Effect of the Combined Compound Probiotics with Glycyrrhinic Acid on Alleviating Cytotoxicity of IPEC-J2 Cells Induced by Multi-Mycotoxins

Abstract
:1. Introduction
2. Results
2.1. Screening the Optimal Microbial Combinations for High Degradation Rate of Multi-Mycotoxins
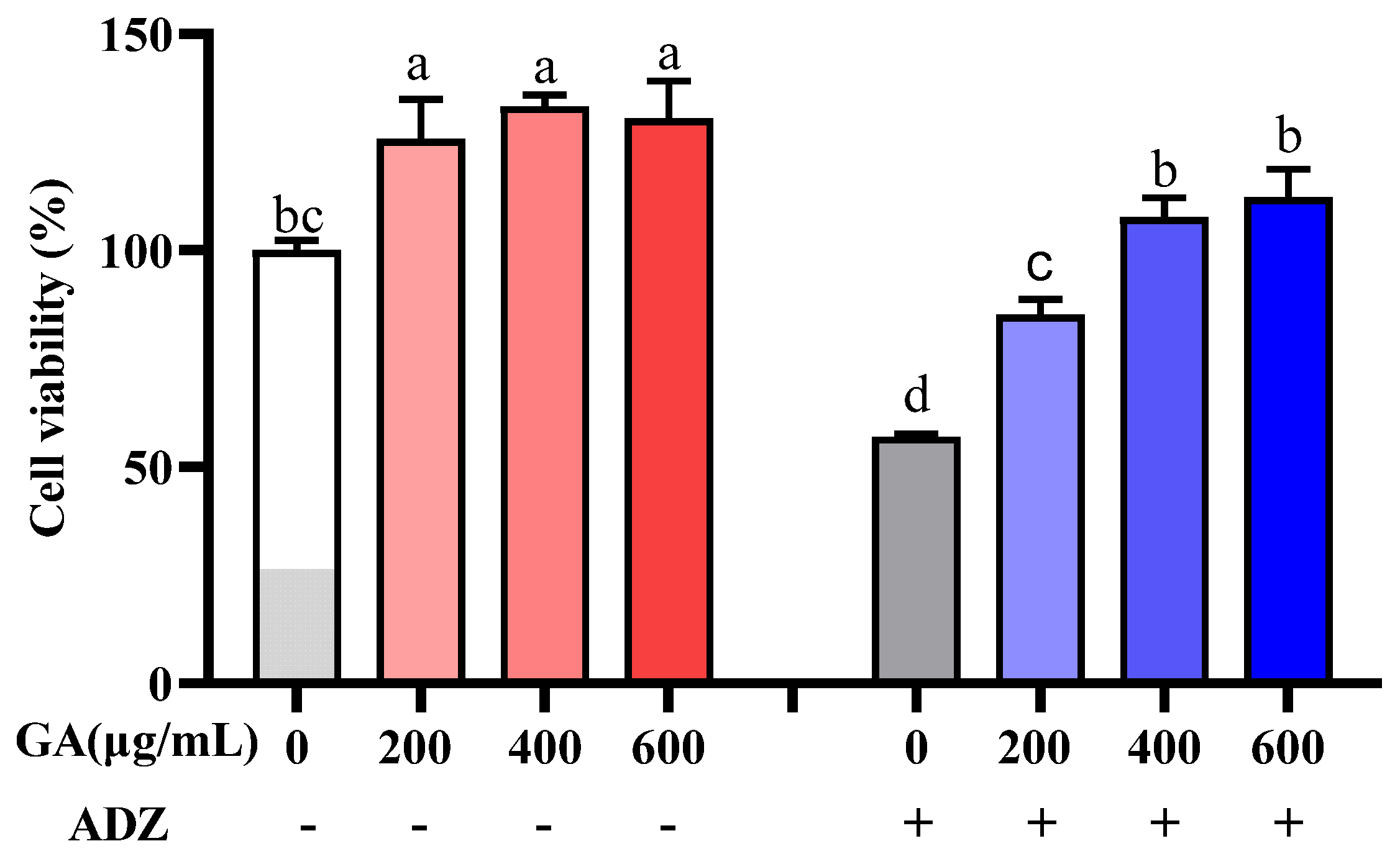
2.2. IPEC-J2 Cell Viability Affected by GA
2.3. Effect of CP, GA or CGA on Alleviating the Viability of IPEC-J2 Cells Induced by ADZ
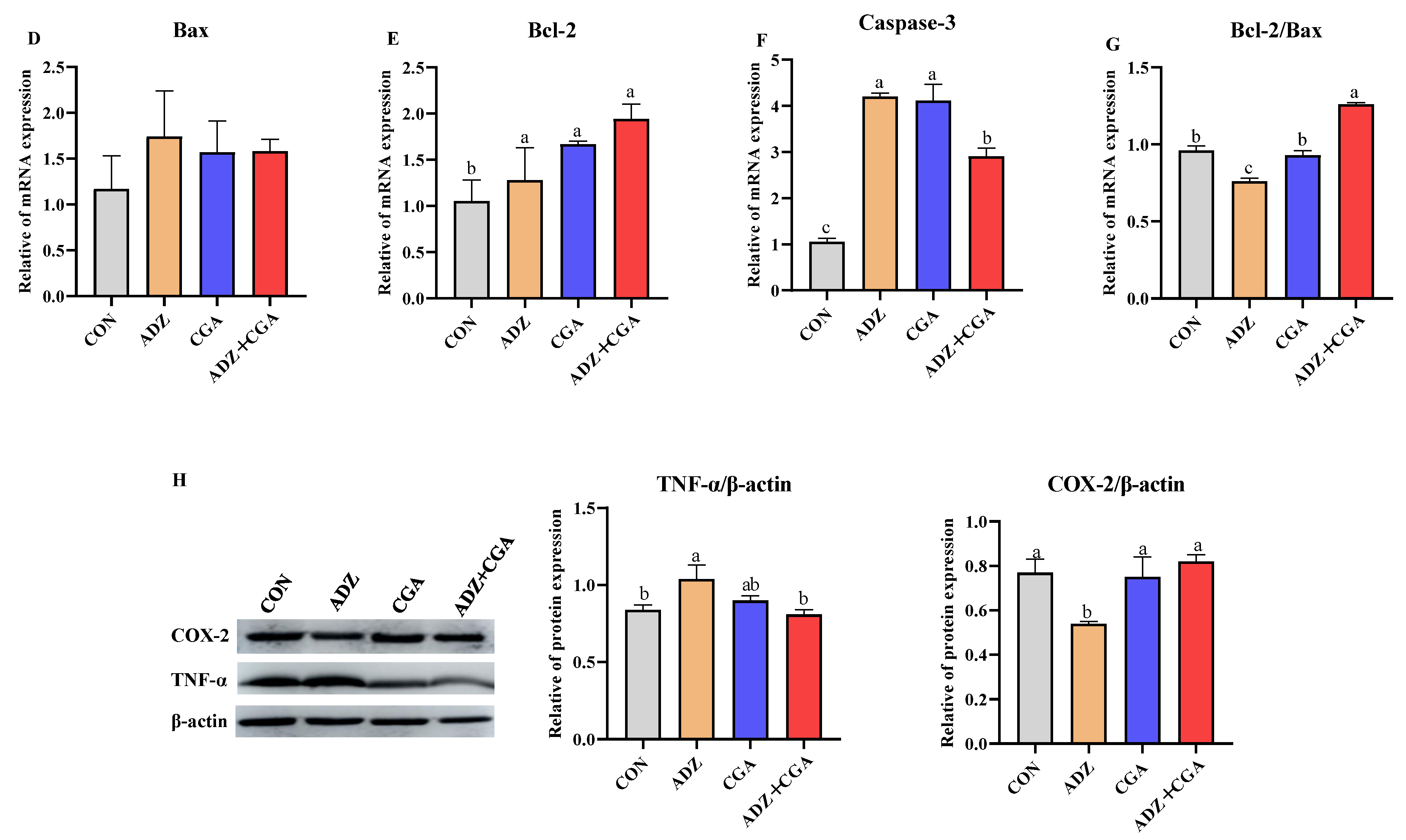
2.4. CGA CGA Alleviates ADZ-Induced Injury in IPEC-J2 Cells
2.5. Effect of CGA on Alleviating IPEC-J2 Cell Inflammation Induced by ADZ
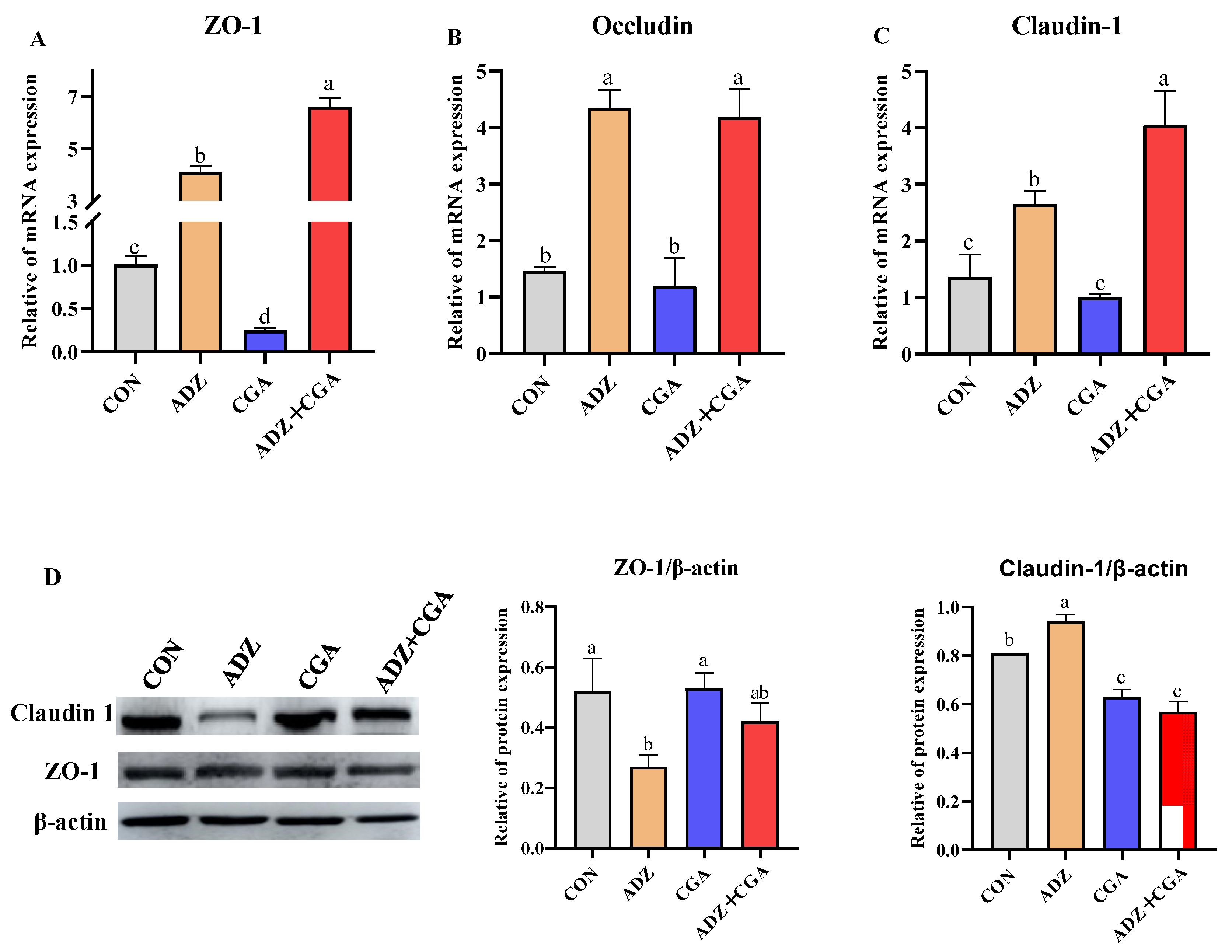
2.6. CGA Increasing mRNA Abundances and Protein Expressions of Tight Junction Protein Genes in Cells with ADZ-Induced IPEC-J2 Injury
2.7. CGA Altering the Nutrient-Transporter Gene mRNA and Protein Expressions in IPEC-J2 Cells Induced by ADZ
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. The Combinations of AFB1, ZEA and DON
5.2. Microbes and Glycyrrhinic Acid (GA) Preparation and Combination
5.3. Cell Culture and Treatments
5.4. Cell Viability Was Measured
5.5. Annexin V-FITC/PI Apoptosis Determination
5.6. RNA Extraction and Quantitative Real-Time PCR
5.7. Western Blotting Analysis
5.8. Data Analysis and Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gajęcki, M.T.; Gajęcka, M.; Zielonka, U. The presence of mycotoxins in feed and their influence on animal health. Toxins 2020, 12, e663. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375–381. [Google Scholar] [CrossRef]
- Ates, E.; Mittendorf, K.; Stroka, J.; Senyuva, H. Determination of fusarium mycotoxins in wheat, maize and animal feed using on-line clean-up with high resolution mass spectrometry. Food Addit. Contam. Part A 2013, 30, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Holanda, D.M.; Yiannikouris, A.; Kim, S.W. Investigation of the efficacy of a postbiotic yeast cell wall-based blend on newly-weaned pigs under a dietary challenge of multiple mycotoxins with emphasis on deoxynivalenol. Toxins 2020, 12, e504. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, S.; Wang, J.; Luo, C.; Zhao, S.; Zheng, N. Modulation of intestinal epithelial permeability in differentiated Caco-2 cells exposed to aflatoxin M1 and ochratoxin a individually or collectively. Toxins 2017, 10, e13. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Guo, C.; Yu, S.; Zhu, L.; Wang, Y.; Hu, H.; Deng, J. Progress in mycotoxins affecting intestinal mucosal barrier function. Int. J. Mol. Sci. 2019, 20, e2777. [Google Scholar] [CrossRef]
