Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity


Abstract
:1. Introduction
2. Bacterial Metabolites
3. Fungal Metabolites
4. Plant Metabolites
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations(FAO). Future of Food and Agriculture-Trends and Challenges; Food and Agriculture Organization of the United Nations(FAO): Rome, Italy, 2017. [Google Scholar]
- Rosegrant, M.W.; Cline, S.A. Global food security: Challenges and policies. Science 2003, 302, 1917–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Caicedo, J.C.; Villamizar, S. Xanthomonas citri ssp. citri pathogenicity, a review. Citrus Res. Dev. Biotech. 2021, 135. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action mechanisms of effectors in plant-pathogen interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Lonigro, S.L.; Evidente, A.; Andolfi, A. Bacteriocin production by Pseudomonas syringae pv. ciccaronei NCPPB2355. Isolation and partial characterization of the antimicrobial compound. J. Appl. Microbiol. 1999, 86, 257–265. [Google Scholar]
- Evidente, A.; Abouzeid, A.M.; Andolfi, A.; Cimmino, A. Recent achievements in the bio-control of Orobanche infesting important crops in the Mediterranean basin. J. Agric. Sci. Technol. 2011, 1, 461–483. [Google Scholar]
- Pires, D.; Vicente, C.S.; Menéndez, E.; Faria, J.M.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The fight against plant-parasitic nematodes: Current status of bacterial and fungal biocontrol agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef]
- Lorenzen, K.; Anke, K. Basidiomycetes as a source for new bioactive natural products. In Current Organic Chemistry; Mori, K., Ed.; Bentham Science Publisher: Miami, FL, USA, 1998; pp. 329–364. [Google Scholar]
- Anke, H.; Sterner, O. Insecticidal and nematocidal metabolites from fungi. In The Mycota; Osiewacz, H.D., Ed.; Springer: Berlin, Germany, 2002; Volume 10, pp. 109–127. [Google Scholar]
- Osmanova, N.; Schultze, W.; Ayoub, N. Azaphilones: A class of fungal metabolites with diverse biological activities. Phytochem. Rev. 2010, 9, 315–342. [Google Scholar] [CrossRef]
- Shen, W.; Mao, H.; Huang, Q.; Dong, J. Benzenediol lactones: A class of fungal metabolites with diverse structural features and biological activities. Eur. J. Med. Chem. 2015, 97, 747–777. [Google Scholar] [CrossRef]
- Mao, Z.; Sun, W.; Fu, L.; Luo, H.; Lai, D.; Zhou, L. Natural dibenzo-α-pyrones and their bioactivities. Molecules 2014, 19, 5088–5108. [Google Scholar] [CrossRef] [PubMed]
- Ghisalberti, E.L. Secondary metabolites with antinematodal activity. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2002; Volume 26, pp. 425–506. [Google Scholar]
- D’addabbo, T.; Carbonara, T.; Leonetti, P.; Radicci, V.; Tava, A.; Avato, P. Control of plant parasitic nematodes with active saponins and biomass from Medicago sativa. Phytochem. Rev. 2011, 10, 503–519. [Google Scholar] [CrossRef]
- Hernández-Carlos, B.; Gamboa-Angulo, M. Insecticidal and nematicidal contributions of Mexican flora in the search for safer biopesticides. Molecules 2019, 24, 897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrar, M.; Ullah, M.W.; Manan, S.; Farooq, U.; Rafiq, M.; Hasan, F. Fungi from the extremes of life: An untapped treasure for bioactive compounds. Appl. Microbiol. Biotechnol. 2020, 104, 2777–2801. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Martínez-Gómez, Á.; Fenoll, C.; Escobar, C. The use of biochar for plant pathogen control. Phytopathology 2021, 111, 1490–1499. [Google Scholar] [CrossRef]
- Snoop, W.; Mrozik, H.; Fisher, M.H. Structure and activity of avermectins and milbemycins in animal health. Vet. Parsitol. 1995, 59, 139–156. [Google Scholar]
- Hoshino, T. Violacein and related tryptophan metabolites produced by Chromobacterium violaceum: Biosynthetic mechanism and pathway for construction of violacein core. Appl. Microbiol. Biotechnol. 2011, 91, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Konzen, M.; De Marco, D.; Cordova, C.A.; Vieira, T.O.; Antonio, R.V.; Creczynski-Pasa, T.B. Antioxidant properties of violacein: Possible relation on its biological function. Bioorg. Med. Chem. 2006, 14, 8307–8313. [Google Scholar] [CrossRef]
- Becker, M.H.; Brucker, R.M.; Schwantes, C.R.; Harris, R.N.; Minbiole, K.P. The bacterially produced metabolite violacein is associated with survival of amphibians infected with a lethal fungus. Appl. Environ. Microbiol. 2009, 75, 6635–6638. [Google Scholar] [CrossRef] [Green Version]
- Matz, C.; Webb, J.S.; Schupp, P.J.; Phang, S.Y.; Penesyan, A.; Egan, S.; Steinberg, P.; Kjelleberg, S. Marine biofilm bacteria evade eukaryotic predation by targeted chemical defense. PLoS ONE 2008, 3, e2744. [Google Scholar] [CrossRef] [Green Version]
