Possible Mechanisms of the Interplay between Drugs and Mycotoxins—Is There a Possible Impact?

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Mycotoxins Are Linked to Human Diseases
2.1. General Overview of Mycotoxins and Their Toxic Effects
2.2. Mycotoxins: A Global Health Problem
3. Mycotoxin–Drug Interactions Are an Emerging Research Topic
3.1. Food and Pharmacology: Food–Drug Interactions
3.2. Mycotoxin Metabolism through The CYP450 Complex
3.2.1. Aflatoxins
3.2.2. Fusarium Toxins
4. Mycotoxin–Drug Interactions: In Vitro and In Vivo Studies
4.1. General Overview
4.2. Aflatoxins
4.3. Fusarium Toxins
4.4. Physiologically-Based Pharmacokinetic Modelling to Predict Mycotoxin-Drug Interactions
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2019, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in food and feed. Adv. Food Nutr. Res. 2019, 89, 297–345. [Google Scholar]
- Sobral, M.; Faria, M.; Cunha, S.; Ferreire, I. Toxicological Interactions between Mycotoxins from Ubiquitous Fungi: Impact on Hepatic and Intestinal Human Epithelial Cells. Chemosphere 2018, 202, 538–548. [Google Scholar] [CrossRef]
- Kebede, H.; Liu, X.; Jin, J.; Xing, F. Current status of major mycotoxins contamination in food and feed in Africa. Food Control 2020, 110, 106975. [Google Scholar] [CrossRef]
- Richard, J.L. Discovery of Aflatoxins and Significant Historical Features. Toxin Rev. 2008, 27, 171–201. [Google Scholar] [CrossRef]
- Mahato, D.K.; Lee, K.E.; Kamle, M.; Devi, S.; Dewangan, K.N.; Kumar, P.; Kang, S.G. Aflatoxins in Food and Feed: An Overview on Prevalence, Detection and Control Strategies. Front. Microbiol. 2019, 10, 2266. [Google Scholar] [CrossRef] [Green Version]
- Bryła, M.; Waśkiewicz, A.; Ksieniewicz-Woźniak, E.; Szymczyk, K.; Jędrzejczak, R. Modified Fusarium Mycotoxins in Cereals and Their Products-Metabolism, Occurrence, and Toxicity: An Updated Review. Molecules 2018, 23, 963. [Google Scholar] [CrossRef] [Green Version]
- Adekoya, I.; Njobeh, P.; Obadina, A.; Chilaka, C.; Okoth, S.; De Boevre, M.; De Saeger, S. Awareness and Prevalence of Mycotoxin Contamination in Selected Nigerian Fermented Foods. Toxins 2017, 9, 363. [Google Scholar] [CrossRef] [Green Version]
- IARC. Monograph IARC Aflatoxins; IARC: Lyon, France, 2002. Available online: https://monographs.iarc.fr/wp-content/uploads/2018/06/mono100F-23.pdf (accessed on 25 May 2020).
- Claeys, L.; Romano, C.; De Ruyck, K.; Wilson, H.; Fervers, B.; Korenjak, M.; Zavadil, J.; Gunter, M.J.; De Saeger, S.; De Boevre, M.; et al. Mycotoxin exposure and human cancer risk: A systematic review of epidemiological studies. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1449–1464. [Google Scholar] [CrossRef] [PubMed]
- Lumsangkul, C.; Chiang, H.-I.; Lo, N.-W.; Fan, Y.-K.; Ju, J.-C. Developmental Toxicity of Mycotoxin Fumonisin B₁ in Animal Embryogenesis: An Overview. Toxins 2019, 11, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IARC. IARC Publications Website—Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1993. Available online: https://publications.iarc.fr/74 (accessed on 28 February 2022).
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Mayer, E.; Novak, B.; Springler, A.; Schwartz-Zimmermann, H.E.; Nagl, V.; Reisinger, N.; Hessenberger, S.; Schatzmayr, G. Effects of deoxynivalenol (DON) and its microbial biotransformation product deepoxy-deoxynivalenol (DOM-1) on a trout, pig, mouse, and human cell line. Mycotoxin Res. 2017, 33, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Sorrenti, V.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Bognanno, M.; Galvano, F. Toxicity of ochratoxin a and its modulation by antioxidants: A review. Toxins 2013, 5, 1742–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 Years of Research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Health & Consumer protection directorate-general Opinion of The Scientific Committee on Food. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/cs_contaminants_catalogue_out88_en.pdf (accessed on 6 August 2020).
- Gilbert, D.L.E.S.G. Principles of Toxicology|Casarett and Doull’s Toxicology: The Basic Science of Poisons, 8e|AccessPharmacy|McGraw-Hill Medical. 2013. Available online: https://accesspharmacy.mhmedical.com/content.aspx?bookid=958§ionid=53483721 (accessed on 11 March 2020).
