Survey of Freshly Harvested Oat Grains from Southern Brazil Reveals High Incidence of Type B Trichothecenes and Associated Fusarium Species

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Water Activity and Mycobiota of Freshly Harvested Oat Grains
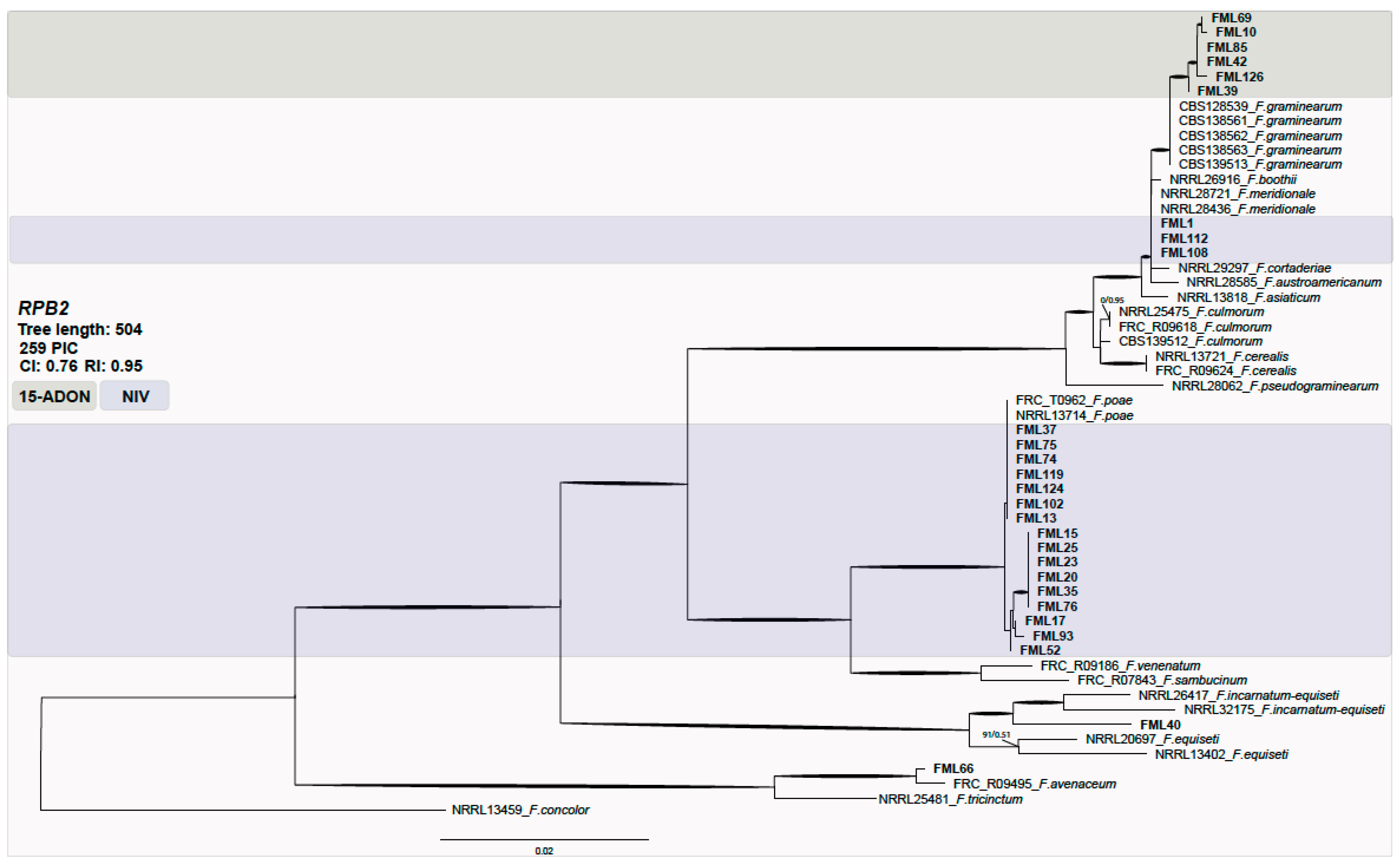
2.2. Molecular Characterization of Fusarium Isolates
2.3. Mycotoxin Analysis
Occurrence of Type B Trichothecenes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Oat Samples
5.2. Water Activity and Identification of Mycobiota
5.3. Characterization of Fusarium Isolates
5.4. DNA Extraction, PCR, and Sequencing Analyses of the RPB2 Gene
5.5. Phylogenetic Analysis
5.6. Mycotoxin Analysis
5.6.1. Mycotoxin Extraction
5.6.2. Chromatography Conditions
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaur, S.; Bhardwaj, R.D.; Kapoor, R.; Grewal, S.K. Biochemical characterization of oat (Avena sativa L.) genotypes with high nutritional potential. LWT 2019, 110, 32–39. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA). Grain: World Markets and Trade. Foreign Agricultural Service/USDA, Global Market Analysis. Available online: https://apps.fas.usda.gov/psdonline/circulars/grain.pdf (accessed on 1 March 2021).