- Yang, C.; Song, G.; Lim, W. Effects of mycotoxin-contaminated feed on farm animals. J. Hazard. Mater. 2020, 389, 122087. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, N.; Liu, J.; Li, F.D.; Li, S.L.; Wang, J.Q. Aflatoxin B1 and aflatoxin M1 induced cytotoxicity and DNA damage in differentiated and undifferentiated Caco-2 cells. Food Chem. Toxicol. 2015, 83, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Akinrinmade, F.J.; Akinrinde, A.S.; Amid, A. Changes in serum cytokine levels, hepatic and intestinal morphology in aflatoxin B1-induced injury: Modulatory roles of melatonin and flavonoid-rich fractions from Chromolena odorata. Mycotoxin Res. 2016, 32, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zha, A.; Yuan, D.; Cui, Z.; Qi, M.; Liao, S.; Liao, P.; Tan, B. The evaluation of the antioxidant and intestinal protective effects of baicalin-copper in deoxynivalenol-challenged piglets. Oxid. Med. Cell. Longev. 2020, 2020, 5363546. [Google Scholar] [CrossRef] [Green Version]
- Liao, P.; Liao, M.; Li, L.; Tan, B.; Yin, Y. Effect of deoxynivalenol on apoptosis, barrier function, and expression levels of genes involved in nutrient transport, mitochondrial biogenesis and function in IPEC-J2 cells. Toxicol. Res. 2017, 6, 866–877. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef]
- Gajęcka, M.; Tarasiuk, M.; Zielonka, Ł.; Dąbrowski, M.; Gajęcki, M. Risk assessment for changes in the metabolic profile and body weights of pre-pubertal gilts during long-term monotonic exposure to low doses of zearalenone (ZEN). Res. Vet. Sci. 2016, 109, 169–180. [Google Scholar] [CrossRef]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of zearalenone in the porcine intestinal tract. Molecules 2016, 22, e18. [Google Scholar] [CrossRef]
- Lewczuk, B.; Przybylska-Gornowicz, B.; Gajęcka, M.; Targońska, K.; Ziółkowska, N.; Prusik, M.; Gajęcki, M. Histological structure of duodenum in gilts receiving low doses of zearalenone and deoxynivalenol in feed. Exp. Toxicol. Pathol. 2016, 68, 157–166. [Google Scholar] [CrossRef]
- Guo, Y.; Huo, X.; Zhao, L.; Ma, Q.; Zhang, J.; Ji, C.; Zhao, L. Protective effects of Bacillus subtilis ANSB060, Bacillus subtilis ANSB01G, and Devosia sp. ANSB714-based mycotoxin biodegradation agent on mice fed with naturally moldy diets. Probiotics Antimicrob. Proteins 2020, 12, 994–1001. [Google Scholar] [CrossRef]
- Zhou, J.; Ao, X.; Lei, Y.; Ji, C.; Ma, Q. Bacillus subtilis ANSB01G culture alleviates oxidative stress and cell apoptosis induced by dietary zearalenone in first-parity gestation sows. Anim. Nutr. 2020, 6, 372–378. [Google Scholar] [CrossRef]
- Suresh, G.; Cabezudo, I.; Pulicharla, R.; Cuprys, A.; Rouissi, T.; Brar, S.K. Biodegradation of aflatoxin B1 with cell-free extracts of Trametes versicolor and Bacillus subtilis. Res. Vet. Sci. 2020, 133, 85–91. [Google Scholar] [CrossRef]