- Wild, C.P. The exposome: From concept to utility. Int. J. Epidemiol. 2012, 41, 24–32. [Google Scholar] [CrossRef]
- Darwish, W.S.; Ikenaka, Y.; Nakayama, S.M.M.; Ishizuka, M. An overview on mycotoxin contamination of foods in Africa. J. Vet. Med. Sci. 2014, 76, 789–797. [Google Scholar] [CrossRef] [Green Version]
- Patrick, N.B.; Dutton, F.; Michael, D.F.; Hussaini, M.A. (PDF) Mycotoxins and Human Health: Significance, Prevention and Control. 2010. Available online: https://www.researchgate.net/publication/281308830_Mycotoxins_and_human_health_significance_prevention_and_control (accessed on 24 March 2020).
- Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre- and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Russell, R.; Paterson, M.; Lima, N. How will climate change affect mycotoxins in food? Food Res. Int. 2010, 43, 1902–1914. [Google Scholar] [CrossRef] [Green Version]
- Azziz-Baumgartner, E.; Lindblade, K.; Gieseker, K.; Rogers, H.S.; Kieszak, S.; Njapau, H.; Schleicher, R.; McCoy, L.F.; Misore, A.; DeCock, K.; et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environ. Health Perspect. 2005, 113, 1779–1783. [Google Scholar] [CrossRef] [PubMed]
- Kamala, A.; Shirima, C.; Jani, B.; Bakari, M.; Sillo, H.; Rusibamayila, N.; De Saeger, S.; Kimanya, M.; Gong, Y.Y.; Simba, A.; et al. Outbreak of an acute aflatoxicosis in Tanzania during 2016. World Mycotoxin J. 2018, 11, 311–320. [Google Scholar] [CrossRef]
- WHO. Health Emergency Information and Risk Assessment. 2019. Available online: https://apps.who.int/iris/bitstream/handle/10665/326465/OEW33-1218082019.pdf (accessed on 24 March 2020).
- Jonsyn-Ellis, F.E. Seasonal variation in exposure frequency and concentration levels of aflatoxins and ochratoxins in urine samples of boys and girls. Mycopathologia 2001, 152, 35–40. [Google Scholar] [CrossRef]
- Gautham, D. Assessment of Aflatoxin B1 Level in Chilli, Maize and Groundnut Samples from Kathmandu Valley. J. Food Sci. Technol. Nepal 2008, 4, 57–60. Available online: https://www.nepjol.info/index.php/JFSTN/article/view/2000 (accessed on 11 March 2020).
- Wu, H.C.; Santella, R. The role of aflatoxins in hepatocellular carcinoma. Hepat. Mon. 2012, 12, e7238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chang, C.-C.H.; Marsh, G.M.; Wu, F. Population attributable risk of aflatoxin-related liver cancer: Systematic review and meta-analysis. Eur. J. Cancer 2012, 48, 2125–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibb, H.; Devleesschauwer, B.; Bolger, P.M.; Wu, F.; Ezendam, J.; Cliff, J.; Zeilmaker, M.; Verger, P.; Pitt, J.; Baines, J.; et al. World Health Organization estimates of the global and regional disease burden of four foodborne chemical toxins, 2010: A data synthesis. F1000Research 2015, 4, 1393. [Google Scholar] [CrossRef]
- Kimanya, M.E.; Routledge, M.N.; Mpolya, E.; Ezekiel, C.N.; Shirima, C.P.; Gong, Y.Y. Estimating the risk of aflatoxin-induced liver cancer in Tanzania based on biomarker data. PLoS ONE 2021, 16, e0247281. [Google Scholar] [CrossRef]
- Sangare-Tigori, B.; Moukha, S.; Kouadio, J.H.; Dano, D.S.; Betbeder, A.-M.; Achour, A.; Creppy, E.E. Ochratoxin A in human blood in Abidjan, Côte d’Ivoire. Toxicon 2006, 47, 894–900. [Google Scholar] [CrossRef]
- Zaied, C.; Bouaziz, C.; Azizi, I.; Bensassi, F.; Chour, A.; Bacha, H.; Abid, S. Presence of ochratoxin A in Tunisian blood nephropathy patients. Exposure level to OTA. Exp. Toxicol. Pathol. 2011, 63, 613–618. [Google Scholar] [CrossRef]
