Ameliorative Effects of Luteolin and Activated Charcoal on Growth Performance, Immunity Function, and Antioxidant Capacity in Broiler Chickens Exposed to Deoxynivalenol

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
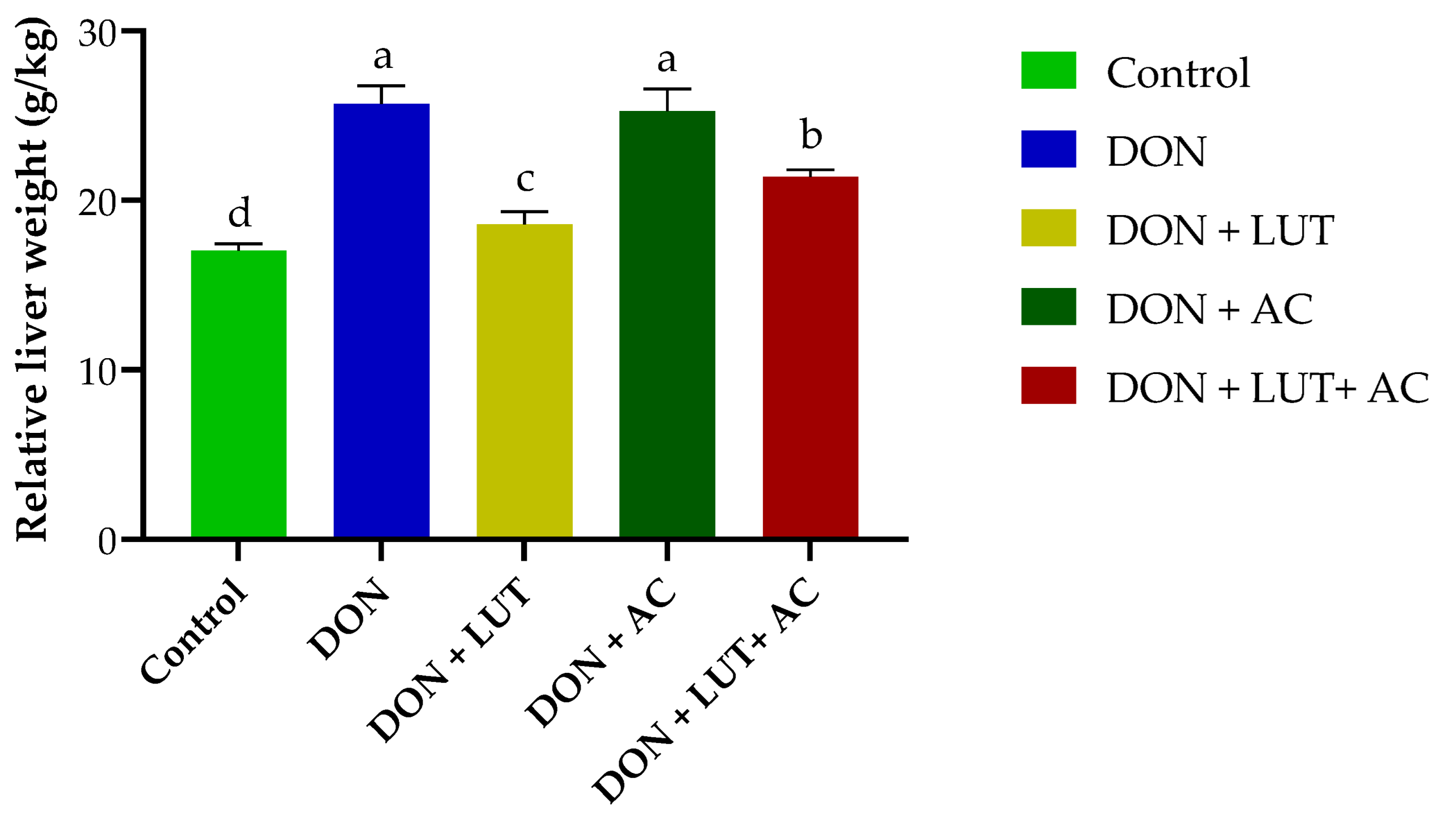
2.1. Compared with the DON Group, LUT and AC Supplementation Significantly Improved the Growth Performance
2.2. Serum Biochemical Analysis Based on Biochemical Criteria
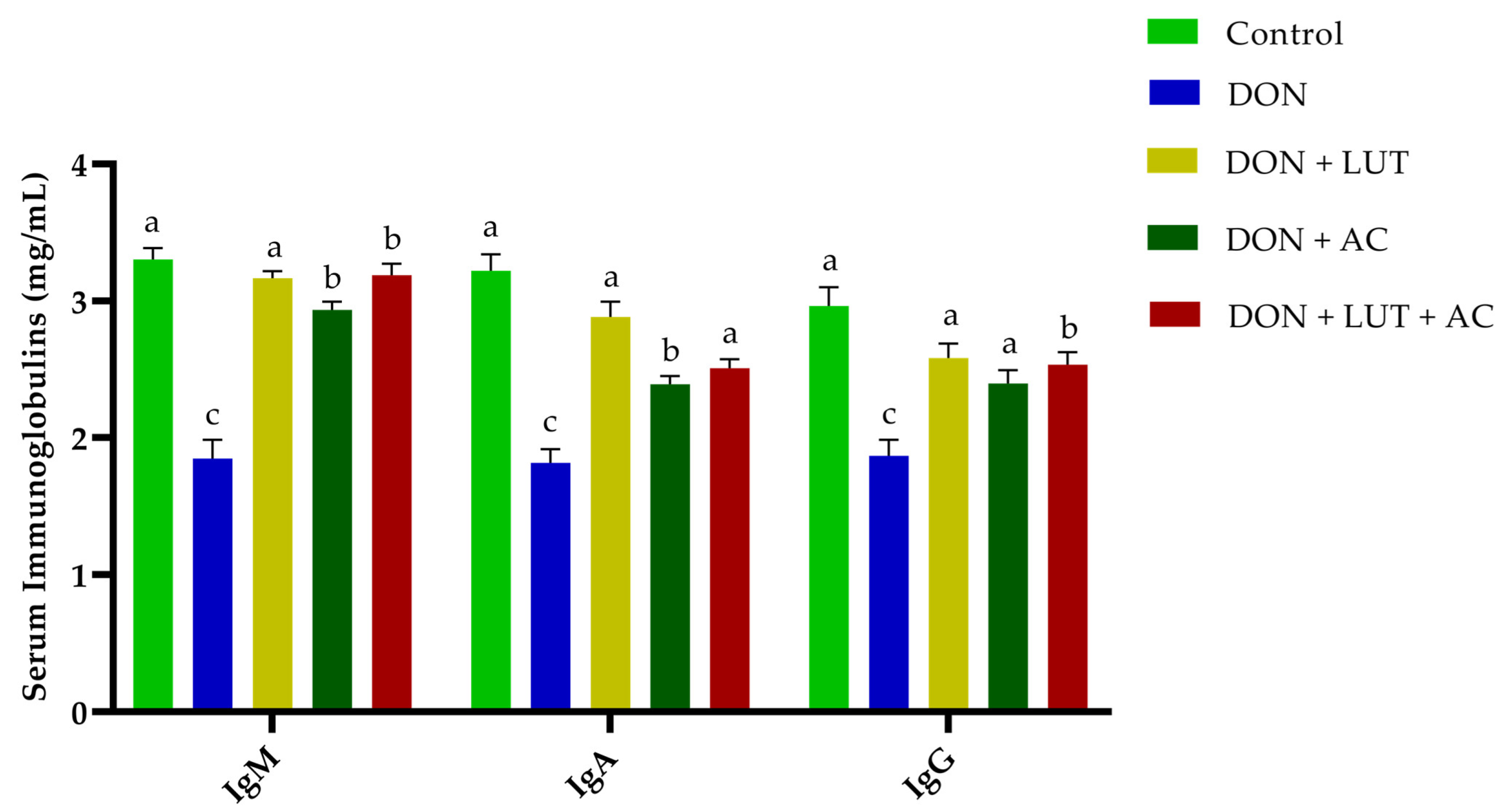
2.3. Immunoglobulin Levels in Serum
2.4. Analyses of Serum Antioxidants
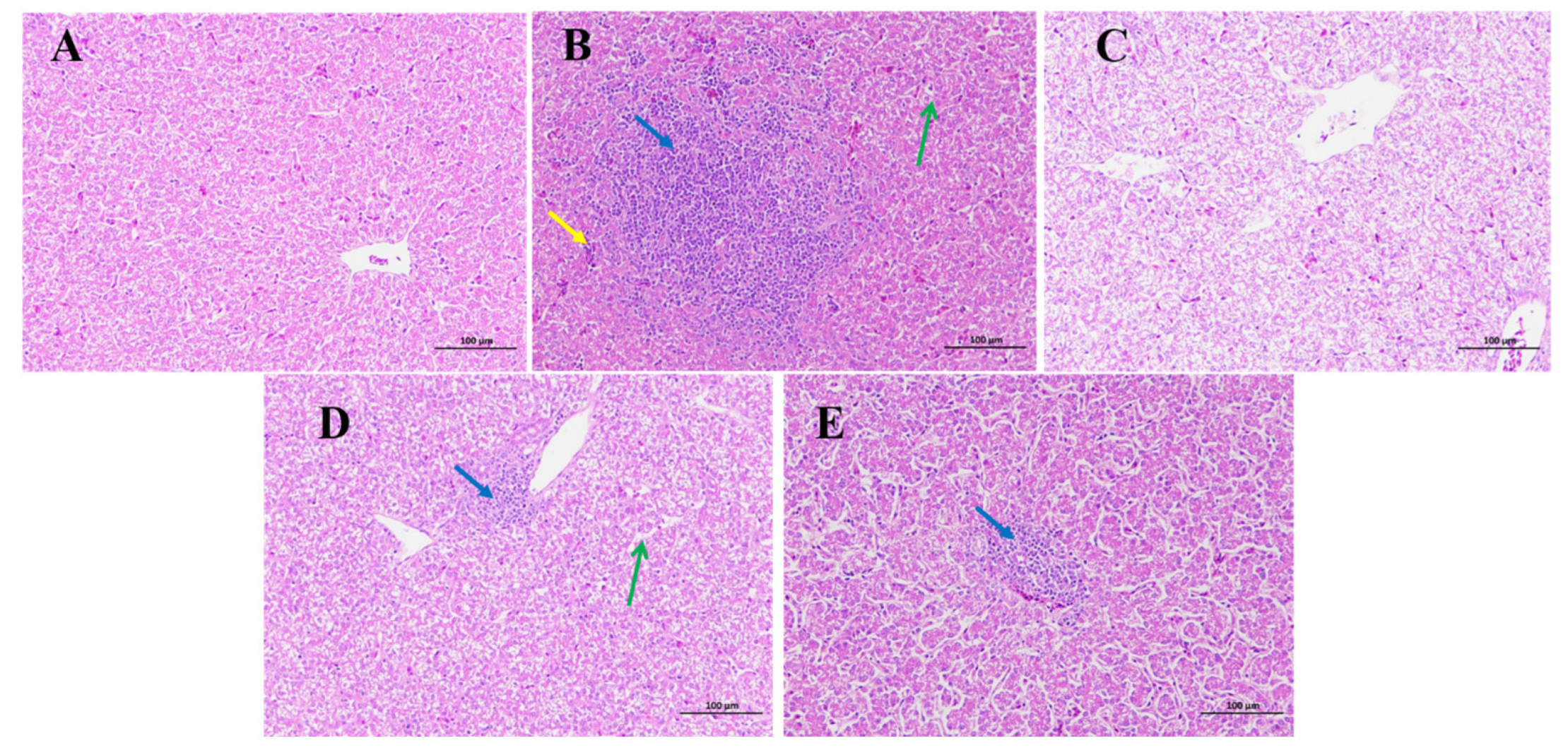
