Whole-Transcriptome Analysis of Non-Coding RNA Alteration in Porcine Alveolar Macrophage Exposed to Aflatoxin B1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
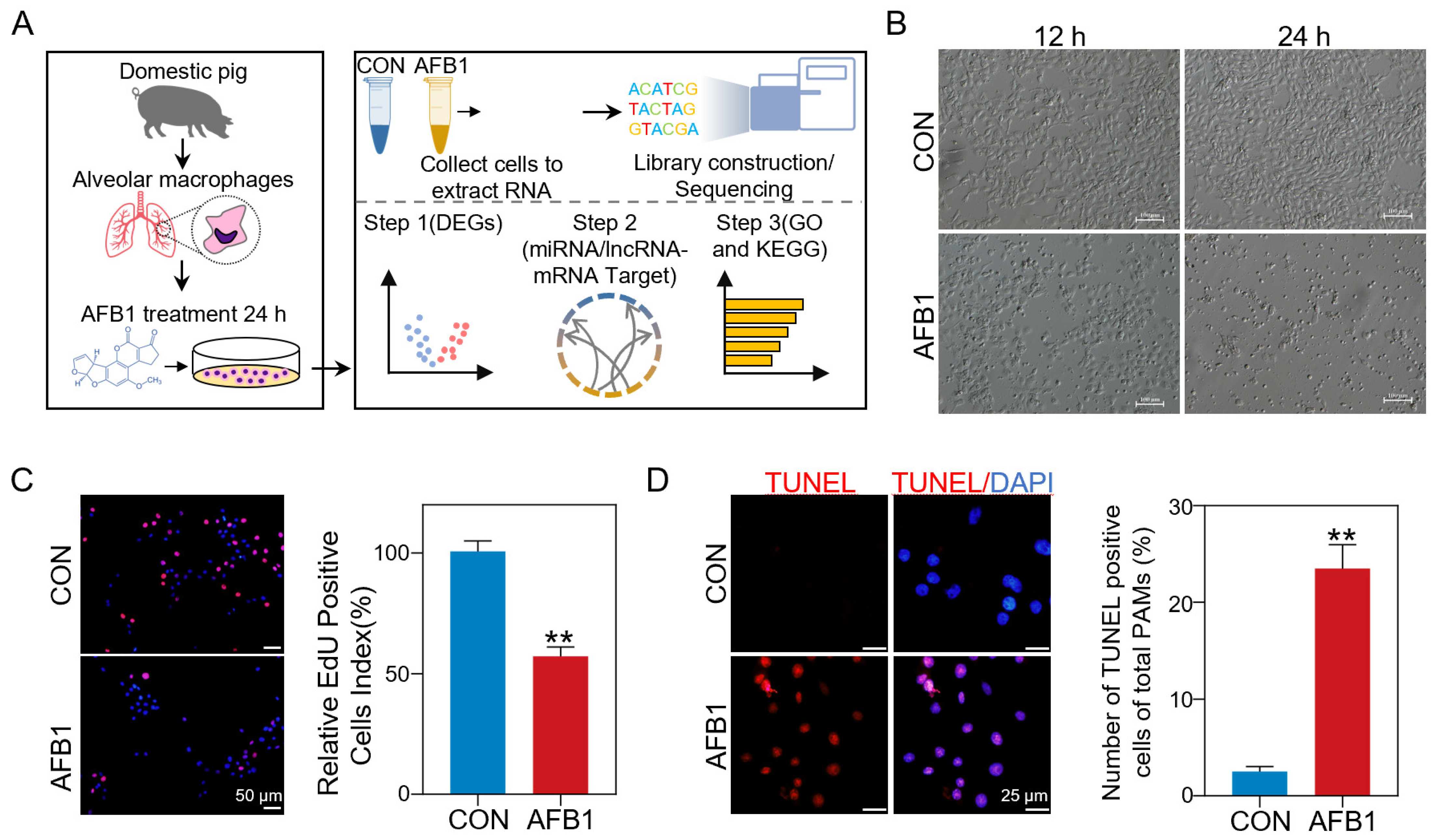
2.1. AFB1 Inhibited Cell Proliferation and Induced Cell Apoptosis
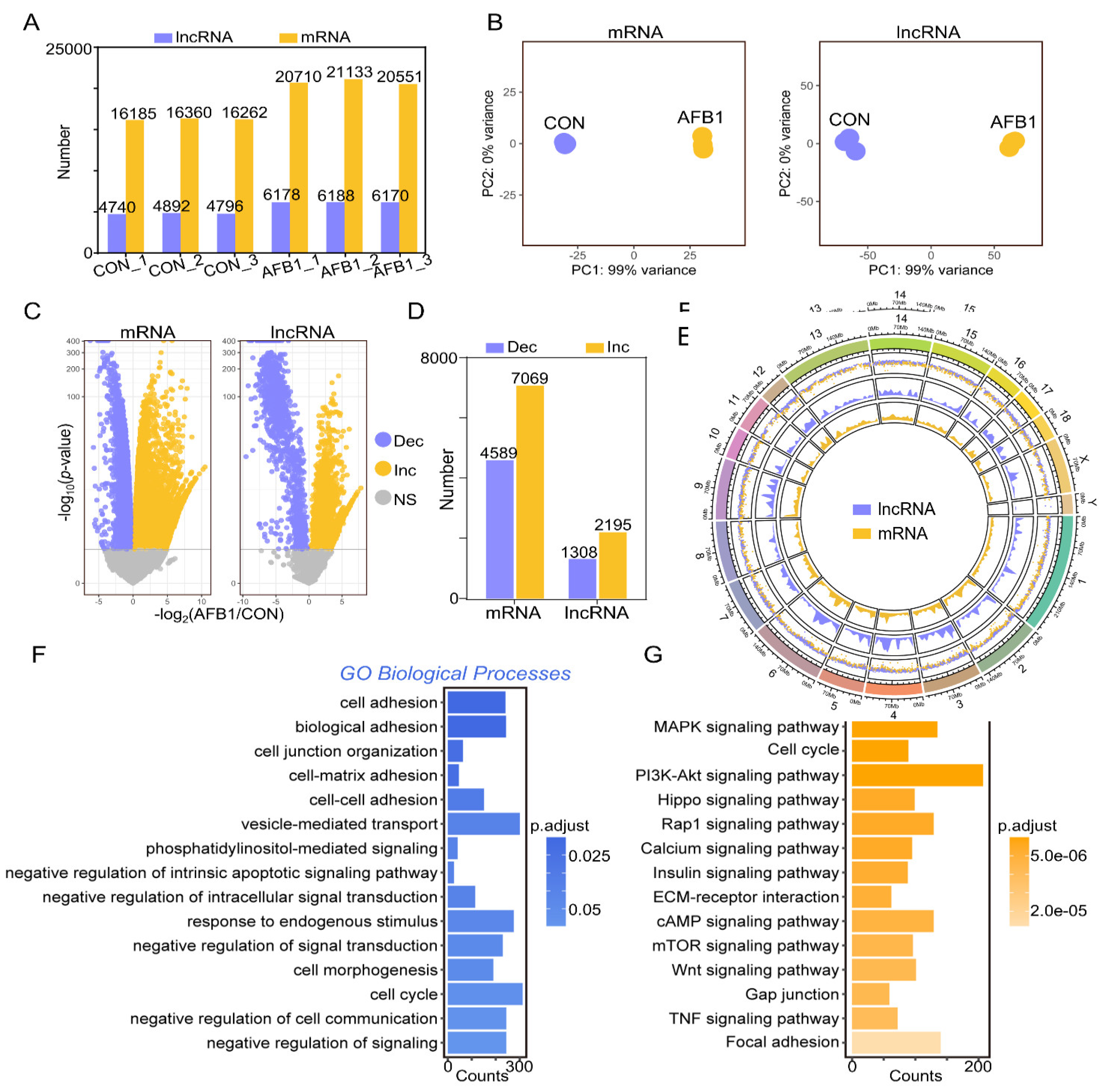
2.2. AFB1 Exposure Affected lncRNA and mRNA Expression of Porcine 3D4/2 Cells
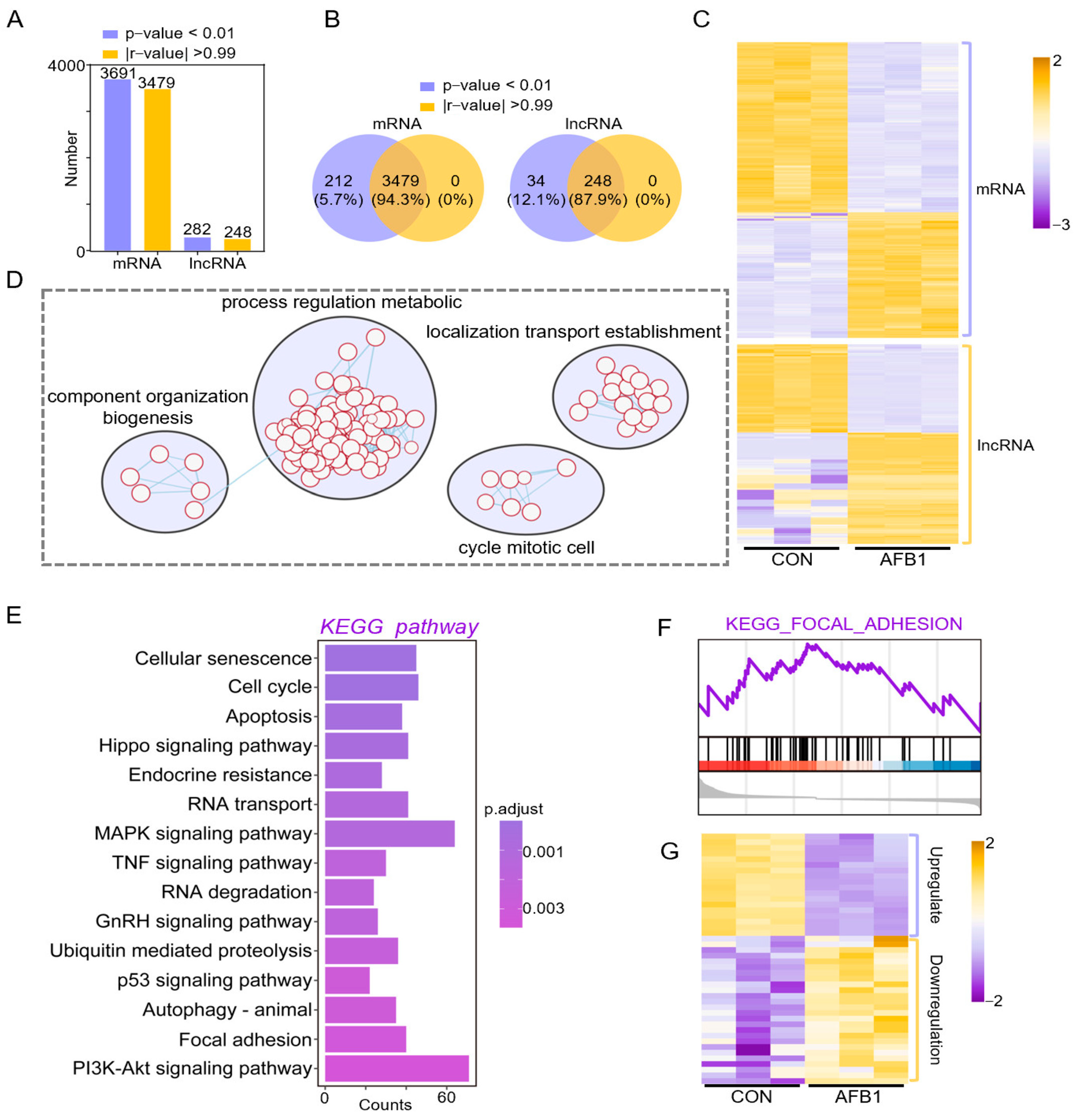
2.3. Co-Expression Analysis of DELs and DEGs in Porcine 3D4/2 Cells
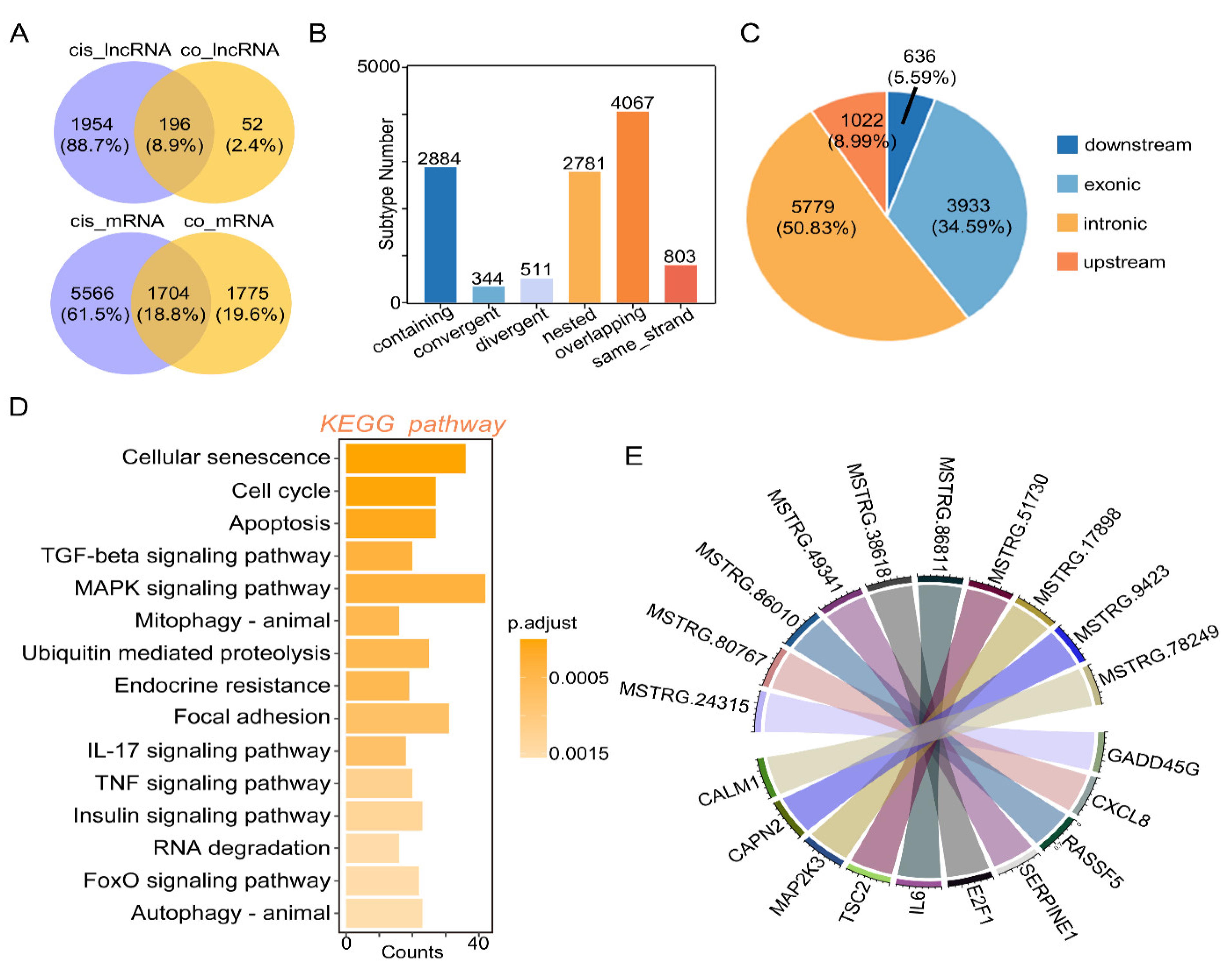
2.4. Cis-Regulation of mRNA and lncRNAs with Target Genes
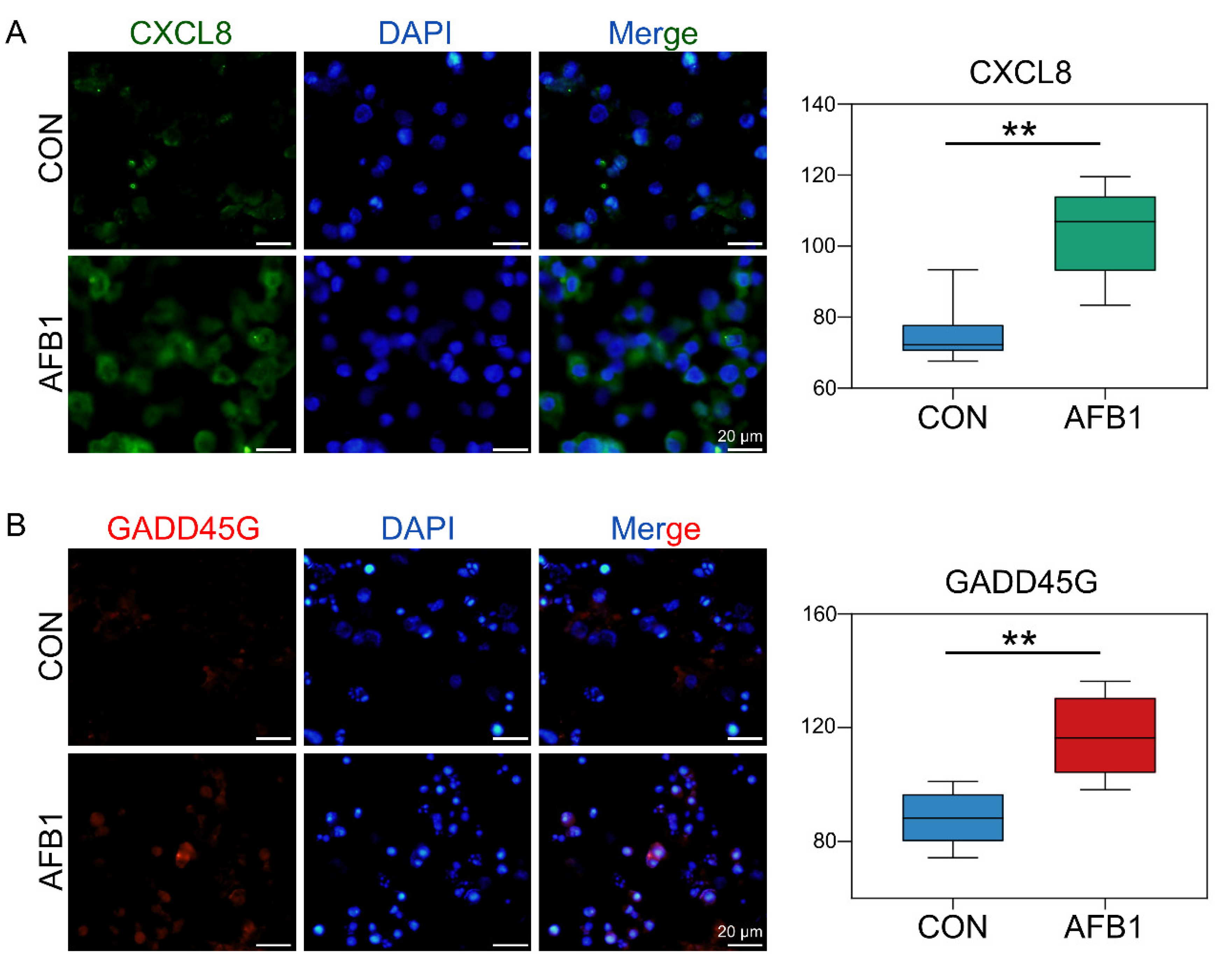
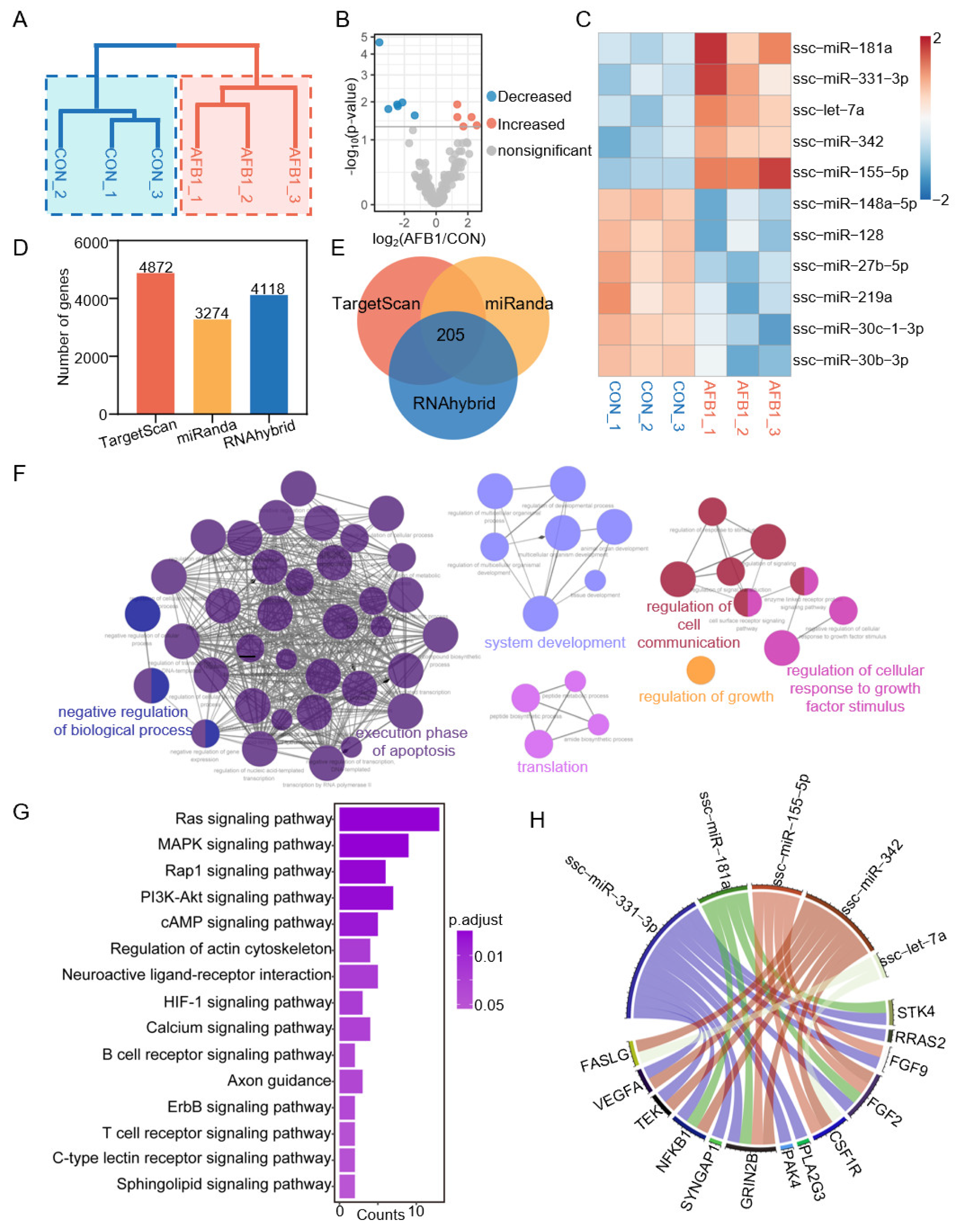
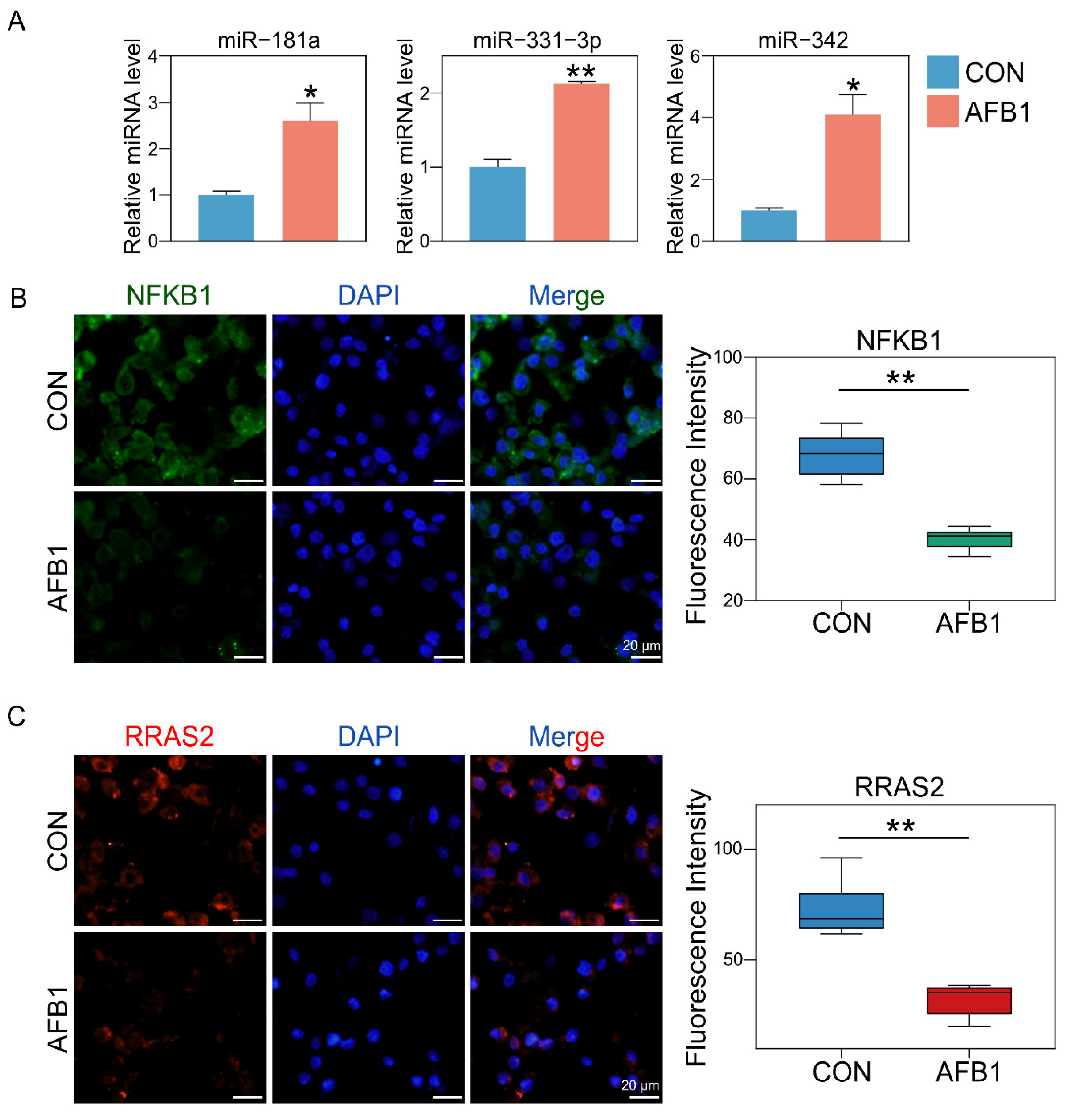
2.5. AFB1 Exposure Altered miRNA Expression
3. Discussion
4. Materials and Methods
4.1. In Vitro AFB1 Treatment of Porcine Alveolar Macrophages
4.2. EdU Staining for Proliferation
4.3. TUNEL Staining
4.4. RNA Extraction and Sequencing of Whole Transcriptome RNA
4.5. Pipeline of Data Processing
4.6. Screening for Candidate lncRNAs
4.7. Discovering Differentially Expressed Genes (DEGs) and RNA Target Prediction
4.8. GO Classification and KEGG Enrichment Analysis
4.9. RT-qPCR
4.10. Immunofluorescence Staining
4.11. Statistical Method
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tkaczyk, A.; Jedziniak, P. Mycotoxin Biomarkers in Pigs-Current State of Knowledge and Analytics. Toxins 2021, 13, 586. [Google Scholar] [CrossRef] [PubMed]
- Liew, W.P.; Mohd-Redzwan, S. Mycotoxin: Its Impact on Gut Health and Microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in Food and Feed. Adv. Food Nutr. Res. 2019, 89, 297–345. [Google Scholar]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [Green Version]
