Recombinant Expression of ABCC2 Variants Confirms the Importance of Mutations in Extracellular Loop 4 for Cry1F Resistance in Fall Armyworm



Abstract
:
1. Introduction
2. Results
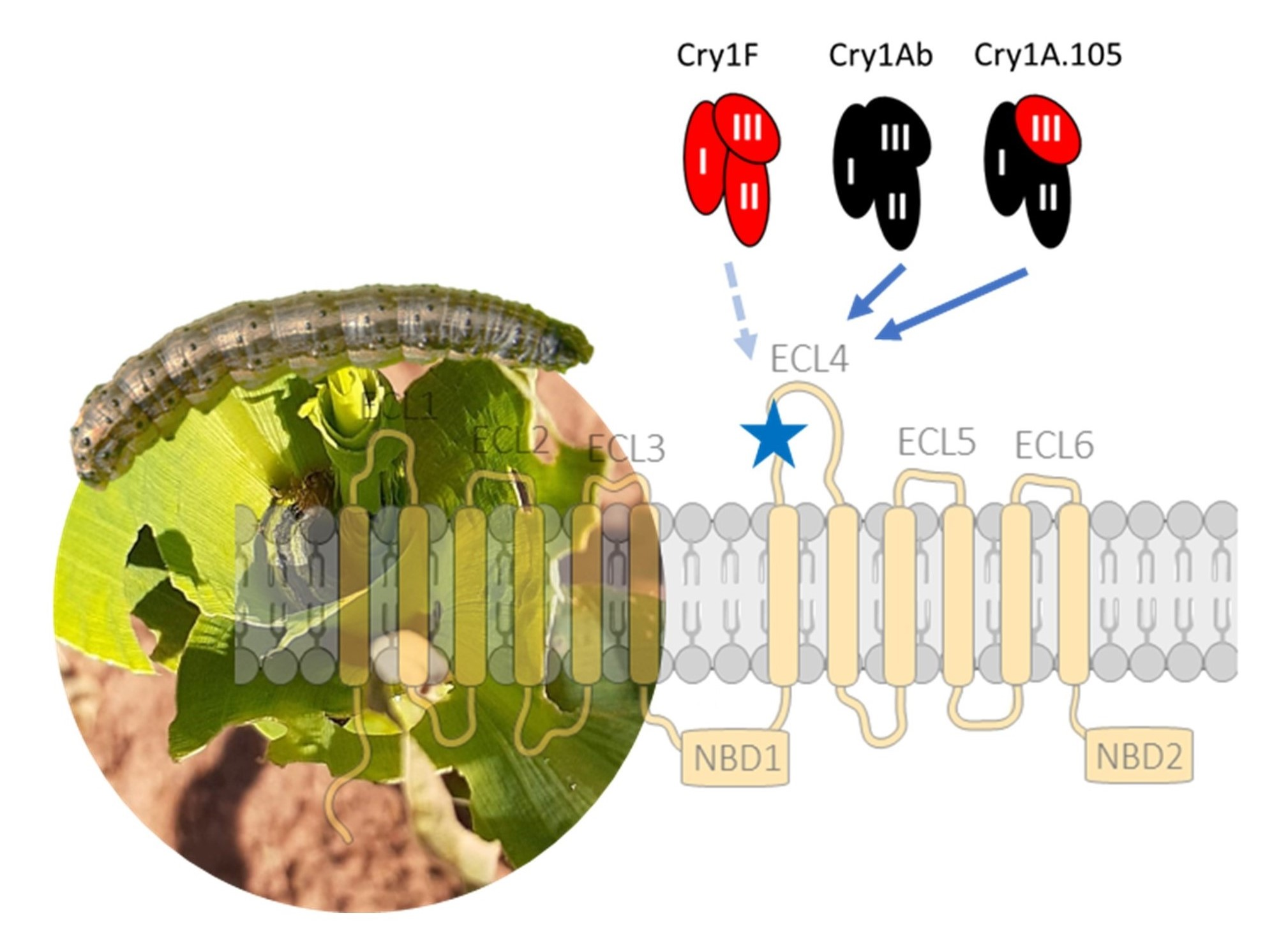
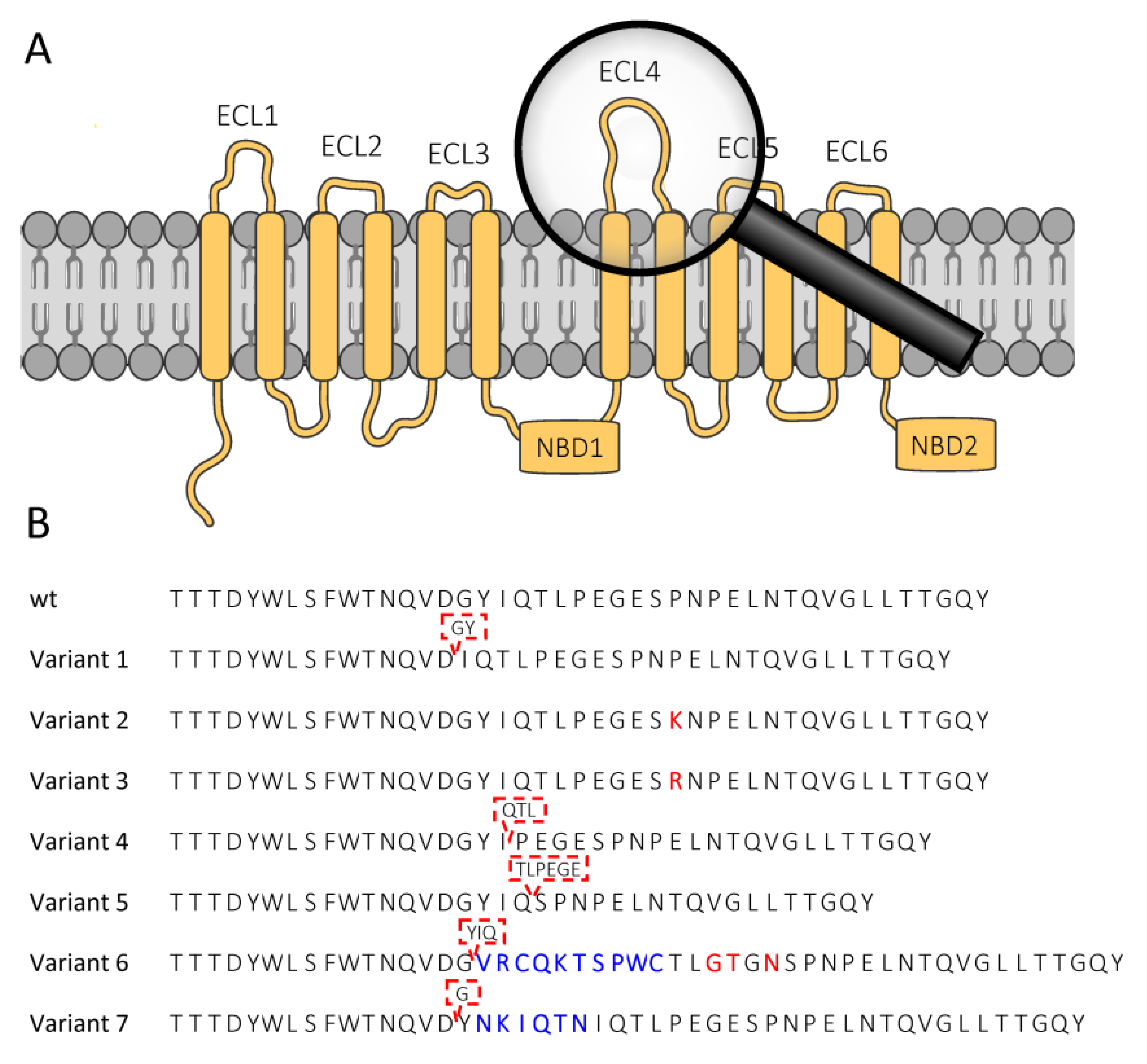
2.1. Effect of SfABCC2 ECL4 Mutations on Cry1F, Cry1A.105 and Cry1Ab Toxicity
2.2. SfABCC3 Is a Receptor for Cry1Ab but Not Cry1F
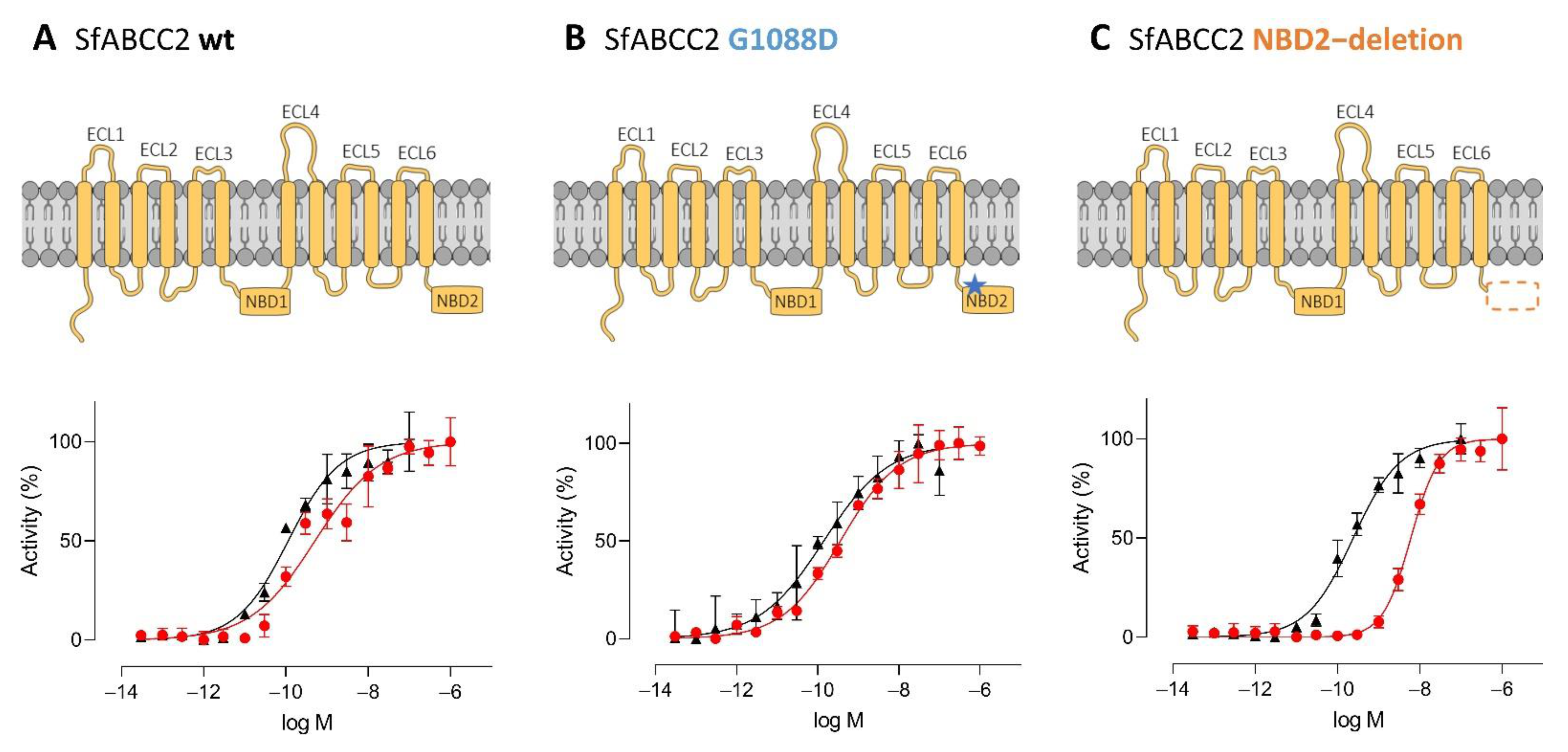
2.3. ABCC2 Gating Activity has no Effect on Cry1 Activity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insect Cell Culture
5.2. Expression of ABC-Transporter Variants in Sf9 Cells
5.3. SYTOXTM Orange Cytotoxicity Assay
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Sparks, A.N. A Review of the Biology of the Fall Armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Barros, E.M.; Torres, J.B.; Ruberson, J.R.; Oliveira, M.D. Development of Spodoptera frugiperda on Different Hosts and Damage to Reproductive Structures in Cotton. Entomol. Exp. Appl. 2010, 137, 237–245. [Google Scholar] [CrossRef]
- Overton, K.; Maino, J.L.; Day, R.; Umina, P.A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O.L. Global Crop Impacts, Yield Losses and Action Thresholds for Fall Armyworm (Spodoptera frugiperda): A Review. Crop Prot. 2021, 145, 105641. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niassy, S.; Agbodzavu, M.K.; Kimathi, E.; Mutune, B.; Abdel-Rahman, E.F.M.; Salifu, D.; Hailu, G.; Belayneh, Y.T.; Felege, E.; Tonnang, H.E.Z.; et al. Bioecology of Fall Armyworm Spodoptera frugiperda (J. E. Smith), Its Management and Potential Patterns of Seasonal Spread in Africa. PLoS ONE 2021, 16, e0249042. [Google Scholar] [CrossRef] [PubMed]
- Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.; Maruthi, M.S.; Pavithra, H.B.; Hegbe, K.; Navi, S.; Prabhu, S.T.; Goergen, G.E. First Report of the Fall Armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an Alien Invasive Pest on Maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Ma, J.; Wang, Y.-P.; Wu, M.-F.; Gao, B.-Y.; Liu, J.; Lee, G.-S.; Otuka, A.; Hu, G. High Risk of the Fall Armyworm Invading Japan and the Korean Peninsula via Overseas Migration. J. Appl. Entomol. 2019, 143, 911–920. [Google Scholar] [CrossRef]
- Tepa-Yotto, G.T.; Tonnang, H.E.Z.; Goergen, G.; Subramanian, S.; Kimathi, E.; Abdel-Rahman, E.M.; Flø, D.; Thunes, K.H.; Fiaboe, K.K.M.; Niassy, S.; et al. Global Habitat Suitability of Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae): Key Parasitoids Considered for Its Biological Control. Insects 2021, 12, 273. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Q.; Zhang, H.; Wu, K. Spread of Invasive Migratory Pest Spodoptera frugiperda and Management Practices throughout China. J. Integr. Agric. 2021, 20, 637–645. [Google Scholar] [CrossRef]
