Mycotoxin Interactions along the Gastrointestinal Tract: In Vitro Semi-Dynamic Digestion and Static Colonic Fermentation of a Contaminated Meal

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Mycotoxins’ Analytical Method Performance
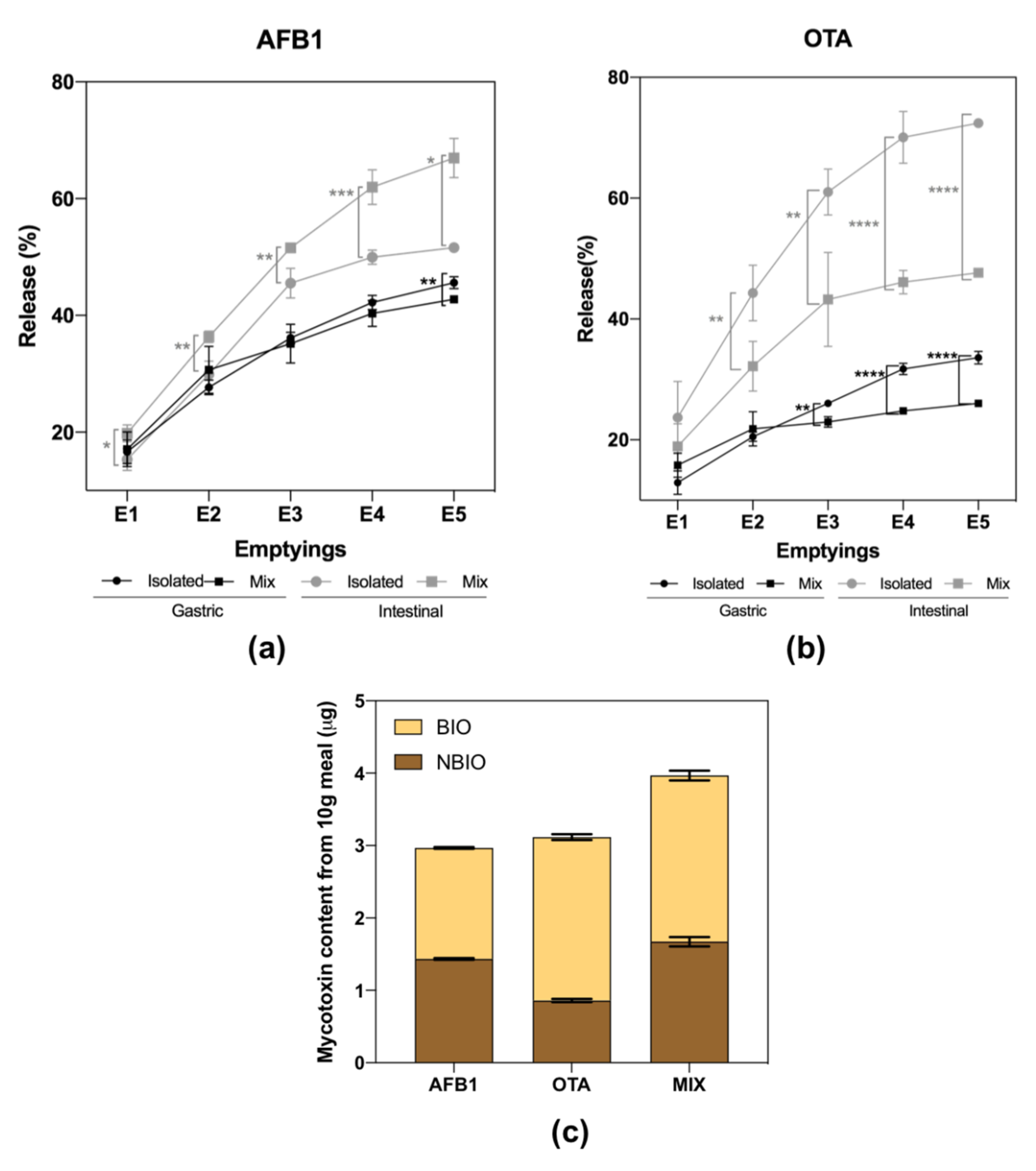
2.2. Mycotoxins Bioaccessibility during Gastrointestinal Digestion
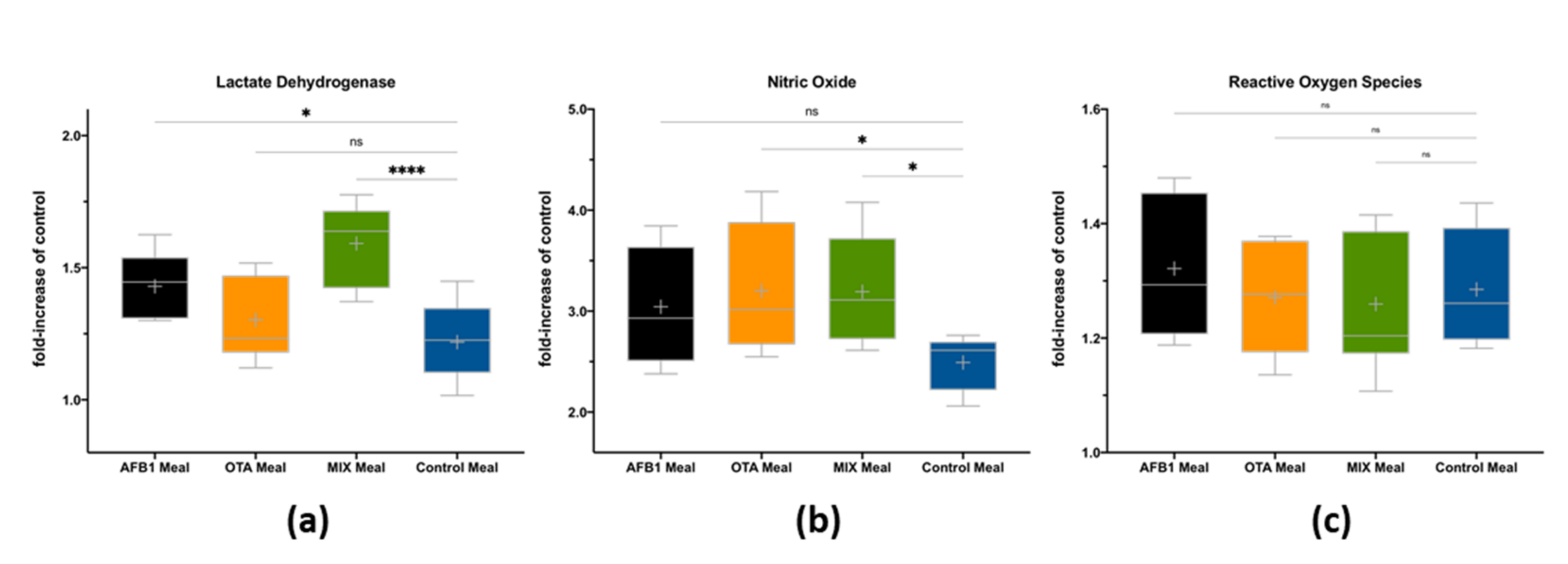
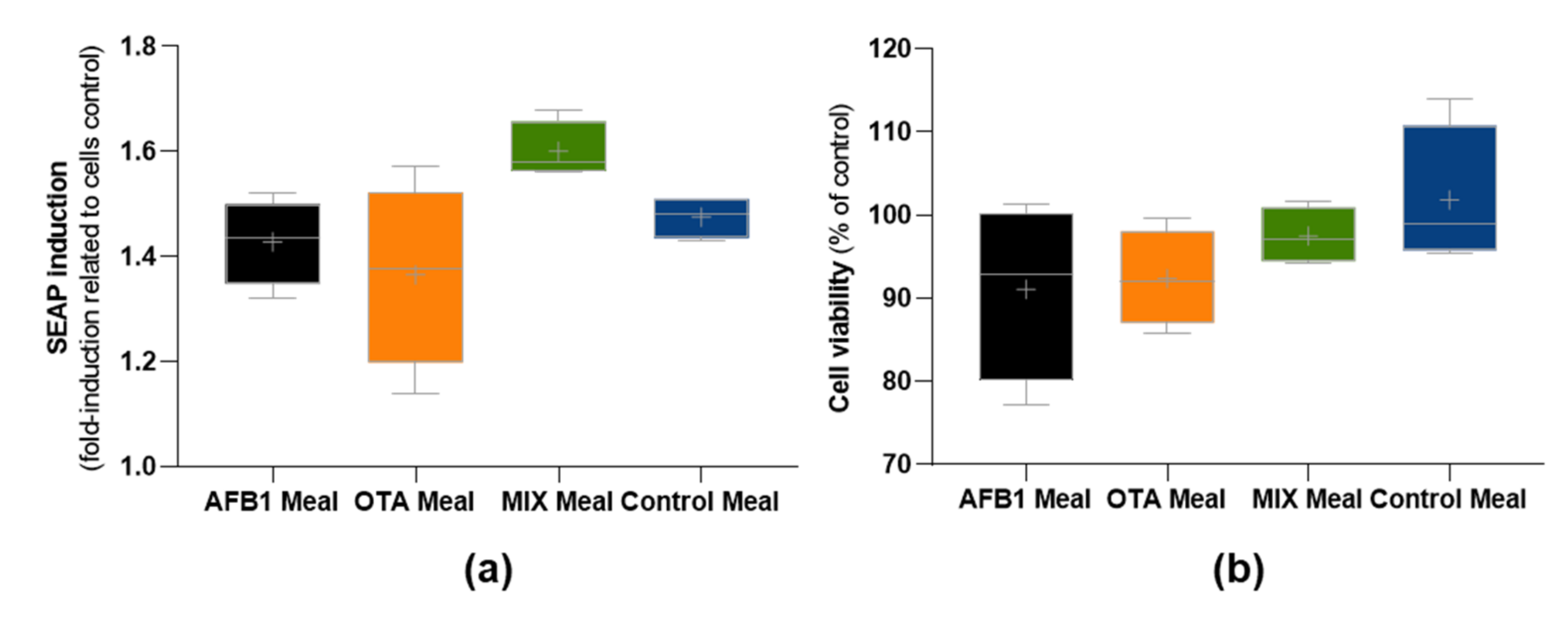
