Contamination, Detection and Control of Mycotoxins in Fruits and Vegetables


Abstract
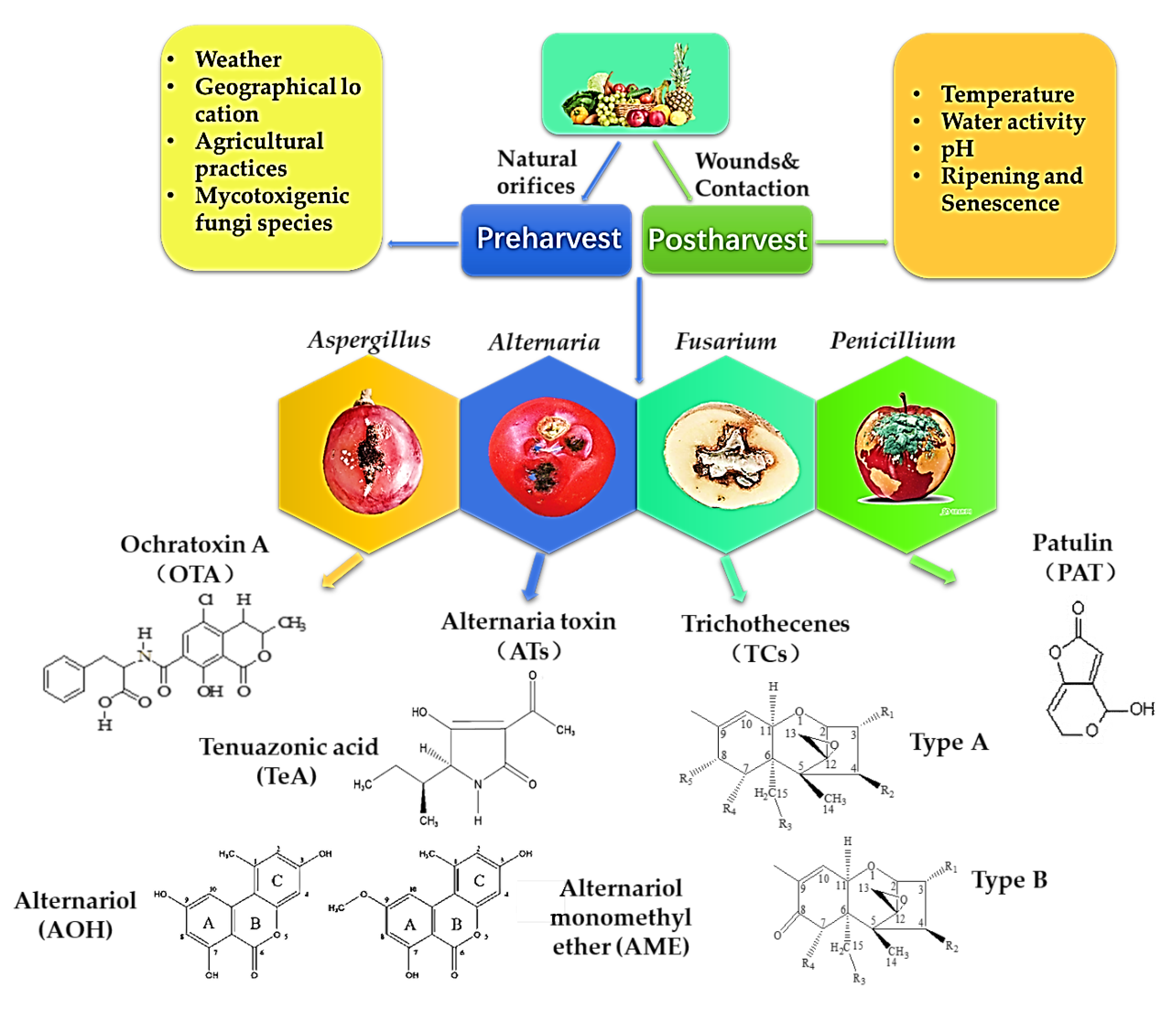
:1. Introduction
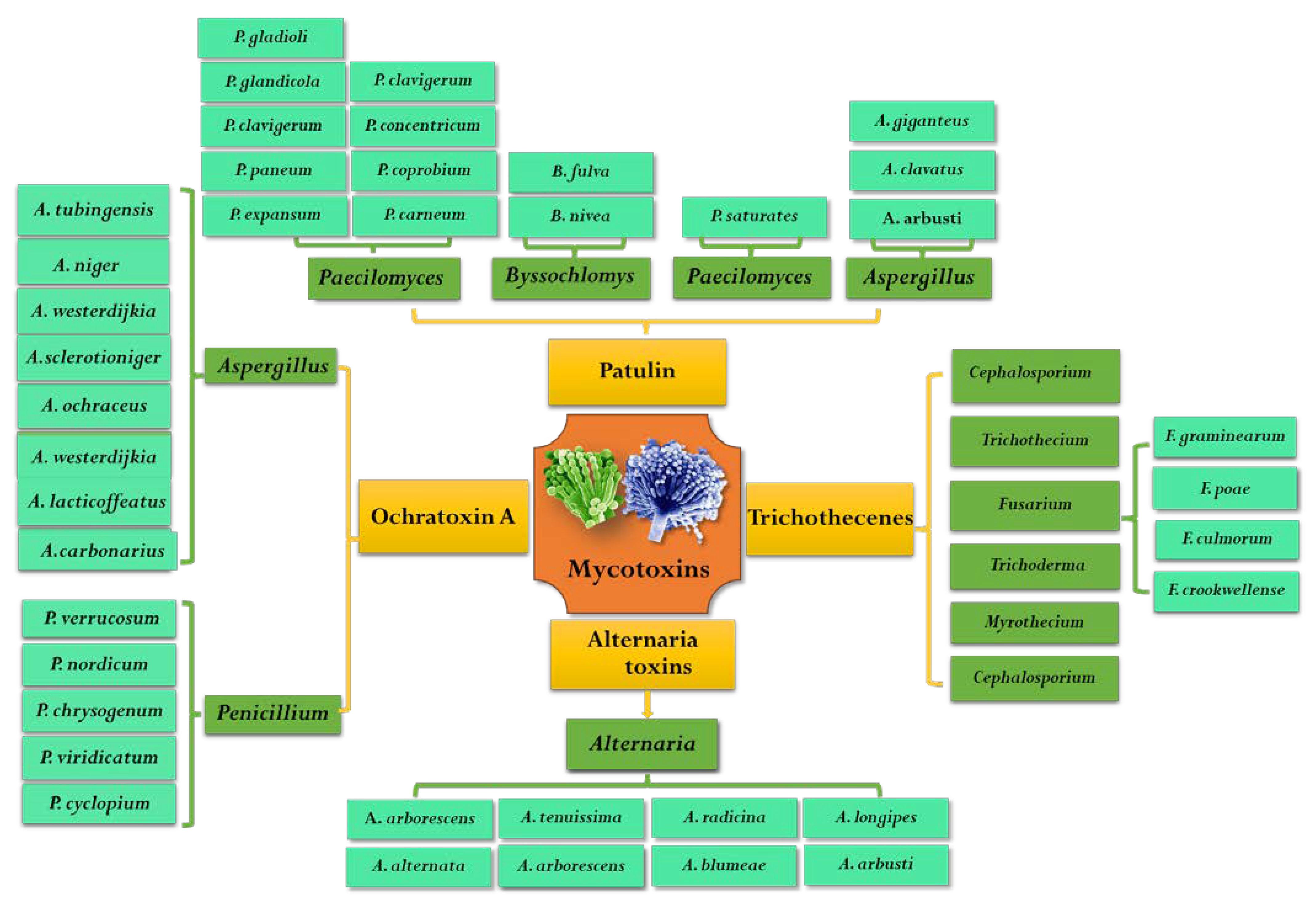
2. Occurrence, Contamination and Toxicity of Mycotoxins
2.1. Ochratoxin A (OTA)
2.2. Patulin (PAT)
2.3. Alternaria Toxins (ATs)
2.4. Trichothecenes (TCs)
3. Determination Technique of Mycotoxins in Fruits and Vegetables
3.1. Chromatographical Technique
3.2. Biosensor Technique
3.2.1. Antibody and Aptamer
3.2.2. SPR, QCM and Colorimetric Sensor
3.2.3. Electrochemical Sensor
3.3. Immunological Technique
4. Degradation Technique of Mycotoxins in Fruits and Vegetables
4.1. Physical Method
4.1.1. Physical Adsorption
4.1.2. Physical Removal and Detoxification
- (1)
- Heating
- (2)
- Microwave
- (3)
- Irradiation
4.2. Chemical Degradation
4.3. Biological Degradation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sanzani, S.M.; Reverberi, M.; Geisen, R. Mycotoxins in harvested fruits and vegetables: Insights in producing fungi, biological role, conducive conditions, and tools to manage postharvest contamination. Postharvest Biol. Technol. 2016, 122, 95–105. [Google Scholar] [CrossRef]
- Yang, J.; Li, J.; Jiang, Y.; Duan, X.; Qu, H.; Yang, B.; Chen, F.; Dharini, S. Natural occurrence, analysis, and prevention of my-cotoxins in fruits and their processed products. Crit. Rev. Food Sci. 2014, 54, 64–83. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Cruz, M.L.; Mansilla, M.L.; Tadeo, J.L. Mycotoxins in fruits and their processed products: Analysis, occurrence and health implications. J. Adv. Res. 2010, 1, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Barkai-golan, R.; Paster, N. Mycotoxins in Fruits and Vegetables; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Ünüsan, N. Systematic review of mycotoxins in food and feeds in Turkey. Food Control. 2018, 97, 1–14. [Google Scholar] [CrossRef]
- Omotayo, O.P.; Omotayo, A.O.; Mwanza, M.; Babalola, O.O. Prevalence of Mycotoxins and Their Consequences on Human Health. Toxicol. Res. 2019, 35, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Da Rocha, M.E.B.; da Chagas Oliveira Freire, F.; Feitosa Maia, F.E.; Florindo Guedes, M.I.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Zhao, Z.; Yang, X.; Zhao, X.; Chen, L.; Bai, B.; Zhou, C.; Wang, J. Method Development and Validation for the Analysis of Emerging and Traditional Fusarium Mycotoxins in Pepper, Potato, Tomato, and Cucumber by UPLC-MS/MS. Food Anal. Methods 2018, 11, 1780–1788. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Nie, J.; Bacha, S.A.S.; Yan, Z.; Gao, G. Occurrence and co-occurrence of mycotoxins in apple and apple products from China. Food Control 2020, 118, 107354. [Google Scholar] [CrossRef]
- Pan, T.-T.; Sun, D.-W.; Pu, H.; Wei, Q.-Y. Simple Approach for the Rapid Detection of Alternariol in Pear Fruit by Surface-Enhanced Raman Scattering with Pyridine-Modified Silver Nanoparticles. J. Agric. Food Chem. 2018, 66, 2180–2187. [Google Scholar] [CrossRef]
- López-Puertollano, D.; Agulló, C.; Mercader, J.V.; Abad-Somovilla, A.; Abad-Fuentes, A. Immunoanalytical methods for ochratoxin A monitoring in wine and must based on innovative immunoreagents. Food Chem. 2020, 345, 128828. [Google Scholar] [CrossRef]
- Myresiotis, C.K.; Testempasis, S.; Vryzas, Z.; Karaoglanidis, G.S.; Papadopoulou-Mourkidou, E. Determination of mycotoxins in pomegranate fruits and juices using a QuEChERS-based method. Food Chem. 2015, 182, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Van Der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a Toxic Metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 Years of Research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabañes, F.J.; Bragulat, M.R.; Castellá, G. Ochratoxin A Producing Species in the Genus Penicillium. Toxins 2010, 2, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, M.; Rodríguez, A.; Peromingo, B.; Núñez, F.; Rodríguez, M. Enterococcus faecium: A promising protective culture to control growth of ochratoxigenic moulds and mycotoxin production in dry-fermented sausages. Mycotoxin Res. 2019, 36, 137–145. [Google Scholar] [CrossRef]
- Amézqueta, S.; Schorr-Galindo, S.; Murillo-Arbizu, M.; González-Peñas, E.; de Cerain, A.L.; Guiraud, J. OTA-producing fungi in foodstuffs: A review. Food Control 2012, 26, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Zimmerli, B.; Dick, R. Ochratoxin A in table wine and grape-juice: Occurrence and risk assessment∗. Food Addit. Contam. 1996, 13, 655–668. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Wine Contamination with Ochratoxins: A Review. Beverages 2018, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.J.; Rodrigues, A.P.; Pereira, A.M.P.T.; Lino, C.; Pena, A. Ochratoxin A in the Portuguese Wine Market, Occurrence and Risk Assessment. Food Addit. Contam. Part B 2019, 12, 145–149. [Google Scholar] [CrossRef]
- Kochman, J.; Jakubczyk, K.; Janda, K. Mycotoxins in red wine: Occurrence and risk assessment. Food Control 2021, 129, 108229. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, L.; Li, J.; Zhu, B.; Ma, L. Occurrence of Ochratoxin A in Chinese wines: Influence of local meteorological parameters. Eur. Food Res. Technol. 2012, 236, 277–283. [Google Scholar] [CrossRef]
- De Jesus, C.L.; Bartley, A.; Welch, A.Z.; Berry, J.P. High Incidence and Levels of Ochratoxin A in Wines Sourced from the United States. Toxins 2018, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajok, I.; Kowalska, A.; Piekut, A.; Ćwieląg-Drabek, M. A risk assessment of dietary exposure to ochratoxin A for the Polish population. Food Chem. 2019, 284, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.Z. Mycotoxins in food, recent development in food analysis and future challenges; a review. Curr. Opin. Food Sci. 2021, 42, 237–247. [Google Scholar] [CrossRef]
- Saadullah, A.A.M.; A Bdullah, S.K. Fungal contamination of dried vine fruits and ochratoxin a detection in grape juice from Duhok, Iraq. Rev. Innovaciencia 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Wei, D.; Wu, X.; Xu, J.; Dong, F.; Liu, X.; Zheng, Y.; Ji, M. Determination of Ochratoxin A contamination in grapes, processed grape products and animal-derived products using ultra-performance liquid chromatography-tandem mass spectroscopy system. Sci. Rep. 2018, 8, 2051. [Google Scholar] [CrossRef]
- Yusefi, J.; Valaee, M.; Nazari, F.; Maleki, J.; Mottaghianpour, E.; Khosrokhavar, R.; Hosseini, M.J. Occurrence of ochratoxin a in grape juice of Iran. Iran. J. Pharm. Res. IJPR 2018, 17, 140–146. [Google Scholar]