- Burtet, L.M.; Bernardi, O.; Melo, A.A.; Pes, M.P.; Strahl, T.T.; Guedes, J.V. Managing Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), with Bt Maize and Insecticides in Southern Brazil. Pest Manag. Sci. 2017, 73, 2569–2577. [Google Scholar] [CrossRef]
- Kulye, M.; Mehlhorn, S.; Boaventura, D.; Godley, N.; Venkatesh, S.; Rudrappa, T.; Charan, T.; Rathi, D.; Nauen, R. Baseline Susceptibility of Spodoptera frugiperda Populations Collected in India towards Different Chemical Classes of Insecticides. Insects 2021, 12, 758. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Sánchez, F.A.; Rivera, G.; Bocanegra-García, V.; Martínez-Padrón, H.Y.; Berrones-Morales, M.; Niño-García, N.; Herrera-Mayorga, V. Advances in Control Strategies against Spodoptera frugiperda. A Review. Molecules 2021, 26, 5587. [Google Scholar] [CrossRef]
- Bernardi, O.; Bernardi, D.; Amado, D.; Sousa, R.S.; Fatoretto, J.; Medeiros, F.C.L.; Conville, J.; Burd, T.; Omoto, C. Resistance Risk Assessment of Spodoptera frugiperda (Lepidoptera: Noctuidae) and Diatraea saccharalis (Lepidoptera: Crambidae) to Vip3Aa20 Insecticidal Protein Expressed in Corn. J. Econ. Entomol. 2015, 108, 2711–2719. [Google Scholar] [CrossRef]
- Moscardini, V.F.; Marques, L.H.; Santos, A.C.; Rossetto, J.; Silva, O.A.B.N.; Rampazzo, P.E.; Castro, B.A. Efficacy of Bacillus thuringiensis (Bt) Maize Expressing Cry1F, Cry1A.105, Cry2Ab2 and Vip3Aa20 Proteins to Manage the Fall Armyworm (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2020, 137, 105269. [Google Scholar] [CrossRef]
- Siebert, M.W.; Babock1, J.M.; Nolting, S.; Santos, A.C.; Jr, J.J.A.; Neese, P.A.; King, J.E.; Jenkins, J.N.; McCarty, J.; Lorenz, G.M.; et al. Efficacy of Cry1F Insecticidal Protein in Maize and Cotton for Control of Fall Armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 2008, 91, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Machado, E.; Junior, G.; Führ, F.M.; Zago, S.; Marques, L.; Santos, A.; Nowatzki, T.; Dahmer, M.; Omoto, C.; Bernardi, O. Cross-Crop Resistance of Spodoptera frugiperda Selected on Bt Maize to Genetically-Modified Soybean Expressing Cry1Ac and Cry1F Proteins in Brazil. Sci. Rep. 2020, 10, 10080. [Google Scholar] [CrossRef]
