Candidalysin Is the Hemolytic Factor of Candida albicans

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
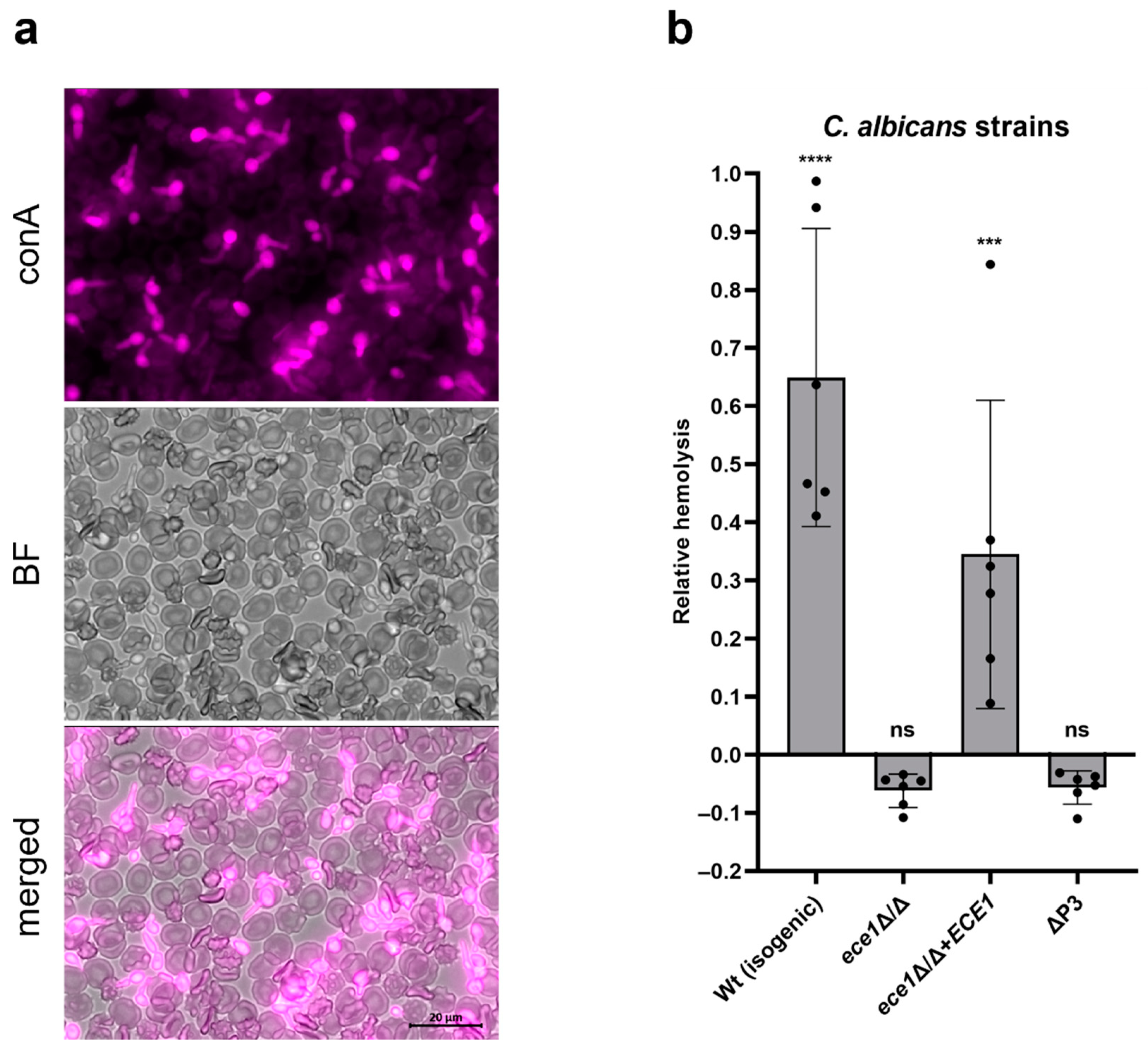
2.1. Ece1 Is Responsible for Human RBC Lysis by C. albicans Hyphae
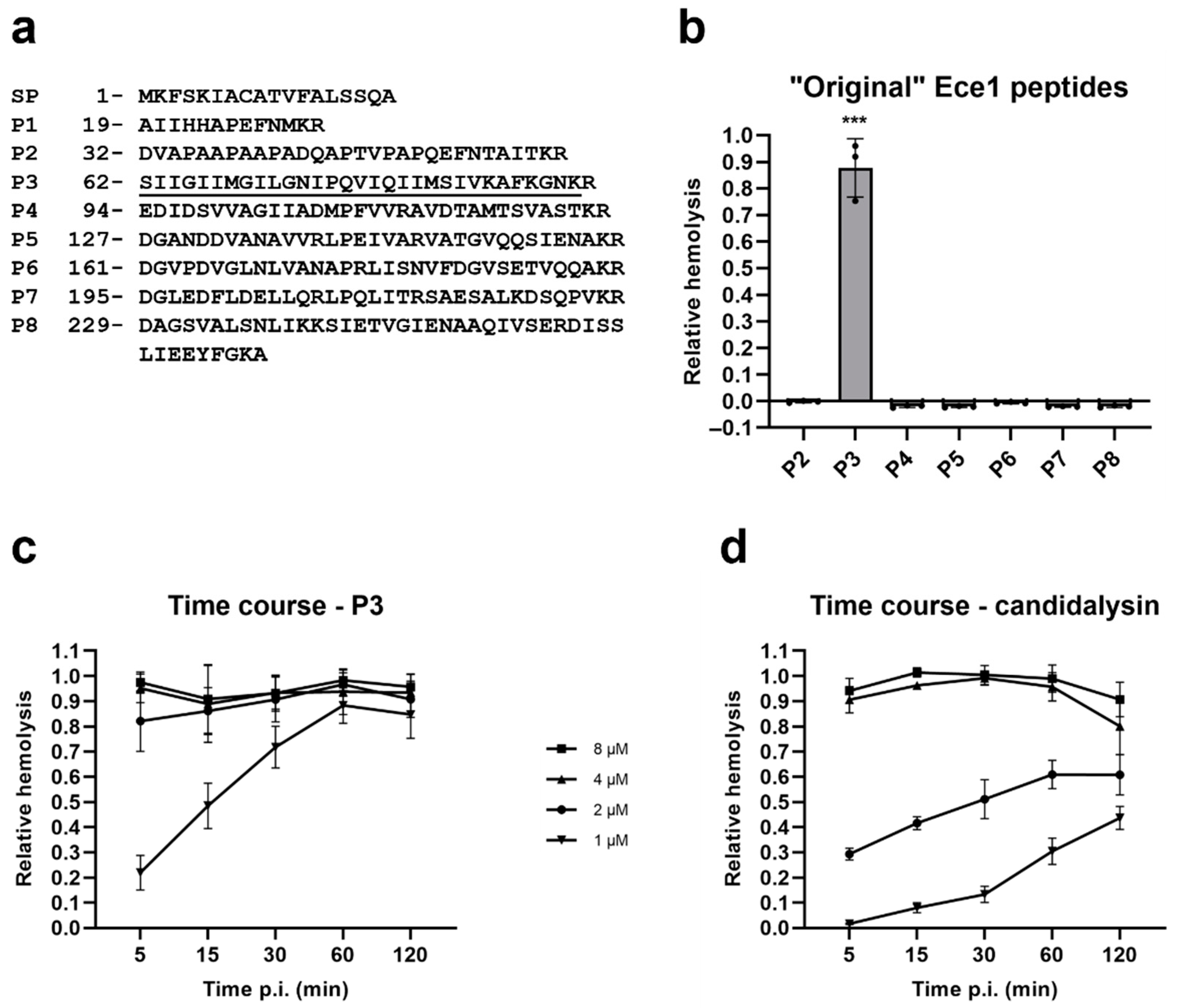
2.2. Candidalysin Is Also a Hemolytic Peptide Toxin
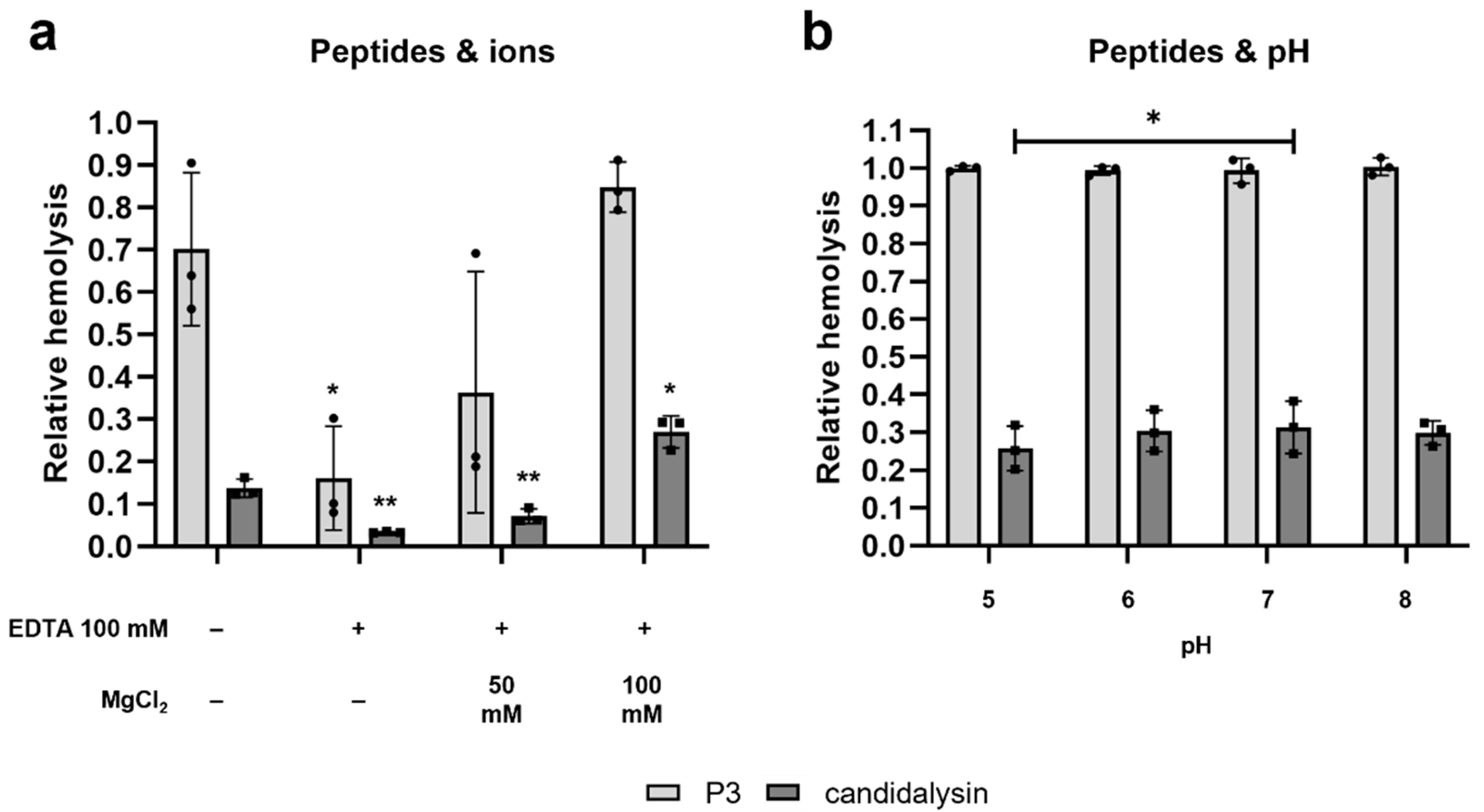
2.3. The Hemolytic Potential of Candidalysin and P3 Is Ion-Dependent and pH-Independent
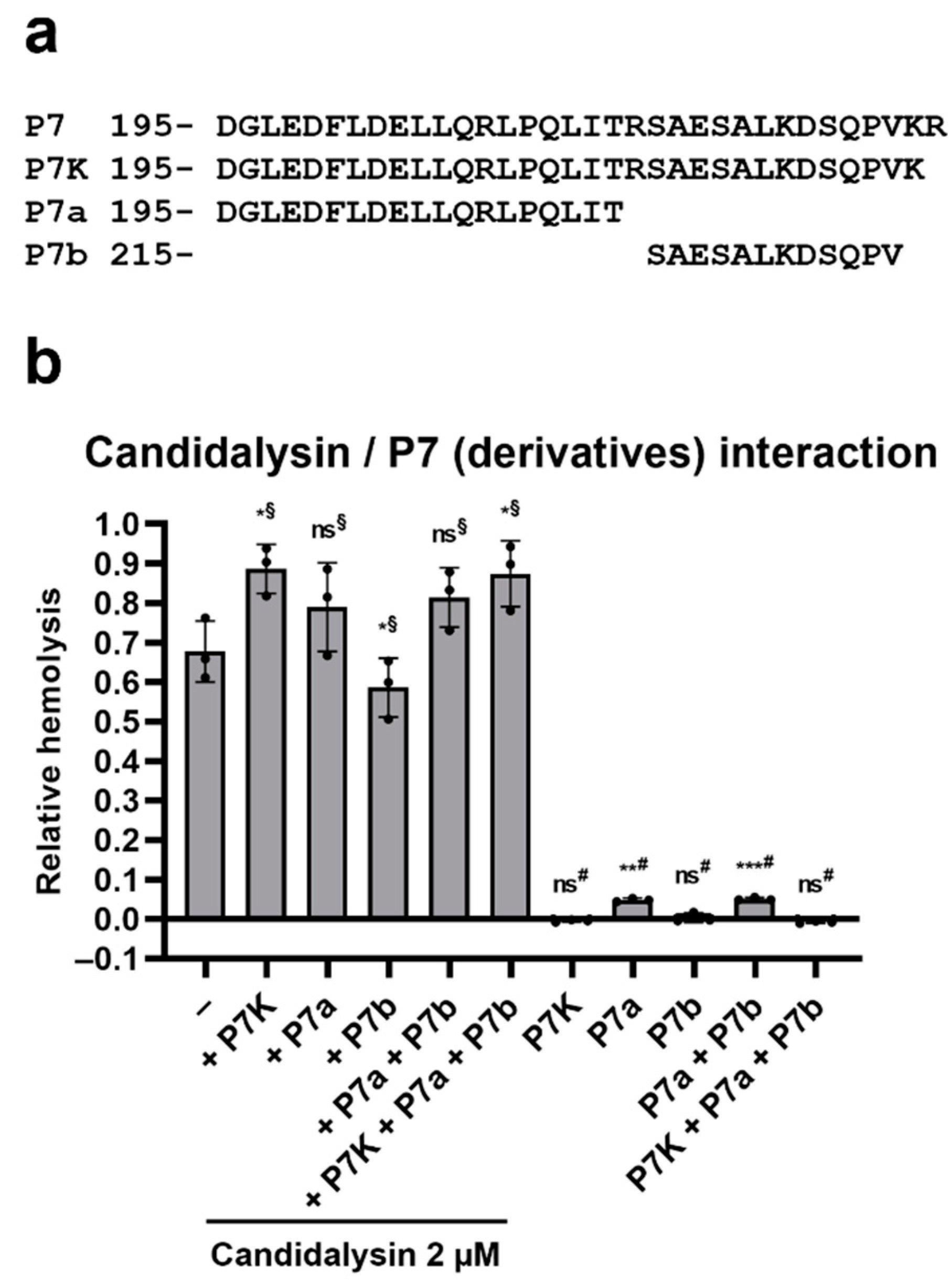
2.4. Ece1 P7 Derivatives Modulate the Hemolytic Potential of Candidalysin
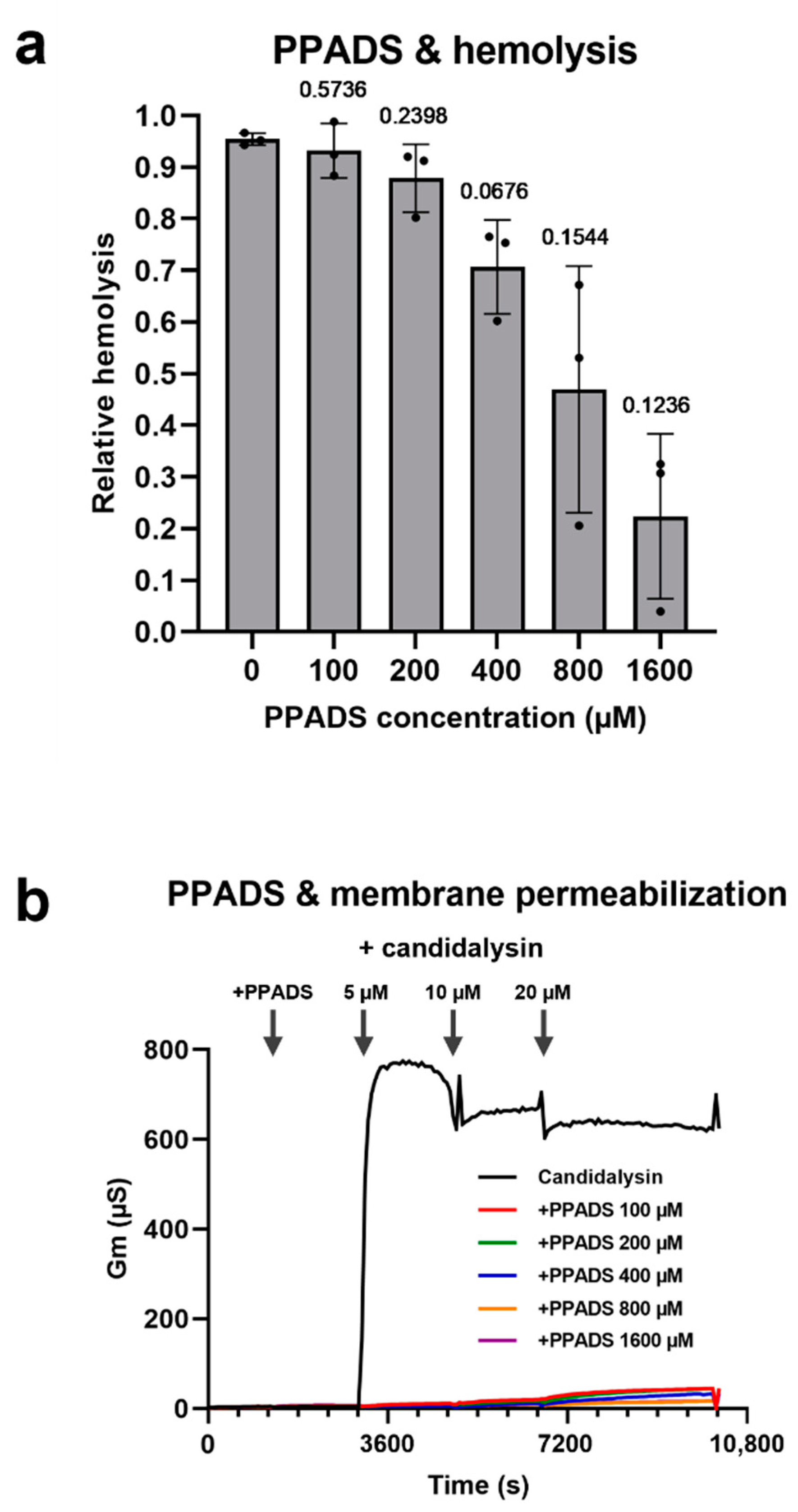
2.5. Purinergic Receptor Antagonists Protect RBCs from Candidalysin-Induced Lysis
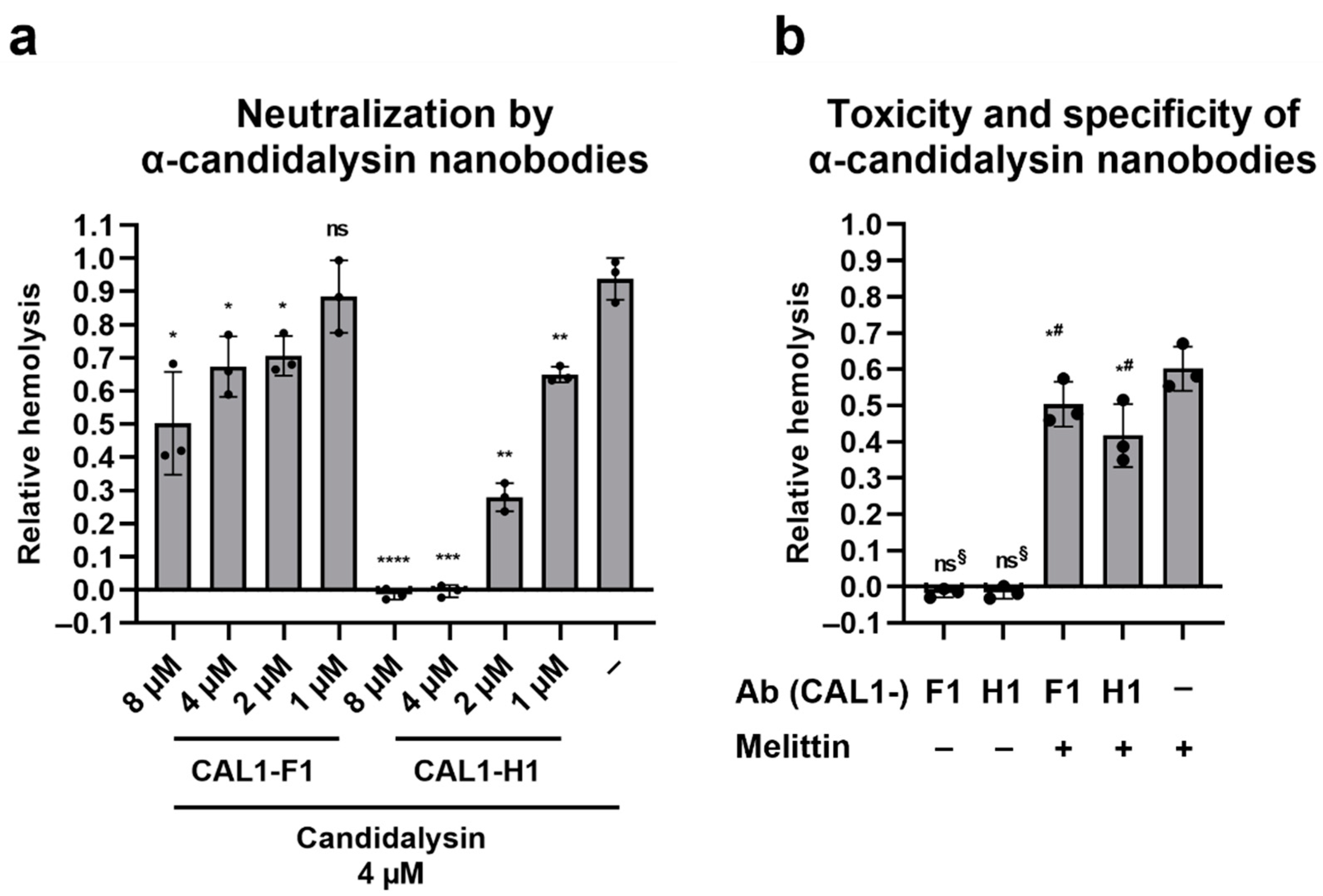
2.6. Llama-Derived Anti-Candidalysin Nanobodies Neutralize the Hemolytic Potential of Candidalysin
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Culture Conditions
4.2. RBC Preparation
4.3. Handling of Peptides
4.4. Hemolysis Assay
4.5. Determination of Membrane Permeabilization
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bougnoux, M.E.; Diogo, D.; Francois, N.; Sendid, B.; Veirmeire, S.; Colombel, J.F.; Bouchier, C.; Van Kruiningen, H.; d’Enfert, C.; Poulain, D. Multilocus sequence typing reveals intrafamilial transmission and microevolutions of Candida albicans isolates from the human digestive tract. J. Clin. Microbiol. 2006, 44, 1810–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Pasqualotto, A.C.; Nedel, W.L.; Machado, T.S.; Severo, L.C. Risk factors and outcome for nosocomial breakthrough candidaemia. J. Infect. 2006, 52, 216–222. [Google Scholar] [CrossRef]