- Oteiza, J.M.; Khaneghah, A.M.; Campagnollo, F.B.; Granato, D.; Mahmoudi, M.R.; Sant’Ana, A.S.; Gianuzzi, L. Influence of production on the presence of patulin and ochratoxin A in fruit juices and wines of Argentina. LWT 2017, 80, 200–207. [Google Scholar] [CrossRef]
- Lucchetta, G.; Bazzo, I.; Cortivo, G.D.; Stringher, L.; Bellotto, D.; Borgo, M.; Angelini, E. Occurrence of Black Aspergilli and Ochratoxin A on Grapes in Italy. Toxins 2010, 2, 840–855. [Google Scholar] [CrossRef] [Green Version]
- Sanzani, S.M.; Djenane, F.; Incerti, O.; Admane, N.; Mincuzzi, A.; Ippolito, A. Mycotoxigenic fungi contaminating greenhouse-grown tomato fruit and their alternative control. Eur. J. Plant Pathol. 2021, 160, 287–300. [Google Scholar] [CrossRef]
- Wei, C.; Dai, X.; Guo, L.; Lei, X. Identification and toxin-producing capability of causing-spoilage fungi in strawberry. J. Food Saf. Food Qual. 2017, 8, 1721–1726. [Google Scholar] [CrossRef]
- Al-Hazmi, N. Determination of Patulin and Ochratoxin A using HPLC in apple juice samples in Saudi Arabia. Saudi J. Biol. Sci. 2010, 17, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heperkan, D.; Güler, F.K.; Oktay, H. Mycoflora and natural occurrence of aflatoxin, cyclopiazonic acid, fumonisin and ochratoxin A in dried figs. Food Addit. Contam. Part A 2012, 29, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, E.; Shakerian, A. Ochratoxin A in dried figs, raisings, apricots, dates on Iranian retail market. Health 2013, 5, 2077–2080. [Google Scholar] [CrossRef] [Green Version]
- Pavón, M.A.; González, I.; Martín, R.; Garcia, T. Competitive direct ELISA based on a monoclonal antibody for detection of Ochratoxin A in dried fig samples. Food Agric. Immunol. 2012, 23, 83–91. [Google Scholar] [CrossRef]
- Gupta, D.; Bala, P.; Sharma, Y.P. Evaluation of Fungal Flora and Mycotoxin Contamination in Whole Dried Apricots (Prunus armeniaca L.) from J&K, India. Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2015, 87, 81–87. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Mehmood, Z.; Asi, M.R.; Shahid, M.; Sehar, M.; Malik, N. Co-occurrence of aflatoxins and ochratoxin A in nuts, dry fruits, and nuty products. J. Food Saf. 2018, 38, e12462. [Google Scholar] [CrossRef]
- Heshmati, A.; Zohrevand, T.; Khaneghah, A.M.; Nejad, A.S.M.; Sant’Ana, A.S. Co-occurrence of aflatoxins and ochratoxin A in dried fruits in Iran: Dietary exposure risk assessment. Food Chem. Toxicol. 2017, 106, 202–208. [Google Scholar] [CrossRef]
- Tosun, A.; Ozden, S. Ochratoxin A in red pepper flakes commercialised in Turkey. Food Addit. Contam. Part B 2015, 9, 46–50. [Google Scholar] [CrossRef]
- Ok, H.E.; Chung, S.H.; Lee, N.; Chun, H.S. Simple High-Performance Liquid Chromatography Method for the Simultaneous Analysis of Aflatoxins, Ochratoxin A, and Zearalenone in Dried and Ground Red Pepper. J. Food Prot. 2015, 78, 1226–1231. [Google Scholar] [CrossRef]
- Spadaro, D.; Patharajan, S.; Lore, A.; Gullino, M.L.; Garibaldi, A. Effect of pH, water activity and temperature on the growth and accumulation of ochratoxin A produced by three strains of Aspergillus carbonarius isolated from Italian vineyards. Phytopathol. Mediterr. 2010, 49, 65–73. [Google Scholar] [CrossRef]
- Welke, J.E. Fungal and mycotoxin problems in grape juice and wine industries. Curr. Opin. Food Sci. 2019, 29, 7–13. [Google Scholar] [CrossRef]
- Erceg, S.; Mateo, E.M.; Zipancic, I.; Jiménez, F.J.R.; Aragó, M.A.P.; Jiménez, M.; Soria, J.M.; Garcia-Esparza, M.Á. Assessment of toxic efects of ochratoxin A in human embryonic stem Cells. Toxins 2019, 11, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirot, V.; Fremy, J.-M.; Leblanc, J.-C. Dietary exposure to mycotoxins and health risk assessment in the second French total diet study. Food Chem. Toxicol. 2012, 52, 1–11. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, Y.S.; Khalil, R.; Saad, T.T. Acute toxicity of ochratoxin-A in marine water-reared sea bass (Dicentrarchus labrax L.). Chemosphere 2009, 75, 878–882. [Google Scholar] [CrossRef]
- Gresham, A.; Done, S.; Livesey, C.; MacDonald, S.; Chan, D.; Sayers, R.; Clark, C.; Kemp, P. Survey of pigs’ kidneys with lesions consistent with PMWS and PDNS and ochratoxicosis. Part 1: Concentrations and prevalence of ochratoxin A. Veter. Rec. 2006, 159, 737–742. [Google Scholar] [CrossRef]
- Ringot, D.; Chango, A.; Schneider, Y.-J.; Larondelle, Y. Toxicokinetics and toxicodynamics of ochratoxin A, an update. Chem. Interact. 2006, 159, 18–46. [Google Scholar] [CrossRef]
- Huff, W.E.; Kubena, L.F.; Harvey, R.B.; Phillips, T.D. Efficacy of Hydrated Sodium Calcium Aluminosilicate to Reduce the Individual and Combined Toxicity of Aflatoxin and Ochratoxin A. Poult. Sci. 1992, 71, 64–69. [Google Scholar] [CrossRef]
- IARC. Monographs on the Evaluation of Carcinogenic Risks to Humans: Some Naturally Occuring Substances: Food Items and Costituents, Heterocyclic Aromatic Amines and Mycotoxins; IARC: Lyon, France, 1993; Volume 56, pp. 489–524. [Google Scholar]
- Li, H.; Mao, X.; Liu, K.; Sun, J.; Li, B.; Malyar, R.M.; Liu, D.; Pan, C.; Gan, F.; Liu, Y.; et al. Ochratoxin A induces nephrotoxicity in vitro and in vivo via pyroptosis. Arch. Toxicol. 2021, 95, 1489–1502. [Google Scholar] [CrossRef]
- Imaoka, T.; Yang, J.; Wang, L.; McDonald, M.G.; Afsharinejad, Z.; Bammler, T.K.; Van Ness, K.; Yeung, C.K.; Rettie, A.E.; Himmelfarb, J.; et al. Microphysiological system modeling of ochratoxin A-associated nephrotoxicity. Toxicology 2020, 444, 152582. [Google Scholar] [CrossRef]
- Niaz, K.; Shah, S.Z.A.; Khan, F.; Bule, M. Ochratoxin A–induced genotoxic and epigenetic mechanisms lead to Alzheimer disease: Its modulation with strategies. Environ. Sci. Pollut. Res. 2020, 27, 44673–44700. [Google Scholar] [CrossRef] [PubMed]
- Raistrick, H. Patulin in the Common Cold Collaborative Research on a Derivative of Penicillium Patulum Bainier: I. Introduction. Lancet 1943, 242, 625. [Google Scholar] [CrossRef]
- Andersen, B.; Smedsgaard, A.J.; Frisvad, J.C. Penicillium expansum: Consistent Production of Patulin, Chaetoglobosins, and Other Secondary Metabolites in Culture and Their Natural Occurrence in Fruit Products. J. Agric. Food Chem. 2004, 52, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Saleh, I.; Goktepe, I. The characteristics, occurrence, and toxicological effects of patulin. Food Chem. Toxicol. 2019, 129, 301–311. [Google Scholar] [CrossRef]
- Paster, N.; Barkai-Golan, R. Mouldy fruits and vegetables as a source of mycotoxins: Part 2. World Mycotoxin J. 2008, 4, 385–396. [Google Scholar] [CrossRef]
- Sajid, M.; Mehmood, S.; Yuan, Y.; Yue, T. Mycotoxin patulin in food matrices: Occurrence and its biological degradation strategies. Drug Metab. Rev. 2019, 51, 105–120. [Google Scholar] [CrossRef]