- Benkerroum, N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int. J. Environ. Res. Public Health 2020, 17, 423. [Google Scholar] [CrossRef] [Green Version]
- Pócsi, I.; Király, G.; Bánfalvi, G. Antineoplastic potential of mycotoxins. Acta Microbiol. Immunol. Hung. 2018, 65, 267–307. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Huang, K.; Zhang, B.; Zhu, L.; Xu, W. Aflatoxin B1-induced epigenetic alterations: An overview. Food Chem Toxicol. 2017, 109, 683–689. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Johnson, W.W.; Shimada, T.; Ueng, Y.F.; Yamazaki, H.; Langouët, S. Activation and Detoxication of Aflatoxin B1. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1998, 402, 121–128. [Google Scholar] [CrossRef]
- Gouas, D.; Shi, H.; Hainaut, P. The Aflatoxin-Induced TP53 Mutation at Codon 249 (R249S): Biomarker of Exposure, Early Detection and Target for Therapy. Cancer Lett. 2009, 286, 29–37. [Google Scholar] [CrossRef]
- Supriya, C.H.; Akhila, B.; Pratap, R.K.; Girish, B.P.; Sreenivasula, R.P. Effects of Maternal Exposure to Aflatoxin B1 during Pregnancy on Fertility Output of Dams and Developmental, Behavioral and Reproductive Consequences in Female Offspring Using a Rat Model. Toxicol. Mech. Methods 2016, 26, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Kourousekos, G.D.; Theodosiadou, E.K.; Lymberopoulos, A.G.; Belibasaki, S.; Boscos, C. Effect of Aflatoxin B1 on Blood Serum Oestradiol-17β and Progesterone Concentrations during the Luteal Phase and the Synchronized Oestrus of Goats. Anim. Reprod. 2018, 15, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Cao, Z.; Zhang, J.; Ji, Q.; Li, Y. Aflatoxin B1 Promotes Autophagy Associated with Oxidative Stress-Related PI3K/AKT/mTOR Signaling Pathway in Mice Testis. Environ. Pollut. 2019, 255, 113317. [Google Scholar] [CrossRef]