- Armando, M.R.; Dogi, C.A.; Rosa, C.A.; Dalcero, A.M.; Cavaglieri, L.R. Saccharomyces cerevisiae strains and the reduction of Aspergillus parasiticus growth and aflatoxin B1 production at different interacting environmental conditions, in vitro. Food Addit. Contam. Part A 2012, 29, 1443–1449. [Google Scholar] [CrossRef]
- Wall-Martínez, H.A.; Pascari, X.; Bigordà, A.; Ramos, A.J.; Marín, S.; Sanchis, V. The fate of fusarium mycotoxins (deoxynivalenol and zearalenone) through wort fermenting by Saccharomyces yeasts (S. cerevisiae and S. pastorianus). Food Res. Int. 2019, 126, 108587. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, J.; Wang, P.; Yin, Q.Q.; Huang, W.W.; Liu, C.Q.; Bai, X.X.; Zhu, Q.; Gao, T.Z.; Zhou, P. Effects of Saccharomyces cerevisiae on alleviating cytotoxicity of porcine jejunal epithelia cells induced by deoxynivalenol. AMB Express 2019, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhang, S.; Xie, Y.; Li, Q. Purification and characteristics of an aflatoxin B1 degradation enzyme isolated from Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2019, 366, fnz034. [Google Scholar] [CrossRef]
- Li, R.; Wu, K.; Li, Y.; Liang, X.; Lai, K.P.; Chen, J. Integrative pharmacological mechanism of vitamin C combined with glycyrrhizic acid against COVID-19: Findings of bioinformatics analyses. Brief. Bioinform. 2021, 22, 1161–1174. [Google Scholar] [CrossRef]
- Wang, H.; Ge, X.; Qu, H.; Wang, N.; Zhou, J.; Xu, W.; Xie, J.; Zhou, Y.; Shi, L.; Qin, Z. Glycyrrhizic acid inhibits proliferation of gastric cancer cells by inducing cell cycle arrest and apoptosis. Cancer Manag. Res. 2020, 12, 2853–2861. [Google Scholar] [CrossRef]
- Honda, H.; Nagai, Y.; Matsunaga, T.; Saitoh, S.; Akashi-Takamura, S.; Hayashi, H.; Fujii, I.; Miyake, K.; Muraguchi, A.; Takatsu, K. Glycyrrhizin and isoliquiritigenin suppress the LPS sensor toll-like receptor 4/MD-2 complex signaling in a different manner. J. Leukoc. Biol. 2012, 91, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Elsherbini, A.M.; Maysarah, N.M.; El-Sherbiny, M.; Al-Gayyar, M.M.; Elsherbiny, N.M. Glycyrrhizic acid ameliorates sodium nitrite-induced lung and salivary gland toxicity: Impact on oxidative stress, inflammation and fibrosis. Hum. Exp. Toxicol. 2021, 40, 707–721. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yan, G.; Chang, J.; Wang, P.; Lu, F. Comparative transcriptome analysis reveals the protective mechanism of glycyrrhinic acid for deoxynivalenol-induced inflammation and apoptosis in IPEC-J2 cells. Oxid. Med. Cell. Longev. 2020, 2020, 5974157. [Google Scholar] [CrossRef] [PubMed]
- Humer, E.; Lucke, A.; Harder, H.; Metzler-Zebeli, B.U.; Böhm, J.; Zebeli, Q. Effects of citric and lactic acid on the reduction of deoxynivalenol and its derivatives in feeds. Toxins 2016, 8, 285. [Google Scholar] [CrossRef]
