Patulin Detoxification by Recombinant Manganese Peroxidase from Moniliophthora roreri Expressed by Pichia pastoris

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
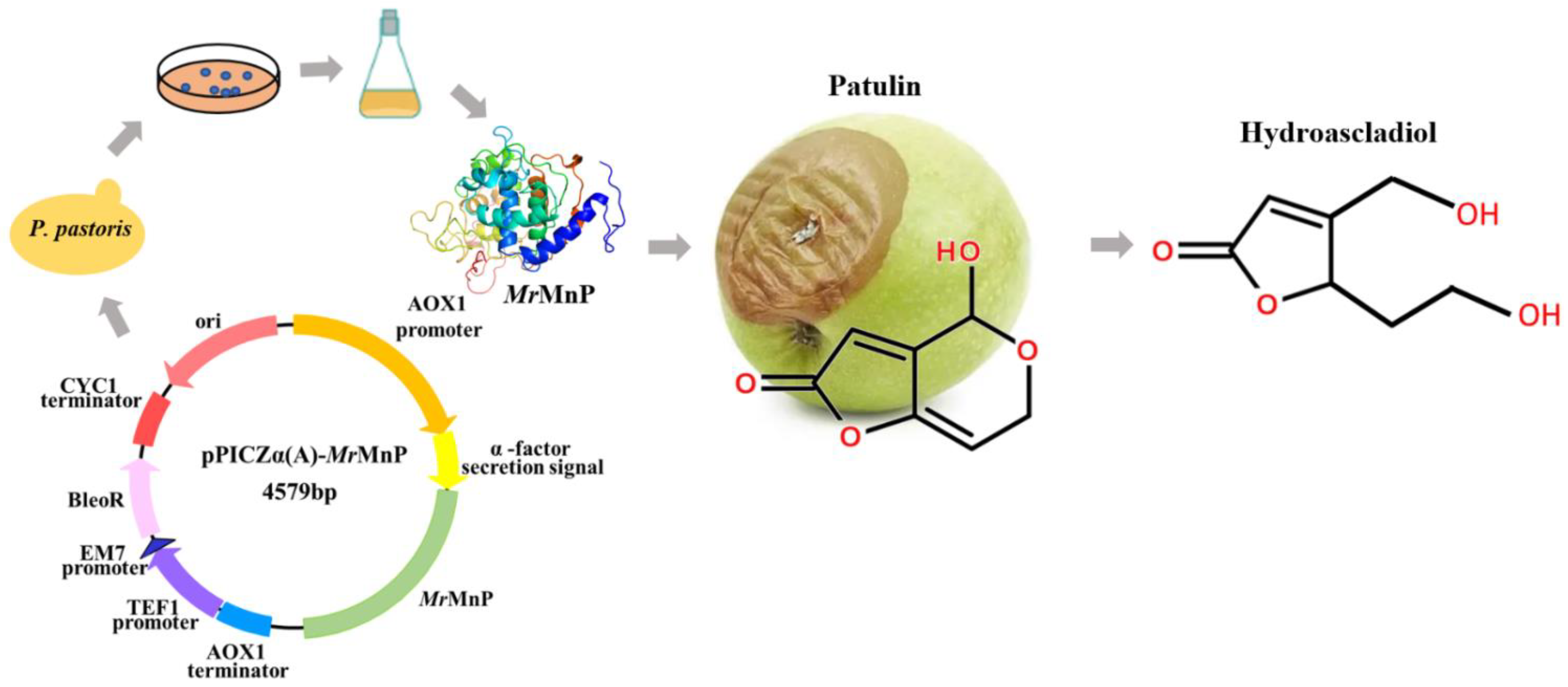
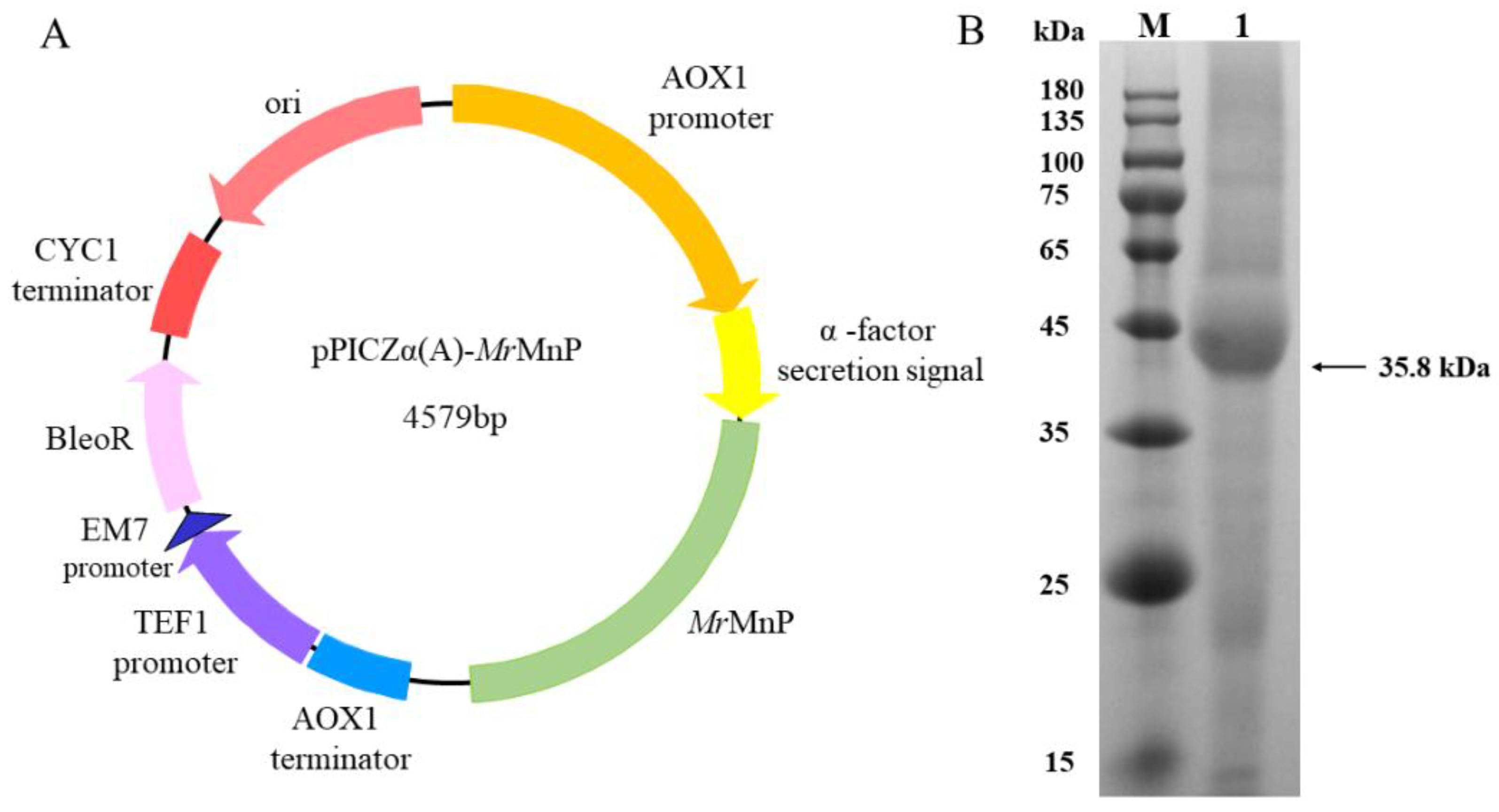
2.1. Recombinant Production of the Manganese Peroxidase MrMnP
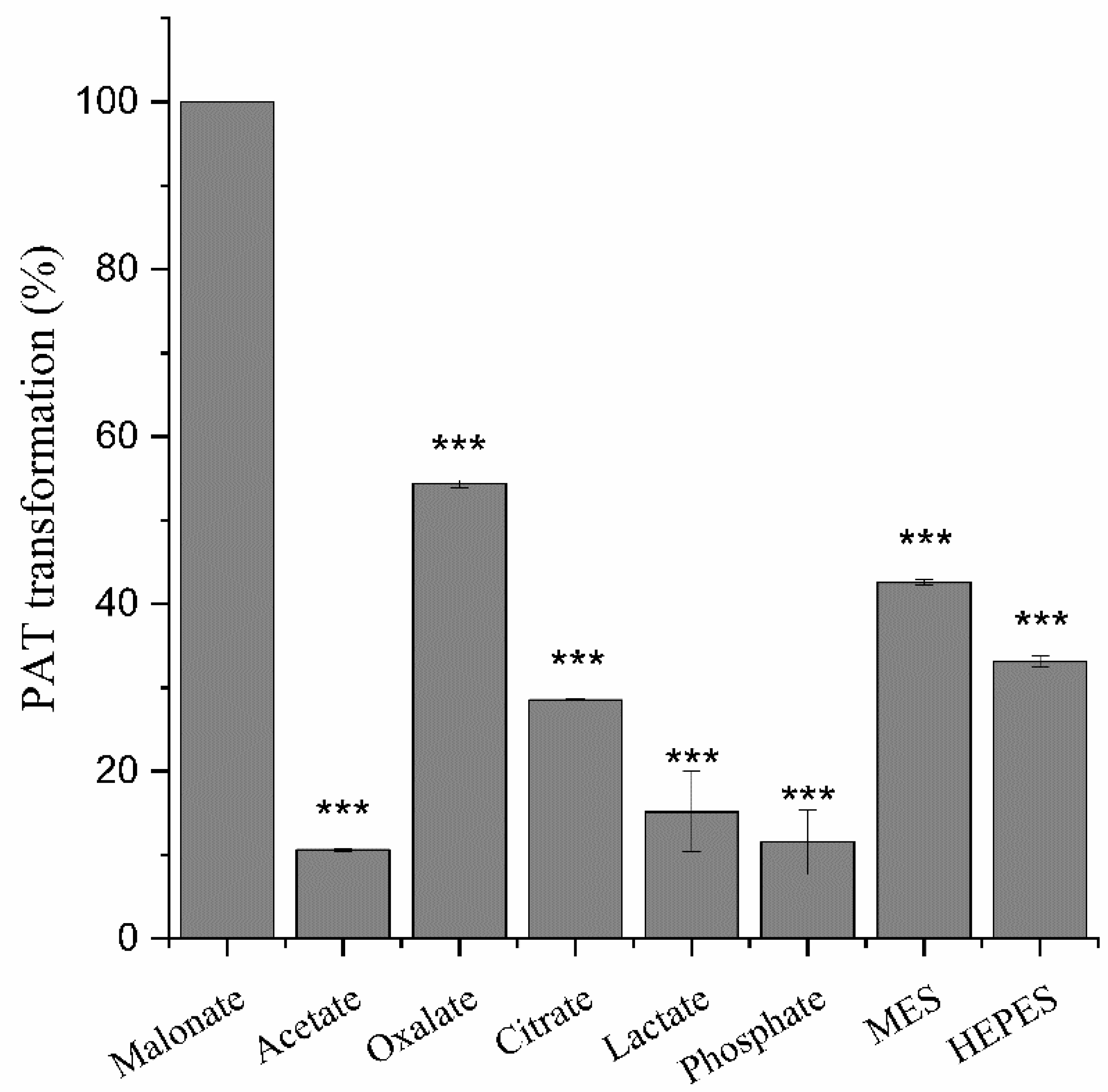
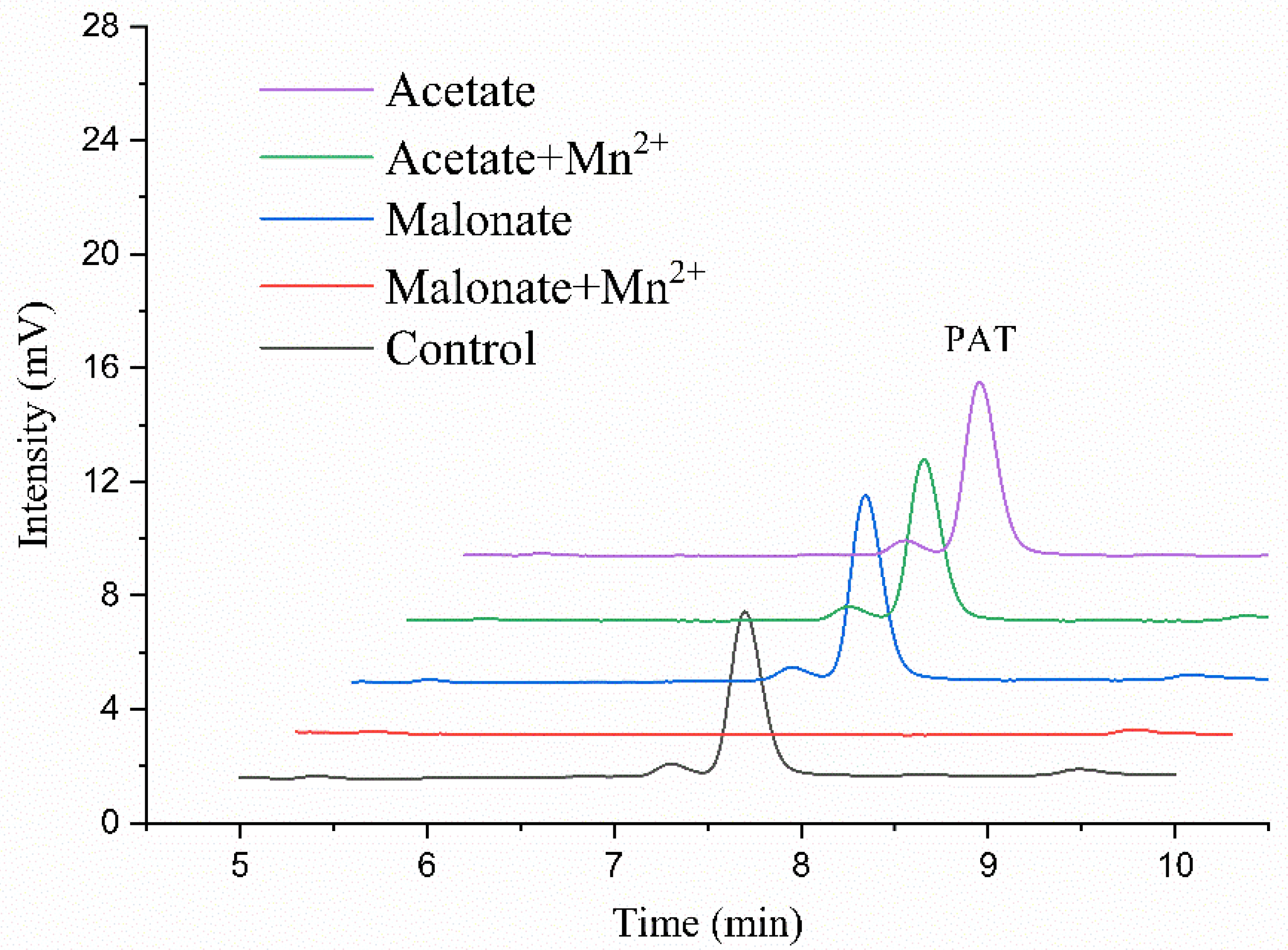
2.2. The Di-Carboxylic Acids Play a Critical Role in Degradation of Patulin by MrMnP
2.3. Mn2+ Is Another Key Determinant in Degradation of Patulin by MrMnP
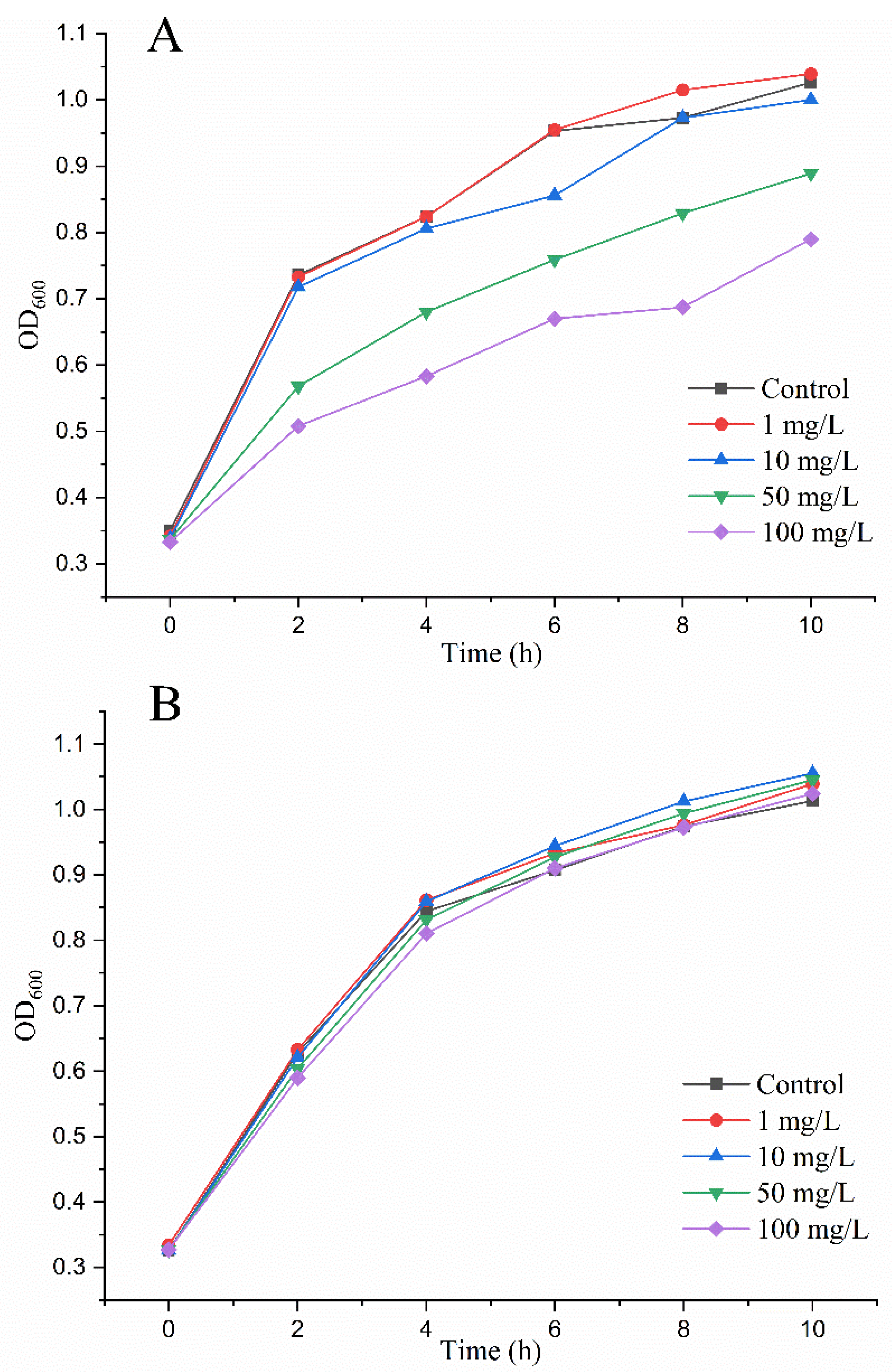
2.4. MrMnP-Catalyzed Degradation of Patulin Led to Detoxification
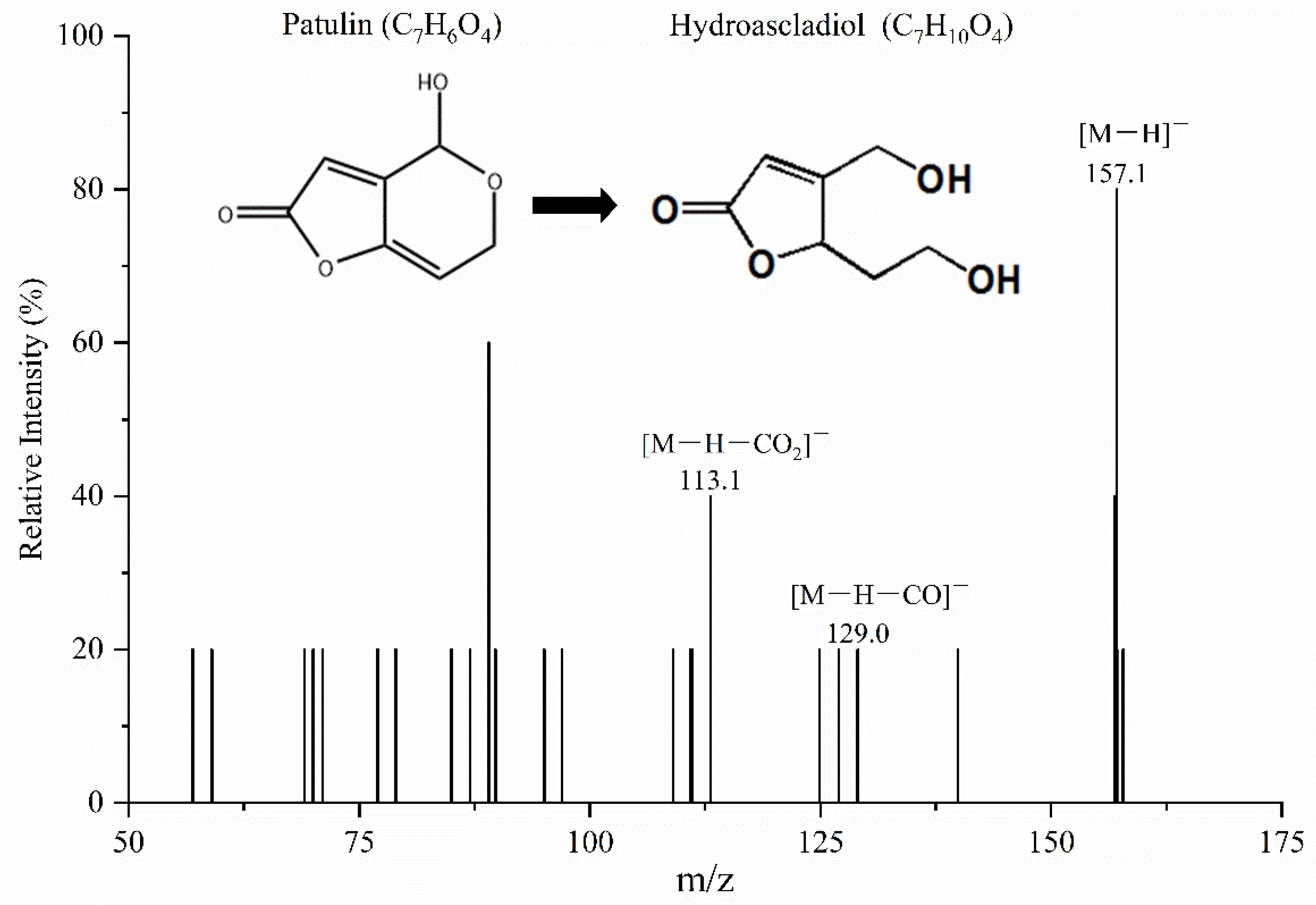
2.5. Structural Analysis of the Degradation Products
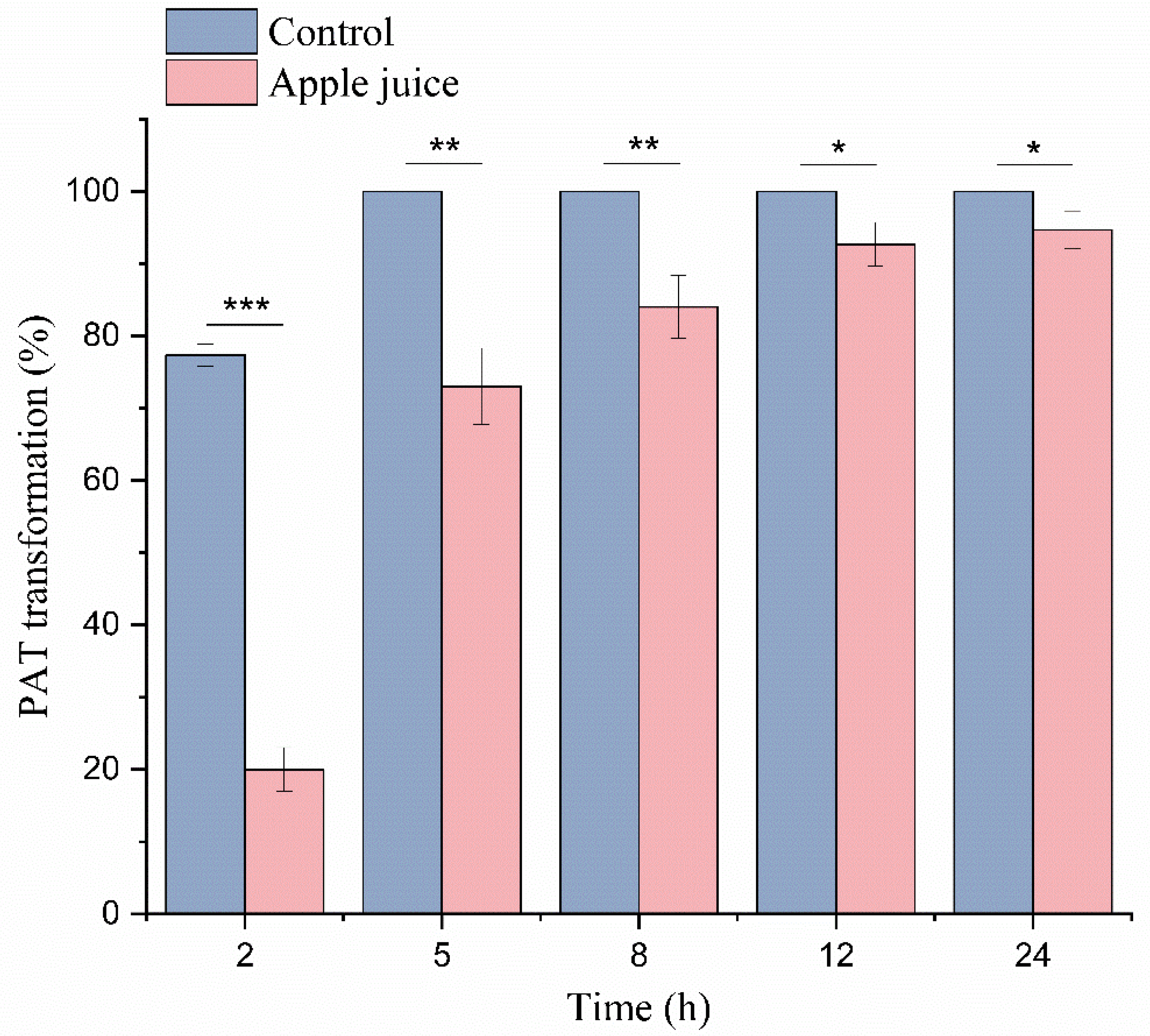
2.6. Degradation in a Simulated Patulin-Contaminated Apple Juice
3. Conclusions
4. Materials and Methods
4.1. Strains and Plasmids
4.2. Cloning and Expression of MrMnP
4.3. Fed-Batch Fermentation of MrMnP in a Bioreactor
4.4. Degradation of Patulin by MrMnP
4.5. Effect of the Buffer Componesnts on Degradation of Patulin by MrMnP
4.6. Effect of Mn2+ on Degradation of Patulin
4.7. Toxicity Assay
4.8. HPLC and LC-MS/MS Analyses
4.9. MrMnP-Catalyzed Degradation of Patulin in a Simulated Patulin-Contaminated Apple Juice
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PAT | Patulin |
| MnP | Manganese Peroxidases |
| ABTS | 2,2′-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| H2O2 | Hydrogen Peroxide |
| MES | 2-Morpholinoethanesulphonic acid |
| HEPES | 2-[4-(2-Hydroxyethyl)-1-piperazinyl] Ethanesulfonic acid |
References
- Iqbal, S.Z.; Malik, S.; Asi, M.R.; Selamat, J.; Malik, N. Natural occurrence of patulin in different fruits, juices and smoothies and evaluation of dietary intake in Punjab, Pakistan. Food Control 2018, 84, 370–374. [Google Scholar] [CrossRef]