- Chen, C.; Mitchell, N.J.; Gratz, J.; Houpt, E.R.; Gong, Y.; Egner, P.A.; Groopman, J.D.; Riley, R.T.; Showker, J.L.; Svensen, E.; et al. Exposure to aflatoxin and fumonisin in children at risk for growth impairment in rural Tanzania. Environ. Int. 2018, 115, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Jolly, P.E.; Preko, P.; Wang, J.-S.; Ellis, W.O.; Phillips, T.D.; Williams, J.H. Aflatoxin-Related Immune Dysfunction in Health and in Human Immunodeficiency Virus Disease. Clin. Dev. Immunol. 2008, 2008, 790309. [Google Scholar] [CrossRef] [PubMed]
- Afum, C.; Cudjoe, L.; Hills, J.; Hunt, R.; Padilla, L.A.; Elmore, S.; Afriyie, A.; Opare-Sem, O.; Phillips, T.; Jolly, P.E. Association between Aflatoxin M₁ and Liver Disease in HBV/HCV Infected Persons in Ghana. Int. J. Environ. Res. Public Health 2016, 13, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Person, S.D.; Funkhouser, E.; Yatich, N.J.; Stiles, J.K.; Ellis, W.O.; Jiang, Y.; Ehiri, J.E.; Williams, J.H.; Jolly, P.E.; Wilson, C.; et al. Association between Anemia and Aflatoxin B1 Biomarker Levels among Pregnant Women in Kumasi, Ghana. Am. J. Trop. Med. Hyg. 2010, 83, 1077–1083. [Google Scholar] [CrossRef] [Green Version]
- Komsky-Elbaz, A.; Saktsier, M.; Roth, Z. Aflatoxin B1 impairs sperm quality and fertilization competence. Toxicology 2018, 393, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, Y.; Bhat, R.V.; Ravindranath, V. Production of Deoxynivalenol by Fusarium Isolates from Samples of Wheat Associated with a Human Mycotoxicosis Outbreak and from Sorghum Cultivars. 1989. Volume 55. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC203133/pdf/aem00103-0199.pdf (accessed on 25 March 2020).
- Aquino, S.; Corre, B. Aflatoxins in Pet Foods: A Risk to Special Consumers. In Aflatoxins—Detection, Measurement and Control; InTech: London, UK, 2011. [Google Scholar]
- González Pereyra, M.L.; Carvalho, E.C.Q.; Tissera, J.L.; Keller, K.M.; Magnoli, C.E.; Rosa, C.A.R.; Dalcero, A.M.; Cavaglieri, L.R. An outbreak of acute aflatoxicosis on a chinchilla (Chinchilla lanigera) farm in Argentina. J. Vet. Diagn. Investig. 2008, 20, 853–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BIOMIN Mycotoxin Survey Q3 2021 Results|Biomin.net. Available online: https://www.biomin.net/science-hub/biomin-mycotoxin-survey-q3-2021-results/ (accessed on 10 November 2022).
- Peter Mshelia, L.; Selamat, J.; Iskandar Putra Samsudin, N.; Rafii, M.Y.; Abdul Mutalib, N.A.; Nordin, N.; Berthiller, F. Effect of Temperature, Water Activity and Carbon Dioxide on Fungal Growth and Mycotoxin Production of Acclimatised Isolates of Fusarium verticillioides and F. graminearum. Toxins 2020, 12, 478. [Google Scholar] [CrossRef]
- Biomin, “AllAboutFeed—New Data Shows Mycotoxins Are Not under Control”. 2018. Available online: https://www.allaboutfeed.net/Mycotoxins/Articles/2018/8/New-data-shows-mycotoxins-are-not-under-control-321012E/ (accessed on 26 March 2020).
- Bushra, R.; Aslam, N.; Khan, A.Y. Food-drug interactions. Oman Med. J. 2011, 26, 77–83. [Google Scholar] [CrossRef]
- Weininger, J. Nutritional Disease—Food-Drug Interactions|Britannica. 2016. Available online: https://www.britannica.com/science/nutritional-disease/Foodborne-illnesses (accessed on 6 December 2019).