- Companhia Nacional de Abastecimento (CONAB). Acompanhamento da Safra Brasileira, Grãos, Safra 2021/22, 2° levantamento. Conab Reports 2021, 9. Available online: https://www.conab.gov.br/info-agro/safras/graos/boletim-da-safra-de-graos (accessed on 23 November 2021).
- Liu, L.; Ma, M.; Liu, Z.; Zhang, L.; Zhou, J. Community structure of fungal pathogens causing spikelet rot disease of naked oat from different ecological regions of China. Sci. Rep. 2021, 11, 1243. [Google Scholar] [CrossRef]
- Tamburic-Ilincic, L. Fusarium species and mycotoxins associated with oat in southwestern Ontario, Canada. Can. J. Plant Sci. 2010, 90, 211–216. [Google Scholar] [CrossRef]
- Hautsalo, J.; Jalli, M.; Manninen, O.; Veteläinen, M. Evaluation of resistance to Fusarium graminearum in oats. Euphytica 2018, 214, 139. [Google Scholar] [CrossRef]
- Schöneberg, T.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Seifert, K.; Gräfenhan, T.; Keller, B.; et al. Occurrence of Fusarium species and mycotoxins in Swiss oats—Impact of cropping factors. Eur. J. Agron. 2018, 92, 123–132. [Google Scholar] [CrossRef]
- Ghimire, B.; Sapkota, S.; Bahri, B.A.; Martinez-Espinoza, A.D.; Buck, J.W.; Mergoum, M. Fusarium Head Blight and rust diseases in soft red winter wheat in the Southeast United States: State of the art, challenges and future perspective for breeding. Front. Plant Sci. 2020, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Mylona, K.; Magan, N. Fusarium langsethiae: Storage environment influences dry matter losses and T2 and HT-2 toxin contamination of oats. J. Stored Prod. Res. 2011, 47, 321–327. [Google Scholar] [CrossRef]
- O’Donnel, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.N.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef]
- Pereira, C.S.; Cunha, S.C.; Fernandes, J.O. Prevalent mycotoxins in animal feed: Occurrence and analytical methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA). Scientific opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar] [CrossRef]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. Toxicity of deoxynivalenol and its acetylated derivatives on the intestine: Differential effects on morphology, barrier function, tight junction proteins, and mitogen-activated protein kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Safety Evaluation of Certain Contaminants in Food; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Evaluation of Certain Food Additives and Contaminants; Fifty-Third Report of the Joint FAO/WHO Expert Committee on Food Additives; World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- Tekauz, A.; Mitchell Fetch, J.W.; Rossnagel, B.G.; Savard, M.E. Progress in assessing the impact of Fusarium head blight on oat in Western Canada and screening of avena germplasm for resistance. Cereal Res. Commun. 2008, 36, 49–56. [Google Scholar] [CrossRef]
- Yli-Mattila, T. Ecology and evolution of toxigenic Fusarium species in cereals in Northern Europe and Asia. J. Plant Pathol. 2010, 92, 7–18. [Google Scholar]
- Schöneberg, T.; Kibler, K.; Wettstein, F.E.; Bucheli, T.D.; Forrer, H.R.; Musa, T.; Mascher, F.; Bertossa, M.; Keller, B.; Vogelgsang, S. Influence of temperature, humidity duration and growth stage on the infection and mycotoxin production by Fusarium langsethiae and Fusarium poae in oats. Plant Pathol. 2019, 68, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Kokkonen, M.; Jestoi, M.; Laitila, A. Mycotoxin production of Fusarium langsethiae and Fusarium sporotrichioides on cereal-based substrates. Mycotoxin Res. 2012, 28, 25–35. [Google Scholar] [CrossRef]