2.5. LUT and AC Prevent Damage to the Liver Caused by DON in Broilers
2.6. LUT and AC Improve Liver Antioxidant Parameters
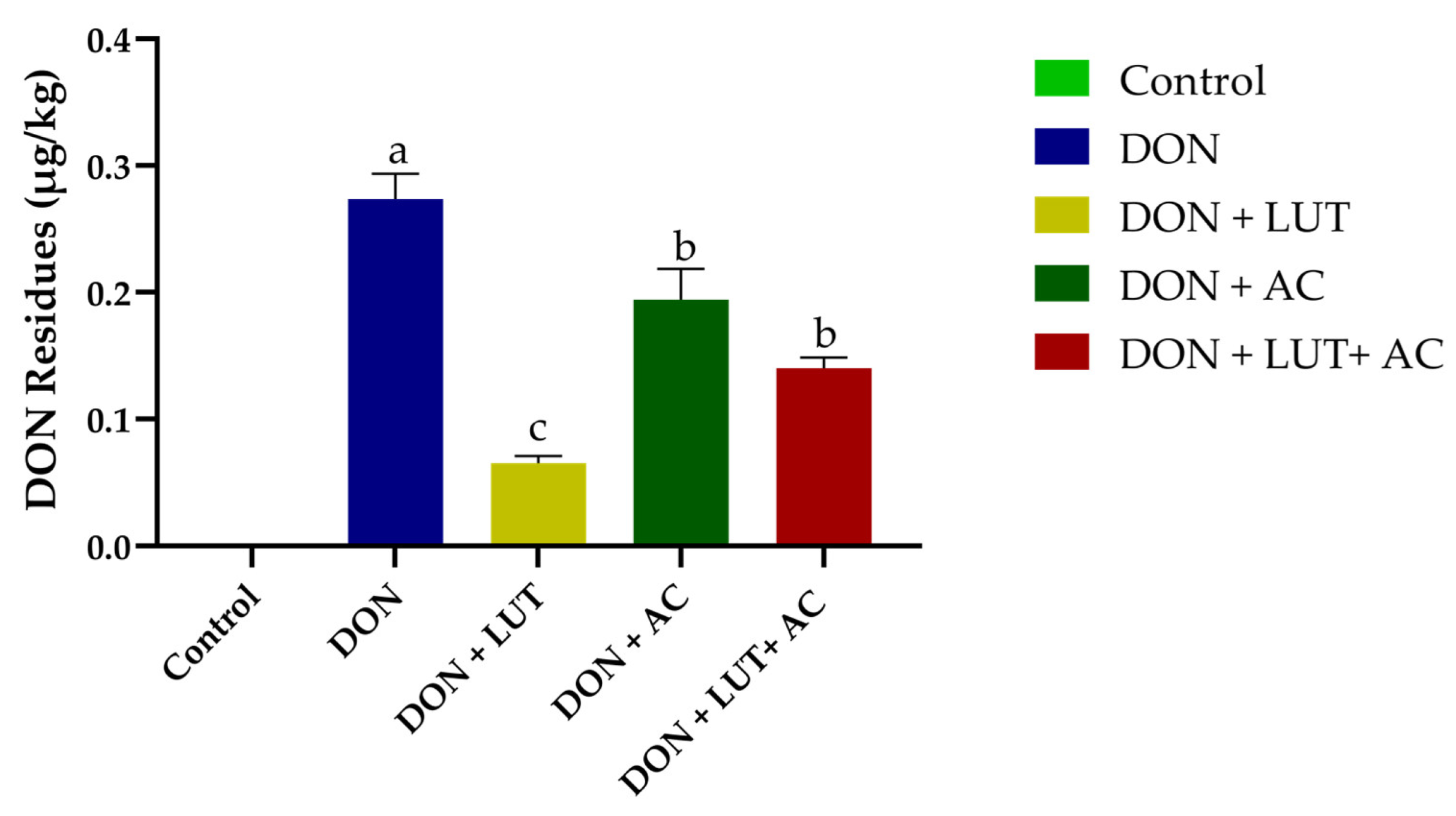
2.7. Residues of DON in the Liver
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Analysis and Preparation for the DON Toxin
5.2. Animals and Experimental Design
5.3. Sample Collection and Measurement
5.4. Histopathological and Biochemical Analysis of Serum
5.5. Detection of Antioxidative Enzymes in Serum and Liver
5.6. Analyses of Serum Immunoglobulin Concentrations
5.7. HPLC Analysis of DON Residues in Liver
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nematbakhsh, S.; Selamat, J.; Idris, L.H.; Abdull Razis, A.F. Chicken Authentication and Discrimination via Live Weight, Body Size, Carcass Traits, and Breast Muscle Fat Content Clustering as Affected by Breed and Sex Varieties in Malaysia. Foods 2021, 10, 1575. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture (Foreign Agricultural Service). Livestock and Poultry: World Markets and Trade China Total Meat Imports Forecast Higher in 2023; United States Department of Agriculture (Foreign Agricultural Service): Washington, DC, USA, 2023; Volume 2022, pp. 8–9.