- Cao, Z.; Shao, B.; Xu, F.; Liu, Y.; Li, Y.; Zhu, Y. Protective Effect of Selenium on Aflatoxin B1-Induced Testicular Toxicity in Mice. Biol. Trac. Elem. Res. 2017, 180, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, W. Aflatoxin B1 Impairs Mitochondrial Functions, Activates Ros Generation, Induces Apoptosis and Involves Nrf2 Signal Pathway in Primary Broiler Hepatocytes. Anim. Sci. J. 2016, 87, 1490–1500. [Google Scholar] [CrossRef]
- Storvik, M.; Huuskonen, P.; Kyllönen, T.; Lehtonen, S.; El-Nezami, H.; Auriola, S.; Pasanen, M. Aflatoxin B1—A Potential Endocrine Disruptor–up-Regulates CYP19A1 in JEG-3 cells. Toxicol. Lett. 2011, 202, 161–167. [Google Scholar] [CrossRef]
- Skinner, M.K.; Ben, M.M.; Sadler-Riggleman, I.; Beck, D.; Nilsson, E.; McBirney, M.; Klukovich, R.; Xie, Y.M.; Tang, C.; Yan, W. Alterations in Sperm DNA Methylation, Non-Coding RNA and Histone Retention Associate with DDT-Induced Epigenetic Transgenerational Inheritance of Disease. Epigenet. Chromatin 2018, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Beck, D.; Ben, M.M.; Skinner, M.K. Integration of Sperm ncRNA-Directed DNA Methylation and DNA Methylation-Directed Histone Retention in Epigenetic Transgenerational Inheritance. Epigenet. Chromatin 2021, 14, 6. [Google Scholar] [CrossRef]
- Mathieu, E.L.; Belhocine, M.; Dao, L.T.; Puthier, D.; Spicuglia, S. Functions of lncRNA in Development and Diseases. Med. Sci. 2014, 30, 790–796. [Google Scholar]
- Ma, Y.; Zhang, J.; Wen, L.; Lin, A. Membrane-Lipid Associated lncRNA: New Regulator in Cancer Signaling. Cancer Lett. 2018, 419, 27–29. [Google Scholar] [CrossRef]
- Liu, X.; Kumar, M.S.; Wang, T.; Xu, Z.; Zhao, X.; Wang, Y.; Yin, H.; Fan, X.; Zeng, B.; Yang, M.; et al. AFB1 Induced Transcriptional Regulation Related to Apoptosis and Lipid Metabolism in Liver of Chicken. Toxins 2020, 12, 290. [Google Scholar] [CrossRef] [PubMed]
