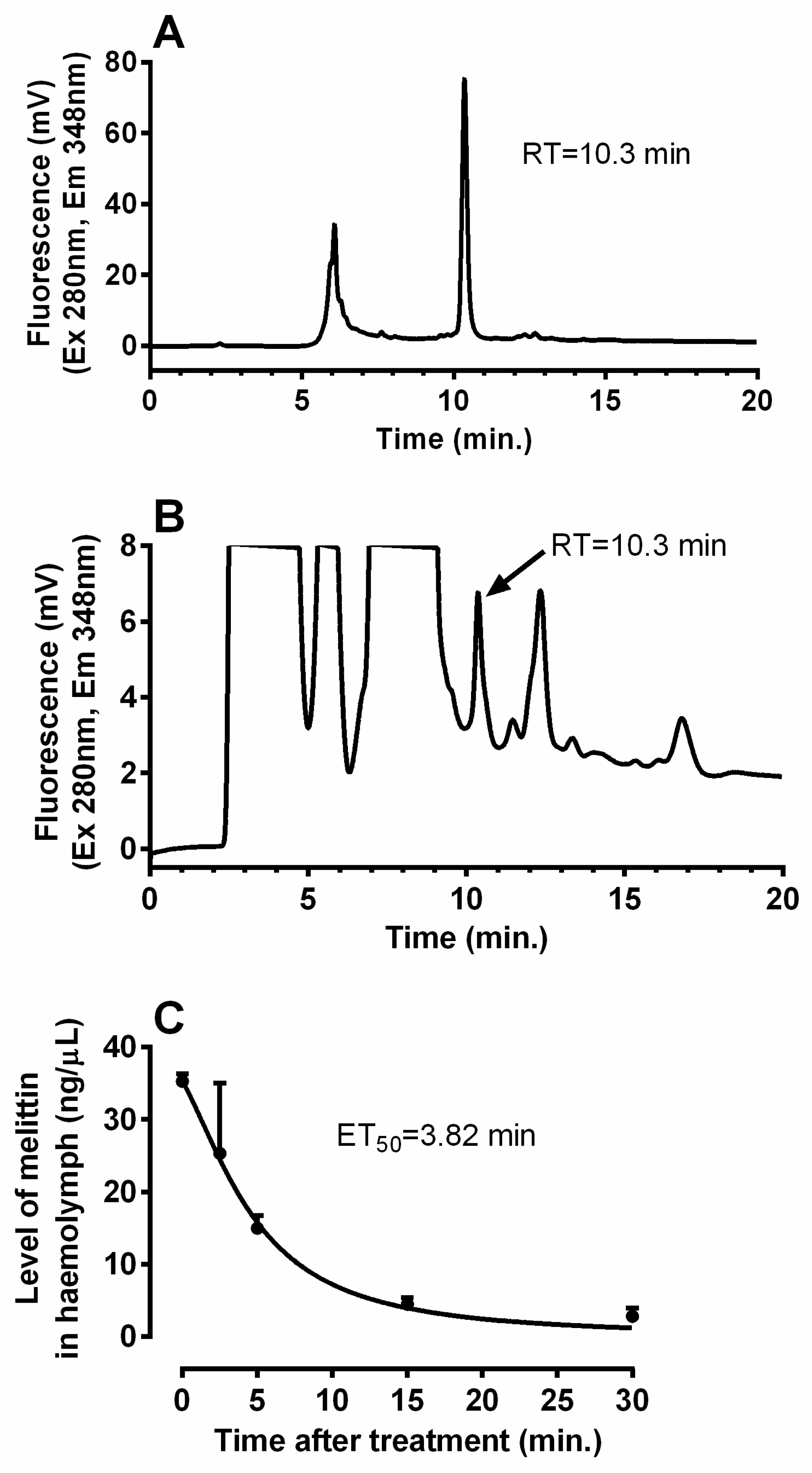
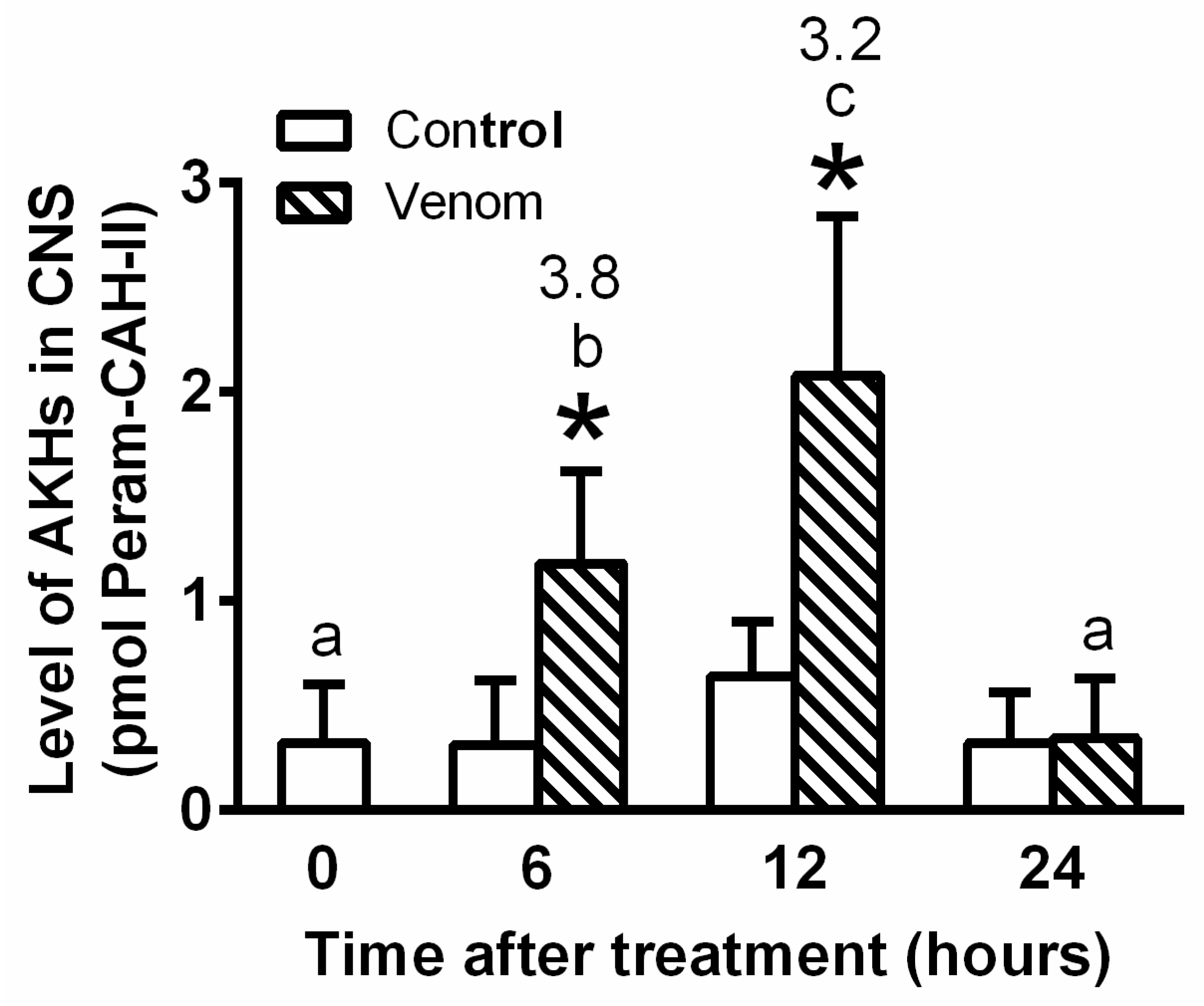
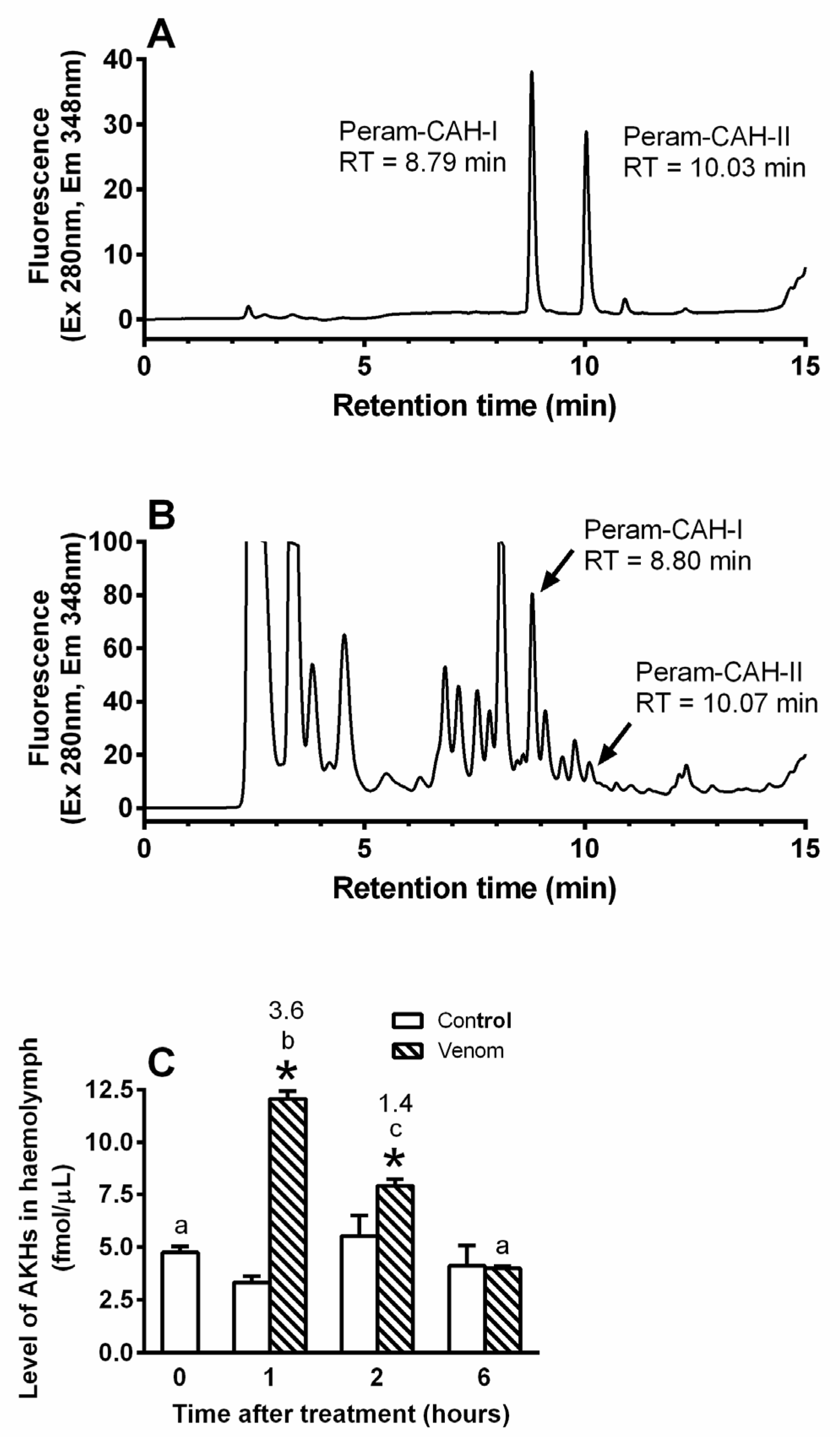
2. Results
2.1. General Interactions between Venom and AKH
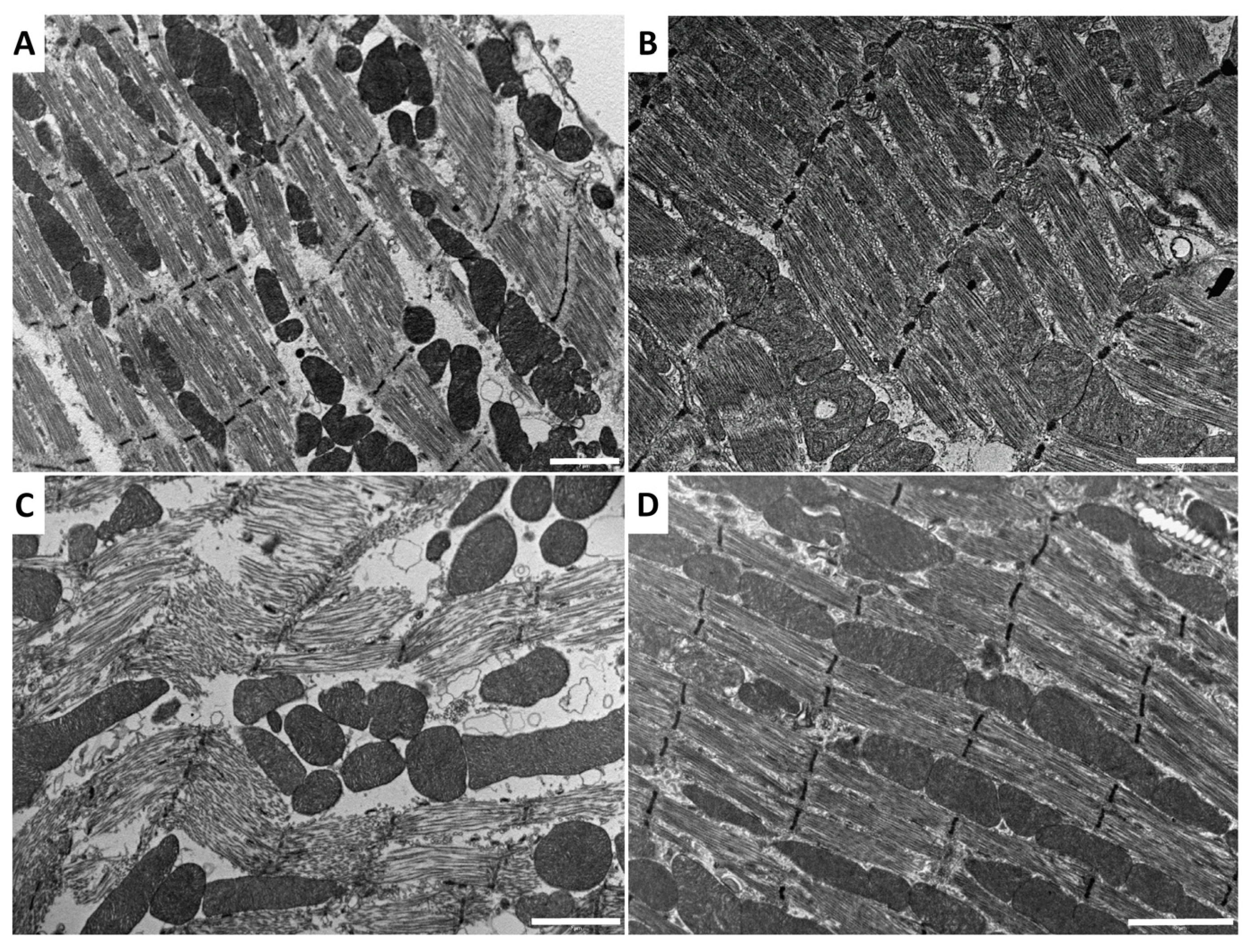
2.2. Effect of Venom and AKH on Muscle Ultrastructure
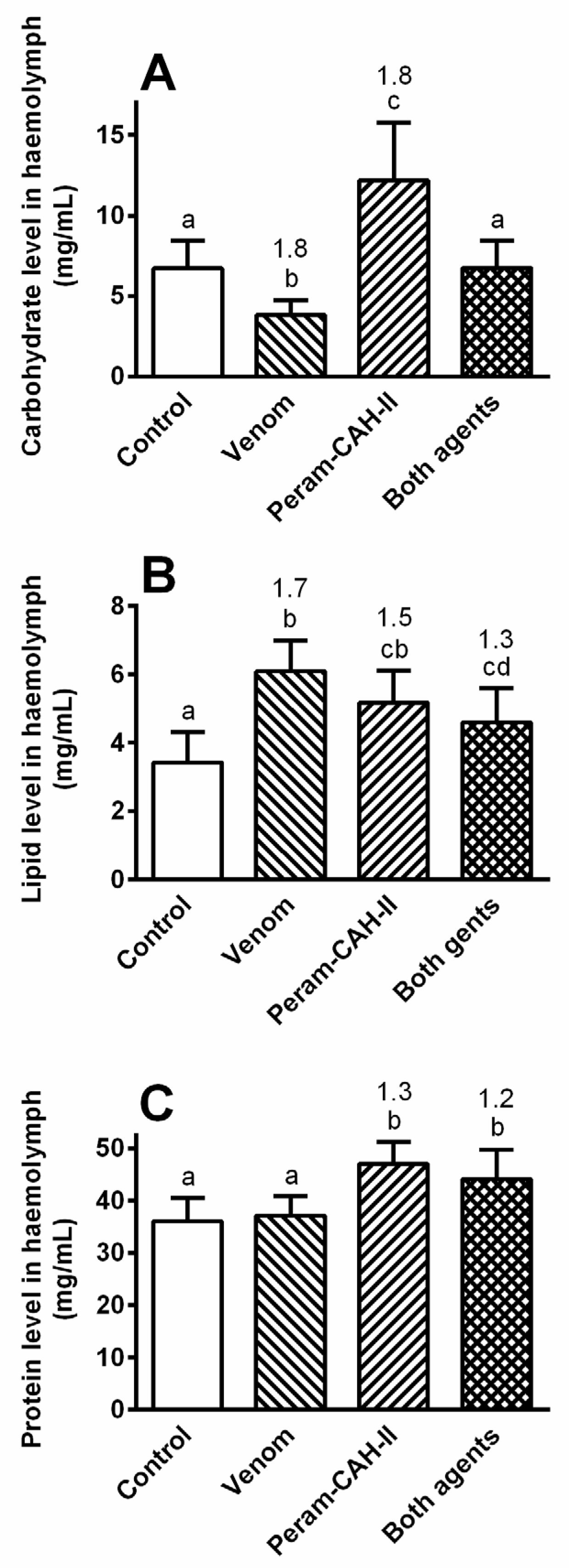
2.3. Effect of Venom and AKH on Selected Metabolites
3. Discussion
3.1. Interactions between Bee Venom and AKH
3.2. Effect of Venom/AKH on Physiological Processes
4. Conclusions
5. Materials and Methods
5.1. Experimental Animals
5.2. Venom and Hormone Treatments
5.3. Turnover of Melittin in the Cockroach Body
5.4. Quantification of Peram-CAH-I and Peram-CAH-II in Cockroach Body
5.5. Transmission Electron Microscopy (TEM) of Cockroach Muscle
5.6. Quantification of Nutrients in Haemolymph
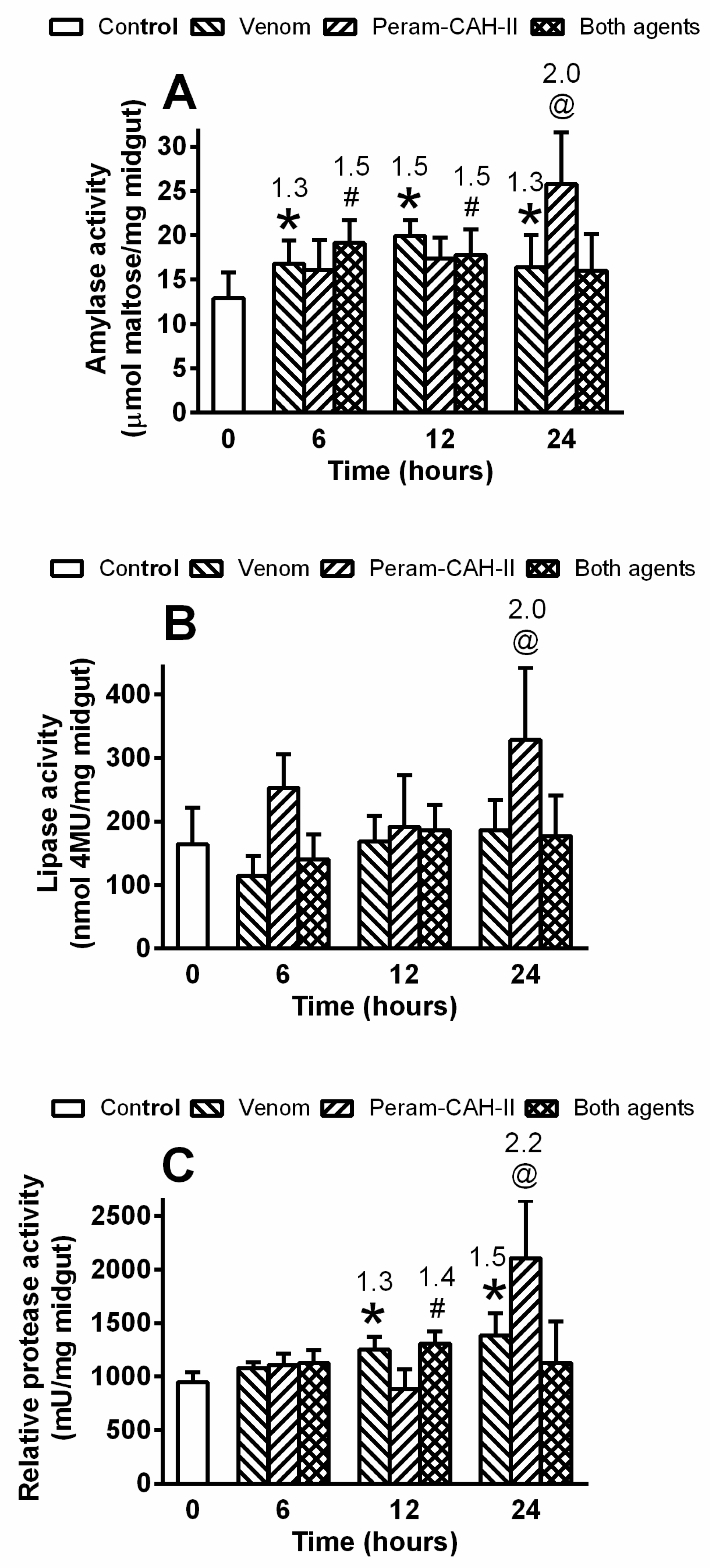
5.7. Activity of Gut Enzymes
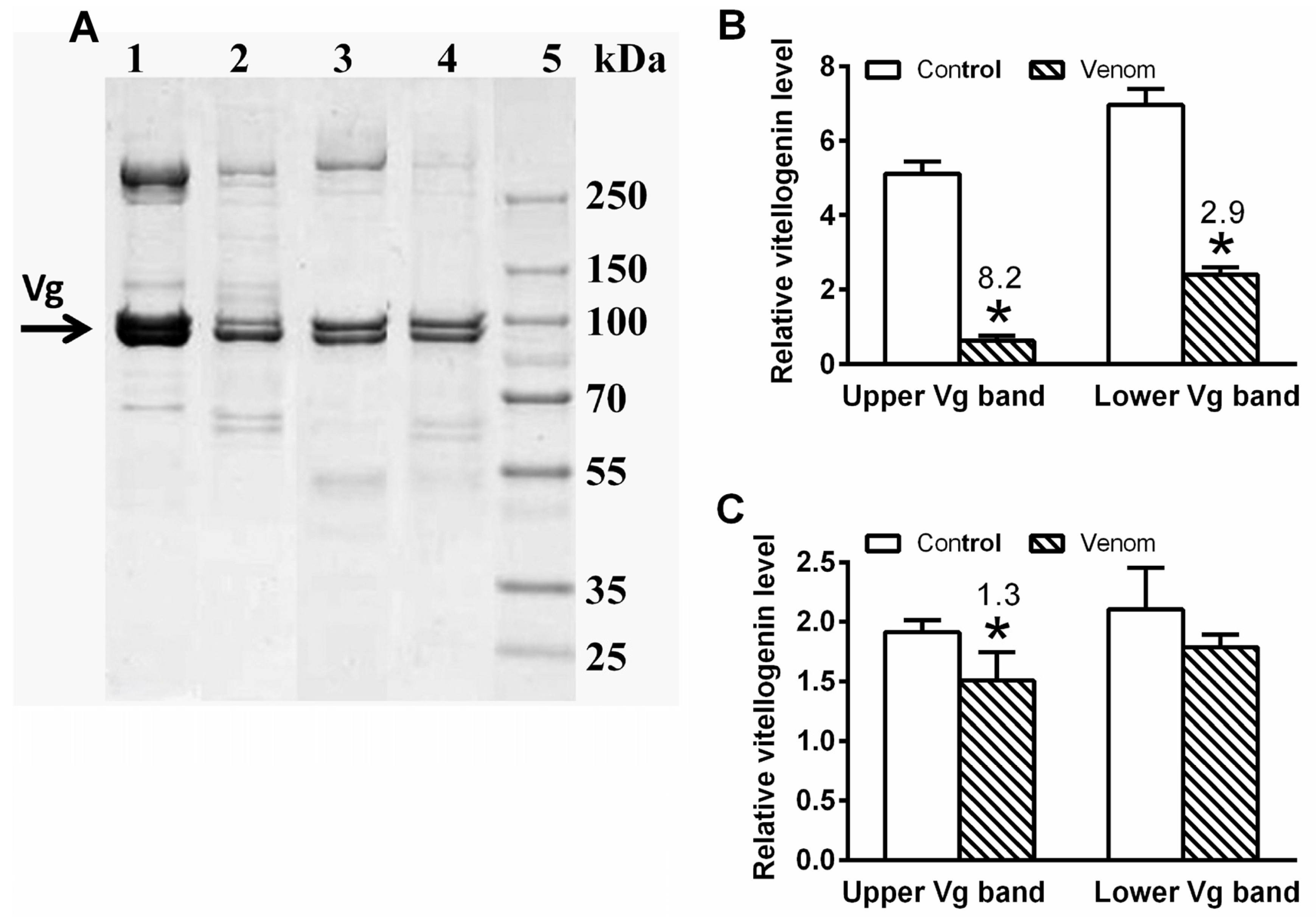
5.8. Vitellogenin Level
5.9. Dopamine and Glutathione Determination
5.10. Data Presentation and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lubawy, J.; Urbanski, A.; Mrówczynska, L.; Matuszewska, E.; Swiatły-Błaszkiewicz, A.; Matysiak, J.; Rosinski, R. The influence of bee venom melittin on the functioning of the immune system and the contractile activity of the insect heart—A preliminary study. Toxins 2019, 11, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.; Girald, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [Green Version]
- Son, D.J.; Lee, J.W.; Lee, Y.H.; Song, H.S.; Lee, C.K.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lariviere, W.R. The nociceptive and anti-nociceptive effects of bee venom injection and therapy: A double-edged sword. Prog. Neurobiol. 2010, 92, 151–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florea, A.; Varga, A.P.; Matei, H.V. Ultrastructural variability of mitochondrial cristae induced in vitro by bee (Apis mellifera) venom and its derivatives, melittin and phospholipase A2, in isolated rat adrenocortical mitochondria. Micron 2018, 112, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Horan, K.L.; Adamski, S.W.; Ayele, W.; Langone, J.J.; Grega, G.J. Evidence that prolonged histamine suffusions produce transient increases in vascular permeability subsequent to the formation of venular macro-molecular leakage sites. Proof of the Majno-Palade hypothesis. Am. J. Pathol. 