- OECD/FAO. 6 Meat. Agricultural Outlook 2021–2030; OECD: Paris, France; FAO: Rome, Italy, 2021; pp. 163–177. [Google Scholar]
- Deng, S.; Xing, T.; Li, C.; Xu, X.; Zhou, G. The Effect of Breed and Age on the Growth Performance, Carcass Traits and Metabolic Profile in Breast Muscle of Chinese Indigenous Chickens. Foods 2022, 11, 483. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, W.K. Dietary Application of Tannins as a Potential Mitigation Strategy for Current Challenges in Poultry Production: A Review. Animals 2020, 10, 2389. [Google Scholar] [CrossRef] [PubMed]
- Ochieng, P.E.; Scippo, M.L.; Kemboi, D.C.; Croubels, S.; Okoth, S.; Kang’ethe, E.K.; Doupovec, B.; Gathumbi, J.K.; Lindahl, J.F.; Antonissen, G. Mycotoxins in Poultry Feed and Feed Ingredients from Sub-Saharan Africa and Their Impact on the Production of Broiler and Layer Chickens: A Review. Toxins 2021, 13, 633. [Google Scholar] [CrossRef]
- Liu, J.D.; Doupovec, B.; Schatzmayr, D.; Murugesan, G.R.; Bortoluzzi, C.; Villegas, A.M.; Applegate, T.J. The Impact of Deoxynivalenol, Fumonisins, and Their Combination on Performance, Nutrient, and Energy Digestibility in Broiler Chickens. Poult. Sci. 2020, 99, 272–279. [Google Scholar] [CrossRef]
- Riahi, I.; Marquis, V.; Pérez-Vendrell, A.M.; Brufau, J.; Esteve-Garcia, E.; Ramos, A.J. Effects of Deoxynivalenol-Contaminated Diets on Metabolic and Immunological Parameters in Broiler Chickens. Animals 2021, 11, 147. [Google Scholar] [CrossRef]
- Grenier, B.; Schwartz-Zimmermann, H.E.; Gruber-Dorninger, C.; Dohnal, I.; Aleschko, M.; Schatzmayr, G.; Moll, W.D.; Applegate, T.J. Enzymatic Hydrolysis of Fumonisins in the Gastrointestinal Tract of Broiler Chickens. Poult. Sci. 2017, 96, 4342–4351. [Google Scholar] [CrossRef]
- Kövesi, B.; Cserháti, M.; Erdélyi, M.; Zándoki, E.; Mézes, M.; Balogh, K. Long-Term Effects of Ochratoxin A on the Glutathione Redox System and Its Regulation in Chicken. Antioxidants 2019, 8, 178. [Google Scholar] [CrossRef] [Green Version]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in Food and Feed, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 89. [Google Scholar]
- Awad, W.A.; Ghareeb, K.; Dadak, A.; Gille, L.; Staniek, K.; Hess, M.; Böhm, J. Genotoxic Effects of Deoxynivalenol in Broiler Chickens Fed Low-Protein Feeds. Poult. Sci. 2012, 91, 550–555. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, H.; Lian, S. Cortisol Excess-Mediated Mitochondrial Damage Following Cold Exposure. Cells 2019, 8, 612. [Google Scholar]
- Deng, N.; Lv, Y.; Bing, Q.; Li, S.; Han, B.; Jiang, H.; Yang, Q.; Wang, X.; Wu, P.; Liu, Y.; et al. Inhibition of the Nrf2 Signaling Pathway Involved in Imidacloprid-Induced Liver Fibrosis in Coturnix japonica. Environ. Toxicol. 2022, 37, 2354–2365. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Kou, J.-J.; Shi, J.-Z.; He, Y.-Y.; Hao, J.-J.; Zhang, H.-Y.; Luo, D.-M.; Song, J.-K.; Yan, Y.; Xie, X.-M.; Du, G.-H.; et al. Luteolin Alleviates Cognitive Impairment in Alzheimer’s Disease Mouse Model via Inhibiting Endoplasmic Reticulum Stress-Dependent Neuroinflammation. Acta Pharmacol. Sin. 2022, 43, 840–849. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Dong, R.; Huang, K.; Wang, C.; Gu, J.; Luo, H.; Liu, K.; Wu, J.; Sun, H.; et al. Luteolin Ameliorates LPS-Induced Acute Liver Injury by Inhibiting TXNIP-NLRP3 Inflammasome in Mice. Phytomedicine 2021, 87, 153586. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Liu, S.; Wu, Y.; Zhou, Q.; Zhang, Y.; Zheng, X.; Han, Y.; Xie, C.; Liu, N. Adsorption of Deoxynivalenol by Pillared Montmorillonite. Food Chem. 2021, 343, 128391. [Google Scholar] [CrossRef]