- Allert, S.; Forster, T.M.; Svensson, C.M.; Richardson, J.P.; Pawlik, T.; Hebecker, B.; Rudolphi, S.; Juraschitz, M.; Schaller, M.; Blagojevic, M.; et al. Candida albicans-induced epithelial damage mediates translocation through intestinal barriers. mBio 2018, 9, e00915-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyes, D.L.; Wilson, D.; Richardson, J.P.; Mogavero, S.; Tang, S.X.; Wernecke, J.; Hofs, S.; Gratacap, R.L.; Robbins, J.; Runglall, M.; et al. Candidalysin is a fungal peptide toxin critical for mucosal infection. Nature 2016, 532, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Swidergall, M.; Khalaji, M.; Solis, N.V.; Moyes, D.L.; Drummond, R.A.; Hube, B.; Lionakis, M.S.; Murdoch, C.; Filler, S.G.; Naglik, J.R. Candidalysin is required for neutrophil recruitment and virulence during systemic Candida albicans infection. J. Infect. Dis. 2019, 220, 1477–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, R.A.; Swamydas, M.; Oikonomou, V.; Zhai, B.; Dambuza, I.M.; Schaefer, B.C.; Bohrer, A.C.; Mayer-Barber, K.D.; Lira, S.A.; Iwakura, Y.; et al. CARD9(+) microglia promote antifungal immunity via IL-1beta- and CXCL1-mediated neutrophil recruitment. Nat. Immunol. 2019, 20, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.P.; Willems, H.M.E.; Moyes, D.L.; Shoaie, S.; Barker, K.S.; Tan, S.L.; Palmer, G.E.; Hube, B.; Naglik, J.R.; Peters, B.M. Candidalysin drives epithelial signaling, neutrophil recruitment, and immunopathology at the vaginal mucosa. Infect. Immun. 