- Stenglein, S.A.; Dinolfo, M.I.; Barros, G.; Bongiorno, F.; Chulze, S.N.; Moreno, M.V. Fusarium poae pathogenicity and mycotoxin accumulation on selected wheat and barley genotypes at a single location in Argentina. Plant Dis. 2014, 98, 1733–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munkvold, G.P. Fusarium species and their associated mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Human Press: Totowa, NJ, USA, 2017; pp. 51–106. [Google Scholar]
- Langseth, W.; Rundberget, T. The occurrence of HT-2 toxin and other trichothecenes in Norwegian cereals. Mycopathologia 1999, 147, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Hietaniemi, V.; Kontturi, M.; Rämö, S.; Eurola, M.; Kangas, A.; Niskanen, M.; Saastamoinen, M. Contents of trichothecenes in oats during official variety, organic cultivation and nitrogen fertilization trials in Finland. Agr. Food Sci. 2004, 13, 54–67. [Google Scholar] [CrossRef]
- Gottschalk, C.; Barthel, J.; Engelhardt, G.; Bauer, J.; Meyes, K. Occurrence of type A trichothecenes in conventionally and organically produced oats and oat products. Mol. Nutr. Food Res. 2007, 51, 1547–1553. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jørgensen, L.N. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef] [Green Version]
- Polišenská, I.; Jirsa, O.; Vaculová, K.; Pospíchalová, M.; Wawroszova, S.; Frydrych, J. Fusarium mycotoxins in two hulles oat and barley cultivars used for food purposes. Foods 2020, 9, 1037. [Google Scholar] [CrossRef] [PubMed]
- Rupollo, G.; Gutkoski, L.C.; Martins, I.R.; Elias, M.C. Efeito da umidade e do período de armazenamento hermético na contaminação natural por fungos e a produção de micotoxinas em grãos de aveia. Cienc. Agrotec. 2006, 30, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Marini, L.J.; Gutkoski, L.C.; Elias, M.C.; Santin, J.A. Qualidade de grãos de aveia sob secagem intermitente em altas temperaturas. Cienc. Rural 2007, 37, 1268–1273. [Google Scholar] [CrossRef]
- Sacchi, C.; González, H.H.L.; Broggi, L.E.; Pacin, A.; Resnik, S.L.; Cano, G.; Taglieri, D. Fungal contamination and mycotoxin natural occurrence in oats for race horses feeding in Argentina. Anim. Feed Sci. Tech. 2009, 152, 330–335. [Google Scholar] [CrossRef]
- Diaz, G.; Perilla, N.; Rojas, Y. Occurrence of aflatoxins in selected Colombian foods. Mycotoxin Res. 2001, 17, 15–20. [Google Scholar] [CrossRef]
- Mallmann, C.A.; Santurio, J.M.; Almeida, C.A.A.; Dilkin, P. Fumonisin B1 levels in cereals and feeds from Southern Brazil. Arq. Inst. Biol. 2001, 68, 41–45. [Google Scholar]
- Ortiz, J.; Camp, J.V.; Mestdagh, F.; Donoso, S.; Meulenaer, B. Mycotoxin co-occurrence in rice, oat flakes and wheat noodles used as staple foods in Ecuador. Food Addit. Contam. A 2013, 30, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.; Jacxsens, L.; Astudillo, G.; Ballesteros, A.; Donoso, S.; Huybregts, L.; Meulenaer, B. Multiple mycotoxin exposure of infants and young children via breastfeeding and complementary/weaning foods consumption in Ecuadorian highlands. Food Chem. Toxicol. 2018, 118, 541–548. [Google Scholar] [CrossRef]
- Graichen, F.A.S.; Martinelli, J.A.; Wesp, C.L.; Federizzi, L.C.; Chaves, M.S. Epidemiological and histological components of crown rust resistance in oat genotypes. Eur. J. Plant Pathol. 2011, 131, 497–510. [Google Scholar] [CrossRef]
- Fredlund, E.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Lindblad, M. Deoxynivalenol and other selected Fusarium toxins in Swedish oats—Occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 276–283. [Google Scholar] [CrossRef]