- Wu, L.; Li, J.; Li, Y.; Li, T.; He, Q.; Tang, Y.; Liu, H.; Su, Y.; Yin, Y.; Liao, P. Aflatoxin B(1), zearalenone and deoxynivalenol in feed ingredients and complete feed from different province in China. J. Anim. Sci. Biotechnol. 2016, 7, e63. [Google Scholar] [CrossRef] [PubMed]
- Weaver, A.; See, M.; Hansen, J.; Kim, Y.; De Souza, A.; Middleton, T.; Kim, S. The use of feed additives to reduce the effects of aflatoxin and deoxynivalenol on pig growth, organ health and immune status during chronic exposure. Toxins 2013, 5, 1261–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Wang, Q.; Wu, H.; Xia, S.; Guo, H.; Blaženović, I.; Zhang, Y.; Sun, X. Insights into cellular metabolic pathways of the combined toxicity responses of Caco-2 cells exposed to deoxynivalenol, zearalenone and aflatoxin B1. Food Chem. Toxicol. 2019, 126, 106–112. [Google Scholar] [CrossRef]
- Huang, W.; Chang, J.; Wang, P.; Liu, C.; Yin, Q.; Song, A.; Gao, T.; Dang, X.; Lu, F. Effect of compound probiotics and mycotoxin degradation enzymes on alleviating cytotoxicity of swine jejunal epithelial cells induced by aflatoxin B1 and zearalenone. Toxins 2019, 11, e12. [Google Scholar] [CrossRef]
- Shetty, P.H.; Hald, B.; Jespersen, L. Surface binding of aflatoxin B1 by Saccharomyces cerevisiae strains with potential decontaminating abilities in indigenous fermented foods. Int. J. Food Microbiol. 2007, 113, 41–46. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Mavumengwana, V. Degradation and detoxification of AFB1 by Staphylocococcus warneri, Sporosarcina sp. and Lysinibacillus fusiformis. Food Control. 2016, 68, 92–96. [Google Scholar] [CrossRef]
- Zanello, G.; Berri, M.; Dupont, J.; Sizaret, P.; D’inca, R. Saccharomyces cerevisiae modulates immune gene expressions and inhibits enterotoxigenic Escherichia coli-mediated ERK1/2 and p38 signaling pathways. PLoS ONE 2011, 6, e18573. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.C.; Agger, S.; Rainey, P.M. Too much of a good thing: A woman with hypertension and hypokalemia. Clin. Chem. 2009, 55, 2093–2096. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yu, M.; Dong, F.; Shi, J.; Xu, J. Esterase activity inspired selection and characterization of zearalenone degrading bacteria Bacillus pumilus ES-21. Food Control 2017, 77, 57–64. [Google Scholar] [CrossRef]
- Khanian, M.; Karimi-Torshizi, M.-A.; Allameh, A. Alleviation of aflatoxin-related oxidative damage to liver and improvement of growth performance in broiler chickens consumed Lactobacillus plantarum 299v for entire growth period. Toxicon 2019, 158, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Chang, J.; Wang, P.; Liu, C.; Gao, T. Effect of the combined compound probiotics with mycotoxin–degradation enzyme on detoxifying aflatoxin B1 and zearalenone. J. Toxicol. Sci. 2018, 43, 377–385. [Google Scholar] [CrossRef]