- Peterson, B.; Bezuidenhout, C.C.; Van den Berg, J. An Overview of Mechanisms of Cry Toxin Resistance in Lepidopteran Insects. J. Econ. Entomol. 2017, 110, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect Resistance to Bt Crops: Lessons from the First Billion Acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Carrière, Y. Evaluating Cross-Resistance between Vip and Cry Toxins of Bacillus thuringiensis. J. Econ. Entomol. 2020, 113, 553–561. [Google Scholar] [CrossRef]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and Characterization of Field Resistance to Bt Maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; dos Santos, A.C.; Omoto, C. Field-Evolved Resistance to Cry1F Maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Chandrasena, D.I.; Signorini, A.M.; Abratti, G.; Storer, N.P.; Olaciregui, M.L.; Alves, A.P.; Pilcher, C.D. Characterization of Field-Evolved Resistance to Bacillus thuringiensis-Derived Cry1F δ-Endotoxin in Spodoptera frugiperda Populations from Argentina. Pest Manag. Sci. 2018, 74, 746–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F. Resistance of the Fall Armyworm, Spodoptera frugiperda, to Transgenic Bacillus thuringiensis Cry1F Corn in the Americas: Lessons and Implications for Bt Corn IRM in China. Insect Sci. 2021, 28, 574–589. [Google Scholar] [CrossRef]
- Bernardi, D.; Salmeron, E.; Horikoshi, R.J.; Bernardi, O.; Dourado, P.M.; Carvalho, R.A.; Martinelli, S.; Head, G.P.; Omoto, C. Cross-Resistance between Cry1 Proteins in Fall Armyworm (Spodoptera frugiperda) May Affect the Durability of Current Pyramided Bt Maize Hybrids in Brazil. PLoS ONE 2015, 10, e0140130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Amaya, O.F.; Tavares, C.S.; Monteiro, H.M.; Teixeira, T.P.M.; Guedes, R.N.C.; Alves, A.P.; Pereira, E.J.G. Genetic Basis of Cry1F Resistance in Two Brazilian Populations of Fall Armyworm, Spodoptera frugiperda. Crop Prot. 2016, 81, 154–162. [Google Scholar] [CrossRef]
- Vélez, A.M.; Spencer, T.A.; Alves, A.P.; Moellenbeck, D.; Meagher, R.L.; Chirakkal, H.; Siegfried, B.D. Inheritance of Cry1F Resistance, Cross-Resistance and Frequency of Resistant Alleles in Spodoptera frugiperda (Lepidoptera: Noctuidae). Bull. Entomol. Res. 2013, 103, 700–713. [Google Scholar] [CrossRef] [Green Version]
- Horikoshi, R.J.; Bernardi, O.; Amaral, S.D.A.; Miraldo, L.L.; Durigan, M.R.; Bernardi, D.; Silva, S.S.; Omoto, C. Lack of Relevant Cross-Resistance to Bt Insecticide XenTari in Strains of Spodoptera frugiperda (J. E. Smith) Resistant to Bt Maize. J. Invertebr. Pathol. 2019, 161, 1–6. [Google Scholar] [CrossRef]