- Li, N.; Cui, R.; Zhang, F.; Meng, X.; Liu, B. A novel enzyme from Rhodotorula mucilaginosa Aldolase: Isolation, identification and degradation for patulin in apple juice. Process Biochem. 2022, 116, 148–156. [Google Scholar] [CrossRef]
- Zhong, L.; Carere, J.; Lu, Z.; Lu, F.; Zhou, T. Patulin in apples and apple-based food products: The burdens and the mitigation strategies. Toxins 2018, 10, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Ma, J.; Solairaj, D.; Fu, Y.; Zhang, H. Efficacy of Meyerozyma guilliermondii in controlling patulin production by Penicillium expansum in shuijing pears. Biol. Control 2022, 168, 104856. [Google Scholar] [CrossRef]
- Pal, S.; Singh, N.; Ansari, K.M. Toxicological effects of patulin mycotoxin on the mammalian system: An overview. Toxicol. Res. 2017, 6, 764–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Egmond, H.P. Worldwide regulations for mycotoxins. Adv. Exp. Med. Biol. 2002, 504, 257–269. [Google Scholar]
- Torovic, L.; Dimitrov, N.; Lopes, A.; Martins, C.; Alvito, P.; Assuncao, R. Patulin in fruit juices: Occurrence, bioaccessibility, and risk assessment for Serbian population. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018, 35, 985–995. [Google Scholar] [CrossRef]
- Cano-Sancho, G.; Marin, S.; Ramos, A.J.; Sanchis, V. Survey of patulin occurrence in apple juice and apple products in Catalonia, Spain, and an estimate of dietary intake. Food Addit. Contam. Part B Surveill. 2009, 2, 59–65. [Google Scholar] [CrossRef]
- Barreira, M.J.; Alvito, P.C.; Almeida, C.M.M. Occurrence of patulin in apple-based-foods in Portugal. Food Chem. 2010, 121, 653–658. [Google Scholar] [CrossRef]