1986, 123, 570–576. [Google Scholar]
- Park, H.G.; Lee, S.K.; Kim, B.Y.; Yoon, H.J.; Choi, Y.S.; Lee, K.Y.; Wan, H.; Li, J.; Jin, B.R. Honeybee (Apis cerana) vitellogenin acts as antimicrobial and antioxidant agent in the body and venom. Dev. Comp. Immunol. 2018, 85, 51–60. [Google Scholar] [CrossRef]
- Salmela, H.; Amdam, G.V.; Freitak, D. Transfer of immunity from mother to offspring is mediated via egg-yolk protein vitellogenin. PLoS Pathog. 2015, 11, e1005015. [Google Scholar] [CrossRef] [Green Version]
- Kodrík, D.; Ibrahim, E.; Gautam, U.K.; Čapková-Frydrychová, R.; Bednářová, A.; Krištůfek, V.; Jedlička, P. Changes in vitellogenin expression caused by nematodal and fungal infections in insects. J. Exp. Biol. 2019, 222, jeb202853. [Google Scholar] [CrossRef] [Green Version]
- Storey, K.B. Adventures in oxygen metabolism. Comp. Biochem. Physiol. C 2004, 139, 359–369. [Google Scholar] [CrossRef]
- Gäde, G.; Hoffmann, K.H.; Spring, J.H. Hormonal regulation in insects: Facts, gaps, and future directions. Physiol. Rev. 1997, 77, 963–1032. [Google Scholar] [CrossRef]
- Kodrík, D. Adipokinetic hormone functions that are not associated with insect flight. Physiol. Entomol. 2008, 33, 171–180. [Google Scholar] [CrossRef]
- Kodrík, D.; Bednářová, A.; Zemanová, M.; Krishnan, N. Hormonal regulation of response to oxidative stress in insects—An update. Int. J. Mol. Sci. 2015, 16, 25788–25816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodrík, D.; Vinokurov, K.; Tomčala, A.; Socha, R. The effect of adipokinetic hormone on midgut characteristics in Pyrrhocoris apterus L. (Heteroptera). J. Insect Physiol. 2012, 58, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Bil, M.; Broeckx, V.; Landuyt, B.; Huybrechts, R. Differential peptidomics highlights adipokinetic hormone as key player in regulating digestion in anautogenous flesh fly, Sarcophaga crassipalpis. Gen. Comp. Endocrinol. 2014, 208, 49–56. [Google Scholar] [CrossRef]