2.3. Impact of Bioaccessible Mycotoxins on Intestinal Cells’ Viability and Inflammation
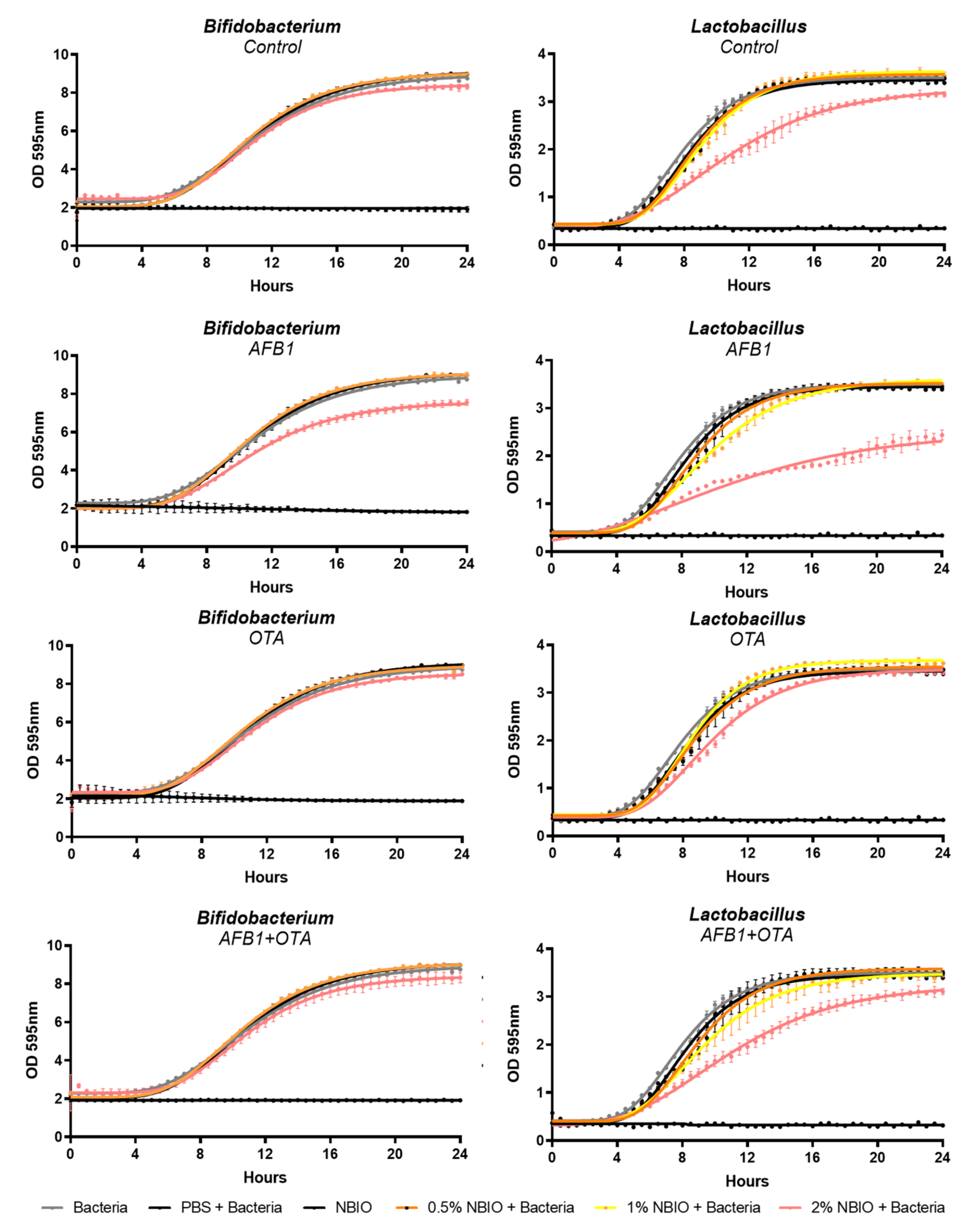
2.4. Impact of Non-Bioaccessible Mycotoxins on Probiotic Lactobacillus casei and Bifidobacterium lactis
- (i)
- Strong negative Pearson correlations between the AFB1 content in the meal and all growth parameters of both L. casei and B. lactis, suggesting a negative impact on bacterial growth (i.e., reduction in the MOD, growth rate, and lag time). Even stronger negative correlations were observed between the non-bioaccessible AFB1 content and probiotic growth parameters, indicating an inhibition of growth in both strains by AFB1.
- (ii)
- Strong positive correlations between the OTA content in the meal and L. casei growth parameters and the B. lactis MOD. Moderate correlations were observed between the non-bioacessible OTA content and L. casei growth, confirming an enhancement of the growth rate.