- Vidal, A.; Marín, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Determination of aflatoxins, deoxynivalenol, ochratoxin A and zearalenone in wheat and oat-based bran supplements sold in the Spanish market. Food Chem. Toxicol. 2013, 53, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Hofgaard, I.S.; Aamot, H.U.; Torp, T.; Jestoi, M.; Lattanzio, V.M.T.; Klemsdal, S.S.; Waalwijk, C.; Van Der Lee, T.; Brodal, G. Associations between Fusarium species and mycotoxins in oats and spring wheat from farmers fields in Norway over a six-year period. World Mycotoxin J. 2016, 9, 365–378. [Google Scholar] [CrossRef]
- Schöneberg, T.; Martin, C.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Keller, B.; Vogelgsang, S. Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. A 2016, 33, 1608–1619. [Google Scholar] [CrossRef] [Green Version]
- Zingales, V.; Fernández-Franzón, M.; Ruiz, M.J. Occurrence, mitigation and in vitro cytotoxicity of nivalenol, a type B trichothecene mycotoxin—Updates from the last decade (2010–2020). Food Chem. Toxicol. 2021, 152, 112182. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Manes, J. Fusarium species, chemotype characterization and trichothecene contamination of durum and soft wheat in an area of Central Italy. J. Sci. Food Agr. 2015, 95, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Sumíková, T.; Chrpová, J.; Džuman, Z.; Salava, J.; Štĕrbová, L.; Palicová, J.; Slavíková, P.; Stránská-Zachariášová, M.; Hajšlová, J. Mycotoxins content and its association with changing patterns of Fusarium pathogens in wheat in the Czech Republic. World Mycotoxin J. 2017, 10, 143–151. [Google Scholar] [CrossRef]
- Almeida, M.I.; Almeida, N.G.; Carvalho, K.L.; Gonçalves, G.A.A.; Silva, C.N.; Santos, E.A.; Garcia, J.C.; Vargas, E.A. Co-occurrence of aflatoxins B1, B2, G1 and G2, ochratoxin A, zearalenone, deoxynivalenol, and citreoviridin in rice in Brazil. Food Addit. Contam. A 2012, 29, 694–703. [Google Scholar] [CrossRef]
- Del Ponte, E.M.; Garda-Buffon, J.; Badiale-Furlong, E. Deoxynivalenol and nivalenol in commercial wheat grain related to Fusarium head blight epidemics in southern Brazil. Food Chem. 2012, 132, 1087–1091. [Google Scholar] [CrossRef] [Green Version]
- Piacentini, K.C.; Savi, G.D.; Pereira, M.E.; Scussel, V.M. Fungi and the natural occurrence of deoxynivalenol and fumonisins in malting barley (Hordeum vulgare L.). Food Chem. 2015, 187, 204–209. [Google Scholar] [CrossRef] [Green Version]
- Calori-Domingues, M.A.; Bernardi, C.M.; Nardin, M.S.; Souza, G.V.; Santos, F.G.R.; Stein, M.A.; Gloria, E.M.; Dias, C.T.S.; Camargo, A.C. Co-occurrence and distribution of deoxynivalenol, nivalenol and zearalenone in wheat from Brazil. Food Addit. Contam. B 2016, 9, 142–151. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Bemvenuti, R.H.; Zorzete, P.; Garcia, F.S.; Corrêa, B. Fungal diversity and natural occurrence of deoxynivalenol and zearalenone in freshly harvested wheat grains from Brazil. Food Chem. 2016, 196, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, K.C.; Rocha, L.O.; Savi, G.D.; Carnielli-Queiroz, L.; Almeida, F.G.; Minella, E.; Corrêa, B. Occurrence of deoxynivalenol and zearalenone in brewing barley grains from Brazil. Mycotoxin Res. 2018, 34, 173–178. [Google Scholar] [CrossRef]
- Savi, G.D.; Piacentini, K.C.; Rocha, L.O.; Carnielli-Queiroz, L.; Furtado, B.G.; Scussel, R.; Zanoni, E.T.; Machado-de-Ávila, R.A.; Corrêa, B.; Angioletto, E. Incidence of toxigenic fungi and zearalenone in rice grains from Brazil. Int. J. Food Microbiol. 2018, 270, 5–13. [Google Scholar] [CrossRef]