- Ioi, J.D.; Zhou, T.; Tsao, R.; Marcone, F.M. Mitigation of patulin in fresh and processed foods and beverages. Toxins 2017, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Karaca, H.; Sedat Velioglu, Y. Effects of some metals and chelating agents on patulin degradation by ozone. Ozone Sci. Eng. 2009, 31, 224–231. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, H.; Yang, Q.; Ren, R. Efficacy of Pichia caribbica in controlling blue mold rot and patulin degradation in apples. Int. J. Food Microbiol. 2013, 162, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Jiang, W.; Li, C.; Ma, N.; Xu, Y.; Meng, X. Patulin biodegradation by marine yeast Kodameae ohmeri. Food Addit Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 352–360. [Google Scholar] [PubMed]
- Castoria, R.; Mannina, L.; Duran-Patron, R.; Maffei, F.; Sobolev, A.P.; De Felice, D.V.; Pinedo-Rivilla, C.; Ritieni, A.; Ferracane, R.; Wright, S.A. Conversion of the mycotoxin patulin to the less toxic desoxypatulinic acid by the biocontrol yeast Rhodosporidium kratochvilovae strain LS11. J. Agric. Food Chem. 2011, 59, 11571–11578. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Tang, H.; Yang, C.; Meng, X.; Liu, B. Detoxification of mycotoxin patulin by the yeast Rhodotorula mucilaginosa. Food Control 2019, 96, 47–52. [Google Scholar] [CrossRef]
- Ito, M.; Sato, I.; Ishizaka, M.; Yoshida, S.; Koitabashi, M.; Yoshida, S.; Tsushima, S. Bacterial cytochrome P450 system catabolizing the Fusarium toxin deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Carere, J.; Hassan, Y.I.; Lepp, D.; Zhou, T. The Identification of DepB: An Enzyme Responsible for the Final Detoxification Step in the Deoxynivalenol Epimerization Pathway in Devosia mutans 17-2-E-8. Front. Microbiol. 2018, 9, 1573. [Google Scholar] [CrossRef]