- Bodláková, K.; Jedlička, P.; Kodrík, D. Adipokinetic hormones control amylase activity in the cockroach (Periplaneta americana) gut. Insect Sci. 2017, 24, 259–269. [Google Scholar] [CrossRef]
- Van der Horst, D.J.; Van Marrewijk, W.J.A.; Diederen, H.B. Adipokinetic hormones of insect: Release, signal transduction, and responses. Int. Rev. Cytol. 2001, 211, 179–240. [Google Scholar]
- Socha, R.; Kodrík, D.; Zemek, R. Stimulatory effects of bioamines norepinephrine and dopamine on locomotion of Pyrrhocoris apterus (L.): Is the adipokinetic hormone involved? Comp. Biochem. Physiol. B 2008, 151, 305–310. [Google Scholar] [CrossRef]
- Scarborough, R.M.; Jamieson, G.C.; Kalish, F.; Kramer, J.S.; McEnroe, G.A.; Miller, C.A.; Schooley, D. Isolation and primary structure of two peptides with cardioacceleratory and hyperglycemic activity from the corpora cardiaca of Periplaneta americana. Proc. Natl. Acad. Sci. USA 1984, 81, 5575–5579. [Google Scholar] [CrossRef] [Green Version]
- Michitsch, J.; Steel, J.E. Carbohydrate and lipid metabolism in cockroach (Periplaneta americana) fat body are both activated by low and similar concentrations of Peram-AKH II. Peptides 2008, 29, 226–234. [Google Scholar] [CrossRef]
- Culliney, T.W. Geological history and evolution of the honey bee. Am. Bee J. 1983, 123, 580–585. [Google Scholar]
- Engel, M.S. Fossil honey bees and evolution in the genus Apis (Hymenoptera: Apidae). Apidologie 1998, 29, 265–281. [Google Scholar] [CrossRef]
- Oudejans, R.C.H.M.; Vroemen, S.F.; Jansen, R.F.R.; Van der Horst, D.J. Locust adipokinetic hormones: Carrier-independent transport and differential inactivation at physiological concentrations during rest and flight. Proc. Natl. Acad. Sci. USA 1996, 93, 8654–8659. [Google Scholar] [CrossRef] [Green Version]