- Heckel, D.G. How Do Toxins from Bacillus thuringiensis Kill Insects? An Evolutionary Perspective. Arch. Insect Biochem. Physiol. 2020, 104, e21673. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.L.D.A.; Soccol, V.T.; Soccol, C.R. Bacillus thuringiensis: Mechanism of Action, Resistance, and New Applications: A Review. Crit. Rev. Biotechnol. 2016, 36, 317–326. [Google Scholar] [CrossRef]
- Adang, M.J.; Crickmore, N.; Jurat-Fuentes, J.L. Chapter Two—Diversity of Bacillus thuringiensis Crystal Toxins and Mechanism of Action. In Advances in Insect Physiology; Dhadialla, T.S., Gill, S.S., Eds.; Insect Midgut and Insecticidal Proteins; Academic Press: Cambridge, MA, USA, 2014; Volume 47, pp. 39–87. [Google Scholar]
- Heckel, D.G. The Essential and Enigmatic Role of ABC Transporters in Bt Resistance of Noctuids and Other Insect Pests of Agriculture. Insects 2021, 12, 389. [Google Scholar] [CrossRef]
- Sato, R.; Adegawa, S.; Li, X.; Tanaka, S.; Endo, H. Function and Role of ATP-Binding Cassette Transporters as Receptors for 3D-Cry Toxins. Toxins 2019, 11, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahan, L.J.; Pauchet, Y.; Vogel, H.; Heckel, D.G. An ABC Transporter Mutation Is Correlated with Insect Resistance to Bacillus thuringiensis Cry1Ac Toxin. PLoS Genet. 2010, 6, e1001248. [Google Scholar] [CrossRef] [Green Version]
- Baxter, S.W.; Badenes-Pérez, F.R.; Morrison, A.; Vogel, H.; Crickmore, N.; Kain, W.; Wang, P.; Heckel, D.G.; Jiggins, C.D. Parallel Evolution of Bacillus thuringiensis Toxin Resistance in Lepidoptera. Genetics 2011, 189, 675–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsumi, S.; Miyamoto, K.; Yamamoto, K.; Narukawa, J.; Kawai, S.; Sezutsu, H.; Kobayashi, I.; Uchino, K.; Tamura, T.; Mita, K.; et al. Single Amino Acid Mutation in an ATP-Binding Cassette Transporter Gene Causes Resistance to Bt Toxin Cry1Ab in the Silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2012, 109, E1591–E1598. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; González-Martínez, R.M.; Navarro-Cerrillo, G.; Chakroun, M.; Kim, Y.; Ziarsolo, P.; Blanca, J.; Cañizares, J.; Ferré, J.; Herrero, S. ABCC Transporters Mediate Insect Resistance to Multiple Bt Toxins Revealed by Bulk Segregant Analysis. BMC Biol. 2014, 12, 46. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Zhang, T.; Liu, C.; Heckel, D.G.; Li, X.; Tabashnik, B.E.; Wu, K. Mis-Splicing of the ABCC2 Gene Linked with Bt Toxin Resistance in Helicoverpa Armigera. Sci. Rep. 2014, 4, 6184. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Hasler, J.; Meagher, R.; Nagoshi, R.; Hietala, L.; Huang, F.; Narva, K.; Jurat-Fuentes, J.L. Mechanism and DNA-Based Detection of Field-Evolved Resistance to Transgenic Bt Corn in Fall Armyworm (Spodoptera frugiperda). Sci. Rep. 2017, 7, 10877. [Google Scholar] [CrossRef] [Green Version]