- (iii)
- No correlation was found between bacterial growth and mycotoxin content when summing up their contents (AFB1 + OTA), suggesting a null effect in the co-exposure situation.
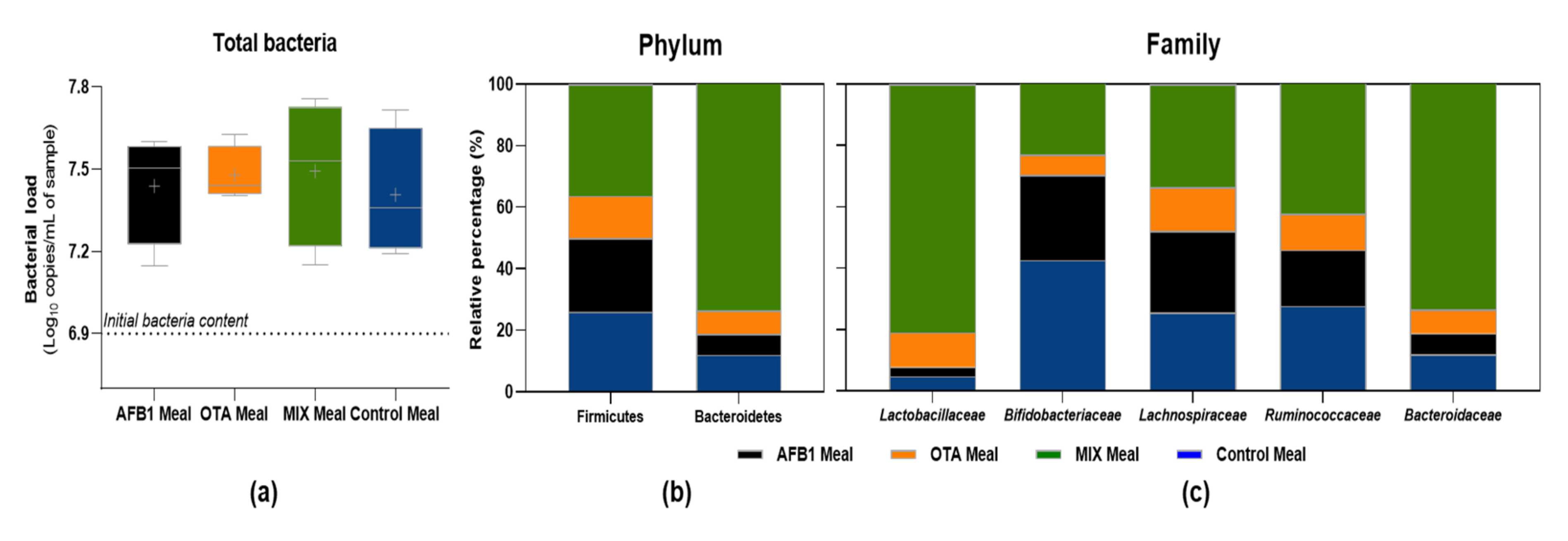
2.5. Impact of Non-Bioaccessible Mycotoxins on the Gut Microbiota, Intestinal Cells’ Viability, and Inflammation
3. Discussion
4. Materials and Methods
4.1. Sample Preparation and Spiking
4.2. In Vitro Upper Gastrointestinal Digestion
4.3. Mycotoxin Analysis
4.4. Impact of Bioaccessible Mycotoxins on Caco-2 Cells’ Viability and on Inflammation
4.5. Effect of Non-Bioaccessible Mycotoxins on Probiotics Lactobacillus casei and Bifidobacterium lactis Growth
4.6. In Vitro Colonic Fermentation
4.7. Total Bacteria Quantification by qPCR and 16S rRNA Amplicon Sequencing
4.8. Impact of Non-Bioaccessible Mycotoxins on HT29 NF-kB Reporter Cells’ Viability and on Inflammation
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens—The IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Becker-Algeri, T.A.; Castagnaro, D.; De Bortoli, K.; De Souza, C.; Drunkler, D.A.; Badiale-Furlong, E. Mycotoxins in Bovine Milk and Dairy Products: A Review. J. Food Sci. 2016, 81, R544–R552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkerroum, N. Mycotoxins in dairy products: A review. Int. Dairy J. 2016, 62, 63–75. [Google Scholar] [CrossRef]
- Sarmast, E.; Fallah, A.A.; Jafari, T.; Khaneghah, A.M. Occurrence and fate of mycotoxins in cereals and cereal-based products: A narrative review of systematic reviews and meta-analyses studies. Curr. Opin. Food Sci. 2021, 39, 68–75. [Google Scholar] [CrossRef]
- Assunção, R.; Martins, C.; Dupont, D.; Alvito, P. Patulin and ochratoxin A co-occurrence and their bioaccessibility in processed cereal-based foods: A contribution for Portuguese children risk assessment. Food Chem. Toxicol. 2016, 96, 205–214. [Google Scholar] [CrossRef]
- Ibáñez-Vea, M.; González-Peñas, E.; Lizarraga, E.; de Cerain, A.L. Co-occurrence of aflatoxins, ochratoxin A and zearalenone in barley from a northern region of Spain. Food Chem. 2012, 132, 35–42. [Google Scholar] [CrossRef]
- Martins, C.; Assunção, R.; Cunha, S.C.; Fernandes, J.O.; Jager, A.; Petta, T.; Oliveira, C.A.; Alvito, P. Assessment of multiple mycotoxins in breakfast cereals available in the Portuguese market. Food Chem. 2018, 239, 132–140. [Google Scholar] [CrossRef]
- De Ruyck, K.; Huybrechts, I.; Yang, S.; Arcella, D.; Claeys, L.; Abbeddou, S.; De Keyzer, W.; De Vries, J.; Ocke, M.; Ruprich, J.; et al. Mycotoxin exposure assessments in a multi-center European validation study by 24-hour dietary recall and biological fluid sampling. Environ. Int. 2020, 137, 105539. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Mulet-Cabero, A.-I.; Rigby, N.M.; Brodkorb, A.; Mackie, A.R. Dairy food structures influence the rates of nutrient digestion through different in vitro gastric behaviour. Food Hydrocoll. 2017, 67, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.; Manyes, L.; Mañes, J.; Meca, G. Influence of prebiotics, probiotics and protein ingredients on mycotoxin bioaccessibility. Food Funct. 2015, 6, 987–994. [Google Scholar] [CrossRef]
- Sobral, M.M.C.; Cunha, S.C.; Faria, M.A.; Martins, Z.; Ferreira, I.M. Influence of oven and microwave cooking with the addition of herbs on the exposure to multi-mycotoxins from chicken breast muscle. Food Chem. 2019, 276, 274–284. [Google Scholar] [CrossRef]
- Tolosa, J.; Font, G.; Mañes, J.; Ferrer, E. Mitigation of enniatins in edible fish tissues by thermal processes and identification of degradation products. Food Chem. Toxicol. 2017, 101, 67–74. [Google Scholar] [CrossRef]
- Faria, M.; Melo, A.; Ferreira, I.I. Influence of dietary patterns on contaminants bioaccessibility and intestinal transport by in vitro assays. Food Res. Int. 2020, 137, 109358. [Google Scholar] [CrossRef]