- Burchacka, E.; Łukaszewicz, M.; Kułażyński, M. Determination of Mechanisms of Action of Active Carbons as a Feed Additive. Bioorg. Chem. 2019, 93, 102804. [Google Scholar] [CrossRef]
- Khatoon, A.; Khan, M.Z.; Abidin, Z.U.; Bhatti, S.A. Effects of Feeding Bentonite Clay upon Ochratoxin A–Induced Immunosuppression in Broiler Chicks. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018, 35, 538–545. [Google Scholar] [CrossRef]
- Oso, A.O.; Akapo, O.; Sanwo, K.A.; Bamgbose, A.M. Utilization of Unpeeled Cassava (Manihot esculenta Crantz) Root Meal Supplemented with or without Charcoal by Broiler Chickens. J. Anim. Physiol. Anim. Nutr. 2014, 98, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, S.A.; Khan, M.Z.; Saleemi, M.K.; Hassan, Z.U. Combating Immunotoxicity of Aflatoxin B1 by Dietary Carbon Supplementation in Broiler Chickens. Environ. Sci. Pollut. Res. 2021, 28, 49089–49101. [Google Scholar] [CrossRef]
- Ruhnau, D.; Hess, C.; Grenier, B.; Doupovec, B.; Schatzmayr, D.; Hess, M.; Awad, W.A. The Mycotoxin Deoxynivalenol (DON) Promotes Campylobacter jejuni Multiplication in the Intestine of Broiler Chickens With Consequences on Bacterial Translocation and Gut Integrity. Front. Vet. Sci. 2020, 7, 573894. [Google Scholar] [CrossRef] [PubMed]
- Andretta, I.; Kipper, M.; Lehnen, C.R.; Hauschild, L.; Vale, M.M.; Lovatto, P.A. Meta-Analytical Study of Productive and Nutritional Interactions of Mycotoxins in Broilers. Poult. Sci. 2011, 90, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Hogan, N.S. Performance Effects of Feed-Borne Fusarium Mycotoxins on Broiler Chickens: Influences of Timing and Duration of Exposure. Anim. Nutr. 2019, 5, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Mgbeahuruike, A.C.; Ejiofor, T.E.; Ashang, M.U.; Ojiako, C.; Obasi, C.C.; Ezema, C.; Okoroafor, O.; Mwanza, M.; Karlsson, M.; Chah, K.F. Reduction of the Adverse Impacts of Fungal Mycotoxin on Proximate Composition of Feed and Growth Performance in Broilers by Combined Adsorbents. Toxins 2021, 13, 430. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Assessment of the Multi-Mycotoxin-Binding Efficacy of a Carbon/Aluminosilicate-Based Product in an in vitro Gastrointestinal Model. J. Agric. Food Chem. 2007, 55, 4810–4819. [Google Scholar] [CrossRef] [PubMed]
- Prapapanpong, J.; Udomkusonsri, P.; Mahavorasirikul, W.; Choochuay, S.; Tansakul, N. in vitro Studies on Gastrointestinal Monogastric and Avian Models to Evaluate the Binding Efficacy of Mycotoxin Adsorbents by Liquid Chromatography-Tandem Mass Spectrometry. J. Adv. Vet. Anim. Res. 2019, 6, 125–132. [Google Scholar] [CrossRef]
- Rajput, S.A.; Shaukat, A.; Wu, K.; Rajput, I.R.; Baloch, D.M.; Akhtar, R.W.; Raza, M.A.; Najda, A.; Rafał, P.; Albrakati, A.; et al. Luteolin Alleviates AflatoxinB1-induced Apoptosis and Oxidative Stress in the Liver of Mice through Activation of Nrf2 Signaling Pathway. Antioxidants 2021, 10, 1268. [Google Scholar] [CrossRef]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailey, W.; Schomaker, S. The Current State of Serum Biomarkers of Hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef]
- Goyarts, T.; Grove, N.; Dänicke, S. Effects of the Fusarium Toxin Deoxynivalenol from Naturally Contaminated Wheat given Subchronically or as One Single Dose on the in vivo Protein Synthesis of Peripheral Blood Lymphocytes and Plasma Proteins in the Pig. Food Chem. Toxicol. 2006, 44, 1953–1965. [Google Scholar] [CrossRef]
- Leung, M.C.K.; Smith, T.K.; Karrow, N.A.; Boermans, H.J. Effects of Foodborne Fusarium Mycotoxins with and without a Polymeric Glucomannan Mycotoxin Adsorbent on Food Intake and Nutrient Digestibility, Body Weight, and Physical and Clinicopathologic Variables of Mature Dogs. Am. J. Vet. Res. 2007, 68, 1122–1129. [Google Scholar] [CrossRef]