- Flagel, L.; Lee, Y.W.; Wanjugi, H.; Swarup, S.; Brown, A.; Wang, J.; Kraft, E.; Greenplate, J.; Simmons, J.; Adams, N.; et al. Mutational Disruption of the ABCC2 Gene in Fall Armyworm, Spodoptera frugiperda, Confers Resistance to the Cry1Fa and Cry1A.105 Insecticidal Proteins. Sci. Rep. 2018, 8, 7255. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Tao, J.; Li, Q.; Cheng, Y.; Sun, X.; Wu, K.; Xiao, Y. Genome Editing of the SfABCC2 Gene Confers Resistance to Cry1F Toxin from Bacillus thuringiensis in Spodoptera frugiperda. J. Integr. Agric. 2021, 20, 815–820. [Google Scholar] [CrossRef]
- Boaventura, D.; Martin, M.; Pozzebon, A.; Mota-Sanchez, D.; Nauen, R. Monitoring of Target-Site Mutations Conferring Insecticide Resistance in Spodoptera frugiperda. Insects 2020, 11, 545. [Google Scholar] [CrossRef]
- Schlum, K.; Lamour, K.; Tandy, P.; Emrich, S.J.; de Bortoli, C.P.; Rao, T.; Viteri Dillon, D.M.; Linares-Ramirez, A.M.; Jurat-Fuentes, J.L. Genetic Screening to Identify Candidate Resistance Alleles to Cry1F Corn in Fall Armyworm Using Targeted Sequencing. Insects 2021, 12, 618. [Google Scholar] [CrossRef]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular Characterization of Cry1F Resistance in Fall Armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2020, 116, 103280. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, M.; Wang, L.; Wang, H.; Xia, Z.; Yang, Y.; Bravo, A.; Soberón, M.; Xiao, Y.; Liu, K. SfABCC2 Transporter Extracellular Loops 2 and 4 Are Responsible for the Cry1Fa Insecticidal Specificity against Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2021, 135, 103608. [Google Scholar] [CrossRef]
- Yainna, S.; Nègre, N.; Silvie, P.J.; Brévault, T.; Tay, W.T.; Gordon, K.; dAlençon, E.; Walsh, T.; Nam, K. Geographic Monitoring of Insecticide Resistance Mutations in Native and Invasive Populations of the Fall Armyworm. Insects 2021, 12, 468. [Google Scholar] [CrossRef]