- Versantvoort, C.H.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.; Sips, A.J. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef]
- Ren, Z.; Guo, C.; Yu, S.; Zhu, L.; Wang, Y.; Hu, H.; Deng, J. Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. Int. J. Mol. Sci. 2019, 20, 2777. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.; Priest, E.; Naglik, J.R.; Richardson, J.P. Fungal Toxins and Host Immune Responses. Front. Microbiol. 2021, 12, 643639. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef]
- Claus, S.P.; Guillou, H.; Ellero-Simatos, S. The gut microbiota: A major player in the toxicity of environmental pollutants? NPJ Biofilms Microbiomes 2016, 2, 16003. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rodrigues, C.; Stojanović-Radić, Z.; Dimitrijević, M.; Aleksić, A.; Neffe-Skocińska, K.; Zielińska, D.; Kołożyn-Krajewska, D.; Salehi, B.; Prabu, S.M.; et al. Probiotics: Versatile Bioactive Components in Promoting Human Health. Medicina 2020, 56, 433. [Google Scholar] [CrossRef]
- Guerre, P. Mycotoxin and Gut Microbiota Interactions. Toxins 2020, 12, 769. [Google Scholar] [CrossRef] [PubMed]
- Izco, M.; Vettorazzi, A.; de Toro, M.; Sáenz, Y.; Alvarez-Erviti, L. Oral Sub-chronic Ochratoxin A Exposure Induces Gut Microbiota Alterations in Mice. Toxins 2021, 13, 106. [Google Scholar] [CrossRef]
- Liew, W.-P.-P.; Mohd-Redzwan, S.; Than, L.T.L. Gut Microbiota Profiling of Aflatoxin B1-Induced Rats Treated with Lactobacillus casei Shirota. Toxins 2019, 11, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tang, L.; Glenn, T.C.; Wang, J.S. Aflatoxin B1 Induced Compositional Changes in Gut Microbial Communities of Male F344 Rats. Toxicol. Sci. 2016, 150, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Angelis, E.; Monaci, L.; Mackie, A.R.; Salt, L.J. Bioaccessibility of T-2 and HT-2 toxins in mycotoxin contaminated bread models submitted to in vitro human digestion. Innov. Food Sci. Emerg. Technol. 2013, 25, 88–96. [Google Scholar] [CrossRef]
- González-Arias, C.A.; Marín, S.; Sanchis, V.; Ramos, A.J. Mycotoxin bioaccessibility/absorption assessment using in vitro digestion models: A review. World Mycotoxin J. 2013, 6, 167–184. [Google Scholar] [CrossRef] [Green Version]
- Kabak, B.; Brandon, E.F.; Var, I.; Blokland, M.; Sips, A.J. Effects of probiotic bacteria on the bioaccessibility of aflatoxin B1 and ochratoxin A using an in vitro digestion model under fed conditions. J. Environ. Sci. Health Part B 2009, 44, 472–480. [Google Scholar] [CrossRef]
- SANTE. Method Validation and Analytical Quality Control Procedures for Pesticide Residues Analysis in Food & Feed; SANTE/12682/2019; European Commission: Brussels, Belgium, 2019. [Google Scholar]
- Raters, M.; Matissek, R. Thermal stability of aflatoxin B1 and ochratoxin A. Mycotoxin Res. 2008, 24, 130–134. [Google Scholar] [CrossRef]
- Broom, L. Mycotoxins and the intestine. Anim. Nutr. 2015, 1, 262–265. [Google Scholar] [CrossRef]
- Carvajal-Moreno, M. Do Gastroenterologists Consider Aflatoxins as Origin of Digestive System Cancers? J. Pharmacovigil. 2017, 5, 242. [Google Scholar] [CrossRef] [Green Version]
- Romero, A.; Ares, I.; Ramos, E.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.-R.; Anadón, A.; Martínez, M.-A. Mycotoxins modify the barrier function of Caco-2 cells through differential gene expression of specific claudin isoforms: Protective effect of illite mineral clay. Toxicology 2016, 353, 21–33. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, N.; Liu, J.; Li, F.; Li, S.; Wang, J. Aflatoxin B1 and aflatoxin M1 induced cytotoxicity and DNA damage in differentiated and undifferentiated Caco-2 cells. Food Chem. Toxicol. 2015, 83, 54–60. [Google Scholar] [CrossRef]