- Merrick, B.A.; Chang, J.S.; Phadke, D.P.; Bostrom, M.A.; Shah, R.R.; Wang, X.; Gordon, O.; Wright, G.M. HAfTs Are Novel lncRNA Transcripts from Aflatoxin Exposure. PLoS ONE 2018, 13, e0190992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meki, A.R.M.; Abdel-Ghaffar, S.K.; El-Gibaly, I. Aflatoxin B1 Induces Apoptosis in Rat Liver: Protective Effect of Melatonin. Neuroendocrinol. Lett. 2001, 22, 417–426. [Google Scholar] [PubMed]
- Jackson, P.E.; Groopman, J.D. Aflatoxin and Liver Cancer. Best Pract. Res. Clin. Gastroenterol. 1999, 13, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ogata, M.; Hirai, H.; Kawagishi, H. Detoxification of Aflatoxin B1 by Manganese Peroxidase from the White-Rot Fungus Phanerochaete Sordida YK-624. FEMS Microbiol. Lett. 2011, 314, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Afshar, P.; Roozbeh, N.L.; Shokrzadeh, M.; Ghorbani, H.A.; Naghizadeh, R.S. Bio-Protective Effects of Lactobacillus plantarum subsp. plantarum against Aflatoxin b1 Genotoxicity on Human Blood Lymphocytes: A Native Probiotic Strain Isolated from Iranian Camel Milk. Curr. Med. Mycol. 2020, 6, 54–61. [Google Scholar]
- Ma, J.; Liu, Y.; Guo, Y.; Ma, Q.; Ji, C.; Zhao, L. Transcriptional Profiling of Aflatoxin B1-Induced Oxidative Stress and Inflammatory Response in Macrophages. Toxins 2021, 13, 401. [Google Scholar] [CrossRef]
- Dugyala, R.R.; Sharma, R.P. The Effect of Aflatoxin B1 on Cytokine mRNA and Corresponding Protein Levels in Peritoneal macrophages and Splenic Lymphocytes. Int. J. Immunopharmacol. 1996, 18, 599–608. [Google Scholar] [CrossRef]
- Marin, D.E.; Taranu, I.; Bunaciu, R.P.; Pascale, F.; Tudor, D.S.; Avram, N.; Sarca, M.; Cureu, I.; Criste, R.D.; Suta, V.; et al. Changes in Performance, Blood Parameters, Humoral and Cellular Immune Responses in Weanling Piglets Exposed To Low Doses of Aflatoxin. J. Anim. Sci. 2002, 80, 1250–1257. [Google Scholar] [CrossRef]