- Wright, S.A. Patulin in food. Curr. Opin. Food Sci. 2015, 5, 105–109. [Google Scholar] [CrossRef]
- Wei, C.; Yu, L.; Qiao, N.; Zhao, J.; Zhang, H.; Zhai, Q.; Tian, F.; Chen, W. Progress in the distribution, toxicity, control, and detoxification of patulin: A review. Toxicon 2020, 184, 83–93. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, H.; Yang, Q.; Ren, R. Efficacy of Pichia caribbica in controlling blue mold rot and patulin degradation in apples. Int. J. Food Microbiol. 2013, 162, 167–173. [Google Scholar] [CrossRef]
- Hammami, W.; Al-Thani, R.; Fiori, S.; Al-Meer, S.; Atia, F.A.; Rabah, D.; Migheli, Q.; Jaoua, S. Patulin and patulin producing Penicillium spp. occurrence in apples and apple-based products including baby food. J. Infect. Dev. Ctries. 2017, 11, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Zaied, C.; Abid, S.; Hlel, W.; Bacha, H. Occurrence of patulin in apple-based-foods largely consumed in Tunisia. Food Control 2012, 31, 263–267. [Google Scholar] [CrossRef]
- Zouaoui, N.; Sbaii, N.; Bacha, H.; Abid-Essefi, S. Occurrence of patulin in various fruit juice marketed in Tunisia. Food Control 2015, 51, 356–360. [Google Scholar] [CrossRef]
- Van de Perre, E.; Jacxsens, L.; Van Der Hauwaert, W.; Haesaert, I.; De Meulenaer, B. Screening for the Presence of Patulin in Molded Fresh Produce and Evaluation of Its Stability in the Production of Tomato Products. J. Agric. Food Chem. 2013, 62, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.Z.; Malik, S.; Asi, M.R.; Selamat, J.; Malik, N. Natural occurrence of patulin in different fruits, juices and smoothies and evaluation of dietary intake in Punjab, Pakistan. Food Control 2018, 84, 370–374. [Google Scholar] [CrossRef]
- Sarubbi, F.; Formisano, G.; Auriemma, G.; Arrichiello, A.; Palomba, R. Patulin in homogenized fruit’s and tomato products. Food Control 2016, 59, 420–423. [Google Scholar] [CrossRef]
- Ji, X.; Li, R.; Yang, H.; Qi, P.; Xiao, Y.; Qian, M. Occurrence of patulin in various fruit products and dietary exposure assessment for consumers in China. Food Control 2017, 78, 100–107. [Google Scholar] [CrossRef]
- Aboud, H.M.; Shatha, A.; Shafiq, S.; Jabbar, I.S. Investigation of mycotoxin patulin in some types of dried fruits in Baghdad governorate. Int. J. Adv. Res. 2015, 3, 1128–1135. [Google Scholar]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and Toxicological Effects of Patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Jin, C.; Gan, J.; Wang, X.; Shi, Z.; Xia, X.; Peng, X. Apigenin attenuates patulin-induced apoptosis in HEK293 cells by modulating ROS-mediated mitochondrial dysfunction and caspase signal pathway. Toxicon 2017, 137, 106–113. [Google Scholar] [CrossRef]
- Boussabbeh, M.; Ben Salem, I.; Belguesmi, F.; Bacha, H.; Abid-Essefi, S. Tissue oxidative stress induced by patulin and protective effect of crocin. NeuroToxicology 2016, 53, 343–349. [Google Scholar] [CrossRef]
- Alves, I.; Oliveira, N.; Laires, A.; Rodrigues, A.; Rueff, J. Induction of micronuclei and chromosomal aberrations by the mycotoxin patulin in mammalian cells: Role of ascorbic acid as a modulator of patulin clastogenicity. Mutagenesis 2000, 15, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Yang, J.; Zhang, B.; Zhang, L.; Wu, K.; Yang, A.; Li, C.; Wang, Y.; Zhang, J.; Qi, D. Potential Link between Gut Microbiota and Deoxynivalenol-Induced Feed Refusal in Weaned Piglets. J. Agric. Food Chem. 2019, 67, 4976–4986. [Google Scholar] [CrossRef] [PubMed]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Nakashima, T.; Ueno, T.; Fukami, H. Structure elucidation of AK-toxins, host-specific phytotoxic metabolites produced by alternaria kikuchiana tanaka. Tetrahedron Lett. 1982, 23, 4469–4472. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Cirlini, M.; Falavigna, C. Mycotoxins from alternaria: Toxicological implications. In Advances in Molecular Toxicology; University of Parma: Parma, Italy, 2014; Volume 8, pp. 107–121. [Google Scholar] [CrossRef]
- King, A.D.; Schade, J.E. Alternaria Toxins and Their Importance in Food. J. Food Prot. 1984, 47, 886–901. [Google Scholar] [CrossRef]
- Mamgain, A.; Roychowdhury, R.; Tah, J. Review alternaria pathogenicity and its strategic controls. Res. J. Biol. 2013, 1, 1–9. [Google Scholar]
- Fleck, S.C.; Burkhardt, B.; Pfeiffer, E.; Metzler, M. Alternaria toxins: Altertoxin II is a much stronger mutagen and DNA strand breaking mycotoxin than alternariol and its methyl ether in cultured mammalian cells. Toxicol. Lett. 2012, 214, 27–32. [Google Scholar] [CrossRef]
- Stinson, E.E.; Osman, S.F.; Heisler, E.G.; Siciliano, J.; Bills, D.D. Mycotoxin production in whole tomatoes, apples, oranges, and lemons. J. Agric. Food Chem. 1981, 29, 790–792. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in Occurrence, Importance, and Mycotoxin Control Strategies: Prevention and Detoxification in Foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef]
- Hickert, S.; Bergmann, M.; Ersen, S.; Cramer, B.; Humpf, H.-U. Survey of Alternaria toxin contamination in food from the German market, using a rapid HPLC-MS/MS approach. Mycotoxin Res. 2015, 32, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Shao, B.; Yang, D.; Li, F. Natural Occurrence of Four Alternaria Mycotoxins in Tomato- and Citrus-Based Foods in China. J. Agric. Food Chem. 2014, 63, 343–348. [Google Scholar] [CrossRef] [PubMed]
- EFSA on Contaminants in the Food Chain (CONTAM). Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Gonçalves, C.; Tölgyesi, Á.; Bouten, K.; Robouch, P.; Emons, H.; Stroka, J. Determination of Alternaria Toxins in Tomato, Wheat, and Sunflower Seeds by SPE and LC-MS/MS—A Method Validation Through a Collaborative Trial. J. AOAC Int. 2021, 105, 80–94. [Google Scholar] [CrossRef]
- Guiso, N.; von Konig, C.H.W.; Becker, C.; Hallander, H. Fimbrial Typing of Bordetella pertussis Isolates: Agglutination with Polyclonal and Monoclonal Antisera. J. Clin. Microbiol. 2001, 39, 1684–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, P.; Venema, D.; De Rijk, T.; De Kok, A.; Scholten, J.M.; Mol, H.G.; De Nijs, M. Occurrence of Alternaria toxins in food products in The Netherlands. Food Control 2016, 60, 196–204. [Google Scholar] [CrossRef]
- Asam, S.; Konitzer, K.; Rychlik, M. Precise determination of the Alternaria mycotoxins alternariol and alternariol monomethyl ether in cereal, fruit and vegetable products using stable isotope dilution assays. Mycotoxin Res. 2010, 27, 23–28. [Google Scholar] [CrossRef]