- Roy, B.P.; Paice, M.G.; Archibald, F.S.; Misra, S.K.; Misiak, L.E. Creation of metal-complexing agents, reduction of manganese dioxide, and promotion of manganese peroxidase-mediated Mn(III) production by cellobiose:quinone oxidoreductase from Trametes versicolor. J. Biol. Chem. 1994, 269, 19745–19750. [Google Scholar] [CrossRef]
- Kondo, R.; Harazono, K.; Sakai, K. Bleaching of hardwood kraft pulp with manganese peroxidase secreted from Phanerochaete sordida YK-624. Appl. Environ. Microbiol. 1994, 60, 4359–4363. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Qin, X.; Hao, Z.; Luo, H.; Yao, B.; Su, X. Degradation of four major mycotoxins by eight manganese peroxidases in presence of a dicarboxylic acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef] [Green Version]
- Bronikowski, A.; Koschorreck, K.; Urlacher, V.B. Redesign of a new manganese peroxidase highly expressed in Pichia pastoris towards a lignin-degrading versatile peroxidase. ChemBioChem 2018, 19, 2481–2489. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Su, X.; Luo, H.; Ma, R.; Yao, B.; Ma, F. Deciphering lignocellulose deconstruction by the white rot fungus Irpex lacteus based on genomic and transcriptomic analyses. Biotechnol. Biofuels 2018, 11, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Ren, K.; Jia, R.; Chen, W.; Sun, R. Expression of a fungal manganese peroxidase in Escherichia coli: A comparison between the soluble and refolded enzymes. BMC Biotechnol. 2016, 16, 87. [Google Scholar] [CrossRef] [Green Version]