- Zhou, S.-F.; Liu, J.-P.; Chowbay, B. Polymorphism of human cytochrome P450 enzymes and its clinical impact. Drug Metab. Rev. 2009, 41, 89–295. [Google Scholar] [CrossRef]
- Jancova, P.; Anzenbacher, P.; Anzenbacherova, E. Phase II drug metabolizing enzymes. Biomed. Pap. 2010, 154, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Hannemann, F.; Bichet, A.; Ewen, K.M.; Bernhardt, R. Cytochrome P450 systems—Biological variations of electron transport chains. Biochim. Biophys. Acta—Gen. Subj. 2007, 1770, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Tracy, T.S.; Chaudhry, A.S.; Prasad, B.; Thummel, K.E.; Schuetz, E.G.; Zhong, X.B.; Tien, Y.C.; Jeong, H.; Pan, X.; Shireman, L.M.; et al. Interindividual Variability in Cytochrome P450-Mediated Drug Metabolism. Drug Metab. Dispos. 2016, 44, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, B.; Molony, C.; Chudin, E.; Hao, K.; Zhu, J.; Gaedigk, A.; Suver, C.; Zhong, H.; Leeder, J.S.; et al. Systematic genetic and genomic analysis of cytochrome P450 enzyme activities in human liver. Genome Res. 2010, 20, 1020–1036. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Zhao, L.; Zhang, N.Y.; Karrow, N.A.; Krumm, C.S.; Qi, D.S.; Sun, L.H. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res. Rev. Mutat. Res. 2018, 778, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Bbosa, G.S.; Kitya, D.; Odda, J.; Ogwal-Okeng, J. Aflatoxins metabolism, effects on epigenetic mechanisms and their role in carcinogenesis. Health 2013, 5, 14–34. [Google Scholar] [CrossRef] [Green Version]
- Jolly, P.; Jiang, Y.; Ellis, W.; Awuah, R.; Nnedu, O.; Phillips, T.; Wang, J.-S.; Afriyie-Gyawu, E.; Tang, L.; Person, S.; et al. Determinants of aflatoxin levels in Ghanaians: Sociodemographic factors, knowledge of aflatoxin and food handling and consumption practices. Int. J. Hyg. Environ.-Health 2006, 209, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Mengelers, M.; Yang, S.; De Saeger, S.; De Boevre, M. Mycotoxin Biomarkers of Exposure: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1127–1155. [Google Scholar] [CrossRef] [Green Version]
- Schwartz-Zimmermann, H.E.; Fruhmann, P.; Dänicke, S.; Wiesenberger, G.; Caha, S.; Weber, J.; Berthiller, F. Metabolism of deoxynivalenol and deepoxy-deoxynivalenol in broiler chickens, pullets, roosters and turkeys. Toxins 2015, 7, 4706–4729. [Google Scholar] [CrossRef]
- Schelstraete, W. Interactions between Fusarium Mycotoxins and Cytochrome P450 Drug Metabolizing Enzymes in a Porcine Animal Model. Available online: https://biblio.ugent.be/publication/8632658/file/8632659 (accessed on 22 August 2022).
- Vidal, A.; Claeys, L.; Mengelers, M.; Vanhoorne, V.; Vervaet, C.; Huybrechts, B.; De Saeger, S.; De Boevre, M. Humans significantly metabolize and excrete the mycotoxin deoxynivalenol and its modified form deoxynivalenol-3-glucoside within 24 hours. Sci. Rep. 2018, 8, 5255. [Google Scholar] [CrossRef] [Green Version]
- Warth, B.; Sulyok, M.; Berthiller, F.; Schuhmacher, R.; Krska, R. New insights into the human metabolism of the Fusarium mycotoxins deoxynivalenol and zearalenone. Toxicol. Lett. 2013, 220, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Sayyari, A.; Kruse Faeste, C.; Hansen, U.; Uhlig, S.; Framstad, T.; Schatzmayr, D.; Sivertsen, T. Effects and biotransformation of the mycotoxin deoxynivalenol in growing pigs fed naturally-contaminated grain pelleted with and without the addition of Coriobacteriaceum DSM 11798. Food Addit. Contam. Part A 2018, 35, 1394–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuca, K.; Dohnal, V.; Jezkova, A.; Jun, D. Metabolic Pathways of T-2 Toxin. Curr. Drug Metab. 2008, 9, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.N.; Chen, J.; Xu, B.; Wei, X.; Guo, L.; Xie, J.W. The roles of carboxylesterase and CYP isozymes on the in vitro metabolism of T-2 toxin. Mil. Med. Res. 2015, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Bravin, F.; Duca, R.C.; Balaguer, P.; Delaforge, M. In Vitro cytochrome P450 formation of a mono-hydroxylated metabolite of zearalenone exhibiting estrogenic activities: Possible occurrence of this metabolite in Vivo. Int. J. Mol. Sci. 2009, 10, 1824–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, E.; Hildebrand, A.; Damm, G.; Rapp, A.; Cramer, B.; Humpf, H.-U.; Metzler, M. Aromatic hydroxylation is a major metabolic pathway of the mycotoxin zearalenone in vitro. Mol. Nutr. Food Res. 2009, 53, 1123–1133. [Google Scholar] [CrossRef]
- Shoshei, S.; Makoto, K. Fumonisin B1—An Overview|ScienceDirect Topics. 2010. Available online: https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/fumonisin-b1 (accessed on 9 December 2019).