- Van der Horst, D.J.; Van Marrewijk, W.J.A.; Vullings, H.G.B.; Diederen, J.H.B. Metabolic neurohormones: Release, signal transduction and physiological responses of adipokinetic hormones in insects. Eur. J. Entomol. 1999, 96, 299–308. [Google Scholar]
- Goldsworthy, G.J.; Kodrík, D.; Comley, R.; Lightfoot, M. A quantitative study of the adipokinetic hormone of the firebug, Pyrrhocoris apterus. J. Insect Physiol. 2002, 48, 1103–1108. [Google Scholar] [CrossRef] [Green Version]
- Kodrík, D.; Socha, R. The effect of insecticide on adipokinetic hormone titre in insect body. Pest Manag. Sci. 2005, 61, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Velki, M.; Kodrík, D.; Večeřa, J.; Hackenberger, B.K.; Socha, R. Oxidative stress elicited by insecticides: A role for the adipokinetic hormone. Gen. Comp. Endocrinol. 2011, 172, 77–84. [Google Scholar] [CrossRef]
- Kodrík, D.; Plavšin, I.; Velki, M.; Stašková, T. Enhancement of insecticide efficacy by adipokinetic hormones. In Insecticides: Occurrence, Global Threats and Ecological Impact; Montgomery, J., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2015; pp. 77–91. [Google Scholar]
- Ibrahim, E.; Hejníková, M.; Shaik, H.A.; Doležel, D.; Kodrík, D. Adipokinetic hormone activities in insect body infected by entomopathogenic nematode. J. Insect Physiol. 2017, 98, 347–355. [Google Scholar] [CrossRef]
- Gautam, U.K.; Bohatá, A.; Shaik, H.A.; Zemek, R.; Kodrík, D. Adipokinetic hormone promotes infection with entomopathogenic fungus Isaria fumosorosea in the cockroach Periplaneta americana. Comp. Biochem. Physiol. C 2020, 229, 108677. [Google Scholar] [CrossRef]
- Gautam, U.K.; Hlávková, D.; Shaik, H.A.; Karaca, I.; Karaca, G.; Sezen, K.; Kodrík, D. Adipokinetic hormones enhance the efficacy of the entomopathogenic fungus Isaria fumosorosea in model and pest insects. Pathogens 2020, 9, 801. [Google Scholar] [CrossRef]
- Kodrík, D.; Bártů, I.; Socha, R. Adipokinetic hormone (Pyrap-AKH) enhances the effect of a pyrethroid insecticide against the firebug Pyrrhocoris apterus. Pest Manag. Sci. 2010, 66, 425–431. [Google Scholar] [CrossRef]