- Ballestriero, F.; Daim, M.; Penesyan, A.; Nappi, J.; Schleheck, D.; Bazzicalupo, P.; Di Schiavi, E.; Egan, S. Antinematode activity of violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans. PLoS ONE 2014, 9, e109201. [Google Scholar] [CrossRef]
- Zeng, L.; Jin, H.; Lu, D.; Yang, X.; Pan, L.; Cui, H.; He, X.; Qiu, H.; Qin, B. Isolation and identification of chemical constituents from the bacterium Bacillus sp. and their nematicidal activities. J. Basic Microbiol. 2015, 55, 1239–1244. [Google Scholar] [CrossRef]
- Kosaka, H.; Aikawa, T.; Ogura, N.; Tabata, K.; Kiyohara, T. Pine wilt disease caused by the pine wood nematode: The induced resistance of pine trees by the avirulent isolates of nematode. Eur. J. Plant Pathol. 2001, 107, 667–675. [Google Scholar] [CrossRef]
- Song, M.J.; Bae, J.; Lee, D.S.; Kim, C.H.; Kim, J.S.; Kim, S.W.; Hong, S.I. Purification and characterization of prodigiosin produced by integrated bioreactor from Serratia sp. KH-95. JBB 2006, 101, 157–161. [Google Scholar]
- Rahul, S.; Chandrashekhar, P.; Hemant, B.; Chandrakant, N.; Laxmikant, S.; Satish, P. Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Zeng, Z.; Xue, B.; Deng, Y.; Sun, M.; Tang, Y.-J.; Ruana, L. Bacillus thuringiensis produces the lipopeptide thumolycin to antagonize microbes and nematodes. Microbiol. Res. 2018, 215, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Cabras, A.; Mannoni, M.A.; Serra, S.; Andolfi, A.; Fiore, M.; Evidente, A. Occurrence, isolation and biological activity of phytotoxic metabolites produced in vitro by Sphaeropsis sapinea, pathogenic fungus of Pinus radiata. Eur. J. Plant Pathol. 2006, 115, 187–193. [Google Scholar] [CrossRef]
- Kang, M.K.; Kim, J.H.; Liu, M.J.; Jin, C.Z.; Park, D.; Kim, J.; Sung, B.-H.; Kim, C.J.; Son, K.-H. New discovery on the nematode activity of aureothin and alloaureothin isolated from endophytic bacteria Streptomyces sp. AE170020. Sci. Rep. 2022, 12, 3947. [Google Scholar] [CrossRef]
- Anke, H.; Stadler, M.; Mayer, A.; Sterner, O. Secondary metabolites with nematicidal and antimicrobial activity from nematophagous fungi and Ascomycetes. Can. J. Bot. 1995, 73, 932–939. [Google Scholar] [CrossRef]
- Dasenbrock, J. Isolierung und Strukturautlclarung neuer Wirkstoffe aus Hijheren Pilzen. Ph.D. Thesis, University of Bonn, Bonn, Germany, 1994. [Google Scholar]
- Lorenzen, K.; Anke, T.; Anders, U.; Hindermayr, H.; Hansske, F. 14-Epidihydrocochlioquinone B, and 14-epicochlioquinone B, antibiotics from fermentations of the Ascomycete Neobulgaria pura: Structure elucidation and effects on platelet aggregation. Z. Naturforsch. 1994, 49C, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, J.M.; Frazier, E.G.; Bergstrom, A.R.; Williamson, J.M.; Liesch, J.M.; Goetz, M.A. Cochlioquinone A, a nematocidal agent which competes for specific [3H]ivermectin binding sites. J. Antibiot. 1990, 43, 1179–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, W.C. Ivermectin: An update. Parasitol. Today 1985, 1, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Wada, K.; Munakata, K. New nematicidal metabolites from a fungus, Irpex lacteus. Agric. Biol. Chem. 1981, 45, 1527–1529. [Google Scholar]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. VII. Structure determination of brominated lachnumon and mycorrhizin A derivatives. J. Antibiot. 1995, 48, 158–161. [Google Scholar]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. 111. Production of novel isocoumarin derivatives. Isolation and biological activities. J. Antibiot. 1995, 48, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceurn (Karst.) Karst. IV. Structural elucidation of novel isocoumarin derivatives. J. Antibiot. 1995, 48, 267–270. [Google Scholar]
- Chexal, K.K.; Tamm, C. Biosynthesis of mikrolin. Helv. Chim. Acta 1978, 61, 2002–2018. [Google Scholar] [CrossRef]
- Chexal, K.K.; Tamm, C.; Clardy, J.; Hirotsu, K. Gilmicolin and mycorrhizinol, two new metabolites of Gilmaniella humicola Barron. Helv. Chim. Acta 1979, 62, 1130–1142. [Google Scholar] [CrossRef]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. V. Production, isolation and biological activities of bromine-containing mycorrhizin and lachnumon derivatives and four additional new bioactive metabolites. J. Antibiot. 1995, 48, 149–153. [Google Scholar]
- Stadler, M.; Anke, H.; Sterner, O. Metabolites with nematicidal and antimicrobial activities from the Ascomycete Lachnum papyraceum (Karst.) Karst. VI. Structure determination of non-halogenated metabolites structurally related to mycorrhizin A. J. Antibiot. 1995, 48, 154–157. [Google Scholar]
- Mayer, A.; Anke, H.; Sterner, O. Omphalotin, a new cyclic peptide with potent nematicidal activity from Omphalotus olearius I. Fermentation and biological activity. Nat. Prod. Lett. 1997, 10, 25–32. [Google Scholar] [CrossRef]