- Mary, V. Effects of AFB1, FB and their mixture on the aryl hydrocarbon receptor and CYP450 1A induction. Food Chem. Toxicol. 2015, 75, 104–111. [Google Scholar] [CrossRef]
- Fodor, J.; Meyer, K.; Gottschalk, C.; Mamet, R.; Kametler, L.; Bauer, J.M.; Horn, P.; Kovacs, F.; Kovacs, M. In vitro microbial metabolism of fumonisin B1. Food Addit. Contam. 2007, 24, 416–420. [Google Scholar] [CrossRef] [Green Version]
- Antonissen, G.; De Baere, S.; Novak, B.; Schatzmayr, D.; den Hollander, D.; Devreese, M.; Croubels, S. Toxicokinetics of Hydrolyzed Fumonisin B1 after Single Oral or Intravenous Bolus to Broiler Chickens Fed a Control or a Fumonisins-Contaminated Diet. Toxins 2020, 12, 413. [Google Scholar] [CrossRef]
- Ivanova, L.; Uhlig, S.; Devreese, M.; Croubels, S.; Fæste, C.K. Biotransformation of the mycotoxin enniatin B1 in pigs: A comparative in vitro and in vivo approach. Food Chem. Toxicol. 2017, 105, 506–517. [Google Scholar] [CrossRef]
- Fæste, C.K.; Ivanova, L.; Uhlig, S. In vitro metabolism of the mycotoxin enniatin B in different species and cytochrome P450 enzyme phenotyping by chemical inhibitors. Drug Metab. Dispos. 2011, 39, 1768–1776. [Google Scholar] [CrossRef] [Green Version]
- Anyanwu, E.C.; Campbell, A.W.; Ehiri, J.E. Mycotoxins and antifungal drug interactions: Implications in the treatment of illnesses due to indoor chronic toxigenic mold exposures. Sci. World J. 2004, 4, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Liew, W.P.P.; Mohd-Redzwan, S. Mycotoxin: Its impact on gut health and microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Galtier, P.; Meissonnier, G.; Laffitte, J.; Oswald, I.P.; Loiseau, N. Molecular Interactions between Mycotoxins and Liver Enzymes Involved in Drug Metabolism in Rodents and Farm Animals. 2008. Available online: https://hrcak.srce.hr/index.php?show=clanak&id_clanak_jezik=45495 (accessed on 6 December 2019).
- Ayed-Boussema, I.; Pascussi, J.M.; Maurel, P.; Bacha, H.; Hassen, W. Zearalenone activates pregnane X receptor, constitutive androstane receptor and aryl hydrocarbon receptor and corresponding phase I target genes mRNA in primary cultures of human hepatocytes. Environ. Toxicol. Pharmacol. 2011, 31, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ayed-Boussema, I.; Pascussi, J.M.; Maurel, P.; Bacha, H.; Hassen, W. Effect of aflatoxin B1 on nuclear receptors PXR, CAR, and AhR and their target cytochromes P450 mRNA expression in primary cultures of human hepatocytes. Int. J. Toxicol. 2012, 31, 86–93. [Google Scholar] [CrossRef]
- Gouze, M.E.; Laffitte, J.; Rouimi, P.; Loiseau, N.; Oswald, I.P.; Galtier, P. Effect of various doses of deoxynivalenol on liver xenobiotic metabolizing enzymes in mice. Food Chem. Toxicol. 2006, 44, 476–483. [Google Scholar] [CrossRef]
- Guerre, P.; Eeckhoutte, C.; Burgat, V.; Galtier, P.; Guerrer, P. The effects of T-2 toxin exposure on liver drug metabolizing enzymes in rabbit. Food Addit. Contam. 2010, 17, 1019–1026. [Google Scholar] [CrossRef]
- Osselaere, A.; Li, S.J.; De Bock, L.; Devreese, M.; Goossens, J.; Vandenbroucke, V.; Van Bocxlaer, J.; Boussery, K.; Pasmans, F.; Martel, A.; et al. Toxic effects of dietary exposure to T-2 toxin on intestinal and hepatic biotransformation enzymes and drug transporter systems in broiler chickens. Food Chem. Toxicol. 2013, 55, 150–155. [Google Scholar] [CrossRef]
- Spotti, M.; Maas, R.F.M.; De Nijs, C.M.; Fink-Gremmels, J. Effect of fumonisin B1 on rat hepatic P450 system. Environ. Toxicol. Pharmacol. 2000, 8, 197–204. [Google Scholar] [CrossRef]
- Antonissen, G.; Devreese, M.; De Baere, S.; Martel, A.; Van Immerseel, F.; Croubels, S. Impact of Fusarium mycotoxins on hepatic and intestinal mRNA expression of cytochrome P450 enzymes and drug transporters, and on the pharmacokinetics of oral enrofloxacin in broiler chickens. Food Chem. Toxicol. 2017, 101, 75–83. [Google Scholar] [CrossRef]
- Bertero, A.; Fossati, P.; Eurosia, D.; Tedesco, A.; Caloni, F. toxins Beauvericin and Enniatins: In Vitro Intestinal Effects. Toxins 2020, 12, 686. [Google Scholar] [CrossRef]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarisco, J.A.; Hall, J.O.; Coulombe, R.A. Mechanisms of butylated hydroxytoluene chemoprevention of aflatoxicosis—Inhibition of aflatoxin B1 metabolism. Toxicol. Appl. Pharmacol. 2008, 227, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; He, X.Y.; Otieno, M.; Egner, P.A.; Jacobson, L.P.; Chen, B.; Wang, J.S. Oltipraz Chemoprevention Trial in Qidong, People’s Republic of China: Modulation of Serum Aflatoxin Albumin Adduct Biomarkers—PubMed. 1998. Available online: https://pubmed.ncbi.nlm.nih.gov/9488587/ (accessed on 26 January 2022).