2017, 86, e00645-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasper, L.; Konig, A.; Koenig, P.A.; Gresnigt, M.S.; Westman, J.; Drummond, R.A.; Lionakis, M.S.; Gross, O.; Ruland, J.; Naglik, J.R.; et al. The fungal peptide toxin candidalysin activates the NLRP3 inflammasome and causes cytolysis in mononuclear phagocytes. Nat. Commun. 2018, 9, 4260. [Google Scholar] [CrossRef] [Green Version]
- Bader, O.; Krauke, Y.; Hube, B. Processing of predicted substrates of fungal Kex2 proteinases from Candida albicans, C. glabrata, Saccharomyces cerevisiae and Pichia pastoris. BMC Microbiol. 2008, 8, 116. [Google Scholar] [CrossRef]
- Richardson, J.P.; Mogavero, S.; Moyes, D.L.; Blagojevic, M.; Kruger, T.; Verma, A.H.; Coleman, B.M.; De La Cruz Diaz, J.; Schulz, D.; Ponde, N.O.; et al. Processing of Candida albicans Ece1p is critical for candidalysin maturation and fungal virulence. mBio 2018, 9, e02178-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birse, C.E.; Irwin, M.Y.; Fonzi, W.A.; Sypherd, P.S. Cloning and characterization of ECE1, a gene expressed in association with cell elongation of the dimorphic pathogen Candida albicans. Infect. Immun. 1993, 61, 3648–3655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.; Albrecht-Eckardt, D.; Brunke, S.; Hube, B.; Hunniger, K.; Kurzai, O. A core filamentation response network in Candida albicans is restricted to eight genes. PLoS ONE 2013, 8, e58613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, R.S.; Brunke, S.; Albrecht, A.; Thewes, S.; Laue, M.; Edwards, J.E.; Filler, S.G.; Hube, B. The hyphal-associated adhesin and invasin Als3 of Candida albicans mediates iron acquisition from host ferritin. PLoS Pathog. 2008, 4, e1000217. [Google Scholar] [CrossRef] [PubMed]