- Whitwam, R.; Tien, M. Heterologous expression and reconstitution of fungal Mn peroxidase. Arch. Biochem. Biophys. 1996, 333, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Kongsaeree, P.; Charron, R.; Lajoie, C.; Xu, H.; Scott, G.; Kelly, C. Production and separation of manganese peroxidase from heme amended yeast cultures. Biotechnol. Bioeng. 2008, 99, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Kongsaeree, P.; Schilke, K.; Lajoie, C.; Kelly, C. Effects of pH and temperature on recombinant manganese peroxidase production and stability. Appl. Biochem. Biotechnol. 2008, 146, 15–27. [Google Scholar] [CrossRef]
- Hilden, K.; Makela, M.R.; Lundell, T.; Kuuskeri, J.; Chernykh, A.; Golovleva, L.; Archer, D.B.; Hatakka, A. Heterologous expression and structural characterization of two low pH laccases from a biopulping white-rot fungus Physisporinus rivulosus. Appl. Microbiol. Biotechnol. 2013, 97, 1589–1599. [Google Scholar] [CrossRef]
- Xu, H.; Guo, M.Y.; Gao, Y.H.; Bai, X.H.; Zhou, X.W. Expression and characteristics of manganese peroxidase from Ganoderma lucidum in Pichia pastoris and its application in the degradation of four dyes and phenol. BMC Biotechnol. 2017, 17, 19. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Punt, P.J.; van den Hondel, C.A. Fungal peroxidases: Molecular aspects and applications. J. Biotechnol. 2002, 93, 143–158. [Google Scholar] [CrossRef]
- Cortes-Espinosa, D.V.; Absalon, A.E.; Sanchez, N.; Loera, O.; Rodriguez-Vazquez, R.; Fernandez, F.J. Heterologous expression of manganese peroxidase in Aspergillus niger and its effect on phenanthrene removal from soil. J. Mol. Microbiol. Biotechnol. 2011, 21, 120–129. [Google Scholar] [CrossRef]
- Ansari, Z.; Karimi, A.; Sedghi, S.; Razzaghi, M.; Ebrahimi, S. Glucose oxidase effect on treatment of textile effluent containing reactive azo dyes by Phanerochaete chrysosporium. J. Chem. Technol. Biotechnol. 2017, 92, 1721–1726. [Google Scholar] [CrossRef]