- Guan, F.; Zhang, J.; Shen, H.; Wang, X.; Padovan, A.; Walsh, T.K.; Tay, W.T.; Gordon, K.H.J.; James, W.; Czepak, C.; et al. Whole-Genome Sequencing to Detect Mutations Associated with Resistance to Insecticides and Bt Proteins in Spodoptera frugiperda. Insect Sci. 2021, 28, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Fu, X.; Nageotte, J.R.; Silverman, J.; Bretsnyder, E.C.; Chen, D.; Rydel, T.J.; Bean, G.J.; Li, K.S.; et al. Bacillus thuringiensis Cry1Da_7 and Cry1B.868 Protein Interactions with Novel Receptors Allow Control of Resistant Fall Armyworms, Spodoptera frugiperda (J.E. Smith). Appl. Env. Microbiol. 2019, 85, e00579-19. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Yang, Y.; Shan, Y.; Chakrabarty, S.; Cheng, Y.; Soberón, M.; Bravo, A.; Liu, K.; Wu, K.; Xiao, Y. Two ABC Transporters Are Differentially Involved in the Toxicity of Two Bacillus thuringiensis Cry1 Toxins to the Invasive Crop-Pest Spodoptera frugiperda (J. E. Smith). Pest Manag. Sci. 2021, 77, 1492–1501. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xu, Y.; Zuo, Y.; Yang, Y.; Tabashnik, B.E.; Wu, Y. Evaluation of Five Candidate Receptors for Three Bt Toxins in the Beet Armyworm Using CRISPR-Mediated Gene Knockouts. Insect Biochem. Mol. Biol. 2020, 121, 103361. [Google Scholar] [CrossRef]
- Wang, Y.; Adegawa, S.; Miyamoto, K.; Takasu, Y.; Iizuka, T.; Wada, S.; Mang, D.; Li, X.; Kim, S.; Sato, R.; et al. ATP-Binding Cassette Transporter Subfamily C Members 2, 3 and Cadherin Protein Are Susceptibility-Determining Factors in Bombyx mori for Multiple Bacillus thuringiensis Cry1 Toxins. Insect Biochem. Mol. Biol. 2021, 139, 103649. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, H.; Zhao, S.; Huang, J.; Yang, Y.; Tabashnik, B.E.; Wu, Y. Functional Redundancy of Two ABC Transporter Proteins in Mediating Toxicity of Bacillus thuringiensis to Cotton Bollworm. PLoS Pathog. 2020, 16, e1008427. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Tanaka, S.; Adegawa, S.; Ichino, F.; Tabunoki, H.; Kikuta, S.; Sato, R. Extracellular Loop Structures in Silkworm ABCC Transporters Determine Their Specificities for Bacillus thuringiensis Cry Toxins. J. Biol. Chem. 2018, 293, 8569–8577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, H.; Azuma, M.; Adegawa, S.; Kikuta, S.; Sato, R. Water Influx via Aquaporin Directly Determines Necrotic Cell Death Induced by the Bacillus thuringiensis Cry Toxin. FEBS Lett. 2017, 591, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, H.; Tanaka, S.; Imamura, K.; Adegawa, S.; Kikuta, S.; Sato, R. Cry Toxin Specificities of Insect ABCC Transporters Closely Related to Lepidopteran ABCC2 Transporters. Peptides 2017, 98, 86–92. [Google Scholar] [CrossRef]