- Kohno, J.; Nishio, M.; Sukarai, M.; Kawano, K.; Hiramatsu, H.; Kameda, N.; Kish, N.; Yamashita, T.; Okuda, T.; Komatsubara, S. Isolation and structure determination of TMC-151s: Novel polyketide antibiotics from Gliocladium catenulatum Gilman & Abbott TC 1280. Tetrahedron 1999, 55, 7771–7786. [Google Scholar]
- Kohno, J.; Nishio, M.; Kish, M.N.; Komatsubara, S. Biosynthesis of the fungal polyketide antibiotics TMC151s: Origin of the carbon skeleton. J. Antibiot. 2000, 53, 1301–1304. [Google Scholar] [CrossRef] [Green Version]
- Kohno, J.; Asai, Y.; Nishio, M.; Sakurai, M.; Kawano, K.; Hiramatsu, H.; Kameda, N.; Kishi, N.; Okuda, T.; Komatsubara, S. TMC-171A, B, C and TMC-154, novel polyketide antibiotics produced by Gliocladium sp. TC 1304 and TC 1282. J. Antibiot. 1999, 52, 1114–1123. [Google Scholar]
- Omura, S.; Tomoda, H.; Tabata, N.; Ohyama, Y.; Abe, T.; Namikoshi, M. Roselipins, novel fungal metabolites having a highly methylated fatty acid modified with a monnose and an arabinitol. J. Antibiot. 1999, 52, 586–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomoda, H.; Ohyama, Y.; Abe, T.; Tabata, N.; Namikoshi, M.; Yamaguchi, Y.; Masuma, R.; Omura, S. Roselipins, inhibitors of diacylglycerol acyltransferase, produced by Gliocladium roseum KF-1040. J. Antibiot. 1999, 52, 689–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabata, N.; Ohyama, Y.; Tomoda, H.; Abe, T.; Namikoshi, M.; Omura, S. Structure elucidation of roselipins, inhibitors of diacylglycerol acyltransferase, produced by Gliocladium roseum KF-1040. J. Antibiot. 1999, 52, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumazawa, S.; Kanda, M.; Utagawa, M.; Chiba, N.; Ohtani, H.; Mikawa, T. MK7924, a novel metabolite with nematocidal activity from Coronophora gregaria. J. Antibiot. 2003, 56, 652–654. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Dong, J.; Wang, L.; Zhou, W.; Li, L.; He, H.; Liu, H.; Zhang, K. Screening and isolation of antinematodal metabolites against Bursaphelenchus xylophilus produced by fungi. Ann. Microbiol. 2008, 58, 375–380. [Google Scholar] [CrossRef]
- Mamiya, Y. The pine wood nematode. In Plant and Insect Nematodes; Nickle, W.R., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1984. [Google Scholar]
- Sutherland, J.R.; Webster, J.M. Nematode pests of forest trees. In Plant-Parasitic Nematodes in Temperate Agriculture; Evans, K., Trudgill, D.L., Webster, J.M., Eds.; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Ferris, H.; Zheng, L. Plant sources of Chinese Herbal Remedies: Effectis on Pratylenchus vulnus and Meloidogyne javanica. J. Nematol. 1999, 31, 241–263. [Google Scholar] [PubMed]
- Dong, J.Y.; Song, H.C.; Li, J.H.; Tang, Y.S.; Sun, R.; Wang, L.; Zhou, Y.P.; Wang, M.L.; Shen, K.Z.; Wang, C.R.; et al. Ymf 1029A-E, preussomerin analogues from the fresh-water-derived fungus YMF 1.01029. J. Nat. Prod. 2008, 71, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.S.; Niu, X.M.; Wang, Y.L.; Guo, J.P.; Pan, W.Z.; Huang, X.-W.; Zhang, K.-Q. Isolation of putative biosynthetic intermediates of prenylated indole alkaloids from a thermophilic fungus Talaromyces thermophilus. Org. Lett. 2010, 12, 4356–4359. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.P.; Zhu, C.Y.; Zhang, C.P.; Chu, Y.S.; Wang, Y.L.; Zhang, J.-X.; Wu, D.-K.; Zhang, K.-Q.; Niu, X.-M. Thermolides, potent nematocidal PKS-NRPS hybrid metabolites from thermophilic fungus Talaromyces thermophilus. J. Am. Chem. Soc. 2012, 134, 20306–20309. [Google Scholar] [CrossRef]
- Zhang, J.M.; Wang, H.H.; Liu, X.; Hu, C.H.; Zou, Y. Heterologous and engineered biosynthesis of nematocidal polyketide-nonribosomal peptide hybrid macrolactone from extreme thermophilic fungi. J. Am. Chem. Soc. 2020, 142, 1957–1965. [Google Scholar] [CrossRef]
- Meng, X.; Mao, Z.; Lou, J.; Xu, L.; Zhong, L.; Peng, Y.; Zhou, L.; Wang, M. Benzopyranones from the endophytic fungus Hyalodendriella sp. Ponipodef12 and their bioactivities. Molecules 2012, 17, 11303–11314. [Google Scholar] [CrossRef]
- Liu, T.; Meyer, S.L.; Chitwood, D.J.; Chauhan, K.R.; Dong, D.; Zhang, T.; Li, J.; Liu, W.C. New nematotoxic indoloditerpenoid produced by Gymnoascus reessii za-130. J. Agric. Food Chem. 2017, 65, 3127–3132. [Google Scholar]
- Ashrafi, S.; Helaly, S.; Schroers, H.J.; Stadler, M.; Richert-Poeggeler, K.R.; Dababat, A.A.; Maier, W. Ijuhya vitellina sp. nov., a novel source for chaetoglobosin A, is a destructive parasite of the cereal cyst nematode Heterodera filipjevi. PLoS ONE 2017, 12, e0180032. [Google Scholar] [CrossRef]
- Kim, T.Y.; Jang, J.Y.; Yu, N.H.; Chi, W.J.; Bae, C.H. Nematicidal activity of grammicin produced by Xylaria grammica KCTC 13121BP against Meloidogyne incognita. Pest Manag. Sci. 2018, 74, 384–391. [Google Scholar] [CrossRef]