- Manns, J.M.; Mosser, D.M.; Buckley, H.R. Production of a hemolytic factor by Candida albicans. Infect. Immun. 1994, 62, 5154–5156. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, W.T.; Nakao, N.; Mikami, T.; Matsumoto, T. Hemoglobin is utilized by Candida albicans in the hyphal form but not yeast form. Biochem. Biophys. Res. Commun. 1997, 232, 350–353. [Google Scholar] [CrossRef]
- Watanabe, T.; Takano, M.; Murakami, M.; Tanaka, H.; Matsuhisa, A.; Nakao, N.; Mikami, T.; Suzuki, M.; Matsumoto, T. Characterization of a haemolytic factor from Candida albicans. Microbiology 1999, 145, 689–694. [Google Scholar] [CrossRef] [Green Version]
- Sacristan, B.; Blanco, M.T.; Galan-Ladero, M.A.; Blanco, J.; Perez-Giraldo, C.; Gomez-Garcia, A.C. Aspartyl proteinase, phospholipase, hemolytic activities and biofilm production of Candida albicans isolated from bronchial aspirates of ICU patients. Med. Mycol. 2011, 49, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Rossoni, R.D.; Barbosa, J.O.; Vilela, S.F.; Jorge, A.O.; Junqueira, J.C. Comparison of the hemolytic activity between C. albicans and non-albicans Candida species. Braz. Oral Res. 2013, 27, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Riceto, E.B.; Menezes Rde, P.; Penatti, M.P.; Pedroso Rdos, S. Enzymatic and hemolytic activity in different Candida species. Rev. Iberoam. Micol. 2015, 32, 79–82. [Google Scholar] [CrossRef]