- Tanaka, S.; Endo, H.; Adegawa, S.; Iizuka, A.; Imamura, K.; Kikuta, S.; Sato, R. Bombyx mori ABC Transporter C2 Structures Responsible for the Receptor Function of Bacillus thuringiensis Cry1Aa Toxin. Insect Biochem. Mol. Biol. 2017, 91, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Adegawa, S.; Yamaguchi, N.; Sato, R. The Base and Root of Domain II Loops of Cry Toxins Contribute to Binding to Bombyx mori ABC Transporter C2. FEBS J. 2021, 289, 965–984. [Google Scholar] [CrossRef]
- Jerga, A.; Evdokimov, A.G.; Moshiri, F.; Haas, J.A.; Chen, M.; Clinton, W.; Fu, X.; Halls, C.; Jimenez-Juarez, N.; Kretzler, C.N.; et al. Disabled Insecticidal Proteins: A Novel Tool to Understand Differences in Insect Receptor Utilization. Insect Biochem. Mol. Biol. 2019, 105, 79–88. [Google Scholar] [CrossRef]
- Boaventura, D.; Buer, B.; Hamaekers, N.; Maiwald, F.; Nauen, R. Toxicological and Molecular Profiling of Insecticide Resistance in a Brazilian Strain of Fall Armyworm Resistant to Bt Cry1 Proteins. Pest Manag. Sci. 2021, 77, 3713–3726. [Google Scholar] [CrossRef]
- Zhang, J.; Jin, M.; Yang, Y.; Liu, L.; Yang, Y.; Gómez, I.; Bravo, A.; Soberón, M.; Xiao, Y.; Liu, K. The Cadherin Protein Is Not Involved in Susceptibility to Bacillus thuringiensis Cry1Ab or Cry1Fa Toxins in Spodoptera frugiperda. Toxins 2020, 12, 375. [Google Scholar] [CrossRef]
- Liu, L.; Chen, Z.; Yang, Y.; Xiao, Y.; Liu, C.; Ma, Y.; Soberón, M.; Bravo, A.; Yang, Y.; Liu, K. A Single Amino Acid Polymorphism in ABCC2 Loop 1 Is Responsible for Differential Toxicity of Bacillus thuringiensis Cry1Ac Toxin in Different Spodoptera (Noctuidae) Species. Insect Biochem. Mol. Biol. 2018, 100, 59–65. [Google Scholar] [CrossRef]
- Pinos, D.; Martínez-Solís, M.; Herrero, S.; Ferré, J.; Hernández-Martínez, P. The Spodoptera exigua ABCC2 Acts as a Cry1A Receptor Independently of Its Nucleotide Binding Domain II. Toxins 2019, 11, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Habbersett, R.C.; Cordek, J.M.; Nolan, J.P.; Yoshida, T.M.; Jett, J.H.; Marrone, B.L. Development of a Mechanism-Based, DNA Staining Protocol Using SYTOX Orange Nucleic Acid Stain and DNA Fragment Sizing Flow Cytometry. Anal. Biochem. 2000, 286, 138–148. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| ABC-Transporter Variant | Cry Protein | EC50 (nM) a | 95% CI b | Slope (±SE) | RR c |
|---|---|---|---|---|---|
| SfABCC2 wt. [GenBank OL955491] | Cry1F | 0.395 | 0.324–0.482 | 0.616 (0.035) | - |
| Cry1Ab | 0.110 | 0.0746–0.163 | 0.638 (0.073) | - | |
| Cry1A.105 | 0.0946 | 0.0700–0.128 | 0.492 (0.036) | - | |
| SfABCC2 Variant 1 (GY-deletion) [GenBank OL955490] | Cry1F | >1000 | - | - | >2532 |