- Betten, D.P.; Richardson, W.H.; Tong, T.C.; Clark, R.F. Massive honey bee envenomation-induced rhabdomyolysis in an adolescent. Pediatrics 2006, 117, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Florea, A.; Craciun, C. Bee (Apis mellifera) venom produced toxic effects of higher amplitude in rat thoracic aorta than in skeletal muscle—An ultrastructural study. Microsc. Microanal. 2012, 18, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Malencik, D.A.; Anderson, S.R. Association of melittin with isolated myosin light-chain. Biochemistry 1988, 27, 1941–1949. [Google Scholar] [CrossRef]
- Ownby, C.L.; Powell, J.R.; Jiang, M.S.; Fletcher, J.E. Melittin and phospholipase A(2) from bee (Apis mellifera) venom cause necrosis of murine skeletal muscle in vivo. Toxicon 1997, 35, 67–80. [Google Scholar] [CrossRef]
- Fletcher, J.E.; Hubert, M.; Wieland, S.J.; Gong, O.H.; Jiang, M.S. Similarities and differences in mechanisms of cardiotoxins, melittin and other myotoxins. Toxicon 1996, 34, 1301–13011. [Google Scholar] [CrossRef]
- Goldsworthy, G.J.; Opoku-Ware, K.; Mullen, L.M. Adipokinetic hormone enhances laminarin and bacterial lipopolysaccharide-induced activation of the prophenoloxidase cascade in the African migratory locust, Locusta migratoria. J. Insect Physiol. 2020, 48, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Goldsworthy, G.J.; Opoku-Ware, K.; Mullen, L.M. Adipokinetic hormone and the immune responses of locusts to infection. Ann. N. Y. Acad. Sci. 2005, 1040, 106–113. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Ann. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [Green Version]
- Bodláková, K.; Beňová, M.; Kodrík, D. The effect of adipokinetic hormones on the activity of digestive enzymes. Physiol. Entomol. 2018, 43, 140–148. [Google Scholar] [CrossRef]
- Havukainen, H.; Münch, D.; Baumann, A.; Zhong, S.; Halskau, Ø.; Krogsgaard, M.; Amdam, G.V. Vitellogenin recognizes cell damage through membrane binding and shields living cells from reactive oxygen species. J. Biol. Chem. 2013, 288, 28369–28381. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.K.; Pakkianathan, B.C.; Kumar, M.; Prasad, T.; Kannan, M.; Konig, S.; Krishnan, M. Vitellogenin from the silkworm, Bombyx mori: An effective anti-bacterial agent. PLoS ONE 2013, 8, e73005. [Google Scholar] [CrossRef] [Green Version]
