Isolation, Purification, and Characterization of a Laccase-Degrading Aflatoxin B1 from Bacillus amyloliquefaciens B10


Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of AFB1-Degrading Bacteria
2.2. Localisation of Degradation Active Substances
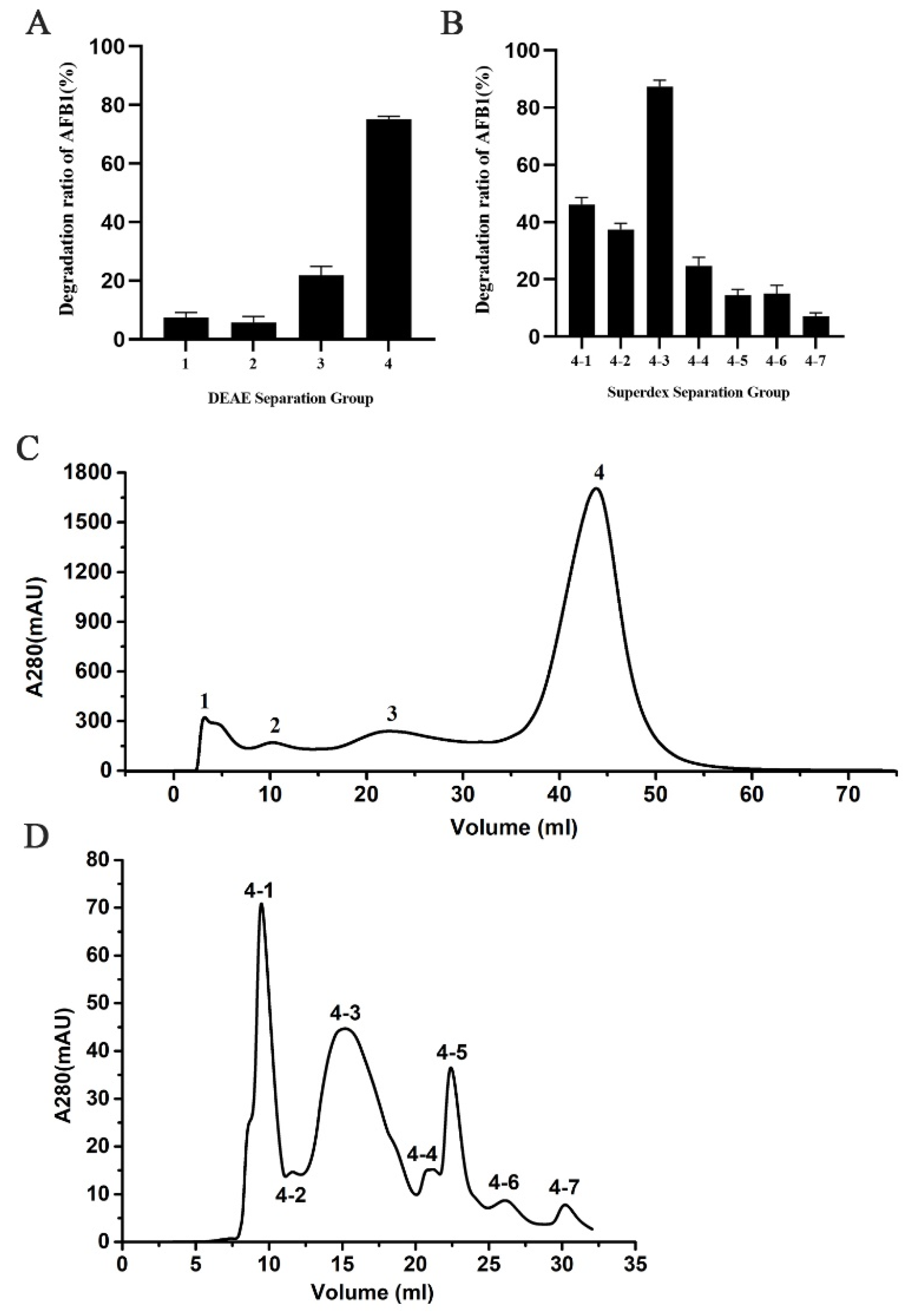
2.3. Isolation of AFB1-Degrading Proteins
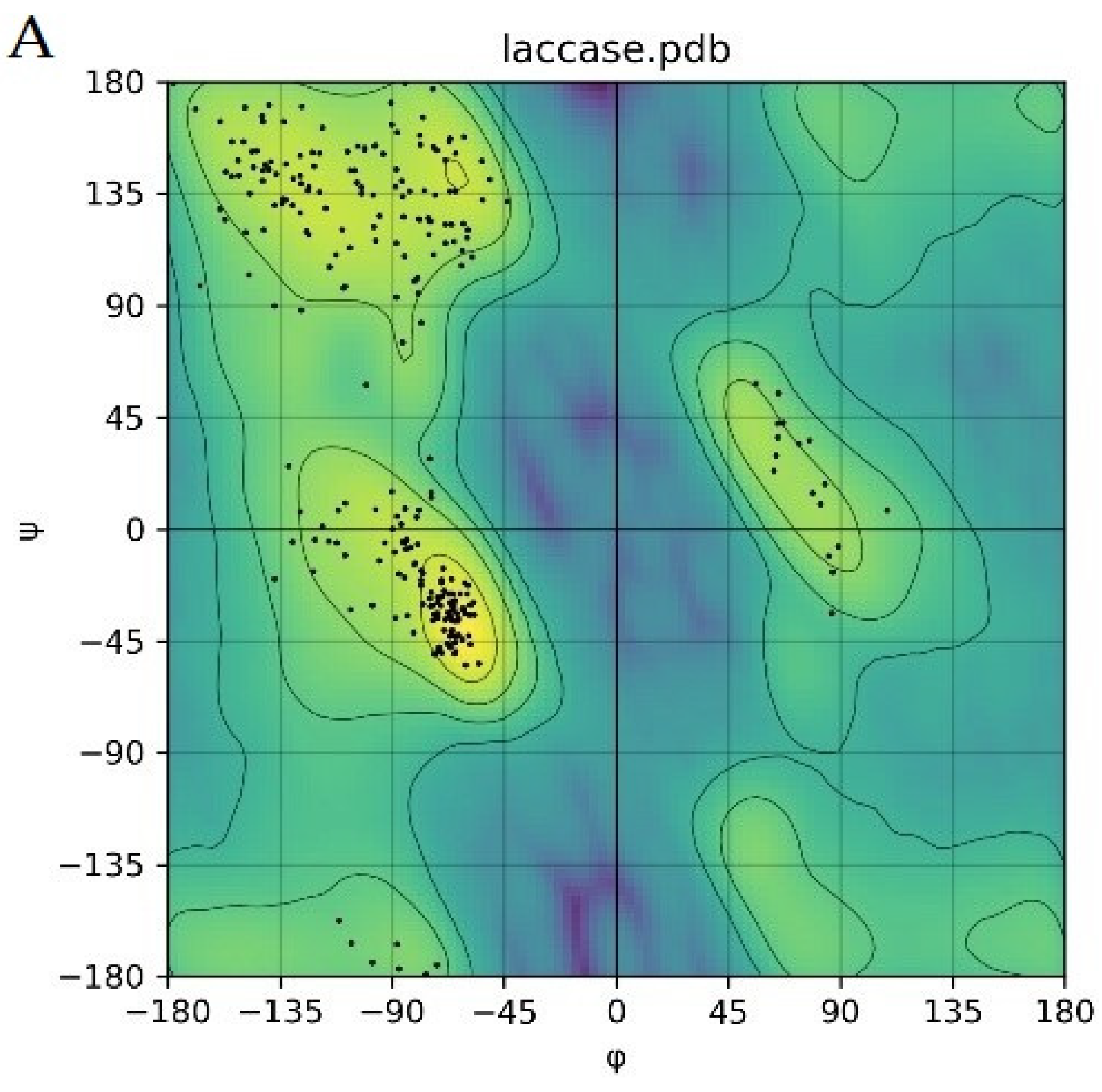
2.4. Molecular Docking for Function Prediction
2.5. Cloning, Expression, and Purification of Laccase from Strain B10
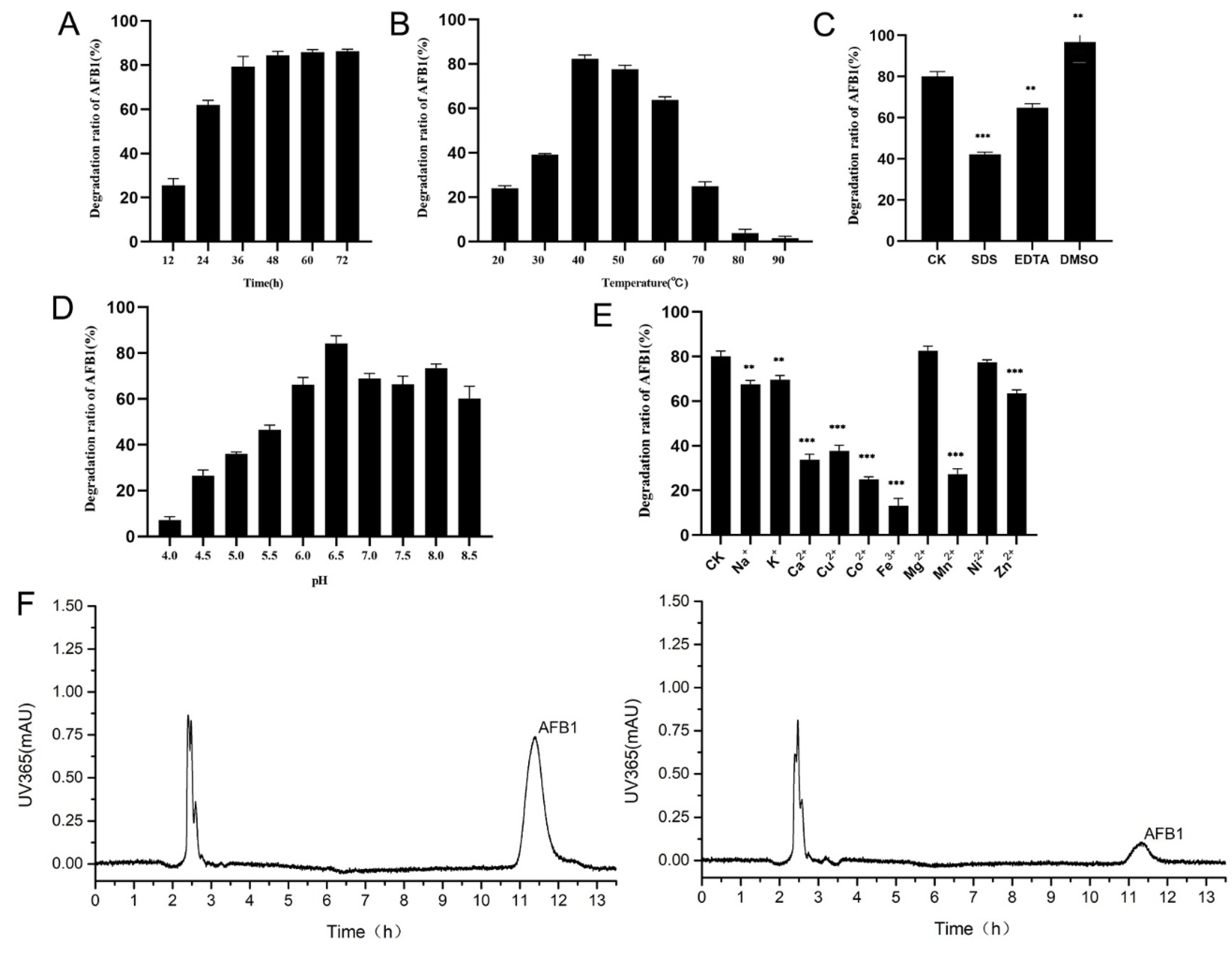
2.6. Efficiency of AFB1 Degradation by the Recombinant Laccase and the Effects of Different Conditions on AFB1 Degradation
2.7. Site-Specific Mutagenesis of B10 Laccases
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Isolation of AFB1-Degrading Strains
5.3. Localisation of AFB1-Degrading Active Substances by Strain B10
5.4. Crude Extraction of Protein from Culture Medium
5.5. Isolation and Identification of AFB1-Degrading Proteins
5.6. Structural Modelling of the Recombinant Laccase and Molecular Docking
5.7. Gene Cloning, Protein Expression, and Purification of the Recombinant Laccase
5.8. Efficiency of AFB1 Degradation by a Recombinant Laccase and Its Effect on AFB1-Degrading Activity under Different Conditions
5.9. Targeted Mutagenesis of the Recombinant Laccase
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kensler, T.W.; Roebuck, B.D.; Wogan, G.N.; Groopman, J.D. Aflatoxin: A 50-Year Odyssey of Mechanistic and Translational Toxicology. Toxicol. Sci. 2011, 120, S28–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peraica, M.; Radić, B.; Lucić, A.; Pavlović, M. Toxic effects of mycotoxins in humans. Bull. World Health Organ. 1999, 77, 754–766. [Google Scholar] [PubMed]
- Nazhand, A.; Durazzo, A.; Lucarini, M.; Souto, E.B.; Santini, A. Characteristics, Occurrence, Detection and Detoxification of Aflatoxins in Foods and Feeds. Foods 2020, 9, 644. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Adebo, O.; Njobeh, P.; Gbashi, S.; Nwinyi, O.; Mavumengwana, V. Review on microbial degradation of aflatoxins. Crit. Rev. Food Sci. Nutr. 2017, 57, 3208–3217. [Google Scholar] [CrossRef] [PubMed]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’H, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From preven-tion in the field to detoxification by adsorption or transformation. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2011, 28, 1590–1609. [Google Scholar] [CrossRef] [PubMed]
- Mishra, H.N.; Das, C. A review on biological control and metabolism of aflatoxin. Crit. Rev. Food Sci. Nutr. 2003, 43, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Naresh, M.; Olsen, M. Mycotoxins in food: Detection and control; Woodhead Publishing: Oxford, UK, 2004; p. 289. [Google Scholar]
- Kološová, A.; Stroka, J. Substances for reduction of the contamination of feed by mycotoxins: A review. World Mycotoxin J. 2011, 4, 225–256. [Google Scholar] [CrossRef]
- Guan, Y.; Chen, J.; Nepovimova, E.; Long, M.; Wu, W.; Kuca, K. Aflatoxin Detoxification Using Microorganisms and Enzymes. Toxins 2021, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Mate, D.; Alcalde, M. Laccase: A multi-purpose biocatalyst at the forefront of biotechnology. Microb. Biotechnol. 2016, 10, 1457–1467. [Google Scholar] [CrossRef]
- Xu, F.; Shin, W.; Brown, S.H.; Wahleithner, J.A.; Sundaram, U.M.; Solomon, E.I. A study of a series of recombinant fungal laccases and bilirubin oxidase that exhibit significant differences in redox potential, substrate specificity, and stability. Biochim. Biophys. Acta Protein Struct. Mol. Enzym. 1996, 1292, 303–311. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Gorbacheva, M.A.; Shleev, S.V.; Yaropolov, A.I. “Blue” laccases. Biochemistry 2007, 72, 1136–1150. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Pawlik, A.; Swiderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkolazka, A.; Paszczynski, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci. 2020, 21, 996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Li, R.; Ng, T.B.; Lai, Y.; Yang, J.; Ye, X. A New Laccase of Lac 2 from the White Rot Fungus Cerrena unicolor 6884 and Lac 2-Mediated Degradation of Aflatoxin B1. Toxins 2020, 12, 476. [Google Scholar] [CrossRef] [PubMed]