- Guo, W.; Fan, K.; Nie, D.; Meng, J.; Huang, Q.; Yang, J.; Shen, Y.; Tangni, E.K.; Zhao, Z.; Wu, Y.; et al. Development of a QuEChERS-Based UHPLC-MS/MS Method for Simultaneous Determination of Six Alternaria Toxins in Grapes. Toxins 2019, 11, 87. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Wang, Y.; Jiang, D.; Feng, X.; Li, J.; Wang, M. Survey of Alternaria Toxins and Other Mycotoxins in Dried Fruits in China. Toxins 2017, 9, 200. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, Y.; Curtui, V.; Dietrich, R.; Gross, M.; Latif, H.; Märtlbauer, E.; Usleber, E. Widespread Occurrence of Low Levels of Alternariol in Apple and Tomato Products, as Determined by Comparative Immunochemical Assessment using Monoclonal and Polyclonal Antibodies. J. Agric. Food Chem. 2011, 59, 6360–6368. [Google Scholar] [CrossRef]
- Scott, P.M.; Lawrence, G.A.; Lau, B.P.Y. Analysis of wines, grape juices and cranberry juices forAlternaria toxins. Mycotoxin Res. 2006, 22, 142–147. [Google Scholar] [CrossRef]
- Walravens, J.; Mikula, H.; Rychlik, M.; Asam, S.; Devos, T.; Ediage, E.N.; Di Mavungu, J.D.; Jacxsens, L.; Van Landschoot, A.; Vanhaecke, L.; et al. Validated UPLC-MS/MS Methods To Quantitate Free and Conjugated Alternaria Toxins in Commercially Available Tomato Products and Fruit and Vegetable Juices in Belgium. J. Agric. Food Chem. 2016, 64, 5101–5109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babič, J.; Tavčar-Kalcher, G.; Celar, F.; Kos, K.; Knific, T.; Jakovac-Strajn, B. Occurrence of Alternaria and Other Toxins in Cereal Grains Intended for Animal Feeding Collected in Slovenia: A Three-Year Study. Toxins 2021, 13, 304. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Ma, L.; Jiang, T.; Jiang, L.Y.; Dai, F.F.; Zhang, Y.H. A review on tenuazonic acid, a toxic produced by alternaria. Food Sci. 2014, 35, 295–301. [Google Scholar] [CrossRef]
- Pero, R.W.; Posner, H.; Blois, M.; Harvan, D.; Spalding, J.W. Toxicity of Metabolites Produced by the Alternaria. Environ. Health Perspect. 1973, 4, 87. [Google Scholar] [CrossRef] [PubMed]
- Sauer, D.B.; Seitz, L.M.; Burroughs, R.; Mohr, H.E.; West, J.L.; Milleret, R.J.; Anthony, H.D. Toxicity of Alternaria metabolites found in weathered sorghum grain at harvest. J. Agric. Food Chem. 1978, 26, 1380–1383. [Google Scholar] [CrossRef]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria Mycotoxins in Food and Feed: An Overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef] [Green Version]
- Fehr, M.; Pahlke, G.; Fritz, J.; Christensen, M.O.; Boege, F.; Altemöller, M.; Podlech, J.; Marko, D. Alternariol acts as a topoisomerase poison, preferentially affecting the IIα isoform. Mol. Nutr. Food Res. 2009, 53, 441–451. [Google Scholar] [CrossRef]
- Víctor-Ortega, M.D.; Lara, F.J.; García-Campaña, A.M.; del Olmo-Iruela, M. Evaluation of dispersive liquid–liquid microextraction for the determination of patulin in apple juices using micellar electrokinetic capillary chromatography. Food Control 2013, 31, 353–358. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.; Korzeniowska, M.; Guiné, R.P. Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, Q.; Kuča, K.; Dohnal, V.; Tian, Z. Deoxynivalenol: Signaling pathways and human exposure risk assessment—An update. Arch. Toxicol. 2014, 88, 1915–1928. [Google Scholar] [CrossRef]
- Udovicki, B.; Audenaert, K.; De Saeger, S.; Rajkovic, A. Overview on the Mycotoxins Incidence in Serbia in the Period 2004–2016. Toxins 2018, 10, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahato, D.K.; Pandhi, S.; Kamle, M.; Gupta, A.; Sharma, B.; Panda, B.K.; Srivastava, S.; Kumar, M.; Selvakumar, R.; Pandey, A.K.; et al. Trichothecenes in food and feed: Occurrence, impact on human health and their detection and management strategies. Toxicon 2022, 208, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Bi, I.; Zong, Y.Y.; Pu, L.; Wang, Y.; Li, Y. Progress of mycotoxins contamination and detection in fruits and vegetables and their products. Food Sci. 2016, 37, 285–290. [Google Scholar] [CrossRef]
- Xue, H.-L.; Bi, Y.; Tang, Y.-M.; Zhao, Y.; Wang, Y. Effect of cultivars, Fusarium strains and storage temperature on trichothecenes production in inoculated potato tubers. Food Chem. 2013, 151, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, X.; Han, S.-Y.; Li, M.; Ma, T.-Z.; Sheng, W.-J.; Zhu, X. Simultaneous Analysis of 20 Mycotoxins in Grapes and Wines from Hexi Corridor Region (China): Based on a QuEChERS–UHPLC–MS/MS Method. Molecules 2018, 23, 1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Xue, H.; Bi, Y.; Li, Y.; Wang, Y.; Zhao, Y.; Shen, K. A method of analysis for T-2 toxin and neosolaniol by UPLC-MS/MS in apple fruit inoculated with Trichothecium roseum. Food Addit. Contam. Part A 2014, 32, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Bi, Y.; Sun, Y.; Hussain, R.; Wang, H.; Zhang, S.; Zhang, R.; Long, H.; Nan, M.; Cheng, X.; et al. Acetylsalicylic acid treatment reduce Fusarium rot development and neosolaniol accumulation in muskmelon fruit. Food Chem. 2019, 289, 278–284. [Google Scholar] [CrossRef]
- Xue, H.L.; Bi, Y.; Hussain, R.; Pu, L.M.; Nan, M.N.; Cheng, X.Y.; Wang, Y.; Li, Y.C. Detection of NEO in muskmelon fruits inoculated with Fusarium sulphureum and its control by postharvest ozone treatment. Food Chem. 2018, 254, 193–200. [Google Scholar] [CrossRef]
- Zhu, M.; Cen, Y.; Ye, W.; Li, S.; Zhang, W. Recent Advances on Macrocyclic Trichothecenes, Their Bioactivities and Biosynthetic Pathway. Toxins 2020, 12, 417. [Google Scholar] [CrossRef]
- Cope, R.B. Chapter 75—Trichothecenes. In Veterinary Toxicology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 75, pp. 1043–1053. [Google Scholar] [CrossRef]
- Sokolović, M.; Garaj-Vrhovac, V.; Šimpraga, B. T-2 Toxin: Incidence and Toxicity in Poultry. Arch. Ind. Hyg. Toxicol. 2008, 59, 43–52. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, H.-R.; Bursian, S.J.; Link, J.E.; Pestka, J.J. Emetic responses to T-2 toxin, HT-2 toxin and emetine correspond to plasma elevations of peptide YY3–36 and 5-hydroxytryptamine. Arch. Toxicol. 2015, 90, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Kuca, K.; Dohnal, V.; Jezkova, A.; Jun, D. Metabolic Pathways of T-2 Toxin. Curr. Drug Metab. 2008, 9, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Daenicke, S.; Keese, C.; Goyarts, T.; Döll, S. Effects of deoxynivalenol (DON) and related compounds on bovine peripheral blood mononuclear cells (PBMC) in vitro and in vivo. Mycotoxin Res. 2010, 27, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Hymery, N.; Sibiril, Y.; Parent-Massin, D. In vitro effects of trichothecenes on human dendritic cells. Toxicol. Vitr. 2006, 20, 899–909. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Dadak, A.; Gille, L.; Staniek, K.; Hess, M.; Böhm, J. Genotoxic effects of deoxynivalenol in broiler chickens fed low-protein feeds. Poult. Sci. 2012, 91, 550–555. [Google Scholar] [CrossRef]