- Pu, J.; Yuan, Q.; Yan, H.; Tian, G.; Chen, D.; He, J.; Zeng, P.; Yu, J.; Mao, X.; Huang, Z.; et al. Effects of Chronic Exposure to Low Levels of Dietary Aflatoxin B1 on Growth Performance, Apparent Total Tract Digestibility and Intestinal Health in Pigs. Animals 2021, 11, 336. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Z.Z.; Zhao, M.; Zhang, Y.; Chen, N.H. Role of Non-Coding RNA in the Pathogenesis of Depression. Gene 2020, 735, 144276. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Q.; Pan, X. MicroRNAs and Their Regulatory Roles in Animals and Plants. J. Cell Physiol. 2007, 210, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jin, J.; Wu, J.; Qu, H.; Wu, S.; Bao, W. Transcriptome and Chromatin Accessibility in Porcine Intestinal Epithelial Cells upon Zearalenone Exposure. Sci. Data 2019, 6, 298. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Wang, H.; Li, X.; Ji, H.; Xu, Y. ZEA Exerts Toxic Effects on Reproduction and Development by Mediating Dio3os in Mouse Endometrial Stromal Cells. J. Biochem. Mol. Toxicol. 2019, 33, e22310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.L.; Li, N.; Wang, H.; Ma, J.M.; Shen, W.; Li, L. Zearalenone Exposure Induces the Apoptosis of Porcine Granulosa Cells and Changes Long Noncoding RNA Expression to Promote Antiapoptosis by Activating the JAK2–STAT3 pathway. J. Agric. Food Chem. 2019, 67, 12117–12128. [Google Scholar] [CrossRef]