- Kim, E.J.; Jeong, S.H.; Cho, J.H.; Ku, H.O.; Pyo, H.M.; Kang, H.G.; Choi, K.H. Plasma Haptoglobin and Immunoglobulins as Diagnostic Indicators of Deoxynivalenol Intoxication. J. Vet. Sci. 2008, 9, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Efrat, S.; Kaempfer, R. Control of Biologically Active Interleukin 2 Messenger RNA Formation in Induced Human Lymphocytes. Proc. Natl. Acad. Sci. USA 1984, 81, 2601–2605. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Naveed, M.; Arain, M.A.; Arif, M.; Abd El-Hack, M.E.; Alagawany, M.; Siyal, F.A.; Soomro, R.N.; Sun, C. Quercetin: Nutritional and Beneficial Effects in Poultry. Worlds Poult. Sci. J. 2017, 73, 355–364. [Google Scholar] [CrossRef]
- Han, B.; Lv, Z.; Han, X.; Li, S.; Han, B.; Yang, Q.; Wang, X.; Wu, P.; Li, J.; Deng, N.; et al. Harmful Effects of Inorganic Mercury Exposure on Kidney Cells: Mitochondrial Dynamics Disorder and Excessive Oxidative Stress. Biol. Trace Elem. Res. 2022, 200, 1591–1597. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, A.; Hanif, S.; Shaukat, I.; Shukat, R.; Rajput, S.A.; Jiang, K.; Akhtar, M.; Yang, Y.; Guo, S.; Shaukat, I.; et al. Upregulated-Gene Expression of pro-Inflammatory Cytokines, Oxidative Stress and Apoptotic Markers through Inflammatory, Oxidative and Apoptosis Mediated Signaling Pathways in Bovine Pneumonia. Microb. Pathog. 2021, 155, 104935. [Google Scholar] [CrossRef]
- Sun, Z.; Li, S.; Yu, Y.; Chen, H.; Ommati, M.M.; Manthari, R.K.; Niu, R.; Wang, J. Alterations in Epididymal Proteomics and Antioxidant Activity of Mice Exposed to Fluoride. Arch. Toxicol. 2018, 92, 169–180. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, C.; Jiang, H.; Han, B.; Wang, X.; Li, S.; Lv, Y.; Lv, Z.; Zhu, Y. Inflammation Response after the Cessation of Chronic Arsenic Exposure and Post-Treatment of Natural Astaxanthin in Liver: Potential Role of Cytokine-Mediated Cell—Cell Interactions. Food Funct. 2020, 11, 9252–9262. [Google Scholar] [CrossRef]
- Shaukat, A.; Yang, C.; Yang, Y.; Guo, Y.-F.; Jiang, K.; Guo, S.; Liu, J.; Zhang, T.; Zhao, G.; Ma, X.; et al. Ginsenoside Rb1: A Novel Therapeutic Agent in Staphylococcus aureus—Induced Acute Lung Injury with Special Reference to Oxidative Stress and Apoptosis. Microb. Pathog. 2020, 143, 104109. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, R.; Su, Y.; Bi, Y.; Li, X.; Zhang, X.; Li, J.; Bao, J. Effects of Acute Cold Stress after Long-Term Cold Stimulation on Antioxidant Status, Heat Shock Proteins, Inflammation and Immune Cytokines in Broiler Heart. Front. Physiol. 2018, 9, 1589. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Chen, S.; Zhou, J.; Chen, J.; Tian, L.; Gao, W.; Zhang, Y.; Ma, A.; Li, L.; Zhou, Z. Luteolin Cocrystals: Characterization, Evaluation of Solubility, Oral Bioavailability and Theoretical Calculation. J. Drug Deliv. Sci. Technol. 2019, 50, 248–254. [Google Scholar] [CrossRef]
- You, L.; Zhao, Y.; Kuca, K.; Wang, X.; Oleksak, P.; Chrienova, Z.; Nepovimova, E.; Jaćević, V.; Wu, Q.; Wu, W. Hypoxia, Oxidative Stress, and Immune Evasion: A Trinity of the Trichothecenes T-2 Toxin and Deoxynivalenol (DON). Arch. Toxicol. 2021, 95, 1899–1915. [Google Scholar] [CrossRef]
- Michalopoulos, G.K. Liver regeneration. J. Cell Physiol. 2007, 213, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Sadiq, F.A.; Liu, W.; Cao, L.; Shen, Z. Protective Effects of Bacillus Subtilis ASAG 216 on Growth Performance, Antioxidant Capacity, Gut Microbiota and Tissues Residues of Weaned Piglets Fed Deoxynivalenol Contaminated Diets. Food Chem. Toxicol. 2021, 148, 111962. [Google Scholar] [CrossRef]