- Edwards, R.L.; Maitland, D.J.; Pittayakhajonwut, A.J.S.; Whalley, J. Metabolites of the higher fungi. Part 33. Grammicin, a novel bicyclic C7H6O4 furanopyranol from the fungus Xylaria grammica (Mont.) Fr. J. Chem. Soc. Perkin Trans. 2001, 1, 1296–1299. [Google Scholar] [CrossRef]
- Birkinshaw, J.H.; Michael, S.E.; Bracken, A.; Raistrick, H. Patulin in the common cold collaborative research on a derivative of Penicillium patulum Bainier. II. Biochem. Chem. Lancet 1943, 245, 625. [Google Scholar]
- Moake, M.M.; Padilla-Zakour, O.I.; Worobo, R.W. Comprehensive review of patulin control methods in foods. Compr. Rev. Food Sci. Food Saf. 2005, 1, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Puel, O.; Galtie, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phukhamsakda, C.; Ariyawansa, H.A.; Phillips, A.J.; Wanasinghe, D.N.; Bhat, D.J.; McKenzie, E.H.C.; Camporesi, E.; Hide, K.D. Additions to Sporormiaceae: Introducing two novel genera, Sparticola and Forliomyces, from Spartium. Cryptogam. Mycol. 2016, 37, 75–97. [Google Scholar]
- Phukhamsakda, C.; Macabeo, A.P.G.; Huch, V.; Cheng, T.; Hyde, K.D. Stadler, M. Sparticolins A-G, biologically active oxidized spirodioxynaphthalene derivatives from the ascomycete. J. Nat. Prod. 2019, 82, 2878–2885. [Google Scholar] [CrossRef]
- Du, F.Y.; Ju, G.L.; Xiao, L.; Zhou, Y.M.; Wu, X. Sesquiterpenes and cyclodepsipeptides from marine-derived fungus Trichoderma longibrachiatum and their antagonistic activities against soil-borne pathogens. Mar. Drugs 2020, 18, 165. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.A.; Estevez-Braun, A.; Estevez-Reyes, R.; Bazzocchi, I.L.; Moujir, L.; Jimenez, I.A.; Ravelo, A.G.; Gonzalez, A.G. Biological activity of secondary metabolites from Bupleurum salicifolium (Umbelliferae). Experientia 1995, 51, 35–39. [Google Scholar]
- Bramwell, D.; BramwelI, Z. Wíld Flowers of the Canary Islands; Cabíldo Insular de Tenerífe: Santa Cruz de Tenerife, Spain, 1974. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons: Chicester, UK, 2002. [Google Scholar]
- Atkinson, H.J.; Taylor, J.D. Evidence for a calcium-binding site on the eggshell of Globodera rostochiensis with a role in hatching. Ann. Appl. Biol. 1980, 96, 307–315. [Google Scholar] [CrossRef]
- Atkinson, H.J.; Taylor, J.D. A calcium-binding sialoglycoprotein associated with an apparent eggshell membrane of Globodera rostochiensis. Ann. Appl. Biol. 1983, 102, 345–354. [Google Scholar] [CrossRef]
- Gonzalez, J.A.; Estevez-Braun, A. Phytonematicidal activity of aromatic compounds related to shikimate pathway. Pestic. Biochem. Phys. 1997, 58, 193–197. [Google Scholar] [CrossRef]
- Zasada, I.A.; Meyer, S.L.F.; Halbrendt, J.M.; Rice, C. Activity of hydroxamic acids from Secale cereale against the plant-parasitic nematodes Meloidogyne incognita and Xiphinema americanum. Phytopathology 2005, 95, 1116–1121. [Google Scholar] [CrossRef] [Green Version]
- Razavi, S.M. Plant counnarins as allelopathic agents. Int. J. Biol. Chem. 2011, 5, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Jin, H.; Liu, Q.; Yan, Z.; Ding, L.; Qin, B. Nematicidal metabolites from roots of Stellera chamaejasme against Bursaphelenchus xylophilus and Bursaphelenchus mucronatus. Pest Manag. Sci. 2014, 70, 827–835. [Google Scholar] [CrossRef]
- Kumarasingha, R.; Karpe, A.V.; Preston, S.; Yeo, T.C.; Lim, D.S.; Tu, C.L.; Kavlene, J.L.; Simpson, J.; Gasser, R.B.; Beale, P.D.; et al. Metabolic profiling and in vitro assessment of anthelmintic fractions of Picria fel-terrae Lour. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 171–178. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, G.; Huang, X.; Wang, Z.; Tan, N. 1, 4-naphthoquinone triggers nematode lethality by inducing oxidative stress and activating insulin/IGF signaling pathway in Caenorhabditis elegans. Molecules 2017, 22, 798. [Google Scholar] [CrossRef] [Green Version]
- Fang, N.; Casida, J.E. Anticancer action of cube’ insecticide: Correlation for rotenoid constituents between inhibition of NADH:ubiquinone oxidoreductase and induced ornithine decarboxylase activities. Proc. Natl. Acad. Sci. USA 1998, 95, 3380–3384. [Google Scholar] [CrossRef] [Green Version]
- Cabizza, M.; Angioni, A.; Melis, M.; Cabras, M.; Tuberoso, C.V.; Cabras, P. Rotenone and rotenoids in cub’e resins, formulations, and residues on olives. J. Agric. Food Chem. 2004, 52, 288–293. [Google Scholar] [CrossRef]
- Wenjie, J.; Fang, Y.; Gan, C.; Wu, Y.; Pang, J. Extraction and purification of deguelin from Derris trifoliata Lour root. Int. J. Agric. Biol. Eng. 2009, 2, 98–103. [Google Scholar]
- Chen, C.S.; Ho, D.R.; Chen, F.Y.; Chen, C.R.; Ke, Y.D.; Su, J.G.J. AKT mediates actinomycin D-induced p53 expression. Oncotarget 2014, 5, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Vats, S.; Kamal, R. Cassia occidentalis L. (a new source of rotenoids): Itsin vitro regulation by feeding precursors and larvicidal efficacy. Plant Cell Tissue Organ Cult. 2014, 116, 403–409. [Google Scholar] [CrossRef]