- Hullo, M.-F.; Moszer, I.; Danchin, A.; Martin-Verstraete, I. CotA of Bacillus subtilis Is a Copper-Dependent Laccase. J. Bacteriol. 2001, 183, 5426–5430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, U.N.; Singh, P.; Pandey, V.P.; Kumar, A. Structure–function relationship among bacterial, fungal and plant laccases. J. Mol. Catal. Enzym. 2011, 68, 117–128. [Google Scholar] [CrossRef]
- Guo, Y.; Qin, X.; Tang, Y.; Ma, Q.; Zhang, J.; Zhao, L. CotA laccase, a novel aflatoxin oxidase from Bacillus licheniformis, transforms aflatoxin B1 to aflatoxin Q1 and epi-aflatoxin Q1. Food Chem. 2020, 325, 126877. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Renaud, J.B.; Rosini, E.; Pollegioni, L.; Vignali, E.; Haidukowski, M.; Sumarah, M.W.; Logrieco, A.F.; Mule, G. Enzy-matic transformation of aflatoxin B1 by Rh_DypB peroxidase and characterization of the reaction products. Chemosphere 2020, 250, 126296. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.L.; Beima, K.M.; Bammler, T.K.; Riley, R.T.; Voss, K.A. 9.24—Hepatotoxic Mycotoxins. In Comprehensive Toxicology, 2nd ed.; Mcqueen, C.A., Ed.; Elsevier: Oxford, UK, 2010; pp. 527–569. [Google Scholar]
- Hsieh, D.P.H.; Salhab, A.S.; Wong, J.J.; Yang, S.L. Toxicity of aflatoxin Q1 as evaluated with the chicken embryo and bacteri-al auxotrophs. Toxicol. Appl. Pharm. 1974, 30, 237–242. [Google Scholar] [CrossRef]
- Martins, L.O.; Durao, P.; Brissos, V.; Lindley, P.F. Laccases of prokaryotic origin: Enzymes at the interface of protein science and protein technology. Celll. Mol. Life Sci. 2015, 72, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- SantAnna, B.M.M.; Marbach, P.P.A.; Rojas-Herrera, M.; de Souza, J.T.; Roque, M.R.A.; Queiroz, A.T.L. High-Quality Draft Genome Sequence of Bacillus amyloliquefaciens Strain 629, an Endophyte from Theobroma cacao. Genome Announc. 2015, 3, e01325-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Zheng, J.; Zhang, Z.; Peng, D.; Sun, M. Nematicidal spore-forming Bacilli share similar virulence factors and mechanisms. Sci. Rep. 2016, 6, 31341. [Google Scholar] [CrossRef]