- Gu, Y.Y.; Chen, M.H.; May, B.H.; Liao, X.Z.; Liu, J.H.; Tao, L.T.; Man-Yuen Sze, D.; Zhang, A.L.; Mo, S.L. Matrine induces apoptosis in multiple colorectal cancer cell lines in vitro and inhibits tumour growth with minimum side effects in vivo via bcl-2 and caspase-3. Phytomedicine 2018, 51, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Lei, M.Y.; Zhang, N.Y.; Zhao, L.; Krumm, C.S.; Qi, D.S. Hepatotoxic effects of mycotoxin combinations in mice. Food Chem. Toxicol. 2014, 74, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Dinu, D.; Bodea, G.O.; Ceapa, C.D.; Munteanu, M.C.; Roming, F.I.; Serban, A.I.; Hermenean, A.; Costache, M.; Zarnescu, O.; Dinischiotu, A. Adapted response of the antioxidant defense system to oxidative stress induced by deoxynivalenol in Hek-293 cells. Toxicon 2011, 57, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Polk, D.B. Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells. J. Biol. Chem. 2002, 277, 50959–50965. [Google Scholar] [CrossRef] [PubMed]
- Roselli, M.; Finamore, A.; Britti, M.S.; Konstantinov, S.R.; Smidt, H.; Vos, W.D.; Mengheri, E. The novel porcine Lactobacillus sobrius strain protects intestinal cells from enterotoxigenic Escherichia coli K88 infection and prevents membrane barrier damage. J. Nutr. 2007, 137, 2709–2716. [Google Scholar] [CrossRef]
- Franz, S.; Rennert, P.; Woznik, M.; Grützke, J.; Lüdde, A.; Arriero Pais, E.M.; Finsterbusch, T.; Geyer, H.; Mankertz, A.; Friedrich, N. Mumps virus SH protein inhibits NF-κB activation by interacting with TNFR1, IL-1R1, and TLR3 complexes. J. Virol. 2017, 91, e01037-17. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Woo, C.; Turner, P.C.; Wan, M.F.; El-Nezami, H. Individual and combined effects of Fusarium toxins on the mRNA expression of pro-inflammatory cytokines in swine jejunal epithelial cells. Toxicol. Lett. 2013, 220, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Vong, L.; Mcknight, W.; Dicay, M.; Martin, G.R. Endogenous and exogenous hydrogen sulfide promotes resolution of colitis in rats. Gastroenterology 2009, 137, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Wang, Q.; He, Q.; Song, X.; Ye, D.; Gao, F.; Jin, S.; Lian, Q.Q. Novel biphasic role of lipoxinA4 on expression of cyclooxygenase-2 in lipopolysaccharide-stimulated lung fibroblasts. Mediators Inflamm. 2011, 2011, e745340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Zheng, S.; Li, W.; Yang, L.; Liu, Y.; Zheng, X.; Yang, Y.; Yang, L.; Wang, Q.; Smith, F.G. Novel biphasic role of resolvin D1 on expression of cyclooxygenase-2 in lipopolysaccharide-stimulated lung fibroblasts is partly through PI3K/AKT and ERK2 pathways. Mediat. Inflamm. 2013, 2013, 964012. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Kojima, F.; Yang, L.; Crofford, L.J. Sequential induction of pro- and anti-inflammatory prostaglandins and peroxisome proliferators-activated receptor-gamma during normal wound healing: A time course study. Prostag. Leukotr. Ess. 2007, 76, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.S.; Hamberg, M.; Balagunaseelan, N.; Goodman, M.; Morgenstern, R.; Stran Dbac, K.E.; Samuelsson, B.; Rinaldo-Matthis, A.; Haeggström, J. A dynamic Asp-Arg interaction is essential for catalysis in microsomal prostaglandin E2 synthase. P. Natl. Acad. Sci. USA 2016, 113, 972–977. [Google Scholar] [CrossRef]
- Liew, W.P.; Mohd-Redzwan, S. Mycotoxin: Its Impact on Gut Health and Microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. Toxicity of deoxynivalenol and its acetylated derivatives on the intestine: Differential effects on morphology, barrier function, tight junction proteins, and mitogen-activated protein kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Nougayrède, J.-P.; Rio, J.C.D.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharm. 2009, 237, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.H.; Wan, M.L.; El-Nezami, H.; Wang, M. Protective capacity of resveratrol, a natural polyphenolic compound, against deoxynivalenol-induced intestinal barrier dysfunction and bacterial translocation. Chem. Res. Toxicol. 2016, 29, 823–833. [Google Scholar] [CrossRef]
- Akbari, P.; Braber, S.; Gremmels, H.; Koelink, P.J.; Verheijden, K.A.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol: A trigger for intestinal integrity breakdown. FASEB J. 2014, 28, 2414–2429. [Google Scholar] [CrossRef] [PubMed]
- Groestlinger, J.; Seidl, C.; Varga, E.; Del Favero, G.; Marko, D. Combinatory exposure to urolithin A, alternariol, and deoxynivalenol affects colon cancer metabolism and epithelial barrier integrity in vitro. Front. Nutr. 2022, 24, 882222. [Google Scholar] [CrossRef] [PubMed]
- Akbari, P.; Braber, S.; Alizadeh, A.; Verheijden, K.A.; Schoterman, M.H.; Kraneveld, A.D.; Garssen, J.; Fink-Gremmels, J. Galacto-oligosaccharides protect the intestinal barrier by maintaining the tight junction network and modulating the inflammatory responses after a challenge with the mycotoxin deoxynivalenol in human Caco-2 cell monolayers and B6C3F1 mice. J. Nutr. 2015, 145, 1604–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Feng, Z.; He, D.K.; Fan, X.M.; Shen, J. MicroRNA-21 is upregulated during intestinal barrier dysfunction induced by ischemia reperfusion. Kaohsiung J. Med. Sci. 2018, 34, 556–563. [Google Scholar] [CrossRef]
- Liu, Z.; Li, C.; Chen, S.; Lin, H.; Zhao, H.; Liu, M.; Weng, J.; Liu, T.; Li, X.; Lei, C. MicroRNA-21 increases the expression level of occludin through regulating ROCK1 in prevention of intestinal barrier dysfunction. J. Cell Biochem. 2019, 120, 4545–4554. [Google Scholar] [CrossRef] [PubMed]