- Pinton, P.; Oswald, I.P. Effect of Deoxynivalenol and Other Type B Trichothecenes on the Intestine: A Review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef]
- Desjardins, A.; Proctor, R. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef]
- Becker, C.; Reiter, M.; Pfaffl, M.W.; Meyer, H.H.D.; Bauer, J.; Meyer, K.H.D. Expression of immune relevant genes in pigs under the influence of low doses of deoxynivalenol (DON). Mycotoxin Res. 2011, 27, 287–293. [Google Scholar] [CrossRef]
- Zhao, Y.R.; Ma, L.Y.; Wang, F.H. Research progress in trichothecenes and removal method. Sci. Technol. Food Ind. 2016, 37, 383–387. [Google Scholar]
- Singh, J.; Mehta, A. Rapid and sensitive detection of mycotoxins by advanced and emerging analytical methods: A review. Food Sci. Nutr. 2020, 8, 2183–2204. [Google Scholar] [CrossRef] [PubMed]
- Rhouati, A.; Yang, C.; Hayat, A.; Marty, J.-L. Aptamers: A Promising Tool for Ochratoxin A Detection in Food Analysis. Toxins 2013, 5, 1988–2008. [Google Scholar] [CrossRef] [PubMed]
- Man, Y.; Liang, G.; Li, A.; Pan, L. Recent Advances in Mycotoxin Determination for Food Monitoring via Microchip. Toxins 2017, 9, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO 8128-1-1993; Apple Juice, Apple Juice Concentrates and Drinks Containing Apple Juice-Determination of Patulincontent. Part1: Method Using High-Performance Liquid Chromatography. ISO International Standard (ISO): Geneva, Switzerland, 1993.
- EN 15829: 2010; Foodstuffs-Determination of Ochratoxin A in Currants, Raisins, Sultanas, Mixed Dried Fruit and Dried Figs-HPLC Method with Immunoaffinity Column Cleanup and Fluorescence Detection. The Spanish Association for Standardization and Certification: Madrid, Spain, 2010.
- AOAC 2001.01; Determination of Ochratoxin A in Wine and Beer Immunoaffinitycolumn Cleanup/Liquid Chromatographic Analysis. Association of Official Agricultural Chemists: Washington, DC, USA, 2001.
- Wang, M.; Jiang, N.; Dai, Y.; Feng, X. Maximum residue levels and testing standards of mycotoxins in fruits in china and abroad. J. Food Saf. Qual. 2016, 7, 459–467. (In Chinese) [Google Scholar]
- Tölgyesi, Á.; Farkas, T.; Bálint, M.; McDonald, T.J.; Sharma, V.K. A Dilute and Shoot Strategy for Determining Alternaria Toxins in Tomato-Based Samples and in Different Flours Using LC-IDMS Separation. Molecules 2021, 26, 1017. [Google Scholar] [CrossRef]
- Pallarés, N.; Sebastià, A.; Martínez-Lucas, V.; González-Angulo, M.; Barba, F.J.; Berrada, H.; Ferrer, E. High Pressure Processing Impact on Alternariol and Aflatoxins of Grape Juice and Fruit Juice-Milk Based Beverages. Molecules 2021, 26, 3769. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Agrawal, D.C.; Yang, W.-G.; Chien, W.-J. A simple HPLC-MS/MS method for the analysis of multi-mycotoxins in betel nut. Int. J. Appl. Sci. Eng. 2021, 18, 1–7. [Google Scholar] [CrossRef]
- Bazin, I.; Tria, S.A.; Hayat, A.; Marty, J.-L. New biorecognition molecules in biosensors for the detection of toxins. Biosens. Bioelectron. 2017, 87, 285–298. [Google Scholar] [CrossRef]
- Vidal, J.C.; Bonel, L.; Ezquerra, A.; Hernández, S.; Bertolín, J.R.; Cubel, C.; Castillo, J.R. Electrochemical affinity biosensors for detection of mycotoxins: A review. Biosens. Bioelectron. 2013, 49, 146–158. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, M.; Kang, Y.; Xie, H.; Wang, X.; Song, H.; Li, X.; Fang, W. Identification of a high-affinity monoclonal antibody against ochratoxin A and its application in enzyme-linked immunosorbent assay. Toxicon 2015, 106, 89–96. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Fu, R.; Liu, H.; Zhou, J.; Zhao, Q.; Wang, C.; Jiao, B.; He, Y. A review of portable biosensor for the field detection of mycotoxins. Food Sci. 2022, in press. [Google Scholar]
- Yang, X.-H.; Kong, W.-J.; Yang, M.-H.; Zhao, M.; Ouyang, Z. Application of Aptamer Identification Technology in Rapid Analysis of Mycotoxins. Chin. J. Anal. Chem. 2013, 41, 297–306. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Y.; Duan, N.; Wu, S.; Xia, Y.; Ma, X.; Zhu, C.; Jiang, Y.; Wang, Z. Screening and Identification of DNA Aptamers against T-2 Toxin Assisted by Graphene Oxide. J. Agric. Food Chem. 2014, 62, 10368–10374. [Google Scholar] [CrossRef]
- Zhu, Z.; Feng, M.; Zuo, L.; Zhu, Z.; Wang, F.; Chen, L.; Li, J.; Shan, G.; Luo, S.-Z. An aptamer based surface plasmon resonance biosensor for the detection of ochratoxin A in wine and peanut oil. Biosens. Bioelectron. 2015, 65, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Annida, R.M.; Zuilhof, H.; Van Beek, T.A.; Nielen, M.W.F. Analysis of Mycotoxins in Beer Using a Portable Nanostructured Imaging Surface Plasmon Resonance Biosensor. J. Agric. Food Chem. 2016, 64, 8263–8271. [Google Scholar] [CrossRef] [PubMed]
- Nolan, P.; Auer, S.; Spehar, A.; Elliott, C.T.; Campbell, K. Current trends in rapid tests for mycotoxins. Food Addit. Contam. Part A 2019, 36, 800–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirinçci, Ş.Ş.; Ertekin, Ö.; Laguna, D.E.; Özen, F.Ş.; Öztürk, Z.Z.; Öztürk, S. Label-Free QCM Immunosensor for the Detection of Ochratoxin A. Sensors 2018, 18, 1161. [Google Scholar] [CrossRef] [Green Version]
- Karczmarczyk, A.; Haupt, K.; Feller, K.H. Development of a QCM-D biosensor for Ochratoxin A detection in red wine. Talanta 2017, 166, 193–197. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Nan, M.; Li, Y.; Yun, J.; Wang, Y.; Bi, Y. Novel colorimetric aptasensor based on unmodified gold nanoparticle and ssDNA for rapid and sensitive detection of T-2 toxin. Food Chem. 2021, 348, 129128. [Google Scholar] [CrossRef]
- Yang, C.; Wang, Y.; Marty, J.-L.; Yang, X. Aptamer-based colorimetric biosensing of Ochratoxin A using unmodified gold nanoparticles indicator. Biosens. Bioelectron. 2011, 26, 2724–2727. [Google Scholar] [CrossRef]
- He, Y.; Tian, F.; Zhou, J.; Zhao, Q.; Fu, R.; Jiao, B. Colorimetric aptasensor for ochratoxin A detection based on enzyme-induced gold nanoparticle aggregation. J. Hazard. Mater. 2019, 388, 121758. [Google Scholar] [CrossRef] [PubMed]
- Rhouati, A.; Catanante, G.; Nunes, G.; Hayat, A.; Marty, J.-L. Label-Free Aptasensors for the Detection of Mycotoxins. Sensors 2016, 16, 2178. [Google Scholar] [CrossRef] [PubMed]
- Rhouati, A.; Hayat, A.; Hernandez, D.B.; Meraihi, Z.; Munoz, R.; Marty, J.-L. Development of an automated flow-based electrochemical aptasensor for on-line detection of Ochratoxin A. Sensors Actuators B Chem. 2013, 176, 1160–1166. [Google Scholar] [CrossRef]
- Abnous, K.; Danesh, N.M.; Alibolandi, M.; Ramezani, M.; Taghdisi, S.M. Amperometric aptasensor for ochratoxin A based on the use of a gold electrode modified with aptamer, complementary DNA, SWCNTs and the redox marker Methylene Blue. Mikrochim. Acta 2017, 184, 1151–1159. [Google Scholar] [CrossRef]