- Wang, X.; Luo, C.; Chen, Z. Genome Sequence of the Plant Growth-Promoting Rhizobacterium Bacillus sp. Strain 916. J. Bacteriol. 2012, 194, 5467–5468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamid, A.S.; Tesfamariam, I.G.; Zhang, Y.; Zhang, Z.G. Aflatoxin B1-induced hepatocellular carcinoma in developing coun-tries: Geographical distribution, mechanism of action and prevention. Oncol. Lett. 2013, 5, 1087–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Gupta, S.; Gupta, M. A review study on biological ill effects and health hazards of aflatoxins. Asian J. Adv. Med. Sci. 2021, 3, 1–8. [Google Scholar]
- Cai, M.; Qian, Y.; Chen, N.; Ling, T.; Wang, J.; Jiang, H.; Wang, X.; Qi, K.; Zhou, Y. Detoxification of aflatoxin B1 by Stenotrophomonas sp. CW117 and characterization the thermophilic degradation process. Environ. Pollut. 2020, 261, 114178. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Eisa, A.M.; Sangare, L.; Zhao, Y.; Selvaraj, J.N.; Xing, F.; Wang, Y.; Yang, H.; Liu, Y. Novel Aflatoxin-Degrading En-zyme from Bacillus shackletonii L7. Toxins 2017, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Suresh, G.; Cabezudo, I.; Pulicharla, R.; Cuprys, A.; Rouissi, T.; Brar, S.K. Biodegradation of aflatoxin B1 with cell-free ex-tracts of Trametes versicolor and Bacillus subtilis. Res. Vet. Sci. 2020, 133, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Yan, H.; Yin, C.; Liu, Y.; Xu, Q.; Liu, X.; Zhang, Z. Effective Biodegradation of Aflatoxin B1 Using the Bacillus licheniformis (BL010) Strain. Toxins 2018, 10, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, X.; Wang, Y.; Zhou, Q.; Li, M.; Hu, H.; Ma, Y.; Chen, X.; Ni, J.; Zhao, W.; Huang, S.; et al. Biological Degradation of Aflatoxin B1 by Cell-Free Extracts of Bacillus velezensis DY3108 with Broad PH Stability and Excellent Thermostability. Toxins 2018, 10, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Guan, S.; Gao, X.; Ma, Q.; Lei, Y.; Bai, X.; Ji, C. Preparation, purification and characteristics of an aflatoxin degradation enzyme from Myxococcus fulvus ANSM068. J. Appl. Microbiol. 2010, 110, 147–155. [Google Scholar] [CrossRef]