- Iwase, C.H.T.; Piacentini, K.C.; Giomo, P.P.; Čumová, M.; Wawroszová, S.; Běláková, S.; Minelli, E.; Rocha, L.O. Characterization of the Fusarium sambucinum species complex and detection of multiple mycotoxins in Brazilian barley samples. Food Res. Int. 2020, 136, 109336. [Google Scholar] [CrossRef] [PubMed]
- Scudamore, K.A.; Bailie, H.; Patel, S.; Edwards, S.G. Occurrence and fate of Fusarium mycotoxins during commercial processing of oats in the UK. Food Addit. Contam. 2007, 24, 1374–1385. [Google Scholar] [CrossRef]
- Ndoye, M.; Zhang, J.B.; Wang, J.H.; Gong, A.D.; Li, H.P.; Qu, B.; Li, S.J.; Liao, Y.C. Nivalenol and 15-acetyldeoxynivalenol chemotypes of Fusarium graminearum clade species are prevalent on maize throughout China. J. Phytopathol. 2012, 160, 519–524. [Google Scholar] [CrossRef]
- Jang, J.Y.; Baek, S.G.; Choi, J.H.; Kim, S.; Kim, J.; Kim, D.W.; Yun, S.H.; Lee, T. Characterization of nivalenol-producing Fusarium asiaticum that causes cereal head blight in Korea. Plant Pathol. J. 2019, 35, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Pinto, V.E.; Terminiello, L.A.; Basilico, J.C.; Ritieni, A. Natural occurrence of nivalenol and mycotoxigenic potential of Fusarium graminearum strains in wheat affected by Head Blight in Argentina. Braz. J. Microbiol. 2008, 39, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Schollenberger, M.; Müller, H.M.; Rüfle, M.; Suchy, S.; Planck, S.; Drochner, W. Survey of Fusarium toxins in foodstuffs of plant origin marketed in Germany. Int. J. Food Microbiol. 2005, 97, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional oats. Food Addit. Contam. A 2009, 26, 1063–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hietaniemi, V.; Rämö, S.; Yli-Matila, T.; Jestoi, M.; Peltonen, S.; Kartio, M.; Sieviläinen, E.; Koivisto, T.; Parikka, P. Update survey of Fusarium species and toxins in Finnish cereal grains. Food Addit. Contam. A 2016, 33, 831–848. [Google Scholar] [CrossRef]
- Yan, P.; Liu, Z.; Liu, S.; Yao, L.; Liu, Y.; Wu, Y.; Gong, Z. Natural occurrence of deoxynivalenol and its acetylated derivatives in Chinese maize and wheat collected in 2017. Toxins 2020, 12, 200. [Google Scholar] [CrossRef] [Green Version]
- Cheat, S.; Gerez, J.R.; Cognié, J.; Alassane-Kpembi, I.; Bracarense, A.P.F.L.; Raymond-Letron, I.; Oswald, I.P.; Kolf-Clauw, M. Nivalenol has a greater impact than deoxynivalenol on pig jejunum mucosa in vitro on explants and in vivo on intestinal loops. Toxins 2015, 7, 1945–1961. [Google Scholar] [CrossRef]
- Spaen, J.; Silva, J.V.C. Oat proteins: Review of extraction methods and techno-functionality for liquid and semi-solid applications. LWT 2021, 147, 111478. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Springer: Boston, MA, USA, 2009. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Wiley-Blackwell: Iowa, NJ, USA, 2006. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Sarver, B.A.J.; Balajee, S.A.; Schroers, H.J.; Summerbell, R.C.; Robert, V.A.R.G.; Crous, P.W.; Zhang, N.; et al. Internet-accessible DNA sequence database for identifying fusaria from human and animal infections. J. Clin. Microbiol. 2010, 48, 3708–3718. [Google Scholar] [CrossRef] [Green Version]
- Laurence, M.H.; Summerell, B.A.; Liew, E.C.Y. Fusarium oxysporum f. sp. canariensis: Evidence for horizontal gene transfer of putative pathogenicity genes. Plant Pathol. 2015, 64, 1068–1075. [Google Scholar]