- He, B.; Lu, X. An electrochemical aptasensor based on tetrahedral DNA nanostructures as a signal probe carrier platform for sensitive detection of patulin. Anal. Chim. Acta 2020, 1138, 123–131. [Google Scholar] [CrossRef]
- Song, X.; Wang, D.; Kim, M. Development of an immuno-electrochemical glass carbon electrode sensor based on graphene oxide/gold nanocomposite and antibody for the detection of patulin. Food Chem. 2020, 342, 128257. [Google Scholar] [CrossRef]
- Xu, J.; Qiao, X.; Wang, Y.; Sheng, Q.; Yue, T.; Zheng, J.; Zhou, M. Electrostatic assembly of gold nanoparticles on black phosphorus nanosheets for electrochemical aptasensing of patulin. Mikrochim. Acta 2019, 186, 238. [Google Scholar] [CrossRef]
- Nan, M.; Bi, Y.; Xue, H.; Xue, S.; Long, H.; Pu, L.; Fu, G. Rapid Determination of Ochratoxin A in Grape and Its Commodities Based on a Label-Free Impedimetric Aptasensor Constructed by Layer-by-Layer Self-Assembly. Toxins 2019, 11, 71. [Google Scholar] [CrossRef] [Green Version]
- Nan, M.-N.; Bi, Y.; Xue, H.-L.; Long, H.-T.; Xue, S.-L.; Pu, L.-M.; Prusky, D. Modification performance and electrochemical characteristics of different groups of modified aptamers applied for label-free electrochemical impedimetric sensors. Food Chem. 2020, 337, 127761. [Google Scholar] [CrossRef]
- Meulenberg, E.P. Immunochemical Methods for Ochratoxin A Detection: A Review. Toxins 2012, 4, 244–266. [Google Scholar] [CrossRef]
- Wang, F.; Wan, D.-B.; Shen, Y.-D.; Tian, Y.-X.; Xiao, Z.-L.; Xu, Z.-L.; Yang, J.-Y.; Sun, Y.-M.; Hammock, B.D.; Wang, H. Development of a chemiluminescence immunoassay for detection of tenuazonic acid mycotoxin in fruit juices with a specific camel polyclonal antibody. Anal. Methods 2021, 13, 1795–1802. [Google Scholar] [CrossRef] [PubMed]
- Pei, K.; Xiong, Y.; Xu, B.; Wu, K.; Li, X.; Jiang, H.; Xiong, Y. Colorimetric ELISA for ochratoxin A detection based on the urease-induced metallization of gold nanoflowers. Sensors Actuators B Chem. 2018, 262, 102–109. [Google Scholar] [CrossRef]
- Mcnamee, S.; Bravin, F.; Rosar, G.; Elliott, C.T.; Campbell, K. Development of a nanoarray capable of the rapid and simultaneous detection of zearalenone, T2-toxin and fumonisin. Talanta 2017, 164, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.; Lim, J.; Mo, C. A Review on Lateral Flow Test Strip for Food Safety. J. Biosyst. Eng. 2015, 40, 277–283. [Google Scholar] [CrossRef]
- Jiang, H.; Li, X.; Xiong, Y.; Pei, K.; Nie, L.; Xiong, Y. Silver Nanoparticle-Based Fluorescence-Quenching Lateral Flow Immunoassay for Sensitive Detection of Ochratoxin A in Grape Juice and Wine. Toxins 2017, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Ma, J.; Cheng, Y.; Wang, H.; Sun, J.; Yan, Y. One-step rapid detection of fumonisin B1, dexyonivalenol and zearalenone in grains. Food Control 2020, 117, 107107. [Google Scholar] [CrossRef]
- Foubert, A.; Beloglazova, N.V.; Gordienko, A.; Tessier, M.D.; Drijvers, E.; Hens, Z.; De Saeger, S. Development of a Rainbow Lateral Flow Immunoassay for the Simultaneous Detection of Four Mycotoxins. J. Agric. Food Chem. 2016, 65, 7121–7130. [Google Scholar] [CrossRef]
- Leggott, N.L.; Shephard, G.S.; Stockenström, S.; Staal, E.; Van Schalkwyk, D.J. The reduction of patulin in apple juice by three different types of activated carbon. Food Addit. Contam. 2001, 18, 825–829. [Google Scholar] [CrossRef]
- Huebner, H.J.; Mayura, K.; Pallaroni, L.; Ake, C.L.; Lemke, S.L.; Herrera, P.; Phillips, T.D. Development and Characterization of a Carbon-Based Composite Material for Reducing Patulin Levels in Apple Juice. J. Food Prot. 2000, 63, 106–110. [Google Scholar] [CrossRef]
- Carrasco-Sánchez, V.; Marican, A.; Vergara-Jaque, A.; Folch-Cano, C.; Comer, J.; Laurie, V.F. Polymeric substances for the removal of ochratoxin A from red wine followed by computational modeling of the complexes formed. Food Chem. 2018, 265, 159–164. [Google Scholar] [CrossRef]
- Liu, H.F.; Han, S.Y.; Sheng, W.Y.; Zhu, X.; Jiang, Y.M. Adsorption patulin from apple juice with 8 Kinds of resin. J. Gansu Agric. Univ. 2010, 3, 126–130. (In Chinese) [Google Scholar]
- Wang, Y.H.; Ma, C.Z.; Guo, Z.Y.; Zheng, Y.Y.; Xue, H.L. Study on the kineticmodel of adsorption patulin in pear juice by macroporous resin. J. Food Saf. Qual. 2021, 11, 472–478. (In Chinese) [Google Scholar]
- Liu, Q.L.; Zhang, S.; Zhang, R.; Zhai, T.T.; Fan, P.F.; Xue, H.L. Study on the adsorption of patulin in pear juice by attapulgite. J. Food Saf. Qual. 2019, 10, 2575–2580. (In Chinese) [Google Scholar]
- Zhang, S.; Zhang, R.; Xue, H.L.; Ma, Y.Y.; Bi, Y.; Zong, Y.Y. Adsorption efficiency of attapulgite towards patulin in pear juice. Food Sci. 2019, 40, 57–63. (In Chinese) [Google Scholar] [CrossRef]
- Ji, J.; Xie, W. Removal of aflatoxin B1 from contaminated peanut oils using magnetic attapulgite. Food Chem. 2020, 339, 128072. [Google Scholar] [CrossRef]
- Nan, M.-N.; Bi, Y.; Qiang, Y.; Xue, H.-L.; Yang, L.; Feng, L.-D.; Pu, L.-M.; Long, H.-T.; Prusky, D. Electrostatic adsorption and removal mechanism of ochratoxin A in wine via a positively charged nano-MgO microporous ceramic membrane. Food Chem. 2021, 371, 131157. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Xu, W.; Wang, H.; Liao, S.; Chen, G. Preparation of magnetic molecularly imprinted polymer for selective identification of patulin in juice. J. Chromatogr. B 2020, 1145, 122101. [Google Scholar] [CrossRef]
- Zhao, D.; Jia, J.; Yu, X.; Sun, X. Preparation and characterization of a molecularly imprinted polymer by grafting on silica supports: A selective sorbent for patulin toxin. Anal. Bioanal. Chem. 2011, 401, 2259–2273. [Google Scholar] [CrossRef]
- Sun, J.; Guo, W.; Ji, J.; Li, Z.; Yuan, X.; Pi, F.; Zhang, Y.; Sun, X. Removal of patulin in apple juice based on novel magnetic molecularly imprinted adsorbent Fe3O4@SiO2@CS-GO@MIP. LWT 2019, 118, 108854. [Google Scholar] [CrossRef]
- Aiko, V.; Mehta, A. Occurrence, detection and detoxification of mycotoxins. J. Biosci. 2015, 40, 943–954. [Google Scholar] [CrossRef]
- Dahal, S.; Lee, H.J.; Gu, K.; Ryu, D. Heat Stability of Ochratoxin A in an Aqueous Buffered Model System. J. Food Prot. 2016, 79, 1748–1752. [Google Scholar] [CrossRef] [PubMed]