- Motomura, M.; Toyomasu, T.; Mizuno, K.; Shinozawa, T. Purification and characterization of an aflatoxin degradation enzyme from Pleurotus ostreatus. Microbiol. Res. 2003, 158, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Del Alamo, D.; Govaerts, C.; Mchaourab, H.S. AlphaFold2 predicts the inward-facing conformation of the multidrug transporter LmrP. Proteins Struct. Funct. Bioinform. 2021, 89, 1226–1228. [Google Scholar] [CrossRef] [PubMed]
- Cramer, P. AlphaFold2 and the future of structural biology. Nat. Struct. Mol. Biol. 2021, 28, 704–705. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.J.; Courtney, J.M.; Shen, Y.; Ying, J.; Bax, A. Concordance of X-ray and AlphaFold2 Models of SARS-CoV-2 Main Protease with Residual Dipolar Couplings Measured in Solution. J. Am. Chem. Soc. 2021, 143, 19306–19310. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mao, H.; Hu, C.; Tron, T.; Lin, J.; Wang, J.; Sun, B. Molecular docking studies and in vitro degradation of four afla-toxins (AFB1, AFB2, AFG1, and AFG2) by a recombinant laccase from Saccharomyces cerevisiae. J. Food Sci. 2020, 85, 1353–1360. [Google Scholar] [CrossRef]
- Jardon-Xicotencatl, S.; Díaz-Torres, R.; Marroquin-Cardona, A.; Villarreal-Barajas, T.; Méndez-Albores, A. Detoxification of Aflatoxin-Contaminated Maize by Neutral Electrolyzed Oxidizing Water. Toxins 2015, 7, 4294–4314. [Google Scholar] [CrossRef] [Green Version]
- Novoa, C.; Dhoke, G.V.; Mate, D.; Martinez, R.; Haarmann, T.; Schreiter, M.; Eidner, J.; Schwerdtfeger, R.; Lorenz, P.; Davari, M.D.; et al. KnowVolution of a Fungal Laccase toward Alkaline pH. ChemBioChem 2019, 20, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Liu, Q.; Ma, J.; Liao, H.; Xiong, X.; Zhang, K.; Wang, T.; Liu, X.; Xu, T.; Yuan, S.; et al. An acid-stable bacterial laccase identified from the endophyte Pantoea ananatis Sd-1 genome exhibiting lignin degradation and dye decolor-ization abilities. Biotechnol. Lett. 2015, 37, 2279–2288. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.H.; Lin, M.C.; Lee, C.C.; Yu, S.M.; Wang, A.H.; Ho, T.D. Enhancement of laccase activity by pre-incubation with or-ganic solvents. Sci. Rep. 2019, 9, 9754. [Google Scholar] [CrossRef] [PubMed]