- Zhang, J.; Presley, G.N.; Hammel, K.E.; Ryu, J.S.; Menke, J.R.; Figueroa, M.; Hu, D.; Orr, G.; Schilling, J.S. Localizing gene regulation reveals a staggered wood decay mechanism for the brown rot fungus Postia placenta. Proc. Natl. Acad. Sci. USA 2016, 113, 10968–10973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumiero, A.; Murphy, E.J.; Metcalfe, C.L.; Moody, P.C.; Raven, E.L. An analysis of substrate binding interactions in the heme peroxidase enzymes: A structural perspective. Arch. Biochem. Biophys. 2010, 500, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Feussner, K.; Wu, T.; Yan, F.; Karlovsky, P.; Zheng, X. Detoxification of mycotoxin patulin by the yeast Rhodosporidium paludigenum. Food Chem. 2015, 179, 1–5. [Google Scholar] [CrossRef]
- Hathout, A.S.; Aly, S.E. Biological detoxification of mycotoxins: A review. Ann. Microbiol. 2014, 64, 905–919. [Google Scholar] [CrossRef]
- Malysheva, S.V.; Diana Di Mavungu, J.; Boonen, J.; De Spiegeleer, B.; Goryacheva, I.Y.; Vanhaecke, L.; De Saeger, S. Improved positive electrospray ionization of patulin by adduct formation: Usefulness in liquid chromatography-tandem mass spectrometry multi-mycotoxin analysis. J. Chromatogr. A 2012, 1270, 334–339. [Google Scholar] [CrossRef] [Green Version]
- Hawar, S.; Vevers, W.; Karieb, S.; Ali, B.K.; Billington, R.; Beal, J. Biotransformation of patulin to hydroascladiol by Lactobacillus plantarum. Food Control 2013, 34, 502–508. [Google Scholar] [CrossRef]
- Diao, E.; Hou, H.; Hu, W.; Dong, H.; Li, X. Removing and detoxifying methods of patulin: A review. Trends Food Sci. Technol. 2018, 81, 139–145. [Google Scholar] [CrossRef]
- Crestini, C.; D’Annibale, A.; Sermanni, G.G.; Saladino, R. The reactivity of phenolic and non-phenolic residual kraft lignin model compounds with Mn(II)-peroxidase from Lentinula edodes. Bioorg. Med. Chem. 2000, 8, 433–438. [Google Scholar] [CrossRef]