- Ward, T.J.; Bielawski, J.P.; Kistler, H.C.; Sullivan, E.; O’Donnell, K. Ancestral polymorphism and adaptative evolution in the trichothecene mycotoxin gene cluster of phytopathogenic Fusarium. Proc. Natl. Acad. Sci. USA 2002, 99, 9278–9283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Sarver, B.A.J.; Brandt, M.; Chang, D.C.; Noble-Wang, J.; Park, B.J.; Sutton, D.A.; Benjamin, L.; Lindsley, M.; Padhye, A.; et al. Phylogenetic diversity and microsphere array-based genotyping of human pathogenic fusaria, including isolates from the multistate contact lens-associated U.S. keratitis outbreaks of 2005 and 2006. J. Clin. Microbiol. 2007, 45, 2235–2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L. PAUP* Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Rambaut, A. FigTree v1.3.1; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, Scotland, 2010. [Google Scholar]
- Arraché, E.R.S.; Fontes, M.R.V.; Buffon, J.G.; Badiale-Furlong, E. Trichothecenes in wheat: Methodology, occurrence and human exposure risk. J. Cereal Sci. 2018, 82, 129–137. [Google Scholar] [CrossRef]


{kind=link}
| Oat Origin | RS | PR | Average Count (CFU/g) |
|---|---|---|---|
| Oat aw a | 0.54 | 0.51 | |
| Genera of Fungi | Frequency (%) | ||
| Fusarium | 37.3 | 33.1 | 1.8 × 105 |
| Phoma | 15.4 | 11.1 | 7.7 × 104 |
| Epicoccum | 13.8 | 11.3 | 7.1 × 104 |
| Alternaria | 9.6 | 16.3 | 5.9 × 104 |
| Cladosporium | 7 | 6.9 | 3.3 × 104 |
| Penicillium | 4.4 | 1.9 | 2.2 × 104 |
| Aspergillus | 3.4 | ND | 2.3 × 104 |
| Drechslera | 3 | 1.7 | 1.4 × 104 |
| Pestalotiopsis | 1.5 | 0.3 | 4 × 103 |
| Mucor | 1.5 | 1.7 | 7 × 103 |
| Rhizopus | 1.5 | 2.6 | 9 × 103 |
| Curvularia | 0.8 | ND | 6 × 103 |
| Trichoderma | 0.8 | ND | 6 × 103 |
| Nigrospora | ND | 13.1 | 4.3 × 104 |
| Fusarium Species Complexes | Frequency (%) | ||
| FSAMSC | 93.8 | 85.5 | 1.7 × 105 |
| FTSC | 3.2 | 0 | 7.6 × 103 |
| FFSC | 3 | 6.7 | 4.3 × 104 |
| FIESC | ND | 5 | 5.6 × 103 |
| FSSC | ND | 2.8 | 3.1 × 103 |
| Region | NIV | DON | 15-ADON | 3-ADON | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration (µg/kg) | Concentration (µg/kg) | Concentration (µg/kg) | Concentration (µg/kg) | |||||||||
| Mean | Median | Range | Mean | Median | Range | Mean | Median | Range | Mean | Median | Range | |
| PR | 28.6 | 330.3 | ND-820 | 45 | 540.1 | ND-1620 | 7.7 | 349.7 | ND-723.3 | 18.8 | 648.6 | ND-2546.7 |
| RS | 46.7 | 778.3 | ND-7716.7 | 35 | 503.2 | ND-1610 | 3.6 | 157.8 | ND-420 | 14.8 | 491.7 | ND-3333.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, M.; Iwase, C.H.T.; Bertozzi, B.G.; Caramês, E.T.S.; Carnielli-Queiroz, L.; Langaro, N.C.; Furlong, E.B.; Correa, B.; Rocha, L.O. Survey of Freshly Harvested Oat Grains from Southern Brazil Reveals High Incidence of Type B Trichothecenes and Associated Fusarium Species. Toxins 2021, 13, 855. https://doi.org/10.3390/toxins13120855
Pinheiro M, Iwase CHT, Bertozzi BG, Caramês ETS, Carnielli-Queiroz L, Langaro NC, Furlong EB, Correa B, Rocha LO. Survey of Freshly Harvested Oat Grains from Southern Brazil Reveals High Incidence of Type B Trichothecenes and Associated Fusarium Species. Toxins. 2021; 13(12):855. https://doi.org/10.3390/toxins13120855
Chicago/Turabian StylePinheiro, Mariana, Caio H. T. Iwase, Bruno G. Bertozzi, Elem T. S. Caramês, Lorena Carnielli-Queiroz, Nádia C. Langaro, Eliana B. Furlong, Benedito Correa, and Liliana O. Rocha. 2021. "Survey of Freshly Harvested Oat Grains from Southern Brazil Reveals High Incidence of Type B Trichothecenes and Associated Fusarium Species" Toxins 13, no. 12: 855. https://doi.org/10.3390/toxins13120855
APA StylePinheiro, M., Iwase, C. H. T., Bertozzi, B. G., Caramês, E. T. S., Carnielli-Queiroz, L., Langaro, N. C., Furlong, E. B., Correa, B., & Rocha, L. O. (2021). Survey of Freshly Harvested Oat Grains from Southern Brazil Reveals High Incidence of Type B Trichothecenes and Associated Fusarium Species. Toxins, 13(12), 855. https://doi.org/10.3390/toxins13120855