- Buetler, T.M.; Bammler, T.K.; Hayes, J.D.; Eaton, D.L. Oltipraz-mediated Changes in Aflatoxin B1 Biotransformation in Rat Liver: Implications for Human Chemointervention. Cancer Res. 1996, 56, 2306–2313. [Google Scholar] [PubMed]
- Bammler, T.K.; Slone, D.H.; Eaton, D.L. Effects of Dietary Oltipraz and Ethoxyquin on Aflatoxin B1 Biotransformation in Non-Human Primates. Toxicol. Sci. 2000, 54, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Langouët, S.; Coles, B.; Morel, F.; Becquemont, L.; Beaune, P.; Guengerich, F.P.; Ketterer, B.; Guillouzo, A. Inhibition of CYP1A2 and CYP3A4 by Oltipraz Results in Reduction of Aflatoxin B1 Metabolism in Human Hepatocytes in Primary Culture—PubMed. 1995. Available online: https://pubmed.ncbi.nlm.nih.gov/7585637/ (accessed on 27 November 2020).
- Schelstraete, W.; Devreese, M.; Croubels, S. Impact of Subacute Exposure to T-2 Toxin and Zearalenone on the Pharmacokinetics of Midazolam as CYP3A Probe Drug in a Porcine Animal Model: A Pilot Study. Front. Pharmacol. 2019, 10, 399. [Google Scholar] [CrossRef] [Green Version]
- Guevara-Gonzalez, R. Aflatoxins—Biochemistry and Molecular Biology; InTech: London, UK, 2011; ISBN 9789533073958. [Google Scholar]
- Watanabe, K.P.; Kawai, Y.K.; Ikenaka, Y.; Kawata, M.; Ikushiro, S.-I.; Sakaki, T.; Ishizuka, M. Avian cytochrome P450 (CYP) 1-3 family genes: Isoforms, evolutionary relationships, and mRNA expression in chicken liver. PLoS ONE 2013, 8, e75689. [Google Scholar] [CrossRef] [Green Version]
- Dohnal, V.; Wu, Q.; Kuča, K. Metabolism of aflatoxins: Key enzymes and interindividual as well as interspecies differences. Arch. Toxicol. 2014, 88, 1635–1644. [Google Scholar] [CrossRef]
- Diaz, G.J.; Murcia, H.W. An unusually high production of hepatic aflatoxin B1-dihydrodiol, the possible explanation for the high susceptibility of ducks to aflatoxin B1. Sci. Rep. 2019, 9, 8010. [Google Scholar] [CrossRef] [Green Version]
- Corcueral, L.-A.; Vettorazzi, A.; Arbillagal, L.; Perez, N.; Gill, A.G.; Azquetal, A.; Penas, E.; Garcia-Jalon, J.A.; Lopez de Cerain, A. Genotoxicity of Aflatoxin B1 and Ochratoxin A after Simultaneous Application of the In Vivo Micronucleus and Comet Assay. Food Chem. Toxicol. 2015, 76, 116–124. [Google Scholar]
- Lootens, O.; De Boevre, M.; Gasthuys, E.; Van Bocxlaer, J.; Vermeulen, A.; De Saeger, S. Unravelling the pharmacokinetics of aflatoxin B1: In vitro determination of Michaelis–Menten constants, intrinsic clearance and the metabolic contribution of CYP1A2 and CYP3A4 in pooled human liver microsomes. Front. Microbiol. 2022, 13, 3258. [Google Scholar] [CrossRef]
- Zhuang, X.; Lu, C. PBPK modeling and simulation in drug research and development. Acta Pharm. Sin. B 2016, 6, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Sager, J.E.; Yu, J.; Ragueneau-Majlessi, I.; Isoherranen, N. Physiologically Based Pharmacokinetic (PBPK) Modeling and Simulation Approaches: A Systematic Review of Published Models, Applications, and Model Verification. Drug Metab. Dispos. 2015, 43, 1823. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Lin, Z.; Zeng, Z.; Fang, B.; Li, M.; Cheng, Y.H.; Sun, Y. Assessing Global Human Exposure to T-2 Toxin via Poultry Meat Consumption Using a Lifetime Physiologically Based Pharmacokinetic Model. J. Agric. Food Chem. 2019, 67, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Alvito, P. Deliverables—HBM4EU—Science and Policy for a Healthy Future. 2021. Available online: https://www.hbm4eu.eu/result/deliverables/ (accessed on 29 August 2022).