- Kapich, A.N.; Korneichik, T.V.; Hatakka, A.; Hammel, K.E. Oxidizability of unsaturated fatty acids and of a non-phenolic lignin structure in the manganese peroxidase-dependent lipid peroxidation system. Enzym. Microb. Technol. 2010, 46, 136–140. [Google Scholar] [CrossRef]
- Kong, W.; Chen, H.; Lyu, S.; Ma, F.; Yu, H.; Zhang, X. Characterization of a novel manganese peroxidase from white-rot fungus Echinodontium taxodii 2538, and its use for the degradation of lignin-related compounds. Process Biochem. 2016, 51, 1776–1783. [Google Scholar] [CrossRef]
- Wang, X.; Yao, B.; Su, X. Linking enzymatic oxidative degradation of lignin to organics detoxification. Int. J. Mol. Sci. 2018, 19, 3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostić, A.; Stojanović, M.; Lopičić, Z.; Milojković, J.; Petrović, M. Biomass waste material as potential adsorbent for sequestering pollutants. Zaštita Materijala 2012, 53, 231–237. [Google Scholar]
- Qin, X.; Zhang, J.; Zhang, X.; Yang, Y. Induction, purification and characterization of a novel manganese peroxidase from Irpex lacteus CD2 and its application in the decolorization of different types of dye. PLoS ONE 2014, 9, e113282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Wu, H.; Liao, X.; Ding, Y.; Sun, J.; Zhang, D. Purification and characterization of novel manganese peroxidase from Rhizoctonia sp. SYBC-M3. Biotechnol. Bioprocess Eng. 2011, 15, 1016–1021. [Google Scholar] [CrossRef]
- Praveen, K.; Usha, K.Y.; Viswanath, B.; Reddy, B.R. Kinetic properties of manganese peroxidase from the mushroom Stereum ostrea and its ability to decolorize dyes. J. Microbiol. Biotechnol. 2012, 22, 1540–1548. [Google Scholar] [CrossRef] [Green Version]