- Liew, W.-P.-P.; Mohd-Redzwan, S. Mycotoxin: Its Impact on Gut Health and Microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Grosu, I.A.; Pistol, G.C.; Taranu, I.; Marin, D. The impact of dietary grape seed meal on healthy and aflatoxin B1 afflicted microbiota of pigs after weaning. Toxins 2019, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, A.; Weese, J.; Bracarense, A.; Alfieri, A.; Oliveira, G.; Kawamura, O.; Hirooka, E.; Itano, E.; Costa, M. Single aflatoxin B1 exposure induces changes in gut microbiota community in C57Bl/6 mice. World Mycotoxin J. 2017, 10, 249–254. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, K.I.; Muroi, M.; Kinoshita, M.; Hamada, O.; Minai, Y.; Sugita-Konishi, Y.; Kamata, Y.; Tanamoto, K.I. NF-κB activation via MyD88-dependent Toll-like receptor signaling is inhibited by trichothecene mycotoxin deoxynivalenol. J. Toxicol. Sci. 2016, 41, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.T.; Lo, Y.C.; Wu, M.S.; Li, C.Y.; Kuo, Y.P.; Lai, Y.H.; Tsai, Y.; Chen, K.C.; Chuang, T.H.; Yao, C.H.; et al. Mycotoxin Patulin Suppresses Innate Immune Responses by Mitochondrial Dysfunction and p62/Sequestosome-1-dependent Mitophagy. J. Biol. Chem. 2016, 291, 19299–19311. [Google Scholar] [CrossRef] [Green Version]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- 2002/657/EC: Commission Decision of 12 August 2002 implementing Council Directive 96/23/EC concerning the performance of analytical methods and the interpretation of results. Off. J. Eur. Communities 2002, 221, 8–36.
- ICH. ICH Harmonized Tripartite Guideline: Validation of Analytical Procedures: Text and Methodology Q2(R1). 2005; Available online: (accessed on 27 November 2021). [Google Scholar]
- Pinho, S.C.; Faria, M.A.; Melo, A.; Pinto, E.; Almeida, A.; Alves, R.; Cabrita, A.R.; Fonseca, A.J.; Ferreira, I.M. Effect of skimmed milk on intestinal tract: Prevention of increased reactive oxygen species and nitric oxide formation. Int. Dairy J. 2021, 118, 105046. [Google Scholar] [CrossRef]
- Kaja, S.; Payne, A.J.; Singh, T.; Ghuman, J.K.; Sieck, E.G.; Koulen, P. An optimized lactate dehydrogenase release assay for screening of drug candidates in neuroscience. J. Pharmacol. Toxicol. Methods 2015, 73, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, M.; Kalam, A.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcántara, C.; Žugčić, T.; Abdelkebir, R.; García-Pérez, J.V.; Jambrak, A.R.; Lorenzo, J.M.; Collado, M.C.; Granato, D.; Barba, F.J. Effects of Ultrasound-Assisted Extraction and Solvent on the Phenolic Profile, Bacterial Growth, and Anti-Inflammatory/Antioxidant Activities of Mediterranean Olive and Fig Leaves Extracts. Molecules 2020, 25, 1718. [Google Scholar] [CrossRef] [Green Version]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; Van’T Riet, K. Modeling of the Bacterial Growth Curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [Green Version]
- Hevia, A.; Delgado, S.; Margolles, A.; Sánchez, B. Application of density gradient for the isolation of the fecal microbial stool component and the potential use thereof. Sci. Rep. 2015, 5, 16807. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Del-Campo, A.; Alcántara, C.; Collado, M.C.; Rodríguez-Díaz, J.; Yebra, M.J. Human milk and mucosa-associated disaccharides impact on cultured infant fecal microbiota. Sci. Rep. 2020, 10, 11845. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516. [Google Scholar] [CrossRef] [Green Version]
- Cruaud, P.; Vigneron, A.; Lucchetti-Miganeh, C.; Ciron, P.E.; Godfroy, A.; Cambon-Bonavita, M.A. Influence of DNA Extraction Method, 16S rRNA Targeted Hypervariable Regions, and Sample Origin on Microbial Diversity Detected by 454 Pyrosequencing in Marine Chemosynthetic Ecosystems. Appl. Environ. Microbiol. 2014, 80, 4626–4639. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- FLASh: Fast Length Adjustments of Short Reads. Available online: http://ccb.jhu.edu/software/FLASH/ (accessed on 27 December 2021).