- Combina, M.; Dalcero, A.; Varsavsky, E.; Torres, A.M.; Etcheverry, M.; Rodriguez, M.; Gonzalez, Q.H. Effect of heat treatments on stability of altemariol, alternariol monomethyl ether and tenuazonic acid in sunflower flour. Mycotoxin Res. 1999, 15, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Blaser, A.R.; Starkopf, L.; Deane, A.M.; Poeze, M.; Starkopf, J. Comparison of Different Definitions of Feeding Intolerance: A Retrospective Observational Study. Clin. Nutr. 2015, 34, 956–961. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, T.; Padan, S.K.; Gupta, R.; Bedi, P.; Singh, G. Comparative efficacy of microwave, visible light and ultrasound irradiation for green synthesis of dihydropyrimidinones in fruit juice medium. Recent Advances in Fundamental and Applied Sciences. In AIP Conference Proceedings; AIP Publishing LLC.: New York, NY, USA, 2017; Volume 1860, p. 020059. [Google Scholar] [CrossRef]
- El-Hawa, M.A. Patulin production of Penicillium digitatum in citrus fruits as affected by Gamma Irradiation Combined with Heat. Delta J. Sci. 1993, 17, 172–182. [Google Scholar]
- Aziz, N.H.; Moussa, L.A.; Far, F.M. reduction of fungi and mycotoxins formation in seeds by gamma-radiation. J. Food Saf. 2004, 24, 109–127. [Google Scholar] [CrossRef]
- Abdelaal, S.S. Effects of Gamma radiation, temperature and water activity on the production of alternaria mycotoxins. Egypt. J. Microbiol. 1998, 32, 379–396. [Google Scholar]
- Ibarz, R.; Garvín, A.; Falguera, V.; Pagán, J.; Garza, S.; Ibarz, A. Modelling of patulin photo-degradation by a UV multi-wavelength emitting lamp. Food Res. Int. 2014, 66, 158–166. [Google Scholar] [CrossRef]
- Ibarz, R.; Garvín, A.; Azuara, E.; Ibarz, A. Modelling of ochratoxin A photo-degradation by a UV multi-wavelength emitting lamp. LWT 2015, 61, 385–392. [Google Scholar] [CrossRef]
- Zou, Z.Y.; Huang, F.; Li, H.J. Removal of deoxynivalenol and T-2 Toxin by ultraviolet irradiation. Food Sci. 2015, 36, 7–11. [Google Scholar] [CrossRef]
- Diao, E.; Wang, J.; Li, X.; Wang, X.; Gao, D. Patulin degradation in apple juice using ozone detoxification equipment and its effects on quality. J. Food Process. Preserv. 2018, 42, e13645. [Google Scholar] [CrossRef]
- Yang, K.; Li, K.; Pan, L.; Luo, X.; Xing, J.; Wang, J.; Wang, L.; Wang, R.; Zhai, Y.; Chen, Z. Effect of Ozone and Electron Beam Irradiation on Degradation of Zearalenone and Ochratoxin A. Toxins 2020, 12, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhao, Q.; Wang, T.; Duan, X.; Li, C.; Lei, X. Degradation of Organic Pollutants Using Atmospheric Pressure Glow Discharge Plasma. Plasma Chem. Plasma Process. 2016, 36, 1011–1020. [Google Scholar] [CrossRef]
- Pu, L.; Bi, Y.; Long, H.; Xue, H.; Lu, J.; Zong, Y.; Kankam, F. Glow Discharge Plasma Efficiently Degrades T-2 Toxin in Aqueous Solution and Patulin in Apple Juice. Adv. Tech. Biol. Med. 2017, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Drusch, S.; Kopka, S.; Kaeding, J. Stability of patulin in a juice-like aqueous model system in the presence of ascorbic acid. Food Chem. 2007, 100, 192–197. [Google Scholar] [CrossRef]
- Liuzzi, V.C.; Fanelli, F.; Tristezza, M.; Haidukowski, M.; Picardi, E.; Manzari, C.; Lionetti, C.; Grieco, F.; Logrieco, A.F.; Thon, M.R.; et al. Transcriptional Analysis of Acinetobacter sp. neg1 Capable of Degrading Ochratoxin A. Front. Microbiol. 2017, 7, 2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.; Yu, L.; Qiao, N.; Wang, S.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. The characteristics of patulin detoxification by Lactobacillus plantarum 13M5. Food Chem. Toxicol. 2020, 146, 111787. [Google Scholar] [CrossRef]
- Hatab, S.; Yue, T.; Mohamad, O. Reduction of Patulin in Aqueous Solution by Lactic Acid Bacteria. J. Food Sci. 2012, 77, M238–M241. [Google Scholar] [CrossRef]
- Fuchs, S.; Sontag, G.; Stidl, R.; Ehrlich, V.; Kundi, M.; Knasmüller, S. Detoxification of patulin and ochratoxin A, two abundant mycotoxins, by lactic acid bacteria. Food Chem. Toxicol. 2008, 46, 1398–1407. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Zehner, F.; Täubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res. 2006, 50, 543–551. [Google Scholar] [CrossRef]
- Eriksen, G.S.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Young, J.C.; Zhou, T.; Yu, H.; Zhu, H.; Gong, J. Degradation of trichothecene mycotoxins by chicken intestinal microbes. Food Chem. Toxicol. 2007, 45, 136–143. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhou, T.; Young, J.C.; Boland, G.J.; Scott, P.M. Chemical and biological transformations for detoxification of trichothecene mycotoxins in human and animal food chains: A review. Trends Food Sci. Technol. 2010, 21, 67–76. [Google Scholar] [CrossRef]
- Xue, H.L.; Bi, Y.; Wang, Y.; Ge, Y.H.; Li, Y.C. Advances in toxicity, detection and detoxification of trichothecenes. Food Sci. 2013, 34, 350–355. (In Chinese) [Google Scholar] [CrossRef]
- Zhu, R.; Feussner, K.; Wu, T.; Yan, F.; Karlovsky, P.; Zheng, X. Detoxification of mycotoxin patulin by the yeast Rhodosporidium paludigenum. Food Chem. 2015, 179, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tang, H.; Yang, C.; Meng, X.; Liu, B. Detoxification of mycotoxin patulin by the yeast Rhodotorula mucilaginosa. Food Control 2018, 96, 47–52. [Google Scholar] [CrossRef]
- Tang, H.; Li, X.H.; Zhang, F.; Meng, X.H.; Liu, B.J. Biodegradation of the mycotoxin patulin in apple juice by orotate phos-phoribosyltransferase from Rhodotorula mucilaginosa. Food Control 2019, 100, 158–164. [Google Scholar] [CrossRef]
- Xing, M.; Chen, Y.; Li, B.; Tian, S. Characterization of a short-chain dehydrogenase/reductase and its function in patulin biodegradation in apple juice. Food Chem. 2021, 348, 129046. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Gu, X.Y.; Zhao, L.; Zheng, X.F.; Yang, H.J.; Li, J.; Zhang, H.Y. Biocontrol of postharvest disease of grapes and OTA accumulation by Yarrowia lipolytica Y-2. J. Chin. Inst. Food Sci. Technol. 2020, 20, 201–206. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, X.; Yang, H.; Apaliya, M.; Zhao, L.; Gu, X.; Zheng, X.; Hu, W.; Zhang, H. The mechanisms involved in ochratoxin A elimination by Yarrowia lipolytica Y-2. Ann. Appl. Biol. 2018, 173, 164–174. [Google Scholar] [CrossRef]
- Gómez-Albarrán, C.; Melguizo, C.; Patiño, B.; Vázquez, C.; Gil-Serna, J. Diversity of Mycobiota in Spanish Grape Berries and Selection of Hanseniaspora uvarum U1 to Prevent Mycotoxin Contamination. Toxins 2021, 13, 649. [Google Scholar] [CrossRef]
- Tang, H.; Peng, X.; Li, X.; Meng, X.; Liu, B. Biodegradation of mycotoxin patulin in apple juice by calcium carbonate immobilized porcine pancreatic lipase. Food Control 2018, 88, 69–74. [Google Scholar] [CrossRef]
- Liu, B.; Peng, X.; Meng, X. Effective Biodegradation of Mycotoxin Patulin by Porcine Pancreatic Lipase. Front. Microbiol. 2018, 9, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foodstuffs | Detection Method | Country | Positives (%) | OTA (µg/Kg) | Ref. | |
|---|---|---|---|---|---|---|
| Fruits and products | Peppers | HPLC-FD | Poland | 37.5% | 1.7–2.4 | [26] |