- Ashack, R.J.; McCarty, L.P.; Malek, R.S.; Goodman, F.R.; Peet, N.P. Evaluation of rotenone and related compounds as antagonists of slow-reacting substance of anaphylaxis. J. Med. Chem. 1980, 23, 1022–1026. [Google Scholar] [CrossRef]
- Okombe Embeya, V.; Lumbu Simbi, J.B.; Stevigny, C.; Vandenput, S.; Pongombo Shongo, C.; Duez, P. Traditional plantbased remedies to control gastrointestinal disorders in livestock in the regions of Kamina and Kaniama (Katanga province, DemocraticRepublic of Congo. J. Ethnopharmacol. 2014, 153, 686–693. [Google Scholar] [CrossRef]
- Preston, S.; Korhonen, P.K.; Mouchiroud, L.; Cornaglia, M.; McGee, S.L.; Young, N.D.; Davis, R.A.; Crawford, S.; Nowell, C.; Ansell, B.R.E.; et al. Deguelin exerts potent nematocidal activity via the mitochondrial respiratory chain. FASEB J. 2017, 31, 4515–4532. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Chávez, J.L.; Rufino-González, Y.; Ponce-Macotela, M.; Delgado, G. In vitro activity of “Mexican Arnica” Heterotheca inuloides Cass natural products and some derivatives against Giardia intestinalis. Parasitology 2015, 142, 576–584. [Google Scholar] [CrossRef]
- Rodríguez-Chávez, J.L.; Coballase-Urrutia, E.; Sicilia-Argumedo, G.; Ramírez-Apan, T.; Delgado, G. Toxicological evaluation of thenatural products and some semisynthetic derivatives of Heterotheca inuloides Cass (Asteraceae). J. Ethnopharmacol. 2015, 175, 256–265. [Google Scholar] [CrossRef]
- Rodríguez-Chávez, J.L.; Franco-Navarro, F.; Delgado, G. In vitro nematicidal activity of natural and semisynthetic cadinenes from Heterotheca inuloides against the plant-parasitic nematode Nacobbus aberrans (Tylenchida: Pratylenchidae). Pest Manag. Sci. 2019, 75, 1734–1742. [Google Scholar] [CrossRef]
- Jang, J.Y.; Le Dang, Q.; Choi, G.J.; Park, H.W.; Kim, J.C. Control of root-knot nematodes using Waltheria indica producing 4-quinolone alkaloids. Pest Manag. Sci. 2019, 75, 2264–2270. [Google Scholar]
- Ismail, M.; Kowsar, A.; Javed, S.; Choudhary, M.I.; Khan, S.W.; Abbas, Q.; Tang, Y.; Wang, W. The antibacterial, insecticidal and nematocidal cctivities and toxicity studies of Tanacetum falconeri Hook. f. Turk. J. Pharm. Sci. 2021, 18, 744. [Google Scholar] [CrossRef]
- Velasco-Azorsa, R.; Zeferino-Díaz, R.; Alvarado-Rodríguez, J.G.; López-Ruiz, H.; Rojas-Lima, S.; Flores-Castro, K.; Cid del Prado-Vera, I.; Alatorre-Rosas, R.; Tut-Pech, F.; Carrillo-Benítez, M.G.; et al. Nematicidal activity of furanoeremophilenes against Meloidogyne incognita and Nacobbus aberrans. Pest Manag. Sci. 2022, 78, 2571–2580. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Source | Biological Activity | References |
|---|---|---|---|
| Metabolites Produced by Bacteria | |||
| Avermectin B1a (1) | Streptomyces avermentilis | Nematocidal activity against Haemonchus contortus, Ostertagia circumcincta; Trichostrongylus axei; Trichostrongylus colubriformis; Cooperia spp.; Oesophagostomum columbianu, (LD50 values in mice approximatively ranking between 15 and 50 mg/kg) and Meloidogyne incognita or Rotylenchulus reniformis (LD50/2 h 1.56 and 32.9 µg/mL , respectively) | [22] |
| milbemycin D (2) | Streptomyces hygroscopicus and Streptomyces cyaneogriseus | Nematocidal activity against Caenorhabditis elegans (EC50 89 nM) | [22] |
| Violacein (3) | Chromobacterium violaceum, Janthinobacterium lividum, and Pseudoalteromonas tunicata D2 Collimonas sp., Duganella sp. and Pseudoalteromonas spp. | Antibacterial, antitrypanocidal, anti-ulcerogenic, anticancer, antioxidant, leishmanicidal, antifungal, and antiviral | [24] [27] |
| Oxidative stress resistance, | [24] | ||
| defense of amphibians from fungal disease | [25] | ||
| Pseudoalteromonas tunicate | Antipredator defense mechanism against protozoan grazers | [26] | |
| Microbulbifer sp. D250 | Nematocidal activity against C. elegans (LC50 > 30 nM) | [27] | |
| 4-Oxabicyclo[3.2.2]nona-1(7), 5,8-triene (4) | Bacillus strain SMrs28 | Nematocidal activity against B. sxylophilus and D. destructor (LD50 904.12 and 1594.0 μg/mL/72 h, respectively) | [28] |
| (3S,8aS)-Hexahydro-3-methylpyrro[1,2-a]pyrazine-1,4-dione (5) | “ | Nematocidal activity against B. sxylophilus and D. destructor (LD50 451.26 and 366.62 μg/mL/72 h, respectively) | “ |
| Phenylacetamide (6) | “ | Nematocidal activity against B. sxylophilus and D. destructor (LD50 232.98 and 206.38 μg/mL/72 h, respectively) “ | “ |
| Cyclo(L-Pro-L-Val) (7) | “ | No tested | “ |
| Lauric acid (8) | “ | “ | “ |
| Methyl elaidate (9) | “ | “ | “ |
| Prodigiosin (10) | Serratia marcescens | Nematocidal activity against R. similis and M. javanica (LC50 83 and 79 μg/mL, respectively) | [31] |