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- SILVA High Quality Ribossomal RNA Databases. Available online: http://www.arb-silva.de (accessed on 27 December 2021).
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linearity | LOD (ng) | LOQ (ng) | ||||
|---|---|---|---|---|---|---|
| Range (ng) | Regression Equation | R2 | ||||
| AFB1 | Meal | 2–40 | y = 0.0428x − 0.111 | 0.9978 | 2.2 | 7.3 |
| Gastric digest | 1–20 | y = 0.0479x + 0.0789 | 0.9963 | |||
| Intestinal digest | 1–20 | y = 0.0697x + 0.145 | 0.9939 | |||
| OTA | Meal | 2–40 | y = 0.314x − 1.01 | 0.9931 | 0.76 | 2.52 |
| Gastric digest | 1–20 | y = 0.655x + 0.143 | 0.9947 | |||
| Intestinal digest | 1–20 | y = 0.328x − 0.106 | 0.9993 | |||
| Spiked Level (ng/mL) | Recovery (Interday, % RSD) | Intraday (% RSD) | |
|---|---|---|---|
| AFB1 | 17 | 92 (9) | 5 |
| 34 | 105 (3) | 8 | |
| 85 | 102 (10) | 9 | |
| OTA | 11 | 86 (25) | 14 |
| 22 | 80 (22) | 19 | |
| 55 | 69 (15) | 28 |
| AFB1 Meal | OTA Meal | AFB1 BIO | OTA BIO | Sum BIO AFB1 + OTA | NO | ROS | LDH | |
|---|---|---|---|---|---|---|---|---|
| AFB1 meal | 1 | |||||||
| OTA meal | −0.317 | 1 | ||||||
| AFB1 BIO | 0.977 | −0.264 | 1 | |||||
| OTA BIO | −0.431 | 0.946 | −0.390 | 1 | ||||
| Sum BIO AFB1 + OTA | 0.345 | 0.731 | 0.404 | 0.685 | 1 | |||
| NO | 0.298 | 0.535 | 0.240 | 0.338 | 0.525 | 1 | ||
| ROS | 0.179 | −0.045 | 0.040 | −0.177 | −0.144 | 0.692 | 1 | |
| LDH | 0.640 | 0.187 | 0.637 | −0.059 | 0.446 | 0.793 | 0.579 | 1 |
| Lactobacillus casei | |||||
|---|---|---|---|---|---|
| NBIO | AFB1 | OTA | MIX | Control | |
| 0.5% | MOD | 3.128 ± 0.036 | 3.114 ± 0.034 | 3.176 ± 0.040 | 3.132 ± 0.023 |
| Growth rate (μmax) (h−1) | 0.4502 ± 0.014 | 0.4574 ± 0.014 | 0.453 ± 0.015 | 0.4735 ± 0.010 | |
| Lag time (h) | 5.29 ± 0.137 | 5.228 ± 0.131 | 5.326 ± 0.149 | 5.298 ± 0.088 | |
| 1% | MOD | 3.23 ± 0.038 | 3.235 ± 0.024 | 3.122 ± 0.052 * | 3.202 ± 0.032 |
| Growth rate (μmax) (h−1) | 0.3527 ± 0.009 **** | 0.5211 ± 0.011 *** | 0.3659 ± 0.014 **** | 0.4555 ± 0.012 | |
| Lag time (h) | 4.92 ± 0.147 ** | 5.44 ± 0.084 | 4.863 ± 0.210 ** | 5.39 ± 0.121 | |
| 2% | MOD | 2.364 ± 0.169 *** | 3.104 ± 0.026 ** | 2.946 ± 0.061 | 2.924 ± 0.059 |
| Growth rate (μmax) (h−1) | 0.131 ± 0.006 **** | 0.3696 ± 0.007 **** | 0.228 ± 0.006 | 0.24 ± 0.007 | |
| Lag time (h) | 1.141 ± 1.049 *** | 5.383 ± 0.103 ** | 4.308 ± 0.258 | 4.398 ± 0.256 | |
| Bifidobacterium lactis | |||||
| NBIO | AFB1 | OTA | MIX | Control | |
| 0.5% | MOD | 7.078 ± 0.025 * | 6.788 ± 0.041 *** | 7.013 ± 0.030 | 7.014 ± 0.026 |
| Growth rate (μmax) (h−1) | 0.7646 ± 0.006 | 0.7319 ± 0.009 | 0.7569 ± 0.007 | 0.764 ± 0.006 | |
| Lag time (h) | 5.858 ± 0.044 | 5.808 ± 0.074 | 5.844 ± 0.053 | 5.88 ± 0.046 | |
| 1% | MOD | 7.078 ± 0.025 * | 6.788 ± 0.041 **** | 7.013 ± 0.030 | 7.014 ± 0.026 |