- Zakikhany, K.; Naglik, J.R.; Schmidt-Westhausen, A.; Holland, G.; Schaller, M.; Hube, B. In vivo transcript profiling of Candida albicans identifies a gene essential for interepithelial dissemination. Cell Microbiol. 2007, 9, 2938–2954. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Mackman, N.; Menestrina, G.; Gray, L.; Hugo, F.; Seeger, W.; Holland, I.B. The hemolysin of Escherichia coli. Eur. J. Epidemiol. 1988, 4, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [CrossRef]
- Skals, M.; Jorgensen, N.R.; Leipziger, J.; Praetorius, H.A. Alpha-hemolysin from Escherichia coli uses endogenous amplification through P2X receptor activation to induce hemolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 4030–4035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skals, M.; Leipziger, J.; Praetorius, H.A. Haemolysis induced by alpha-toxin from Staphylococcus aureus requires P2X receptor activation. Pflugers Arch. 2011, 462, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Mogavero, S.; Sauer, F.M.; Brunke, S.; Allert, S.; Schulz, D.; Wisgott, S.; Jablonowski, N.; Elshafee, O.; Kruger, T.; Kniemeyer, O.; et al. Candidalysin delivery to the invasion pocket is critical for host epithelial damage induced by Candida albicans. Cell Microbiol. 2021, 23, e13378. [Google Scholar] [CrossRef] [PubMed]
- Salvin, S.B. Hemolysin from the yeast-like phases of some pathogenic fungi. Proc. Soc. Exp. Biol. Med. 1951, 76, 852–854. [Google Scholar] [CrossRef] [PubMed]
- Kuprowski, M. The influence of the quality of agar on the occurrence of haemolysis in cultures of Candida albicans. Zentralbl. Veterinarmed B 1966, 13, 364–368. [Google Scholar] [CrossRef]