- Klibanov, A.M. Improving enzymes by using them in organic solvents. Nature 2001, 409, 241–246. [Google Scholar] [CrossRef]
- Kim, Y.; Maltseva, N.; Dementieva, I.; Collart, F.; Holzle, D.; Joachimiak, A. Crystal Structure of Hypothetical Protein YfiH From Shigella flexneri at 2 Å Resolution. Proteins Struct. Funct. Bioinform. 2006, 63, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Gunne, M.; Al-Sultani, D.; Urlacher, V.B. Enhancement of copper content and specific activity of CotA laccase from Bacillus licheniformis by coexpression with CopZ copper chaperone in E. coli. J. Biotechnol. 2013, 168, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Cader, M.Z.; de Almeida, R.R.; West, J.A.; Sewell, G.W.; Md-Ibrahim, M.N.; Reikine, S.; Sirago, G.; Unger, L.W.; Iglesias-Romero, A.B.; Ramshorn, K.; et al. FAMIN Is a Multifunctional Purine Enzyme Enabling the Purine Nucle-otide Cycle. Cell 2020, 180, 278–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liu, X.; Zhao, J.; Yue, Q.; Yan, Y.; Gao, Z.; Dong, Y.; Zhang, Z.; Fan, Y.; Tian, J.; et al. Crystal struc-tures of multicopper oxidase CueO G304K mutant: Structural basis of the increased laccase activity. Sci. Rep. 2018, 8, 14252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djoko, K.Y.; Chong, L.X.; Wedd, A.G.; Xiao, Z. Reaction Mechanisms of the Multicopper Oxidase CueO from Escherichia coli Support Its Functional Role as a Cuprous Oxidase. J. Am. Chem. Soc. 2010, 132, 2005–2015. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K.; Komori, H.; Ueki, Y.; Konno, Y.; Kamitaka, Y.; Kurose, S.; Tsujimura, S.; Higuchi, Y.; Kano, K.; Seo, D.; et al. Structure and Function of the Engineered Multicopper Oxidase CueO from Escherichia coli—Deletion of the Methionine-Rich Helical Region Covering the Substrate-Binding Site. J. Mol. Biol. 2007, 373, 141–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Result |
|---|---|
| Methyl Red assay | + |
| Anaerobic | − |
| Oxidase | + |
| Contact enzyme assay | + |