| Growth rate (μmax) (h−1) | 0.7646 ± 0.006 | 0.7319 ± 0.009 ** | 0.7569 ± 0.007 | 0.764 ± 0.006 | |
| Lag time (h) | 5.858 ± 0.044 | 5.808 ± 0.074 | 5.844 ± 0.053 | 5.88 ± 0.046 | |
| 2% | MOD | 5.585 ± 0.042 **** | 6.224 ± 0.058 *** | 6.095 ± 0.087 | 5.978 ± 0.050 |
| Growth rate (μmax) (h−1) | 0.5585 ± 0.008 **** | 0.6715 ± 0.013 | 0.6742 ± 0.021 | 0.6902 ± 0.013 | |
| Lag time (h) | 5.689 ± 0.092 **** | 6.281 ± 0.115 * | 6.289 ± 0.173 | 6.515 ± 0.101 | |
| AFB1 Meal | OTA Meal | AFB1 NBIO | OTA NBIO | Sum NBIO AFB1 + OTA | L. casei | B. lactis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MOD | Growth Rate | Lag Time | MOD | Growth Rate | Lag Time | |||||||
| AFB1 meal | 1 | |||||||||||
| OTA meal | −0.309 | 1 | ||||||||||
| AFB1 NBIO | 0.954 | −0.424 | 1 | |||||||||
| OTA NBIO | 0.044 | 0.739 | −0.181 | 1 | ||||||||
| Sum NBIO AFB1 + OTA | 0.772 | 0.256 | 0.629 | 0.65 | 1 | |||||||
| L. casei | MOD | −0.765 | 0.641 | −0.874 | 0.556 | −0.236 | 1 | |||||
| Growth rate | −0.8 | 0.786 | −0.839 | 0.437 | −0.304 | 0.871 | 1 | |||||
| Lag time | −0.792 | 0.63 | −0.889 | 0.516 | −0.279 | 0.998 | 0.878 | 1 | ||||
| B. lactis | MOD | −0.686 | 0.733 | −0.817 | 0.67 | −0.102 | 0.969 | 0.896 | 0.957 | 1 | ||
| Growth rate | −0.753 | 0.373 | −0.883 | 0.431 | −0.342 | 0.919 | 0.695 | 0.912 | 0.883 | 1 | ||
| Lag time | −0.772 | 0.216 | −0.868 | 0.271 | −0.457 | 0.863 | 0.614 | 0.86 | 0.798 | 0.981 | 1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobral, M.M.C.; Gonçalves, T.; Martins, Z.E.; Bäuerl, C.; Cortés-Macías, E.; Collado, M.C.; Ferreira, I.M.P.L.V.O. Mycotoxin Interactions along the Gastrointestinal Tract: In Vitro Semi-Dynamic Digestion and Static Colonic Fermentation of a Contaminated Meal. Toxins 2022, 14, 28. https://doi.org/10.3390/toxins14010028
Sobral MMC, Gonçalves T, Martins ZE, Bäuerl C, Cortés-Macías E, Collado MC, Ferreira IMPLVO. Mycotoxin Interactions along the Gastrointestinal Tract: In Vitro Semi-Dynamic Digestion and Static Colonic Fermentation of a Contaminated Meal. Toxins. 2022; 14(1):28. https://doi.org/10.3390/toxins14010028
Chicago/Turabian StyleSobral, Maria Madalena Costa, Tiago Gonçalves, Zita E. Martins, Christine Bäuerl, Erika Cortés-Macías, Maria Carmen Collado, and Isabel M. P. L. V. O. Ferreira. 2022. "Mycotoxin Interactions along the Gastrointestinal Tract: In Vitro Semi-Dynamic Digestion and Static Colonic Fermentation of a Contaminated Meal" Toxins 14, no. 1: 28. https://doi.org/10.3390/toxins14010028
APA StyleSobral, M. M. C., Gonçalves, T., Martins, Z. E., Bäuerl, C., Cortés-Macías, E., Collado, M. C., & Ferreira, I. M. P. L. V. O. (2022). Mycotoxin Interactions along the Gastrointestinal Tract: In Vitro Semi-Dynamic Digestion and Static Colonic Fermentation of a Contaminated Meal. Toxins, 14(1), 28. https://doi.org/10.3390/toxins14010028