- Resta-Lenert, S.; Barrett, K.E. Live probiotics protect intestinal epithelial cells from the effects of infection with enteroinvasive Escherichia coli (EIEC). Gut 2003, 52, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Broom, L. Mycotoxins and the intestine. Anim. Nutr. 2015, 1, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, M.J.; Chorfi, Y.; Létourneau-Montminy, M.P.; Guay, F. Effects of deoxynivalenol and sodium meta-bisulphite on nutrient digestibility in growing pigs. Arch. Anim. Nutr. 2019, 73, 360–373. [Google Scholar] [CrossRef] [PubMed]
- Hunder, G.; Schümann, K.; Strugala, G.; Gropp, J.; Forth, W. Influence of subchronic exposure to low dietary deoxynivalenol, a trichothecene mycotoxin, on intestinal absorption of nutrients in mice. Food Chem. Toxicol. 1991, 29, 809–814. [Google Scholar] [CrossRef]
- Scalise, M.; Pochini, L.; Console, L.; Losso, M.A.; Indiveri, C. The human SLC1A5 (ASCT2) amino acid transporter: From function to structure and role in cell biology. Front. Cell. Dev. Biol. 2018, 6, e96. [Google Scholar] [CrossRef] [PubMed]
- Javed, K.; Fairweather, S.J. Amino acid transporters in the regulation of insulin secretion and signalling. Biochem. Soc. Trans. 2019, 47, 571–590. [Google Scholar] [CrossRef]
- Okamura, A.; Koyanagi, S.; Dilxiat, A.; Kusunose, N.; Chen, J.J.; Matsunaga, N.; Shibata, S.; Ohdo, S. Bile acid-regulated peroxisome proliferator-activated receptor-α (PPARα) activity underlies circadian expression of intestinal peptide absorption transporter PepT1/Slc15a1. J. Biol. Chem. 2014, 289, 25296–25305. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Factors | Mycotoxin-Degradation Rates | |||||
|---|---|---|---|---|---|---|---|
| A | B | C | AFB1 | ZEA | DON | Total | |
| 1 | 1 | 1 | 1 | 43.05 ± 2.40 a | 38.26 ± 4.35 ab | 80.49 ± 0.75 ab | 161.80 ± 6.89 a |
| 2 | 1 | 2 | 2 | 26.85 ± 0.64 b | 39.21 ± 9.14 ab | 78.37 ± 0.40 ab | 144.44 ± 8.91 a |
| 3 | 1 | 3 | 3 | 28.16 ± 5.63 b | 46.66 ± 3.66 a | 82.62 ± 1.29 a | 157.44 ± 10.43 a |
| 4 | 2 | 1 | 2 | 42.55 ± 3.18 a | 28.18 ± 1.72 bcd | 75.69 ± 3.05 bc | 146.42 ± 8.99 a |
| 5 | 2 | 2 | 3 | 31.12 ± 8.92 b | 4.78 ± 2.21 e | 76.37 ± 1.26 bc | 112.36 ± 8.38 b |
| 6 | 2 | 3 | 1 | 8.08 ± 2.46 d | 37.03 ± 10.85 abc | 79.39 ± 0.15 ab | 124.50 ± 8.34 b |
| 7 | 3 | 1 | 3 | 28.34 ± 7.30 b | 16.91 ± 5.35 d | 71.52 ± 7.40 c | 116.77 ± 15.23 b |
| 8 | 3 | 2 | 1 | 14.82 ± 8.41 cd | 25.35 ± 7.23 cd | 77.22 ± 1.16 abc | 117.39 ± 15.62 b |
| 9 | 3 | 3 | 2 | 20.36 ± 5.98 bc | 23.08 ± 4.00 d | 76.97 ± 4.52 abc | 120.41 ± 12.56 b |
| K1 | 463.68 | 424.99 | 403.69 | ||||
| K2 | 383.29 | 374.19 | 411.27 | ||||
| K3 | 354.57 | 402.35 | 386.58 | ||||
| R | 109.11 | 50.80 | 24.69 | ||||
| Importance order | A > B > C | ||||||
| The optimal solution | A1B1C1 | ||||||
| Error Source | Sum of Squares | Degree of Freedom | Mean Square | F Value | p-Value |