- Bannert, E.; Tesch, T.; Kluess, J.; Valenta, H.; Frahm, J.; Kersten, S.; Kahlert, S.; Renner, L.; Rothkötter, H.J.; Dänicke, S. Plasma Kinetics and Matrix Residues of Deoxynivalenol (DON) and Zearalenone (ZEN) Are Altered in Endotoxaemic Pigs Independent of LPS Entry Site. Mycotoxin Res. 2017, 33, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Ueberschär, K.H.; Valenta, H.; Matthes, S.; Matthäus, K.; Halle, I. Effects of Graded Levels of Fusarium-Toxin-Contaminated Wheat in Pekin Duck Diets on Performance, Health and Metabolism of Deoxynivalenol and Zearalenone. Br. Poult. Sci. 2004, 45, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, Y.; Beier, R.C.; Zhang, H.; De Ruyck, K.; Sun, F.; Cao, X.; Shen, J.; Zhang, S.; Wang, Z. Simultaneous Determination of Type A and B Trichothecenes and Their Main Metabolites in Food Animal Tissues by Ultraperformance Liquid Chromatography Coupled with Triple-Quadrupole Mass Spectrometry. J. Agric. Food Chem. 2015, 63, 8592–8600. [Google Scholar] [CrossRef]
- Wan, D.; Huang, L.; Pan, Y.; Wu, Q.; Chen, D.; Tao, Y.; Wang, X.; Liu, Z.; Li, J.; Wang, L.; et al. Metabolism, Distribution, and Excretion of Deoxynivalenol with Combined Techniques of Radiotracing, High-Performance Liquid Chromatography Ion Trap Time-of-Flight Mass Spectrometry, and Online Radiometric Detection. J. Agric. Food Chem. 2014, 62, 288–296. [Google Scholar] [CrossRef]
- Wang, S.; Yang, J.; Zhang, B.; Wu, K.; Yang, A.; Li, C.; Zhang, J.; Zhang, C.; Rajput, S.A.; Zhang, N.; et al. Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets and IPEC-J2 Cells. Toxins 2018, 10, 541. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Sun, M.; Hong, X.; Duan, J.; Du, D. Survey of Deoxynivalenol Contamination in Agricultural Products in the Chinese Market Using An ELISA Kit. Toxins 2019, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, H.; Yamamoto, K.; Mori, Y.; Asao, N.; Fukunaka, R.; Deguchi, K.; Iida, K.; Miyazaki, S.; Goto, T. Modified Use of a Commercial ELISA Kit for Deoxynivalenol Determination in Rice and Corn Silage. Mycotoxin Res. 2013, 29, 79–88. [Google Scholar] [CrossRef]
- Ji, F.; Li, H.; Xu, J.; Shi, J. Enzyme-Linked Immunosorbent-Assay for Deoxynivalenol (DON). Toxins 2011, 3, 968–978. [Google Scholar] [CrossRef] [Green Version]
- Zachariasova, M.; Hajslova, J.; Kostelanska, M.; Poustka, J.; Krplova, A.; Cuhra, P.; Hochel, I. Deoxynivalenol and Its Conjugates in Beer: A Critical Assessment of Data Obtained by Enzyme-Linked Immunosorbent Assay and Liquid Chromatography Coupled to Tandem Mass Spectrometry. Anal. Chim. Acta 2008, 625, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Habler, K.; Rychlik, M. Multi-Mycotoxin Stable Isotope Dilution LC-MS/MS Method for Fusarium Toxins in Cereals. Anal. Bioanal. Chem. 2016, 408, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Habler, K.; Frank, O.; Rychlik, M. Chemical Synthesis of Deoxynivalenol-3-β-D-[13C6]-Glucoside and Application in Stable Isotope Dilution Assays. Molecules 2016, 21, 838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiby, I.; Sopel, M.M.; Michlmayr, H.; Adam, G.; Berthiller, F. Development and Validation of an Lc-Ms/Ms Based Method for the Determination of Deoxynivalenol and Its Modified Forms in Maize. Toxins 2021, 13, 600. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | DON | DON + LUT | DON + AC | DON + LUT + AC |
|---|---|---|---|---|---|
| ADFI (g/day) | 67.73 ± 0.77 a | 47.77 ± 1.29 d | 63.90 ± 2.34 c | 65.68 ± 1.17 b | 64.86 ± 0.64 b c |
| ADG (g/day) | 34.59 ± 0.88 a | 25.46 ± 0.32 c | 30.31 ± 0.64 b | 33.74 ± 0.28 a | 31.57 ± 1.01 b |
| FCR (feed: gain) | 1.45 ± 0.012 d | 1.63 ± 0.014 a | 1.56 ± 0.07 b | 1.48 ± 0.012 d | 1.52 ± 0.04 c |
| Parameters | Control | DON | DON + LUT | DON + AC | DON + LUT + AC |
|---|---|---|---|---|---|