| Grapes | ELISA | Italy | 30.4% | 0.02–9.2 | [30] | |
| Strawberries | HPLC-FD | China | - | - | [32] | |
| Apple juice | HPLC-FD | Saudi Arabia | 29.41% | 100–200 | [33] | |
| Dried fruits | Raisins | HPLC-FD | Poland | 47% | 1.1–34 | [26] |
| Figs | HPLC-FD | Turkey | 48% | 0.1–15.3 | [34] | |
| Figs | ELISA | Iran | 10.4 | 2.3–14.2 | [35] | |
| Apricots | ELISA | Iran | 6.7% | 2.8 | [35] | |
| Raisins | ELISA | Iran | 44.7% | 2.9–18.2 | [35] | |
| Dates | ELISA | Iran | 10% | 1.4–3.6 | [35] | |
| Figs | ELISA | Spain | 54.3% | 3.15–277 | [36] | |
| Apricots | HPLC-UV–VIS | India | 28.57% | 194 ± 0.001 | [37] | |
| Raisins | HPLC-FD | Pakistan | 23.5% | 5.60 ± 1.34 | [38] | |
| Apricots | HPLC-FD | Pakistan | 23.1% | 3.10 ± 0.70 | [38] | |
| Plums | HPLC-FD | Pakistan | 25% | 3.90 ± 0.95 | [38] | |
| Figs | HPLC-FD | Pakistan | 21.4% | 2.10 ± 0.79 | [38] | |
| Mulberries | HPLC-FD | Iran | 45.5% | 0.4–3.4 | [39] | |
| Dates | HPLC-FD | Iran | 22.3% | 0.5–2.1 | [39] | |
| Figs | HPLC-FD | Iran | 45.5% | 0.4–12.2 | [39] | |
| Apricots | HPLC-FD | Iran | 50% | 0.75–5.5 | [39] | |
| Dried vegetables | Packed red peppers | HPLC-FD | Turkey | 87.1% | 0.6–1.0 | [40] |
| Unpacked red peppers | HPLC-FD | Turkey | 100% | 1.1–31.7 | [40] | |
| Packed red peppers | HPLC-FD | Korea | 48% | 0.23–56.30 | [41] | |
| Unpacked red peppers | HPLC-FD | Korea | 4% | 0.15–0.20 | [41] |
| Foodstuffs | Detection Method | Country | Positives (%) | PAT (µg/Kg) | Ref. | |
|---|---|---|---|---|---|---|
| Fresh fruits | Tomatoes | HPLC-UV | Belgium | 10.8 | - | [65] |
| Sweet bell peppers | HPLC-UV | Belgium | 11.4 | - | [65] | |
| Onions | HPLC-UV | Belgium | - | - | [65] | |
| Apricots | HPLC-UV-VIS | Argentina | 4.5 | 0.7 | [29] | |
| Grapes | HPLC-UV-VIS | Argentina | 10 | 28.3 | [29] | |
| Pears | HPLC-UV-VIS | Argentina | 10.7 | 54 | [29] | |
| Peaches | HPLC-UV-VIS | Argentina | 9.7 | 5 | [29] | |
| Pineapples | HPLC-UV-VIS | Argentina | - | - | [29] | |
| Oranges | HPLC-UV-VIS | Argentina | 50 | 0.1 | [29] | |
| Seedless grapes | HPLC-UV | Pakistan | 70 | 286.1 | [66] | |
| Red globe grapes | HPLC-UV | Pakistan | 75 | 921.1 | [66] | |
| Flame grapes | HPLC-UV | Pakistan | 66.7 | 190.1 | [66] | |
| Pineapples | HPLC-UV | Pakistan | 81.8 | 254.1 | [66] | |
| Pears | HPLC-UV | Pakistan | 66.7 | 232.1 | [66] | |
| Tomatoes | HPLC-UV | Pakistan | 80 | 410.2 | [66] | |
| Dried fruits | Figs | HPLC-UV | China | 65 | 87.6 | [68] |
| Longans | HPLC-UV | China | 90.5 | 68.4 | [68] | |
| Apricots, dates, plums, peaches and bananas | HPLC-UV | China | 8.3 | 7.4 | [68] | |
| Apricots | HPLC-UV | Iraq | 100 | 0.008–2.84 | [69] | |
| Grapes | HPLC-UV | Iraq | 100 | 0.0198–30.5 | [69] | |
| Fruit products | Pear juice | HPLC-UV | Tunisia | 47.61% | 62.5 | [64] |
| Pear jams | HPLC-UV | Tunisia | 43.75% | 123.7 | [64] | |
| Fruit juice | HPLC-UV | Pakistan | 58.3 | 110.3 | [66] | |
| Smoothie of tomatoes, mint and carrots | HPLC-UV | Pakistan | 44.4 | 50.7 | [66] | |
| Smoothie of pineapple and watermelon | HPLC-UV | Pakistan | 42.9 | 60.6 | [66] | |
| Smoothie of oranges, carrots and mint | HPLC-UV | Pakistan | 50 | 110.4 | [66] | |
| Smoothie of banana, mangoes and strawberry | HPLC-UV | Pakistan | 50 | 20.3 | [66] | |
| Hawthorn products | HPLC-UV | China | 10 | 5.1 | [68] | |
| Fruit juice | HPLC-UV | China | 15 | 5.4 | [68] | |
| Fruit jams | HPLC-UV | China | 10 | 5.0 | [68] |
| Foodstuffs | Detection Method | Country | AOH | AME | TeA | Ref. | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Positives (%) | AOH (µg/Kg) | Positives (%) | AME (µg/Kg) | Positives (%) | TeA (µg/Kg) | |||||
| Fresh fruits | Apples | LC–MS/MS | China | 27.88 | 6.71–8517 | 16.35 | 4.97–2623 | 66.35 | 36–145276 | [9] |
| Apples | LC–MS/MS | Netherlands | - | 0 | - | 0 | - | 0 | [88] | |
| Tomatoes | LC–MS/MS | Netherlands | - | 0 | - | 0 | - | 0 | [88] | |
| Grapes | UPLC–MS/MS | China | 26.8% | 0.09–7.15 | 3.6% | 0.11–0.15 | 28.6% | 0.25–46.97 | [91] | |
| Dried fruits | Raisins | UPLC–MS/MS | China | 5.3 | 3.5~15.6 | 19.3 | 0.3~13.5 | 35.1 | 6.9~594.4 | [91] |
| Dates | UPLC–MS/MS | China | - | - | - | - | 34.0 | 9.6~4411.4 | [91] | |
| Apricots | UPLC–MS/MS | China | - | - | 5.4 | 0.5~2.1 | 37.5 | 10.4~1231.8 | [91] | |
| Figs | LC–MS/MS | Netherlands | 0 | - | 0 | - | 100 | 25–2345 | [88] | |
| Wolfberries | UPLC–MS/MS | China | 3.7 | 5.9~27.4 | 7.4 | 0.2~15.0 | 64.8 | 23.8~5665.3 | [91] | |
| Fruits products | Apple juice | LC–MS/MS | China | 2.44 | 3.70 | 4.88 | <LOQ | 9.76 | <LOQ | [9] |
| Apple jams | LC–MS/MS | China | 23.53 | <LOQ-4.4 | - | - | - | - | [9] | |
| Apple vinegar | LC–MS/MS | China | 2.94 | <LOQ | - | - | 2.94 | 14.5 | [9] | |
| Juice | EIA | Germany | 56.5 | 0.65–16 | 43.5 | 0.14–4.9 | 52.2 | 21–250 | [92] | |
| Red wine | LC-UV and LC–MS/MS | Canada | 83.3 | 0.03–19.4 | 83.3 | 0.01–0.23 | - | - | [93] | |
| Tomato sauce | HPLC–MS/MS | Belgium | 86 | <LOQ-42 | 78 | <LOQ-3.8 | 84.3 | 7.7–330.6 | [94] | |
| Tomato concentrate | HPLC–MS/MS | Belgium | 85 | <LOQ-31 | 67 | <LOQ-6.1 | 100 | <LOQ-174 | [94] | |
| Tomato juice | HPLC–MS/MS | Belgium | 71 | <LOQ-7.0 | 54 | <LOQ-3.3 | 100 | 3.7–333.1 | [94] | |
| Trockenbeerenauslese | LC–MS/MS | Germany | 66.7 | 1.2–4.9 | 66.7 | 0.1–0.3 | - | - | [95] | |
| Vegetable products | LC–MS/MS | Germany | 50 | 2.6–25 | 60 | 0.1–5 | [95] | |||
| Mycotoxin | Sequence | Dissociation Constant (nmol/L) | Reference |
|---|---|---|---|
| OTA | GATCGGGTGTGGGTGGCGTAAAGGGAGCATCGGACA | 200 | [139] |
| PAT | GGCCCGCCAACCCGCATCATCTACACTGAT ATTTTACCT T | 21.83 ± 5.022 | [138] |
| ZEN | CGTGCTACCGTGAAATACCAGCTTATTCAATTCTACCAGCTTTG AGGCTCGATCCAGCTTATTCAATTATACCAGCTTATTCAATTATACCAGCACAATCGTAATCAGTTAG | 15.2 ± 3.4 | [138] |
| FB1 | ATACCAGCTTATTCAATTAATCGCATTACCTTATACCAGCTTATTCAATTACGTCTGCACATACCAGCTTATTCAATTAGATAGTAAGTGCAATCT | 100 ± 30 | [139] |
| T-2 | GTATATCAAGCATCGCGTGTTTACACATGCGAGAGGTGAA | 20.8 ± 3.1 | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nan, M.; Xue, H.; Bi, Y. Contamination, Detection and Control of Mycotoxins in Fruits and Vegetables. Toxins 2022, 14, 309. https://doi.org/10.3390/toxins14050309
Nan M, Xue H, Bi Y. Contamination, Detection and Control of Mycotoxins in Fruits and Vegetables. Toxins. 2022; 14(5):309. https://doi.org/10.3390/toxins14050309
Chicago/Turabian StyleNan, Mina, Huali Xue, and Yang Bi. 2022. "Contamination, Detection and Control of Mycotoxins in Fruits and Vegetables" Toxins 14, no. 5: 309. https://doi.org/10.3390/toxins14050309
APA StyleNan, M., Xue, H., & Bi, Y. (2022). Contamination, Detection and Control of Mycotoxins in Fruits and Vegetables. Toxins, 14(5), 309. https://doi.org/10.3390/toxins14050309