- Salmela, H.; Sundstrom, L. Vitellogenin in inflammation and immunity in social insects. Inflamm. Cell Signal. 2017, 4, e1506. [Google Scholar]
- Tufail, M.; Hatakeyama, M.; Takeda, M. Molecular evidence for two vitellogenin genes and processing of vitellogenins in the American cockroach, Periplaneta americana. Arch. Insect Biochem. Physiol. 2001, 48, 72–80. [Google Scholar] [CrossRef]
- Carlisle, J.; Loughton, B.G. The inhibition of protein synthesis in Locusta migratoria by adipokinetic hormone. J. Insect Physiol. 1986, 32, 573–578. [Google Scholar] [CrossRef]
- Lorenz, M.W. Adipokinetic hormone inhibits the formation of energy stores and egg production in the cricket Gryllus bimaculatus. Comp. Biochem. Physiol. B 2003, 136, 197–206. [Google Scholar] [CrossRef]
- Pichon, Y.; Manaranche, R. Biochemistry of the nervous system. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; Volume 10, pp. 417–450. [Google Scholar]
- Janner, D.E.; Gomes, N.S.; Poetini, M.R.; Poleto, K.H.; Musachio, E.A.S.; de Almeida, F.P.; Amador, E.C.D.; Reginaldo, J.C.; Ramborger, B.P.; Roehrs, R.; et al. Oxidative stress and decreased dopamine levels induced by imidacloprid exposure cause behavioral changes in a neurodevelopmental disorder model in Drosophila melanogaster. Neurotoxicology 2021, 85, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Weisel-Eichler, A.; Haspel, G.; Libersat, F. Venom of a parasitoid wasp induces prolonged grooming in the cockroach. J. Exp. Biol. 1999, 202, 957–964. [Google Scholar] [CrossRef]
- Večeřa, J.; Krishnan, N.; Alquicer, G.; Kodrík, D.; Socha, R. Adipokinetic hormone-induced enhancement of antioxidant capacity of Pyrrhocoris apterus hemolymph in response to oxidative stress. Comp. Biochem. Physiol. C 2007, 146, 336–342. [Google Scholar] [CrossRef]