- Wang, M.; Wu, W.; Li, L.; He, J.; Huang, S.; Chen, S.; Chen, J.; Long, M.; Yang, S.; Li, P. Analysis of the miRNA Expression Profiles in the Zearalenone-Exposed TM3 Leydig Cell Line. Int. J. Mol. Sci. 2019, 20, 635. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.Y.; Yuhan, J.Y.; Huang, K.L.; He, X.Y.; Liang, Z.H.; Xu, W.T. Multidimensional Analysis of the Epigenetic Alterations in Toxicities induced by Mycotoxins. Food Chem. Toxicol. 2021, 153, 112251. [Google Scholar] [CrossRef]
- Long, X.D.; Huang, X.Y.; Ya, O.J.G.; Liao, P.H.; Tang, Y.J.; Ma, Y.; Xia, Q. Polymorphisms in the Precursor microRNAs and Aflatoxin B1-Related Hepatocellular Carcinoma. Mol. Carcinog. 2016, 55, 1060–1072. [Google Scholar] [CrossRef]
- Russo, R.C.; Garcia, C.C.; Teixeira, M.M.; Amaral, F.A. The CXCL8/IL-8 Chemokine Family and Its Receptors in Inflammatory Diseases. Expert Rev. Clin. Immu. 2014, 10, 593–619. [Google Scholar] [CrossRef] [Green Version]
- Krieger, M.; Brunner, T.; Bischoff, S.C.; von Tscharner, V.; Walz, A.; Moser, B.; Baggiolini, M.; Dahinden, C.A. Activation of Human Basophils through the IL-8 Receptor. J. Immunol. 1992, 149, 2662–2667. [Google Scholar]
- Zhang, L.; Yang, Z.; Liu, Y. GADD45 Proteins: Roles in Cellular Senescence and Tumor Development. Exp. Biol. Med. 2014, 239, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Rosemary, S.A.; Richardson, D.R. Growth Arrest and DNA Damage-45 Alpha (GADD45 Alpha). Int. J. Biochem. Cell B 2009, 41, 986–989. [Google Scholar] [CrossRef] [PubMed]
- Pauletto, M.; Tolosi, R.; Giantin, M.; Guerra, G.; Barbarossa, A.; Zaghini, A.; Dacasto, M. Insights into Aflatoxin B1 Toxicity in Cattle: An in vitro Whole-Transcriptomic Approach. Toxins 2020, 12, 429. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Li, Y.; Fan, Y.; Zhao, L.; Wei, H.; Ji, C.; Zhang, J. Molecular Mechanisms of Lipoic Acid Protection against Aflatoxin b1-Induced Liver Oxidative Damage and Inflammatory Responses in Broilers. Toxins 2015, 7, 5435–5447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, Inflammation, and