| Thumolycin (11) | Bacillus thuringiensis | Nematocidal activity against C. elegans (600 U) | [32] |
| Aureothin (12) | Streptomyces sp. AE170020 | Nematocidal activity against B. xylophilus J2s, J3s and J4s (LC50 0.81, 1.15 and 1.54 μg/ L) | [34] |
| Alloaureothin (13) | “ | Nematocidal activity against B. xylophilus J2s, J3s and J4s (LC50 0.83, 1.10 and 1.47 μg/ L) | “ |
| Metabolites produced by fungi | |||
| 1-Methoxy-8-hydroxynaphthalene (14), | Daldinia concentrica | Cytotoxic, antibiotic, and nematocidal activity against C. elegans (LD50 10 μg/mL) | [35] [36] |
| 1,8-Dimethoxynaphthalene (15) | “ | Cytotoxic, antibiotic, and nematocidal activity against C. elegans (LD50 25 μg/mL) | “ |
| 5-Hydroxy-2-methylchromanone (16) | “ | Nematocidal activity C. elegans (LD50 > 100 μg/mL) | [36] |
| 14-epi-Dihydrocochlioquinone B (17) | Neobulgaria pura | Nematocidal against C. elegans (LD50 10 μg/mL) and M. incognita (LD30 100 μg/mL) | [35,37] |
| 14-epi-Cochlioquinone B (18) | “ | Nematocidal activity against C. elegans and M. incognita | “ |
| Cochlioquinone A (19) | Helminthosporium sp. | “ | [38] |
| Ivermectin (20) | Streptomyces avermitilis | “ | [39] |
| 5-Pentyl-2-furaldehyde (21) | Dermateaceae sp. Irpex lacteus | Nematocidal activity against C. elegans (LD50 60 μg/mL) and M. incognita (LD50 75 μg/mL) | [40] |
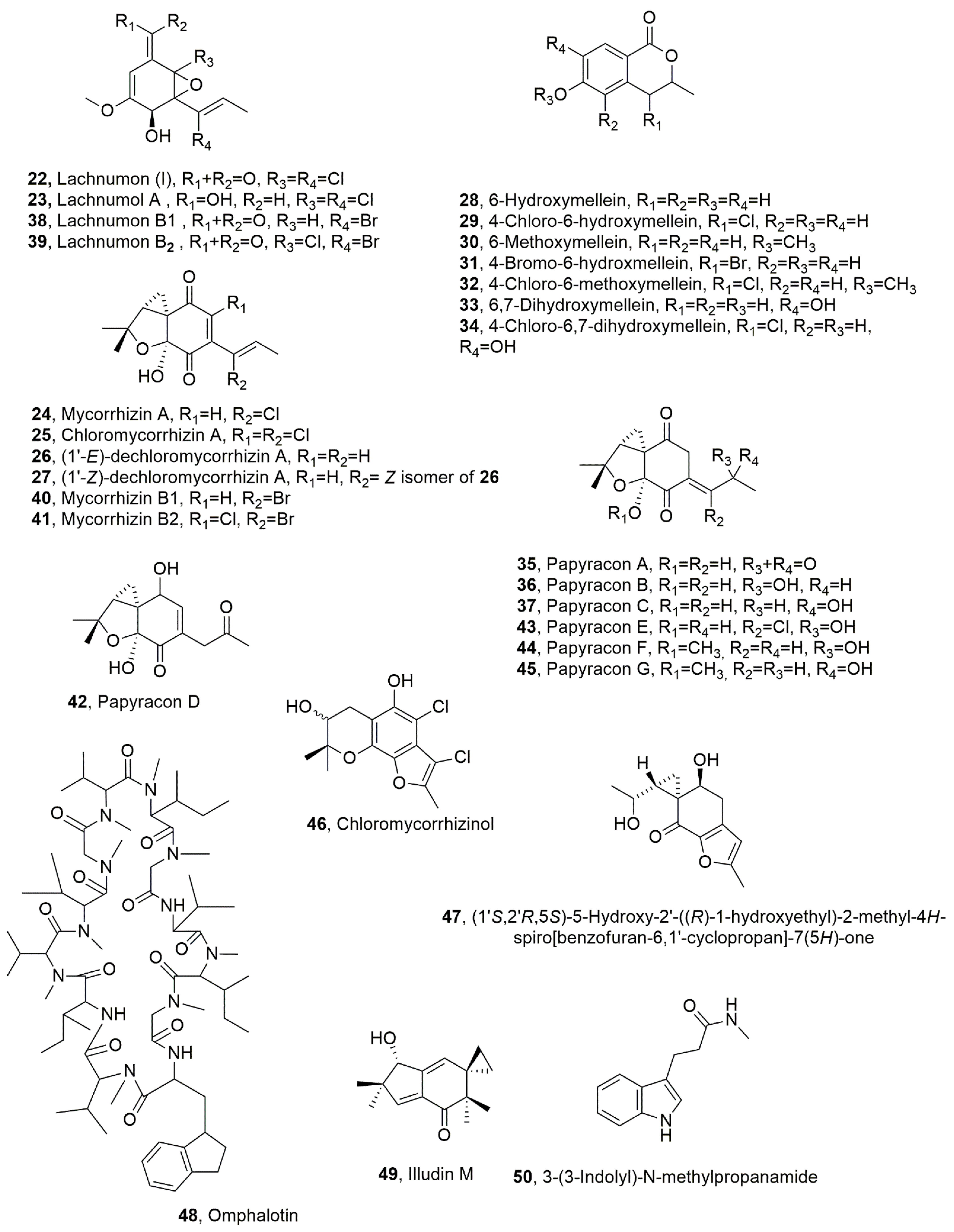
| Lachnumon (I) (22) | Lachnum papyraceum | Nematocidal activity against C. elegans (LD50 25 μg/mL) and M. incognita (LD50 > 100μg/mL) | [41,42] |
| Lachnumol A (23) | “ | Nematocidal activity against C. elegans (LD50 5 μg/mL) and M.incognita (LD50 100 μg/mL) | “ |
| Mycorrhizin A (24), | “ | Nematocidal activity against C. elegans (LD50 1 μg/mL) and M. incognita (LD50 100 μg/mL) | “ |
| Chloromycorrhizin A (25) | “ | Nematocidal activity against C. elegans (LD50 100 μg/mL) and M. incognita (LD50 100 μg/mL) | “ |
| (1’-E)-dechloromycorrhizin A (26) | “ | Nematocidal activity against C. elegans (LD50 2 μg/mL) and M. incognita (LD50 > 100μg/mL) | “ |
| (1’-Z)-dechloromycorrhizin A (27) | “ | Nematocidal activity against C. elegans (LD50 2 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| 6-Hydroxymellein (28) | “ | No nematocidal activity | [35] |
| 4-Chloro-6-hydroxymellein (29) | “ | “ | “ |
| 6-Methoxymellein (30) | “ | “ | “ |
| 4-Bromo-6-hydroxmellein (31) | “ | “ | “ |
| 4-Chloro-6-methoxymellein (23) | “ | “ | “ |
| 6,7-dihydroxymellein (33) | “ | ” | “ |
| 4-Chloro-6,7-dihydroxymellein (34) | “ | Nematocidal activity against C. elegans | “ |
| Papyracon A (35) | “ | Moderate nematocidal activity against C. elegans (LD50 25 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon B (36) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon C (37) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Lachnumon B1 (38) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Lachnumon B2 (39) | “ | Moderate nematocidal activity against C. elegans (LD50 25 μg/mL) and M. incognita (Not tested) | “ |
| Mycorrhizin B1 (40) | “ | Moderate nematocidal activity against C. elegans (LD50 2 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Mycorrhizin B2 (41) | “ | Moderate nematocidal activity against C. elegans (LD50 5 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon D (42) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon E (43) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon F (44) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Papyracon G (45) | “ | Moderate nematocidal activity against C. elegans (LD50 50 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| Chloromycorrhizinol (46) | “ | Nematocidal activity against C. elegans (LD50 100 μg/mL) and M. incognita (LD50 > 100 μg/mL) | “ |
| (1’S,2’R,5S)-5-Hydroxy-2’-((R)-1-hydroxyethyl)-2-methyl-4H-spiro[benzofuran-6,1’-cyclopropan]-7(5H)-one (47) | “ | Not tested | “ |
| Omphalotin (48) | Omphalotus olearius | Nematocidal activity against C. elegans (LD50 0.57 μg/mL) and M. incognita (LD50 18.95 μg/mL) | [48]. |
| Illudin M the (49) | “ | No activity | “ |
| 3-(3-Indolyl)-N-methylpropanamide (50) | “ | “ | “ |
| MK7924 (51) | Coronophora gregaria | Nematocidal activity against C. elegans at 100 μg/mL | [55] |
| 4,8-Dihydroxy-3,4-dihydronaphthalen-1(2H)-one (52), | Caryospora callicarpa | Nematocidal activity against B. xylophilus (LD50 540.2, 436.6 and 209.0 at 12, 24, and 36 h, respectively) | [56] |
| 4,5-Dihydroxy-3,4-dihydronaphthalen-1(2H)-one (53) | “ | Nematocidal activity against B. xylophilus (LD50 1169.8, 461.3, and 229.6 at 12, 24, and 36 h) | “ |
| 4,6,8-Trihydroxy-3,4-dihydronaphthalen-1(2H)-one (54) | “ | Nematocidal activity against B. xylophilus (LD50 1011.6, 522.5, and 220.3 at 12, 24, and 36 h) | “ |