- Ürek, R.Ö.; Pazarlioğlu, N.K. Purification and partial characterization of manganese peroxidase from immobilized Phanerochaete chrysosporium. Process Biochem. 2004, 39, 2061–2068. [Google Scholar] [CrossRef]
- Qin, X.; Sun, X.; Huang, H.; Bai, Y.; Wang, Y.; Luo, H.; Yao, B.; Zhang, X.; Su, X. Oxidation of a non-phenolic lignin model compound by two Irpex lacteus manganese peroxidases: Evidence for implication of carboxylate and radicals. Biotechnol. Biofuels 2017, 10, 103. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wang, X.; Penttinen, L.; Luo, H.; Zhang, Y.; Liu, B.; Yao, B.; Hakulinen, N.; Zhang, W.; Su, X. Patulin Detoxification by Recombinant Manganese Peroxidase from Moniliophthora roreri Expressed by Pichia pastoris. Toxins 2022, 14, 440. https://doi.org/10.3390/toxins14070440
Wang S, Wang X, Penttinen L, Luo H, Zhang Y, Liu B, Yao B, Hakulinen N, Zhang W, Su X. Patulin Detoxification by Recombinant Manganese Peroxidase from Moniliophthora roreri Expressed by Pichia pastoris. Toxins. 2022; 14(7):440. https://doi.org/10.3390/toxins14070440
Chicago/Turabian StyleWang, Shuai, Xiaolu Wang, Leena Penttinen, Huiying Luo, Yuhong Zhang, Bo Liu, Bin Yao, Nina Hakulinen, Wei Zhang, and Xiaoyun Su. 2022. "Patulin Detoxification by Recombinant Manganese Peroxidase from Moniliophthora roreri Expressed by Pichia pastoris" Toxins 14, no. 7: 440. https://doi.org/10.3390/toxins14070440
APA StyleWang, S., Wang, X., Penttinen, L., Luo, H., Zhang, Y., Liu, B., Yao, B., Hakulinen, N., Zhang, W., & Su, X. (2022). Patulin Detoxification by Recombinant Manganese Peroxidase from Moniliophthora roreri Expressed by Pichia pastoris. Toxins, 14(7), 440. https://doi.org/10.3390/toxins14070440