| ALT (U/L) | 1.74 ± 0.01 b | 2.85 ± 0.075 a | 1.68 ± 0.15 c | 1.53 ± 0.20 e | 1.57 ± 0.28 d |
| AST (U/L) | 197.89 ± 1.09 d | 265.64 ± 32.22 c | 173.60 ± 1.04 e | 223.78 ± 1.04 a | 213.68 ± 0.77 b |
| ALP (U/L) | 1346.63 ± 55.39 b | 1874.98 ± 80.98 a | 1329.78 ± 90.32 b c | 1237.51 ± 56.37 c | 1298.35 ± 64.05 b c |
| TG (mmol/L) | 0.93 ± 0.08 b | 1.37 ± 0.01 a | 0.87 ± 0.07 c | 0.52 ± 0.07 e | 0.76 ± 0.07 d |
| TP (g/L) | 33.42 ± 0.74 a | 20.77 ± 1.95 d | 32.52 ± 1.40 a | 27.92 ± 0.52 c | 30.51 ± 0.77 b |
| Albumin (g/L) | 17.94 ± 0.93 a | 14.83 ± 0.47 b | 17.84 ± 0.68 a | 15.83 ± 0.59 b | 17.15 ± 0.17 a |
| Globulin (g/L) | 12.79 ± 0.58 a | 8.49 ± 0.43 d | 11.76 ± 0.73 a b | 10.467 ± 0.65 c | 11.26 ± 0.84 b c |
| Parameters | Control | DON | DON + LUT | DON + AC | DON + LUT + AC |
|---|---|---|---|---|---|
| MDA (nmol/mL) | 3.98 ± 0.24 b | 5.40 ± 0.52 a | 3.75 ± 0.23 bc | 3.25 ± 0.35 d | 3.57 ± 0.34 c |
| T-SOD (U/mL) | 142.11 ± 0.73 b | 105.58 ± 0.50 e | 140.67 ± 0.45 c | 132.28 ± 0.35 d | 148.43 ± 0.45 a |
| GSH-Px (U/mL) | 1420.05 ± 59.06 a | 894.37 ± 78.47 c | 1390.17 ± 125.35 a | 1586.67 ± 95.29 b | 1480.81 ± 85.48 b |
| CAT (U/mL) | 4.72 ± 0.18 a | 1.98 ± 0.19 c | 3.14 ± 0.21 a b | 5.96 ± 0.67 b | 4.02 ± 0.54 b |
| Parameters | Control | DON | DON + LUT | DON + AC | DON + LUT + AC |
|---|---|---|---|---|---|
| CAT (U/mgprot) | 82.76 ± 0.43 a | 38.73 ± 0.43 e | 78.63 ± 0.07 b | 69.43 ± 0.62 d | 75.83 ± 0.35 c |
| GSH-Px (U/mgprot) | 52.05 ± 0.28 a | 28.37 ± 0.330 e | 49.67 ± 0.47 b | 42.17 ± 0.51 d | 45.81 ± 0.27 c |
| T-SOD (U/mgprot) | 69.50 ± 0.33 a | 47.84 ± 0.51 e | 67.64 ± 0.47 b | 62.28 ± 0.56 d | 65.44 ± 0.31 c |
| MDA (nmol/mgprot) | 1.83 ± 0.02 b | 2.83 ± 0.29 a | 1.71 ± 0.06 b | 1.64 ± 0.35 b | 1.67 ± 0.38 b |
| Ingredient | Percentage (%) |
|---|---|
| Corn | 58.3 |
| Soybean meal | 30.2 |
| Fish meal | 5.6 |
| Soybean oil | 2.3 |
| Dicalcium phosphate | 1.2 |
| Limestone | 1.00 |
| Salt | 0.2 |
| Methionine | 0.2 |
| Premix 1 | 1.00 |
| Total | 100 |
| Calculated chemical composition | |
| Crude protein | 21.87 |
| Metabolizable energy (MJ/kg) | 13.45 |
| Lysine | 1.14 |
| Methionine | 0.40 |
| Methionine + Cystine | 0.94 |
| Calcium | 0.95 |
| Available phosphorus | 0.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.; Wang, Y.; Rajput, S.A.; Shaukat, A.; Yang, P.; Farooq, M.Z.; Cheng, Q.; Ali, M.; Mi, X.; An, Y.; et al. Ameliorative Effects of Luteolin and Activated Charcoal on Growth Performance, Immunity Function, and Antioxidant Capacity in Broiler Chickens Exposed to Deoxynivalenol. Toxins 2023, 15, 478. https://doi.org/10.3390/toxins15080478
Hassan M, Wang Y, Rajput SA, Shaukat A, Yang P, Farooq MZ, Cheng Q, Ali M, Mi X, An Y, et al. Ameliorative Effects of Luteolin and Activated Charcoal on Growth Performance, Immunity Function, and Antioxidant Capacity in Broiler Chickens Exposed to Deoxynivalenol. Toxins. 2023; 15(8):478. https://doi.org/10.3390/toxins15080478
Chicago/Turabian StyleHassan, Mubashar, Yanan Wang, Shahid Ali Rajput, Aftab Shaukat, Ping Yang, Muhammad Zahid Farooq, Qianhui Cheng, Mehboob Ali, Xiaomei Mi, Yu An, and et al. 2023. "Ameliorative Effects of Luteolin and Activated Charcoal on Growth Performance, Immunity Function, and Antioxidant Capacity in Broiler Chickens Exposed to Deoxynivalenol" Toxins 15, no. 8: 478. https://doi.org/10.3390/toxins15080478
APA StyleHassan, M., Wang, Y., Rajput, S. A., Shaukat, A., Yang, P., Farooq, M. Z., Cheng, Q., Ali, M., Mi, X., An, Y., & Qi, D. (2023). Ameliorative Effects of Luteolin and Activated Charcoal on Growth Performance, Immunity Function, and Antioxidant Capacity in Broiler Chickens Exposed to Deoxynivalenol. Toxins, 15(8), 478. https://doi.org/10.3390/toxins15080478