- Gilbert-Sandoval, I.; Wesseling, S.; Rietjens, I.M.C.M. Predicting the Acute Liver Toxicity of Aflatoxin B1 in Rats and Humans by an In Vitro–In Silico Testing Strategy. Mol. Nutr. Food Res. 2020, 64, e2000063. [Google Scholar] [CrossRef] [PubMed]
- Youdim, A. Nutrient-Drug Interactions–Nutritional Disorders–MSD Manual Professional Edition. 2019. Available online: https://www.msdmanuals.com/professional/nutritional-disorders/nutrition-general-considerations/nutrient-drug-interactions (accessed on 10 December 2019).


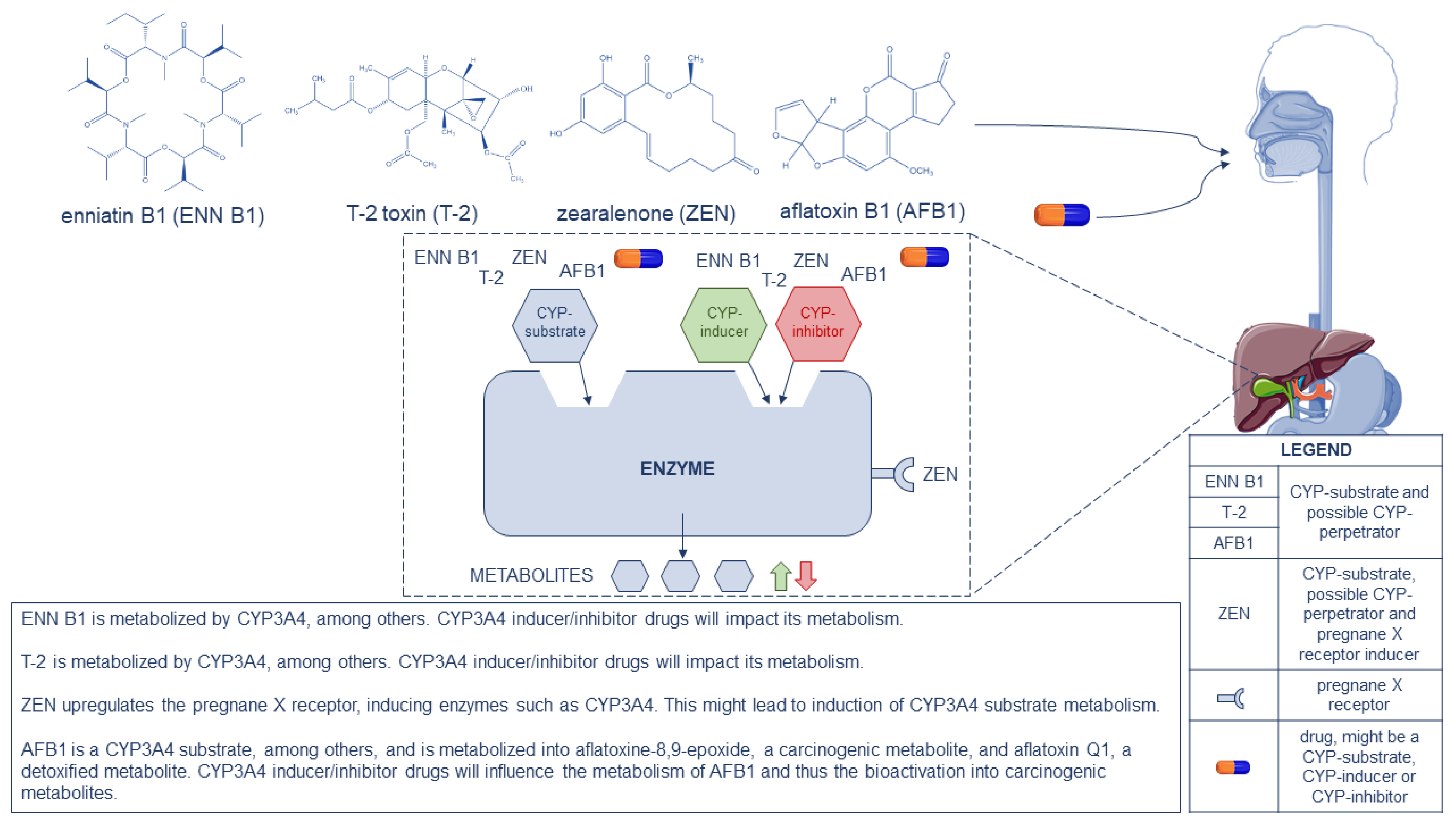{kind=link}
| Mycotoxin | IARC Classification * | Toxic Effects |
|---|---|---|
| Aflatoxin B1 | Group 1 [11] | Cancer, hepatotoxicity, immunosuppression [12] |
| Fumonisin B1 | Group 2B [11] | Cancer, hepatotoxicity, leuko-encephalomalacia, teratogenic effects [12,13] |
| Zearalenone | Group 3 [14] | Infertility, abortion, cervical and breast cancer [15] |
| Deoxynivalenol | Group 3 [14] | Gastrointestinal toxicity, inflammation of central nervous system [16] |
| Ochratoxin A | Group 2B [14] | Nephrotoxicity, cancer, teratogenic and immunotoxic effects [17,18] |
| T-2-toxin | Group 3 [14] | Dermatitis, diarrhea, hemorrhages, necrosis of bone marrow, spleen, ovary, testis and gastrointestinal lining. [19] |
| Mycotoxin | Involved CYP-Enzymes | References |
|---|---|---|
| AFB1 | CYP3A4, CYP3A5, CYP3A7, CYP1A2, CYP2A13 | [55,56,57,58] |
| DON | No phase I metabolism | [59,60,61,62,63] |
| T-2 | CYP3A4, CYP2E1, CYP1A2, CYP2B6, CYP2D6 and CYP2C19 | [64,65] |
| ZEN | CYP3A4, CYP3A5, CYP2C8, CYP1A2 | [66,67] |
| FB1 | No phase I metabolism | [68,69,70,71] |
| ENN B1 | CYP3A4, CYP3A5, CYP1A2, CYP2C19 | [72,73] |
| Mycotoxin | Induction | Inhibition | In Vitro/In Vivo | References |
|---|---|---|---|---|