| Cry1Ab | 0.497 | 0.379–0.652 | 0.624 (0.049) | 4.5 | |
| Cry1A.105 | 0.340 | 0.242–0.476 | 0.632 (0.062) | 3.6 | |
| SfABCC2 Variant 2 (P799K) [GenBank OL955488] | Cry1F | 249 | 192–324 | 1.17 (0.10) | 630 |
| Cry1Ab | 0.980 | 0.845–1.14 | 0.828 (0.045) | 8.9 | |
| Cry1A.105 | 1.03 | 0.868–1.21 | 0.844 (0.053) | 10.9 | |
| SfABCC2 Variant 3 (P799R) [GenBank OL955487] | Cry1F | 649 | 269–1569 | 0.922 (0.118) | 1643 |
| Cry1Ab | 1.32 | 1.12–1.56 | 0.828 (0.051) | 12.0 | |
| Cry1A.105 | 1.58 | 1.38–1.80 | 0.789 (0.036) | 16.7 | |
| SfABCC2 Variant 4 (QTL-deletion) [GenBank OL955486] | Cry1F | >1000 | - | - | >2532 |
| Cry1Ab | 0.975 | 0.826–1.15 | 0.675 (0.034) | 8.9 | |
| Cry1A.105 | 1.48 | 1.24–1.78 | 0.735 (0.044) | 15.6 | |
| SfABCC2 Variant 5 (TLPEGE-deletion) [GenBank OL955485] | Cry1F | >1000 | - | - | >2532 |
| Cry1Ab | 5.09 | 4.21–6.16 | 0.864 (0.061) | 46.3 | |
| Cry1A.105 | 6.41 | 5.65–7.28 | 1.11 (0.067) | 67.8 | |
| SfABCC2 Variant 6 (long-insert) [GenBank OL955484] | Cry1F | >1000 | - | - | >2532 |
| Cry1Ab | 5.47 | 4.79–6.25 | 1.00 (0.057) | 49.7 | |
| Cry1A.105 | 6.61 | 6.03–7.26 | 1.36 (0.072) | 69.9 | |
| SfABCC2 Variant 7 (short-insert) [GenBank OL955489] | Cry1F | >1000 | - | - | >2532 |
| Cry1Ab | 20.9 | 17.8–24.4 | 0.984 (0.063) | 190.0 | |
| Cry1A.105 | 28.5 | 23.5–34.5 | 1.06 (0.089) | 301.3 | |
| SfABCC3 wt. [sequence from [48]] | Cry1F | >1000 | - | - | - |
| Cry1Ab | 3.32 | 2.96–3.72 | 1.30 (0.085) | - | |
| Cry1A.105 | <10 d | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franz, L.; Raming, K.; Nauen, R. Recombinant Expression of ABCC2 Variants Confirms the Importance of Mutations in Extracellular Loop 4 for Cry1F Resistance in Fall Armyworm. Toxins 2022, 14, 157. https://doi.org/10.3390/toxins14020157
Franz L, Raming K, Nauen R. Recombinant Expression of ABCC2 Variants Confirms the Importance of Mutations in Extracellular Loop 4 for Cry1F Resistance in Fall Armyworm. Toxins. 2022; 14(2):157. https://doi.org/10.3390/toxins14020157
Chicago/Turabian StyleFranz, Laura, Klaus Raming, and Ralf Nauen. 2022. "Recombinant Expression of ABCC2 Variants Confirms the Importance of Mutations in Extracellular Loop 4 for Cry1F Resistance in Fall Armyworm" Toxins 14, no. 2: 157. https://doi.org/10.3390/toxins14020157
APA StyleFranz, L., Raming, K., & Nauen, R. (2022). Recombinant Expression of ABCC2 Variants Confirms the Importance of Mutations in Extracellular Loop 4 for Cry1F Resistance in Fall Armyworm. Toxins, 14(2), 157. https://doi.org/10.3390/toxins14020157