- Kodrík, D.; Krishnan, N.; Habuštová, O. Is the titer of adipokinetic peptides in Leptinotarsa decemlineata fed on genetically modified potatoes increased by oxidative stress? Peptides 2007, 28, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Koubová, J.; Sábová, M.; Brejcha, M.; Kodrík, D.; Čapková Frydrychová, R. Seasonal changes in telomerase activity in relation to cell size, DNA replication, and nutrient contents in the fat body of Apis mellifera. Sci. Rep. 2021, 11, 592. [Google Scholar] [CrossRef] [PubMed]
- Kodrík, D.; Krištůfek, V.; Svobodová, Z. Bee year: Basic physiological strategies to cope with seasonality. Comp. Biochem. Physiol. A 2022, 264, 111115. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.J.B. Determining the hemolymph volume of the cockroach. In Tested Studies for Laboratory Teaching, Proceedings of the 15th Workshop/Conference of the Association for Biology Laboratory Education, Atlanta, GA, USA, 7–11 June 1994; Goldman, C.A., Ed.; Association for Biology Laboratory Education (ABLE): Toronto, ON, Canada, 1994; pp. 119–139. [Google Scholar]
- Karbusová, N.; Gautam, U.K.; Kodrík, D. Effect of natural toxins and adipokinetic hormones on the activity of digestive enzymes in the midgut of the cockroach Periplaneta americana. Arch. Insect Biochem. Physiol. 2019, 101, e21586. [Google Scholar] [CrossRef]
- Stoscheck, C.M. Quantitation of protein. In Methods in Enzymology; Deutscher, M.P., Ed.; Academic Press: San Diego, CA, USA, 1990; Volume 182, pp. 50–68. [Google Scholar]
- Zöllner, N.; Kirsch, K. Über die quantitative Bestimmung von Lipoide (Mikromethode) mittels der vielen natürlichen Lipoiden (allen bekannten Plasmalipoiden) gemeinsamen Sulfo-phospho-vanillin-Reaktion. Z. Ges. Exp. Medizin. 1962, 135, 545–561. [Google Scholar] [CrossRef]
- Kodrík, D.; Socha, R.; Šimek, P.; Zemek, R.; Goldsworthy, G.J. A new member of the AKH/RPCH family that stimulates locomotory activity in the firebug, Pyrrhocoris apterus (Heteroptera). Insect Biochem. Mol. Biol. 2000, 30, 489–498. [Google Scholar] [CrossRef]