|---|---|---|---|---|---|
| Corrected Model | 8012.57 | 6 | 1335.43 | 9.32 | 0.00 |
| A | 6397.16 | 2 | 3198.58 | 22.33 | 0.00 |
| B | 1295.41 | 2 | 647.71 | 4.52 | 0.02 |
| C | 320.01 | 2 | 160.00 | 1.12 | 0.35 |
| Error | 2864.83 | 20 | 143.24 | ||
| Corrected Error | 10,877.40 | 26 |
| Primer | Primer Sequence (5′-3′) | Accession Number |
|---|---|---|
| GAPDH | F: ATGACCACAGTCCATGCCATC | XM-004387206 |
| R: CCTGCTTCACCACCTTCTTG | ||
| Bcl-2 | F: AGAGCCGTTTCGTCCCTTTC | XM-003122573.2 |
| R: GCACGTTTCCTAGASTGCAT | ||
| Bax | F: ATGATCGCAGCCGTGGACACG | XM-003355975.1 |
| R: AASTAGATGGTCACCGTCTGC | ||
| Casepase-3 | F: TTGGACTGTGGGATTGAGACG | NM-214131.1 |
| R: CGCTGCACAAAGTGACTGGA | ||
| IL-8 | F: GACCCCAAGGAAAAGTGGGT | NM_213867.1 |
| R: TGACCAGCACAGGAATGAGG | ||
| TNF-α | F: TTCCAGCTGGCCCCTTGAGC | NM_214022 |
| R: GAGGGCATTGGCATACCCAC | ||
| NF-κB | F: CTCGCACAAGGAGACATGAA | NC_006509021 |
| R: ACTCAGCCGGAAGGCATTAT | ||
| ZO-1 | F: CCTGAGTTTGATAGTGGCGTTGA | XM-003353439.2 |
| R: AAATAGATTTCCTGCCCAATTCC | ||
| Occludin | F: ACCCAGCAAASTCATA | NM_001163647.2 |
| R: TCAASTTAAASTGCATA | ||
| Claudin-1 | F: ATTTCAGGTCTGGCTATCTTAGTTGC | NM_001244539.1 |
| R: AGGGCCTTGGTGTTGGGTAA | ||
| ASCT2 | F: CTGGTCTCCTGGATCATGTGG | DQ231578.1 |
| R: CAGGAAGCGGTAGGGGTTTT | ||
| PepT1 | F: CAGACTTASTCCACAACGGA | NM_214347.1 |
| R: TTATCCCGCCAGTACCCAGA | ||
| SGLT1 | F: TCATCATCGTCCTGGTCGTCTC | NM-34044.1 |
| R: CTTCTGGGGCTTCTTGAATGTC | ||
| GLUT2 | F: ATTGTCACAGGCATTCTTGTTAGTCA | NM_001097417.1 |
| R: TTCACTTGATGCTTCTTCCCTTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wang, X.; Chang, J.; Wang, P.; Liu, C.; Yuan, L.; Yin, Q.; Zhu, Q.; Lu, F. Effect of the Combined Compound Probiotics with Glycyrrhinic Acid on Alleviating Cytotoxicity of IPEC-J2 Cells Induced by Multi-Mycotoxins. Toxins 2022, 14, 670. https://doi.org/10.3390/toxins14100670
Wang L, Wang X, Chang J, Wang P, Liu C, Yuan L, Yin Q, Zhu Q, Lu F. Effect of the Combined Compound Probiotics with Glycyrrhinic Acid on Alleviating Cytotoxicity of IPEC-J2 Cells Induced by Multi-Mycotoxins. Toxins. 2022; 14(10):670. https://doi.org/10.3390/toxins14100670
Chicago/Turabian StyleWang, Lijun, Xiaomin Wang, Juan Chang, Ping Wang, Chaoqi Liu, Lin Yuan, Qingqiang Yin, Qun Zhu, and Fushan Lu. 2022. "Effect of the Combined Compound Probiotics with Glycyrrhinic Acid on Alleviating Cytotoxicity of IPEC-J2 Cells Induced by Multi-Mycotoxins" Toxins 14, no. 10: 670. https://doi.org/10.3390/toxins14100670
APA StyleWang, L., Wang, X., Chang, J., Wang, P., Liu, C., Yuan, L., Yin, Q., Zhu, Q., & Lu, F. (2022). Effect of the Combined Compound Probiotics with Glycyrrhinic Acid on Alleviating Cytotoxicity of IPEC-J2 Cells Induced by Multi-Mycotoxins. Toxins, 14(10), 670. https://doi.org/10.3390/toxins14100670