Metabolic Disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; He, J.; Lin, J.; Sun, X.; Sun, C.; Ou, C.; Jiang, C. Distinct Response of the Hepatic Transcriptome to Aflatoxin B1 Induced Hepatocellular Carcinogenesis and Resistance in Rats. Sci. Rep. 2016, 6, 31898. [Google Scholar] [CrossRef]
- Somma, D.; Kok, F.O.; Kerrigan, D.; Wells, C.A.; Carmody, R.J. Defining the Role of Nuclear Factor (NF)-κB p105 Subunit in Human Macrophage by Transcriptomic Analysis of NFKB1 Knockout THP1 Cells. Front Immunol. 2021, 12, 669906. [Google Scholar] [CrossRef]
- Hortal, A.M.; Oeste, C.L.; Cifuentes, C.; Alcoceba, M.; Fernández-Pisonero, I.; Clavaín, L.; Tercero, R.; Mendoza, P.; Domínguez, V.; García-Flores, M.; et al. Overexpression of Wild Type RRAS2, without Oncogenic Mutations, Drives Chronic Lymphocytic Leukemia. Mol. Cancer 2022, 21, 35. [Google Scholar] [CrossRef]
- Li, S.; Liu, R.; Wei, G.; Guo, G.; Yu, H.; Zhang, Y.; Ishfaq, M.; Fazilani, S.A.; Zhang, X. Curcumin Protects against Aflatoxin B1-Induced Liver Injury in Broilers via the Modulation of Long Non-Coding RNA Expression. Ecotoxicol. Environ. Saf. 2021, 208, 111725. [Google Scholar] [CrossRef]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A Fast and Accurate Coding Potential Calculator Based on Sequence Intrinsic Features. Nucleic Acids Res. 2017, 45, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing Sequence Intrinsic Composition to Classify Protein-Coding and Long Non-Coding Transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam Protein Families Database: Towards a More Sustainable Future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Wucher, V.; Legeai, F.; Hedan, B.; Rizk, G.; Lagoutte, L.; Leeb, T.; Jagannathan, V.; Cadieu, E.; David, A.; Lohi, H.; et al. FEELnc: A Tool for Long Non-Coding RNA Annotation and Its Application to the Dog Transcriptome. Nucleic Acids Res. 2017, 45, e57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting Effective microRNA Target Sites in Mammalian mRNAs. elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.; Rehmsmeier, M. RNAhybrid: microRNA Target Prediction Easy, Fast and Flexible. Nucleic Acids Res. 2006, 34, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, M.Y.; Li, N.; Wang, J.J.; Ge, W.; Tan, S.J.; Shen, W.; Li, L. Zearalenone Exposure Triggered Porcine Granulosa Cells Apoptosis via microRNAs-Mediated Focal Adhesion Pathway. Toxicol. Lett. 2020, 330, 80–89. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chao, H.; Ma, H.; Sun, J.; Yuan, S.; Dong, P.; Zhao, A.; Li, L.; Shen, W.; Zhang, X. Whole-Transcriptome Analysis of Non-Coding RNA Alteration in Porcine Alveolar Macrophage Exposed to Aflatoxin B1. Toxins 2022, 14, 373. https://doi.org/10.3390/toxins14060373
Chao H, Ma H, Sun J, Yuan S, Dong P, Zhao A, Li L, Shen W, Zhang X. Whole-Transcriptome Analysis of Non-Coding RNA Alteration in Porcine Alveolar Macrophage Exposed to Aflatoxin B1. Toxins. 2022; 14(6):373. https://doi.org/10.3390/toxins14060373
Chicago/Turabian StyleChao, Huhe, Haohai Ma, Jiadong Sun, Shuai Yuan, Peiyu Dong, Aihong Zhao, Lan Li, Wei Shen, and Xifeng Zhang. 2022. "Whole-Transcriptome Analysis of Non-Coding RNA Alteration in Porcine Alveolar Macrophage Exposed to Aflatoxin B1" Toxins 14, no. 6: 373. https://doi.org/10.3390/toxins14060373
APA StyleChao, H., Ma, H., Sun, J., Yuan, S., Dong, P., Zhao, A., Li, L., Shen, W., & Zhang, X. (2022). Whole-Transcriptome Analysis of Non-Coding RNA Alteration in Porcine Alveolar Macrophage Exposed to Aflatoxin B1. Toxins, 14(6), 373. https://doi.org/10.3390/toxins14060373