- Carroll, N.V.; Longley, R.W.; Roe, J.H. The determination of glycogen in liver and muscle by use of anthrone reagent. J. Biol. Chem. 1956, 220, 583–593. [Google Scholar] [CrossRef]
- Socha, R.; Kodrík, D.; Šimek, P.; Patočková, M. The kind of AKH-mobilized energy substrates in insects can be predicted without a knowledge of the hormone structure. Eur. J. Entomol. 2004, 101, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Bernfeld, P. Amylases, α and β. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; Volume 1, pp. 149–158. [Google Scholar]
- Roberts, I.M. Hydrolysis of 4-methylumbelliferyl butyrate: A convenient and sensitive fluorescent assay for lipase activity. Lipids 1985, 20, 243–247. [Google Scholar] [CrossRef]
- Elpidina, E.N.; Vinokurov, K.S.; Gromenko, V.A.; Rudenskaya, Y.A.; Dunaevsky, Y.E.; Zhuzhikov, D.P. Compartmentalization of proteinases and amylases in Nauphoeta cinerea midgut. Arch. Insect Biochem. Physiol. 2001, 48, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Socha, R.; Šula, J.; Kodrík, D.; Gelbič, I. Hormonal control of vitellogenin synthesis in Pyrrhocoris apterus. J. Insect Physiol. 1991, 37, 805–816. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodláková, K.; Černý, J.; Štěrbová, H.; Guráň, R.; Zítka, O.; Kodrík, D. Insect Body Defence Reactions against Bee Venom: Do Adipokinetic Hormones Play a Role? Toxins 2022, 14, 11. https://doi.org/10.3390/toxins14010011
Bodláková K, Černý J, Štěrbová H, Guráň R, Zítka O, Kodrík D. Insect Body Defence Reactions against Bee Venom: Do Adipokinetic Hormones Play a Role? Toxins. 2022; 14(1):11. https://doi.org/10.3390/toxins14010011
Chicago/Turabian StyleBodláková, Karolina, Jan Černý, Helena Štěrbová, Roman Guráň, Ondřej Zítka, and Dalibor Kodrík. 2022. "Insect Body Defence Reactions against Bee Venom: Do Adipokinetic Hormones Play a Role?" Toxins 14, no. 1: 11. https://doi.org/10.3390/toxins14010011
APA StyleBodláková, K., Černý, J., Štěrbová, H., Guráň, R., Zítka, O., & Kodrík, D. (2022). Insect Body Defence Reactions against Bee Venom: Do Adipokinetic Hormones Play a Role? Toxins, 14(1), 11. https://doi.org/10.3390/toxins14010011