| aflatoxin B1 | CYP1A2 CYP2B6 CYP2C9 CYP3A4 CYP3A5 | CYP1A1 (rabbits) CYP3A6 (rabbits) | In vitro | [76,78] |
| deoxynivalenol | CYP2B | / | In vitro (mice) | [76,79] |
| T-2 toxin | / | CYP1A1 CYP1A2 CYP2A1 CYP2B4 CYP1A4 CYP1A5 CYP3A37 | In vivo (rabbits) In vivo (chicken) | [80] [81] |
| zearalenone | PXR-receptor | / | In vitro | [77] |
| fumonisin B1 | CYP1A4 (chicken) CYP2E1 (rats) | CYP1A2 (rats) CYP2C11 (rats) | In vivo | [76,82,83] |
| enniatin B1 | / | CYP2C19 | In vitro | [84] |
| Perpetrator | Victim | References |
|---|---|---|
| antifungal drugs with azole structure | mycotoxins with an azole structure | [74] |
| butylated hydroxytoluene | aflatoxin B1 | [86] |
| oltipraz | aflatoxin B1 | [87,88,89,90] |
| zearalenone | midazolam | [91] * |
| T-2 toxin | midazolam | [91] * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lootens, O.; Vermeulen, A.; Croubels, S.; De Saeger, S.; Van Bocxlaer, J.; De Boevre, M. Possible Mechanisms of the Interplay between Drugs and Mycotoxins—Is There a Possible Impact? Toxins 2022, 14, 873. https://doi.org/10.3390/toxins14120873
Lootens O, Vermeulen A, Croubels S, De Saeger S, Van Bocxlaer J, De Boevre M. Possible Mechanisms of the Interplay between Drugs and Mycotoxins—Is There a Possible Impact? Toxins. 2022; 14(12):873. https://doi.org/10.3390/toxins14120873
Chicago/Turabian StyleLootens, Orphélie, An Vermeulen, Siska Croubels, Sarah De Saeger, Jan Van Bocxlaer, and Marthe De Boevre. 2022. "Possible Mechanisms of the Interplay between Drugs and Mycotoxins—Is There a Possible Impact?" Toxins 14, no. 12: 873. https://doi.org/10.3390/toxins14120873
APA StyleLootens, O., Vermeulen, A., Croubels, S., De Saeger, S., Van Bocxlaer, J., & De Boevre, M. (2022). Possible Mechanisms of the Interplay between Drugs and Mycotoxins—Is There a Possible Impact? Toxins, 14(12), 873. https://doi.org/10.3390/toxins14120873