- Buxton, R. Blood Agar Plates and Hemolysis Protocols; ASM: Washington, DC, USA, 2005. [Google Scholar]
- Matrai, A.A.; Varga, G.; Tanczos, B.; Barath, B.; Varga, A.; Horvath, L.; Bereczky, Z.; Deak, A.; Nemeth, N. In vitro effects of temperature on red blood cell deformability and membrane stability in human and various vertebrate species. Clin. Hemorheol. Microcirc. 2021, 78, 291–300. [Google Scholar] [CrossRef]
- Pendrak, M.L.; Roberts, D.D. Hemoglobin is an effective inducer of hyphal differentiation in Candida albicans. Med. Mycol. 2007, 45, 61–71. [Google Scholar] [CrossRef]
- Cardoso, P.H.d.O.; Boleti, A.P.d.A.; Silva, P.S.e.; Mukoyama, L.T.H.; Guindo, A.S.; Moraes, L.F.R.N.d.; Oliveira, C.F.R.d.; Macedo, M.L.R.; Carvalho, C.M.E.; de Castro, A.P.; et al. Evaluation of a novel synthetic peptide derived from cytolytic mycotoxin candidalysin. Toxins 2022, 14, 696. [Google Scholar] [CrossRef] [PubMed]
- Moors, M.A.; Stull, T.L.; Blank, K.J.; Buckley, H.R.; Mosser, D.M. A role for complement receptor-like molecules in iron acquisition by Candida albicans. J. Exp. Med. 1992, 175, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Kornitzer, D.; Roy, U. Pathways of heme utilization in fungi. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118817. [Google Scholar] [CrossRef]
- Mak, P. Hemocidins in a functional and structural context of human antimicrobial peptides. Front. Biosci. 2008, 13, 6859–6871. [Google Scholar] [CrossRef] [PubMed]
- Mak, P.; Wojcik, K.; Wicherek, L.; Suder, P.; Dubin, A. Antibacterial hemoglobin peptides in human menstrual blood. Peptides 2004, 25, 1839–1847. [Google Scholar] [CrossRef]