| 3,4,6,8-Tetrahydroxy-3,4-dihydronaphthalen-1(2H)-one (55) | “ | Nematocidal activity against B. xylophilus (LD50 854, 468, and 206.1 at 12, 24 and 36 h) | “ |
| Ymf 1029 A (56) | Unidentified freshwater fungus YMF 1.01029 | Nematocidal activity against B. xylophilus | [60] |
| Ymf 1029 B (57) | “ | “ | “ |
| Ymf 1029 C (58) | “ | “ | “ |
| Ymf 1029 D (59) | “ | “ | “ |
| Ymf 1029 E (60) | “ | “ | “ |
| Preussomerin C (61) | “ | “ | “ |
| Preussomerin D (62) | “ | “ | “ |
| (4RS)-4,8-dihydroxy-3,4-dihydronaphthalen-1(2H)-one (63) | “ | “ | “ |
| 4,6,8-Trihydroxy-3,4-dihydronaphthalen-1(2H)-one (64) | “ | “ | “ |
| Thermolide A (65) | Talaromyces thermophilus | Nematocidal activity against M. incognita, B. siylopilus, and P. redivevus | [62] |
| Thermolide B (66) | “ | “ | “ |
| Thermolide C (67) | “ | “ | “ |
| Thermolide D (68) | “ | “ | “ |
| Thermolide E (69) | “ | Not tested | “ |
| Thermolide F (70) | “ | “ | “ |
| Palmariol B (71) | Hyalodendriella sp. | Antimicrobial, inhibition of acetylcholinesterase, and nematocidal activity against C. elegans (IC50 56.21 μg/mL) | [64] |
| 4-Hydroxymellein (72) | “ | Antibacterial and nematocidal activity against C. elegans (IC50 86.86 μg/mL) | “ |
| Alternariol 9-methyl ether (73) | Nematocidal activity against C. elegans (IC50 93.99 μg/mL) | “ | |
| Botrallin (74) | “ | Nematocidal activity against C. elegans (IC50 84.51 μg/mL) | “ |
| Gymnoascole acetate (75)) | Gymnoascus reessii za-130 | Nematocidal activity against M. incognita J2s (EC50 47.5 μg/mL) | [65] |
| Chaetoglobosin A (76) | Ijuhya vitellina | Nematocidal activity against H. filipjevi (at 50 μg/mL, it paralyzed the nematode) | [66] |
| 19-O-Acetylchaetoglobosin A (77) | “ | Nematocidal activity against H. filipjevi (at 100 μg/mL, it paralyzed the nematode) | “ |
| Grammicin (78) | Xylaria grammica | Low antibacterial and cytotoxic activity, but nematocidal activity against M. incognita J2 juvenile mortality and egg-hatching inhibition (EC50 15.95 and 5.87 μg/mL) | [67] |
| Patulin (79) | Penicillium griseofulvum and Penicillium expansum, and other fungal genera | Antibacterial, cytotoxic, and dematocidal activity against M. incognita J2 juvenile mortality and eggs-hatching inhibition (EC50/72 h 115.67 μg/mL) | [67,69,70] |
| Sparticolin A (80) | Sparticola junci | Nematocidal activity against C. elegans (LD50 50 μg/mL) | [73] |
| Sparticolin B (81) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) | “ |
| Sparticolin C (82) | “ | Nematocidal activity against C. elegans (LD50 25 μg/mL) | “ |
| Sparticolin D (83) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) | “ |
| Sparticolin E (84) | “ | Nematocidal activity against C. elegans (LD50 50 μg/mL) | “ |
| Sparticolin F (85) | “ | Nematocidal activity against C. elegans (LD50 12.5 μg/mL) | “ |
| Sparticolin G (86) | “ | Antifungal and cytotoxic activity, but not tested for nematocidal activity | “ |
| Trichodermene (87) | Trichoderma longibrachiatum | Antifungal activity against Colletotrichum lagenarium | [74] |
| Sesquiterpene (88) | “ | “ | “ |
| Sesquiterpene (89) | “ | “ | “ |
| Sesquiterpene (90) | “ | No activity | “ |
| Sesquiterpene (91) | “ | No activity | “ |
| Sesquiterpene (92) | “ | No activity | “ |
| Cyclodepsipeptide (93) | “ | Nematocidal against M. incognita (IC50 149.2 μg/mL) | “ |
| Cyclodepsipeptide (94) | “ | Nematocidal against M. incognita (IC50 140.6 μg/mL) | “ |
| Cyclodepsipeptide (95) | “ | Nematocidal against M. incognita (IC50 198.7 μg/mL) | “ |
| Metabolites produced by plants | |||
| Guayarol (96) | Bupleurum salicifolium | Nematocidal activity against G. pallida and G. rostochiensis (LC50 range 2 × 10−6 to 1.26 × 10−3) | [80] |
| Buplerol (97) | “ | “ | “ |
| Matairesinol (98) | “ | Nematocidal activity against G. pallida and G. rostochiensis (70% hatching reduction) | “ |
| Matairesinoldimethyl ether (99) | “ | Nematocidal activity against G. pallida and G. rostochiensis (LC50 range 2 × 10−6 to 1.26 × 10−3) | “ |
| Bursehernin (100) | “ | Nematocidal activity against G. pallida and G. rostochiensis (550% hatching reduction; HID 16.42) | “ |
| Pliviatolide (101) | “ | Nematocidal activity against G. pallida and G. rostochiensis (LC50 range 2 × 10−6 to 1.26 × 10−3) | “ |
| Thujaplicatin, methyl ether (102) | “ | “ | “ |
| 2-Chloro-matairesinol (103) | “ | “ | “ |
| Nortrachelogenin (104) | “ | “ | “ |
| Nortrachelogenin triacetate (105) | “ | “ | “ |
| 2-Hydroxythujaplican methyl ether (106) | “ | “ | |
| DIBOA (107) | Plant of Poaceae family | Nematocidal activity against M. incognita (LD50/168 h 74.3 μg/mL; J2 mortality LD50 20.9 μg/mL) and X. americanum (LD50/24 h 18.4 μg/mL) | [81] |
| DIMBOA (108) | “ | Nematocidal activity against M. incognita J2 mortality (LD 50 46.1 μg/mL) and X. americanum (LD50/24 h 48.3 μg/mL) | “ |
| BOA (109) | “ | No activity | “ |