| V-P test | + |
| Indole test | − |
| Starch hydrolysis | + |
| Glucose fermentation | + |
| Glucose gas production | − |
| Gelatine liquefaction | + |
| Citrate | + |
| Xylose | + |
| Nitrate | + |
| Propionate | − |
| 7% NaCl growth | + |
| Compound Name | Structure | Docking Score (kcal/mol) | Hydrogen Bond | Hydrophobic Effect | ||
|---|---|---|---|---|---|---|
| Enzyme Residues | Distance D-A (Å) | Acceptor Atom | ||||
| AFB1 |  | −5.60 | Trp-152 | 3.17 | O-2 | His-87 |
| Lys-153 | 2.82 | O-1 | ||||
| Arg-268 | 2.89 | O-5 | ||||
| AFB2 |  | −6.82 | Tyr-191 | 3.50 | O-2 | His-87 |
| Arg-268 | 2.63 | O-5 | ||||
| 2.62 | O-5 | |||||
| AFG1 |  | −6.58 | Lys-153 | 2.84 | O-6 | His-87 |
| 3.15 | O-8 | |||||
| Arg-268 | 2.79 | O-1 | ||||
| 2.77 | O-1 | |||||
| AFG2 |  | −5.31 | Lys-153 | 2.77 | O-5 | His-87 |
| Arg-268 | 2.65 | O-2 | ||||
| 3.09 | O-1 | |||||
| AFM1 |  | −6.30 | Tyr-191 | 3.16 | O-2 | His-87 |
| His-259 | 2.87 | O-3 | ||||
| Arg-268 | 2.84 | O-7 | ||||
| 2.82 | O-7 | |||||
| AFM2 |  | −6.13 | Trp-152 | 2.78 | O-7 | His-87 Cys-132 |
| Lys-153 | 2.76 | O-6 | ||||
| Tyr-191 | 2.65 | O-3 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, D.; Wen, J.; Lu, G.; Li, T.; Long, M. Isolation, Purification, and Characterization of a Laccase-Degrading Aflatoxin B1 from Bacillus amyloliquefaciens B10. Toxins 2022, 14, 250. https://doi.org/10.3390/toxins14040250
Xiong D, Wen J, Lu G, Li T, Long M. Isolation, Purification, and Characterization of a Laccase-Degrading Aflatoxin B1 from Bacillus amyloliquefaciens B10. Toxins. 2022; 14(4):250. https://doi.org/10.3390/toxins14040250
Chicago/Turabian StyleXiong, Dongwei, Jun Wen, Gen Lu, Tianxi Li, and Miao Long. 2022. "Isolation, Purification, and Characterization of a Laccase-Degrading Aflatoxin B1 from Bacillus amyloliquefaciens B10" Toxins 14, no. 4: 250. https://doi.org/10.3390/toxins14040250
APA StyleXiong, D., Wen, J., Lu, G., Li, T., & Long, M. (2022). Isolation, Purification, and Characterization of a Laccase-Degrading Aflatoxin B1 from Bacillus amyloliquefaciens B10. Toxins, 14(4), 250. https://doi.org/10.3390/toxins14040250