- Bochenska, O.; Rapala-Kozik, M.; Wolak, N.; Bras, G.; Kozik, A.; Dubin, A.; Aoki, W.; Ueda, M.; Mak, P. Secreted aspartic peptidases of Candida albicans liberate bactericidal hemocidins from human hemoglobin. Peptides 2013, 48, 49–58. [Google Scholar] [CrossRef]
- Kirouac, M.; Vachon, V.; Quievy, D.; Schwartz, J.L.; Laprade, R. Protease inhibitors fail to prevent pore formation by the activated Bacillus thuringiensis toxin Cry1Aa in insect brush border membrane vesicles. Appl. Environ. Microbiol. 2006, 72, 506–515. [Google Scholar] [CrossRef] [Green Version]
- Chaigne-Delalande, B.; Lenardo, M.J. Divalent cation signaling in immune cells. Trends Immunol. 2014, 35, 332–344. [Google Scholar] [CrossRef] [Green Version]
- Legge, D.G.; Shortman, K. The effect of pH on the volume, density and shape of erythrocytes and thymic lymphocytes. Br. J. Haematol. 1968, 14, 323–335. [Google Scholar] [CrossRef]
- de Bernard, M.; Papini, E.; de Filippis, V.; Gottardi, E.; Telford, J.; Manetti, R.; Fontana, A.; Rappuoli, R.; Montecucco, C. Low pH activates the vacuolating toxin of Helicobacter pylori, which becomes acid and pepsin resistant. J. Biol. Chem. 1995, 270, 23937–23940. [Google Scholar] [CrossRef]
- Schwiering, M.; Husmann, M.; Hellmann, N. P2X-receptor antagonists inhibit the interaction of S. aureus Hemolysin A with membranes. Toxins 2017, 9, 332. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Description | Internal Name | Relevant Genotype | References |
|---|---|---|---|---|