| MBOA (110) | Nematocidal activity against M. incognita J2 mortality (LD50 49.2 μg/mL) | “ | |
| 8-Geranylpsoralen, (111) | Apiaceae, Rutacea, Asteraceae and Fabaceae plant families | Nematocidal activity against B. xylophilus LD50/72 h 188.3 μg/mL) and P. redivivus (LD50/72 h 117.5 μg/mL) | [82] |
| Imperatorin (112) | “ | Nematocidal activity against B. xylophilus LD50/72 h 161.7 μg/mL) and P. redivivus (LD50/72 h 179.0 μg/mL) | “ |
| Heraclenin (113) | “ | Nematocidal activity against B. xylophilus LD50/72 h 114.7 μg/mL) and P. redivivus (LD50/72 h 184.7 μg/mL) | “ |
| Ruixianglangdusu B (114) | Stellera chamaejasme L. | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure at 227.4, 71.6, and 15.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure at 1.8 × 103, 160.2, and 0.6 μM, respectively) | [83] |
| Umbelliferone (115) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 1.3 × 107, 5.7 × 107, and 3.3 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 2.6 × 103, 851, and 33.4 μM, respectively) | “ |
| Daphnoretin (116) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure 47.8, 3.1, and 2.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 2.3 × 106, 169.9, and 3.1 μM, respectively) | “ |
| Chamaejasmenine C (117) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 1.7 × 104, 1.1 × 104, and 65.3 μM) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 463.5, 156.7, and 0.05 μM, respectively) | “ |
| 7-Methoxyneochaejasmin A (118) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24 and 72 h exposure <0.001, 3.4 and 167.3 μM) and B. mucronatus (LC50 at 12, 24 and 72 h exposure 1.8 × 104, 384.2, and 151.1 μM) | “ |
| (+)-Chamaejasmine (119) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 16.5, 8.8, and 4.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 1.8 × 103, 1.6 × 103, and 5.1 × 103 μM, respectively) | “ |
| Chamaechromone (120) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 0.7, 10.3, and 36.7 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 327, 5.7, and 0.003 μM, respectively) | “ |
| Isosikokianin A (121) | “ | Nematocidal activity against B. xylophilus (LC50 at 12, 24, and 72 h exposure of 147.7, 385.2, and 2.2 × 102 μM, respectively) and B. mucronatus (LC50 at 12, 24, and 72 h exposure of 2.6 × 104, 32.5, and 2.3 μM, respectively) | |
| 1,4-Naphthoquinone (122) | Rubia wallichiana | Nematocidal activity against N2L4 (kill LC50 value 42.26 μg/mL), N2 (inhibition of egg hatching LC50 34.83 μg/mL), M. incognita (LC50 33.51 μg/mL) | [85] |
| Denguelin (123) | Leguminosae spp. | Nematocidal activity against H. contortus (L3 mortality IC50/24,48,72 h 81, 54 and 21 μM) (L4 mortality IC50/24,48,72 h 11.39, 25.4 and 0.004 μM) | [93] |
| 4-Methoxyisocadalene (124) | Heterotheca inuloides | No activity | [96] |
| 7-Hydroxycadalene (125) | “ | Nematocidal activity against N. aberrans (mortality at J2 stage (LC50 31.30 mg/mL) | “ |
| (4R)-7-hydroxy-3,4-dihydrocadalene (126) | “ | Nematocidal activity against N. aberrans (mortality at J2 stage LC50 26.30 mg/mL) | “ |
| 1𝛼-Hydroxy-1(4H)-isocadalen-4-one (127) | “ | No activity | “ |
| (1R,4R)-1-Hydroxy-4H-1,2, 3,4-tetrahydrocadalen-15-oic acid (128) | “ | “ | “ |
| rac-3,7-Dihydroxy 3(4H)-isocadalen-4-one (129) | “ | “ | |
| 5′-Methoxywaltherione A (130) | Waltheria indica | Nematocidal activity against M. arenaria (EC50 0.25 μg/mL), M. hapla (E50 0.09 μg/mL), M. incognita a(E50 0.09 μg/mL), and B. xylophilus (E50 2.13 μg/mL) | [97] |
| Waltherione A (131) | “ | Nematocidal activity against M. arenaria (EC50 0.63 μg/mL), M. hapla (E50 1.74 μg/mL), M. incognita a(E50 0.27 μg/mL), and B. xylophilus (E50 3.54 μg/mL) | “ |
| Waltherione C (132) | “ | Nematocidal activity against M. arenaria (EC50 10.67 μg/mL), M. hapla (E50 19.79 μg/mL), M. incognita (E50 16.59 μg/mL), and B. xylophilus (EC50 790.85 μg/mL) | “ |
| cis-Dehydromatricaria ester (133) | Tanacetum falconeri | Nematocidal activity against M. incognita (EC50/24,36, 72 h 3.4, 0.18, and 0.04 mg/L) | [98] |
| 3β-Angeloyloxy-6β-hydroxyfuranoeremophil-1(10)-ene (134) | Senecio sinuatos | Nematocidal activity against M. incognita (at 10 mg/mL) | [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evidente, A. Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity. Toxins 2022, 14, 849. https://doi.org/10.3390/toxins14120849
Evidente A. Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity. Toxins. 2022; 14(12):849. https://doi.org/10.3390/toxins14120849
Chicago/Turabian StyleEvidente, Antonio. 2022. "Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity" Toxins 14, no. 12: 849. https://doi.org/10.3390/toxins14120849
APA StyleEvidente, A. (2022). Microbial and Plant Derived Low Risk Pesticides Having Nematocidal Activity. Toxins, 14(12), 849. https://doi.org/10.3390/toxins14120849