| Wt (isogenic) | BWP17/CIp30 Isogenic wild type | M1477 | ura3::λimm434/ura3::λimm434 iro1::λimm434/iro1::λimm434 his1::hisG/his1::hisG arg4::hisG/arg4::hisG RPS1/rps1::(URA3-HIS1-ARG4) | [21] |
| ece1Δ/Δ | ece1 double knock-out mutant | M2057 | ece1::ARG4/ece1::HIS1 RPS1/rps1::URA3 | [5] |
| ece1Δ/Δ + ECE1 | ECE1 revertant | M2059 | ece1::ARG4/ece1::HIS1 RPS1/rps1::(URA3-ECE1) | [5] |
| ΔP3 | Mutant lacking only the P3 sequence (all other Ece1 peptides expressed) | M2174 | ece1::ARG4/ece1::HIS1 RPS1/rps1::(URA3-ECE1Δ184–279) | [5] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mogavero, S.; Höfs, S.; Lauer, A.N.; Müller, R.; Brunke, S.; Allert, S.; Gerwien, F.; Groth, S.; Dolk, E.; Wilson, D.; et al. Candidalysin Is the Hemolytic Factor of Candida albicans. Toxins 2022, 14, 874. https://doi.org/10.3390/toxins14120874
Mogavero S, Höfs S, Lauer AN, Müller R, Brunke S, Allert S, Gerwien F, Groth S, Dolk E, Wilson D, et al. Candidalysin Is the Hemolytic Factor of Candida albicans. Toxins. 2022; 14(12):874. https://doi.org/10.3390/toxins14120874
Chicago/Turabian StyleMogavero, Selene, Sarah Höfs, Alexa N. Lauer, Rita Müller, Sascha Brunke, Stefanie Allert, Franziska Gerwien, Sabrina Groth, Edward Dolk, Duncan Wilson, and et al. 2022. "Candidalysin Is the Hemolytic Factor of Candida albicans" Toxins 14, no. 12: 874. https://doi.org/10.3390/toxins14120874
APA StyleMogavero, S., Höfs, S., Lauer, A. N., Müller, R., Brunke, S., Allert, S., Gerwien, F., Groth, S., Dolk, E., Wilson, D., Gutsmann, T., & Hube, B. (2022). Candidalysin Is the Hemolytic Factor of Candida albicans. Toxins, 14(12), 874. https://doi.org/10.3390/toxins14120874